
Analysis of Differentially Expressed MicroRNAs in Serum and Lung Tissues from Individuals with Severe Asthma Treated with Oral Glucocorticoids
 , , , , , , , , ,
, , , , , , , , ,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Study Population
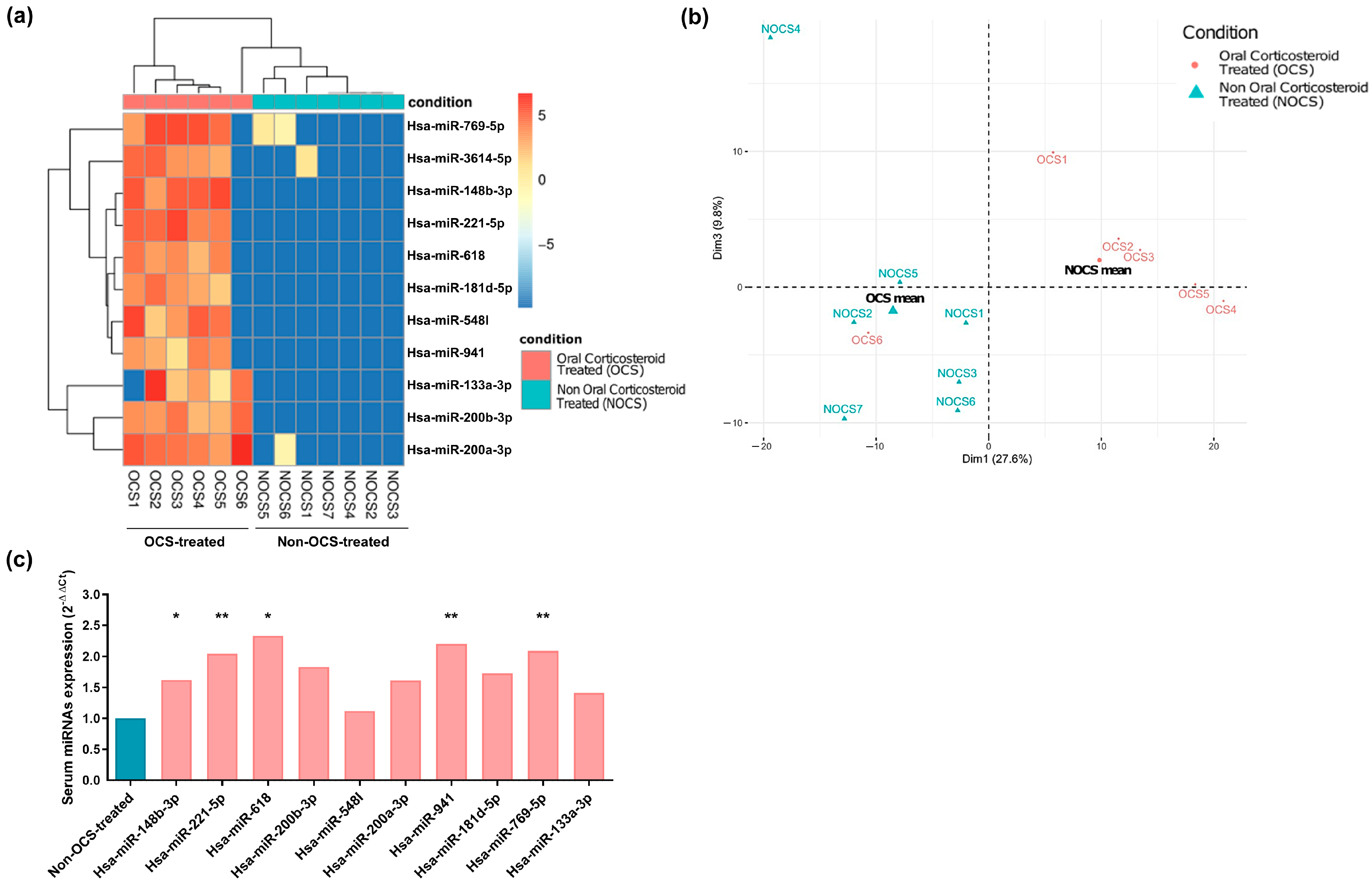
2.2. MiRNA Profile in Serum Samples
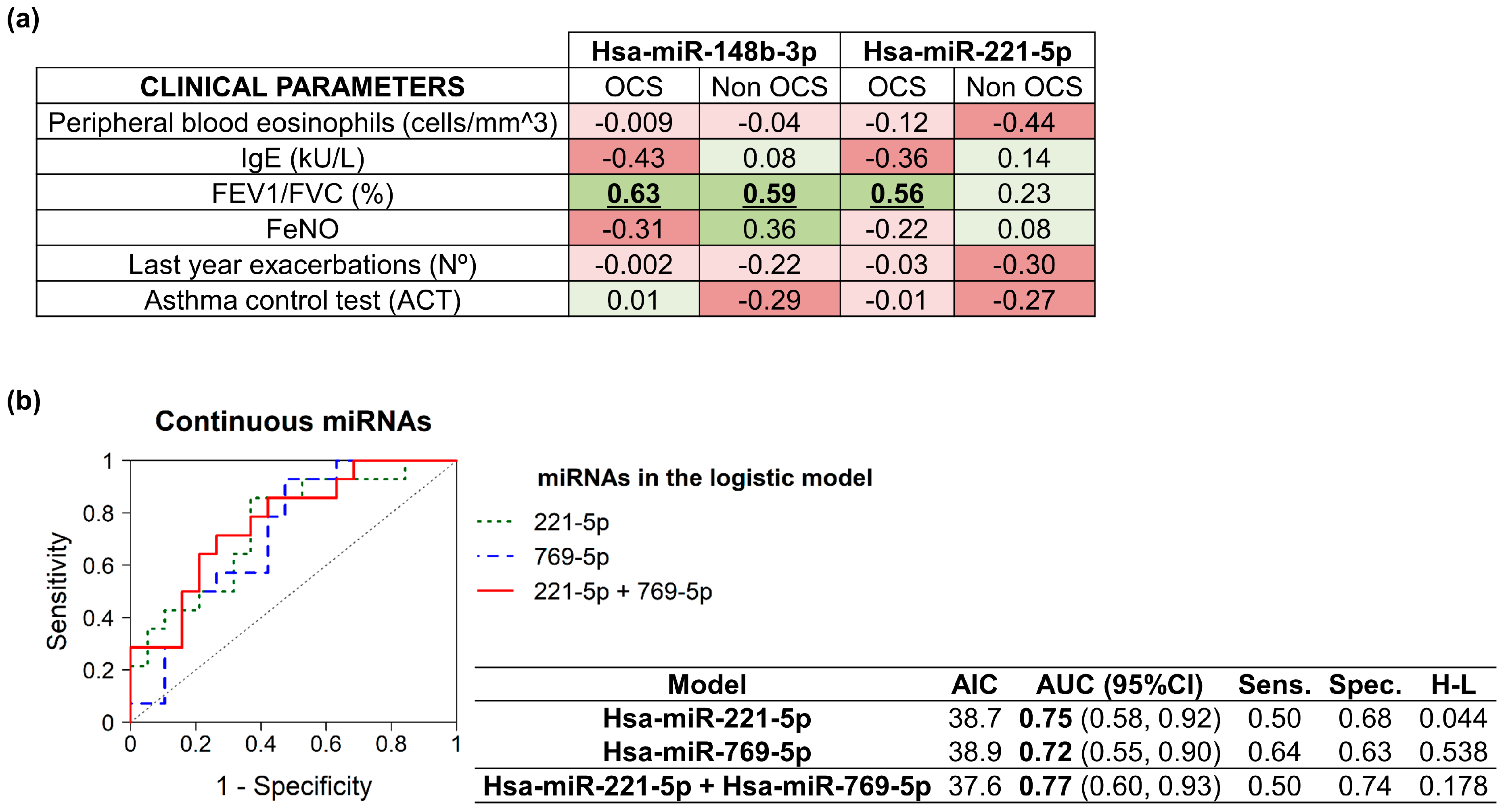
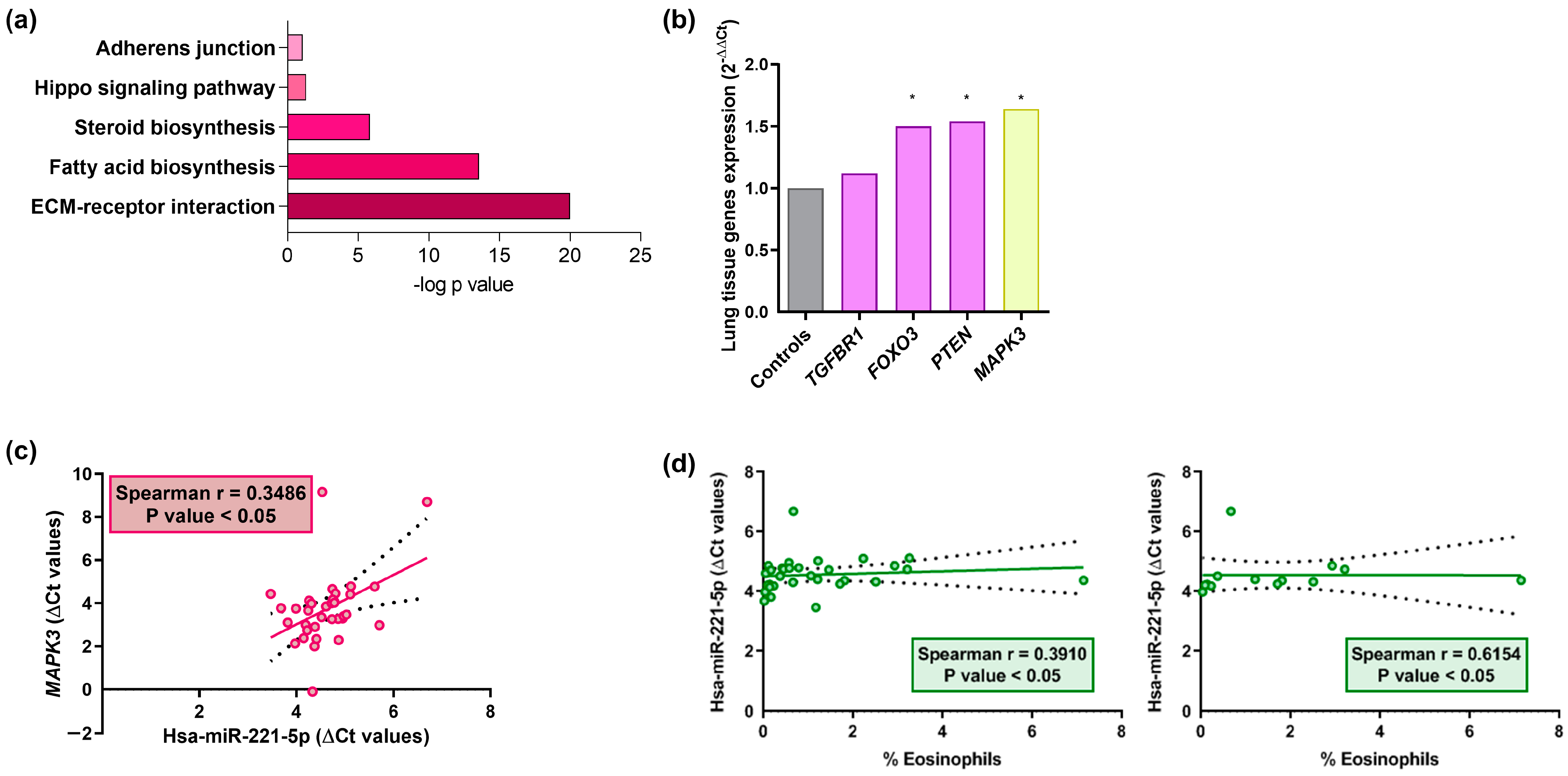
2.3. MiRNAs Correlation and Signaling Pathways
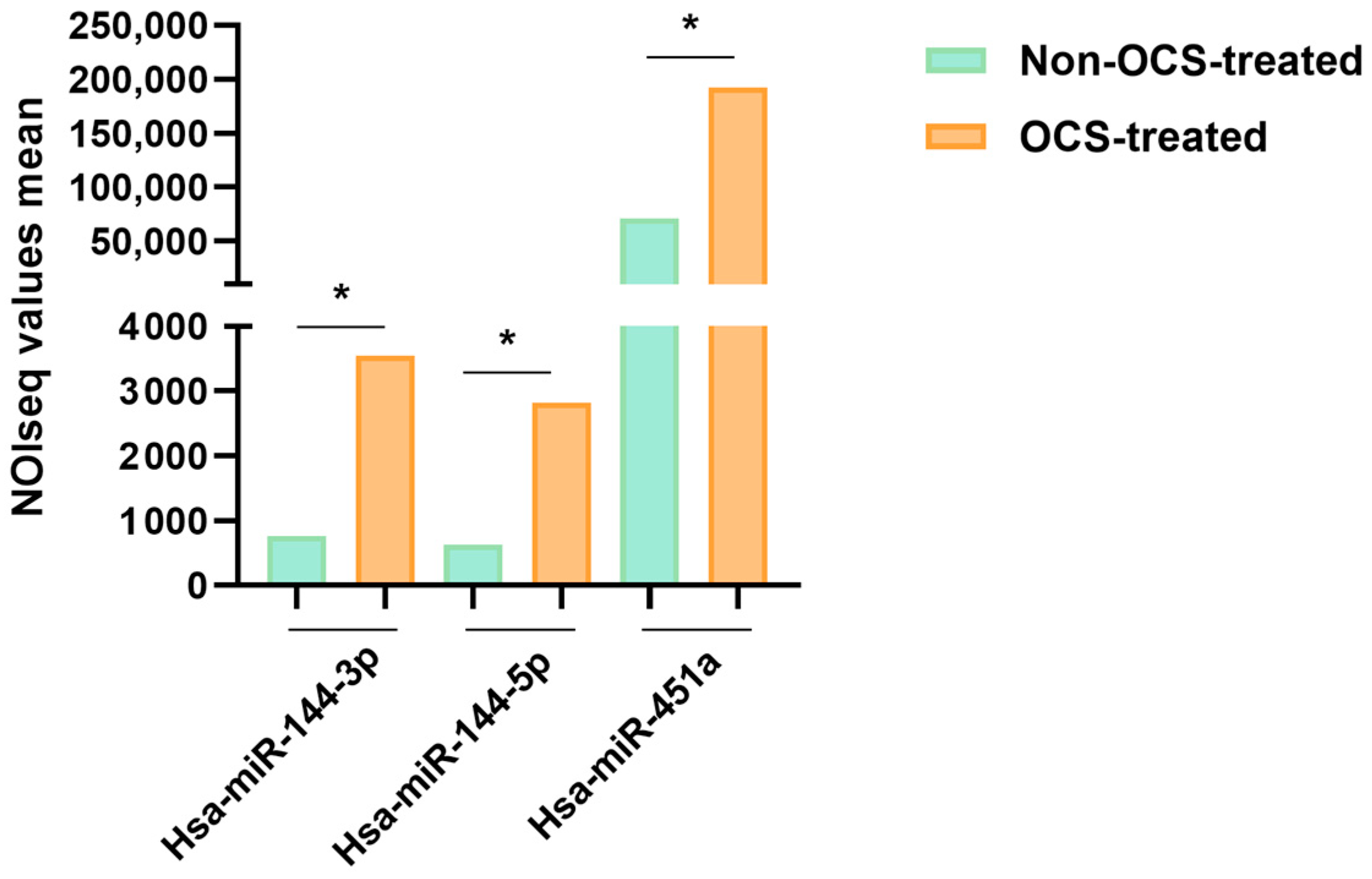
2.4. MiRNA Profile in Lung Samples
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Sample Collection
4.2. Library Preparation, MiRNAs-Seq, Bioinformatic Analysis
4.3. RT-qPCR and Pathway Enrichment Analyses
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hough, K.P.; Curtiss, M.L.; Blain, T.J.; Liu, R.M.; Trevor, J.; Deshane, J.S.; Thannickal, V.J. Airway Remodeling in Asthma. Front. Med. 2020, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F.; Wenzel, S.E.; Brozek, J.L.; Bush, A.; Castro, M.; Sterk, P.J.; Adcock, I.M.; Bateman, E.D.; Bel, E.H.; Bleecker, E.R.; et al. International ERS/ATS guidelines on definition, evaluation and treatment of severe asthma task force report ERS/ATS guidelines on severe asthma Executive Summary. Eur. Respir. J. 2014, 43, 343–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Côté, A.; Godbout, K.; Boulet, L.-P. The management of severe asthma in 2020. Biochem. Pharmacol. 2020, 179, 114112. [Google Scholar] [CrossRef] [PubMed]
- Agache, I.; Akdis, C.A. Precision medicine and phenotypes, endotypes, genotypes, regiotypes, and theratypes of allergic diseases. J. Clin. Investig. 2019, 129, 1493–1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheikhpour, M.; Maleki, M.; Ebrahimi Vargoorani, M.; Amiri, V. A review of epigenetic changes in asthma: Methylation and acetylation. Clin. Epigenet. 2021, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Kuruvilla, M.E.; Lee, F.E.H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Busse, W.W. Biological treatments for severe asthma: A major advance in asthma care. Allergol. Int. 2019, 68, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Suehs, C.M.; Menzies-Gow, A.; Price, D.; Bleecker, E.R.; Canonica, G.W.; Gurnell, M.; Bourdin, A. Expert consensus on the tapering of oral corticosteroids for the treatment of asthma: A delphi study. Am. J. Respir. Crit. Care Med. 2021, 203, 871–881. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Q.; Wang, J.; Gao, S.; Li, C.; Wang, J.; Zhang, S.; Lin, J. Variability of Type 2 inflammatory markers guiding biologic therapy of severe asthma: A 5-year retrospective study from a single tertiary hospital. World Allergy Organ. J. 2021, 14, 100547. [Google Scholar] [CrossRef]
- Weidner, J.; Bartel, S.; Kılıç, A.; Zissler, U.M.; Renz, H.; Schwarze, J.; Schmidt-Weber, C.B.; Maes, T.; Rebane, A.; Krauss-Etschmann, S.; et al. Spotlight on microRNAs in allergy and asthma. Allergy Eur. J. Allergy Clin. Immunol. 2021, 76, 1661–1678. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.Y.; Chang, D.C.; Lin, S.L. The MicroRNA (miRNA): Overview of the RNA Genes that Modulate Gene Function. Mol. Biotechnol. 2008, 38, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Dexheimer, P.J.; Cochella, L. MicroRNAs: From Mechanism to Organism. Front. Cell Dev. Biol. 2020, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.A.; Ludwig, R.G.; Garcia-Martin, R.; Brandão, B.B.; Kahn, C.R. Extracellular miRNAs: From Biomarkers to Mediators of Physiology and Disease. Cell Metab. 2019, 30, 656–673. [Google Scholar] [CrossRef] [PubMed]
- Gil-Martínez, M.; Rodrigo-Muñoz, J.; Sastre, B.; Cañas, J.; García-Latorre, R.; Redondo, N.; de la Fuente, L.; Mínguez, P.; Mahíllo-Fernández, I.; Sastre, J.; et al. Serum microRNAs catalog asthmatic patients by phenotype. J. Investig. Allergol. Clin. Immunol. 2021, 32, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Popović-Grle, S.; Štajduhar, A.; Lampalo, M.; Rnjak, D. Biomarkers in Different Asthma Phenotypes. Genes 2021, 12, 801. [Google Scholar] [CrossRef] [PubMed]
- Mirra, D.; Cione, E.; Spaziano, G.; Esposito, R.; Sorgenti, M.; Granato, E.; Cerqua, I.; Muraca, L.; Iovino, P.; Gallelli, L.; et al. Circulating MicroRNAs Expression Profile in Lung Inflammation: A Preliminary Study. J. Clin. Med. 2022, 11, 5446. [Google Scholar] [CrossRef]
- Carr, T.F.; Bleecker, E. Asthma heterogeneity and severity. World Allergy Organ. J. 2016, 9, 41. [Google Scholar] [CrossRef] [Green Version]
- Licari, A.; Castagnoli, R.; Brambilla, I.; Marseglia, A.; Tosca, M.A.; Marseglia, G.L.; Ciprandi, G. Asthma endotyping and biomarkers in childhood asthma. Pediatr. Allergy Immunol. Pulmonol. 2018, 31, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Xtara, X.; Carr, F.; Xmd, X.; Kraft, X. Use of biomarkers to identify phenotypes and endotypes of severe asthma. Ann. Allergy Asthma Immunol. 2018, 121, 414–420. [Google Scholar] [CrossRef]
- Tiotiu, A. Biomarkers in asthma: State of the art. Asthma Res. Pract. 2018, 4, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonica, G.W.; Blasi, F.; Crimi, N.; Paggiaro, P.; Papi, A.; Fanelli, F.; Stassaldi, A.; Furneri, G. Defining type 2 asthma and patients eligible for dupilumab in Italy: A biomarker-based analysis. Clin. Mol. Allergy 2021, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Zieg, G.; Lack, G.; Harbeck, R.J.; Gelfand, E.W.; Leung, D.Y. In vivo effects of glucocorticoids on IgE production. J. Allergy Clin. Immunol. 1994, 94, 222–230. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, J.M.; Lemiere, C.; Lougheed, M.D.; Ducharme, F.M.; Dell, S.D.; Ramsey, C.; Yang, M.C.L.; Côté, A.; Watson, W.; Olivenstein, R.; et al. Recognition and management of severe asthma: A Canadian Thoracic Society position statement. Can. J. Respir. Crit. Care Sleep Med. 2017, 1, 199–221. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Li, Q.; Zhang, R.; Dai, X.; Chen, W.; Xing, D. Circulating microRNAs: Biomarkers of disease. Clin. Chim. Acta 2021, 516, 46–54. [Google Scholar] [CrossRef]
- Potla, P.; Ali, S.A.; Kapoor, M. A bioinformatics approach to microRNA-sequencing analysis. Osteoarthr. Cartil. Open 2021, 3, 100131. [Google Scholar] [CrossRef]
- Rodrigo-Muñoz, J.M.; Gil-Martínez, M.; Lorente-Sorolla, C.; García-Latorre, R.; Valverde-Monge, M.; Quirce, S.; Sastre, J.; del Pozo, V. miR-144-3p Is a Biomarker Related to Severe Corticosteroid-Dependent Asthma. Front. Immunol. 2022, 13, 858722. [Google Scholar] [CrossRef]
- Ong, J.; Faiz, A.; Timens, W.; van den Berge, M.; Terpstra, M.M.; Kok, K.; van den Berg, A.; Kluiver, J.; Brandsma, C.A. Marked TGF-β-regulated miRNA expression changes in both COPD and control lung fibroblasts. Sci. Rep. 2019, 9, 18214. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Ma, Y.; Tang, Y.; Liu, X.; Zhao, Y.; An, L. MiRNA-221-5p suppressed the Th17/Treg ratio in asthma via RORγt/Foxp3 by targeting SOCS1. Allergy Asthma Clin. Immunol. 2021, 17, 123. [Google Scholar] [CrossRef]
- Li, C.; Hu, X.; Li, L.; Li, J. hui Differential microRNA expression in the peripheral blood from human patients with COVID-19. J. Clin. Lab. Anal. 2020, 34, e23590. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.F.; Yan, P.; Zhao, D.H.; Shi, W.F.; Meng, S.; Liu, Y.; Liu, B.; Li, G.F.; Xie, L.X. The effect of environmental factors on the differential expression of miRNAs in patients with chronic obstructive pulmonary disease: A pilot clinical study. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 741–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigo-Muñoz, J.M.; Cañas, J.A.; Sastre, B.; Rego, N.; Greif, G.; Rial, M.; Mínguez, P.; Mahíllo-Fernández, I.; Fernández-Nieto, M.; Mora, I.; et al. Asthma diagnosis using integrated analysis of eosinophil microRNAs. Allergy Eur. J. Allergy Clin. Immunol. 2019, 74, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Jovičić, A.; Roshan, R.; Moisoi, N.; Pradervand, S.; Moser, R.; Pillai, B.; Luthi-Carter, R. Comprehensive Expression Analyses of Neural Cell-Type-Specific miRNAs Identify New Determinants of the Specification and Maintenance of Neuronal Phenotypes. J. Neurosci. 2013, 33, 5127–5137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imraish, A.; Abu-Thiab, T.; Zihlif, M. IL-13 and FOXO3 genes polymorphisms regulate IgE levels in asthmatic patients. Biomed. Rep. 2021, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Yan, J.; Han, X.R.; Zheng, G.H.; Tang, R.; Liu, L.F.; Wu, D.M.; Lu, J.; Zheng, Y.L. PTEN gene silencing contributes to airway remodeling and induces airway smooth muscle cell proliferation in mice with allergic asthma. J. Thorac. Dis. 2018, 10, 202–211. [Google Scholar] [CrossRef] [Green Version]
- Alam, R.; Gorska, M.M. MAPK Signaling and ERK1/2 bistability in Asthma. Clin. Exp. Allergy 2011, 41, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, X.; Álvarez-Puebla, M.J.; Arismendi, E.; Arochena, L.; del Pilar Ausín, M.; Barranco, P.; Bobolea, I.; Cañas, J.A.; Cardaba, B.; Crespo, A.; et al. Estudio de los mecanismos implicados en la génesis y evolución del asma (proyecto MEGA): Creación y seguimiento a largo plazo de una cohorte de pacientes asmáticos. Arch. Bronconeumol. 2018, 54, 378–385. [Google Scholar] [CrossRef]
- Global Initiative for Asthma-Global Initiative for Asthma-GINA. Available online: https://ginasthma.org/ (accessed on 10 May 2021).
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarazona, S.; García-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 10 January 2023).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OCS-Treated (n = 20) | Non-OCS-Treated (n = 26) | p-Value | ||
|---|---|---|---|---|
| Age (years) † | 54.5 (44.3–59.5) | 53.0 (42.8–61.3) | N.S. | |
| Sex (%) | Female | 17 (85.0%) | 23 (88.5%) | N.S. |
| BMI † | 26.7 (23.1–30.4) | 28.7 (24.2–33.4) | N.S. | |
| Smoking habit (%) | Smokers | 1 (5.0%) | 4/25 (16.0%) | N.S. |
| Passive | 2 (10.0%) | 2/25 (8.0%) | N.S. | |
| Ex-smokers | 7 (35.0%) | 9/25 (36.0%) | N.S. | |
| Non-smokers | 10 (50.0%) | 10/25 (40.0%) | N.S. | |
| Blood eosinophils (cells/µL) † | 300.0 (100.0–500.0) | 300.0 (200.0–525.0) | N.S. | |
| Sputum eosinophils (%) † | 0.0% (0.0–35.0) | 3.1% (2.0–22.1) | N.S. | |
| Atopy (%) | 12 (60.0%) | 17 (65.4%) | N.S. | |
| IgE (IU) † | 366.0 (110.0–690.0) | 96.8 (36.6–285.8) | * | |
| FEV1/FVC (%) # | 67.4% (±15.9) | 73.3% (±23.5) | N.S. | |
| FeNO (ppb) † | 23.0 (13.5–78.3) | 32.0 (13.8–62.0) | N.S. | |
| Exacerbations (%) | 17 (85.0%) | 16 (61.5%) | N.S. | |
| Severity (%) | Severe | 20 (100.0%) | 26 (100.0%) | N.S. |
| Moderate | 0 (0.0%) | 0 (0.0%) | N.S. | |
| Mild | 0 (0.0%) | 0 (0.0%) | N.S. | |
| Intermittent | 0 (0.0%) | 0 (0.0%) | N.S. | |
| ACT # | 16.8 (±5.9) | 17.2 (±5.5) | N.S. | |
| ICS and LABA (%) | 20 (100.0%) | 26 (100.0%) | N.S. | |
| Systemic corticosteroid (%) | 20 (100.0%) | 0 (0.0%) | **** | |
| OCS-Treated (n = 6) | Non-OCS-Treated (n = 7) | p-Value | ||
|---|---|---|---|---|
| Age (years) † | 63.0 (56.8–66.3) | 34.0 (27.0–74.0) | N.S. | |
| Sex (%) | Female | 3 (50.0%) | 1 (14.3%) | N.S. |
| Smoking habit (%) | Smokers | 1 (16.7%) | 1/6 (16.7%) | N.S. |
| Ex-smokers | 3 (50.0%) | 4/6 (66.7%) | N.S. | |
| Non-smokers | 2 (33.3%) | 1/6 (16.7%) | N.S. | |
| Neutrophils (%) † | 61.9% (50.3–72.5) | 83.6% (70.6–89.5) | N.S. | |
| Lymphocytes (%) † | 24.1% (16.5–35.5) | 8.7% (5.9–16.5) | N.S. | |
| Monocytes (%) † | 10.0% (8.9–12.2) | 5.9% (4.4–9.0) | * | |
| Eosinophils (%) † | 2.1% (0.3–3.0) | 0.5% (0.1–3.1) | N.S. | |
| Basophils (%) † | 0.4% (0.3–0.7) | 0.2% (0.1–0.5) | N.S. | |
| Atopy (%) | 1 (16.7%) | 1 (14.3%) | N.S. | |
| OCS (%) | 6 (100.0%) | 0 (0.0%) | ** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Martínez, M.; Lorente-Sorolla, C.; Rodrigo-Muñoz, J.M.; Lendínez, M.Á.; Núñez-Moreno, G.; de la Fuente, L.; Mínguez, P.; Mahíllo-Fernández, I.; Sastre, J.; Valverde-Monge, M.; et al. Analysis of Differentially Expressed MicroRNAs in Serum and Lung Tissues from Individuals with Severe Asthma Treated with Oral Glucocorticoids. Int. J. Mol. Sci. 2023, 24, 1611. https://doi.org/10.3390/ijms24021611
Gil-Martínez M, Lorente-Sorolla C, Rodrigo-Muñoz JM, Lendínez MÁ, Núñez-Moreno G, de la Fuente L, Mínguez P, Mahíllo-Fernández I, Sastre J, Valverde-Monge M, et al. Analysis of Differentially Expressed MicroRNAs in Serum and Lung Tissues from Individuals with Severe Asthma Treated with Oral Glucocorticoids. International Journal of Molecular Sciences. 2023; 24(2):1611. https://doi.org/10.3390/ijms24021611
Chicago/Turabian StyleGil-Martínez, Marta, Clara Lorente-Sorolla, José M. Rodrigo-Muñoz, Miguel Ángel Lendínez, Gonzalo Núñez-Moreno, Lorena de la Fuente, Pablo Mínguez, Ignacio Mahíllo-Fernández, Joaquín Sastre, Marcela Valverde-Monge, and et al. 2023. "Analysis of Differentially Expressed MicroRNAs in Serum and Lung Tissues from Individuals with Severe Asthma Treated with Oral Glucocorticoids" International Journal of Molecular Sciences 24, no. 2: 1611. https://doi.org/10.3390/ijms24021611