
Zymosan Particle-Induced Hemodynamic, Cytokine and Blood Cell Changes in Pigs: An Innate Immune Stimulation Model with Relevance to Cytokine Storm Syndrome and Severe COVID-19

 ,
,
Abstract
:1. Introduction
2. Results
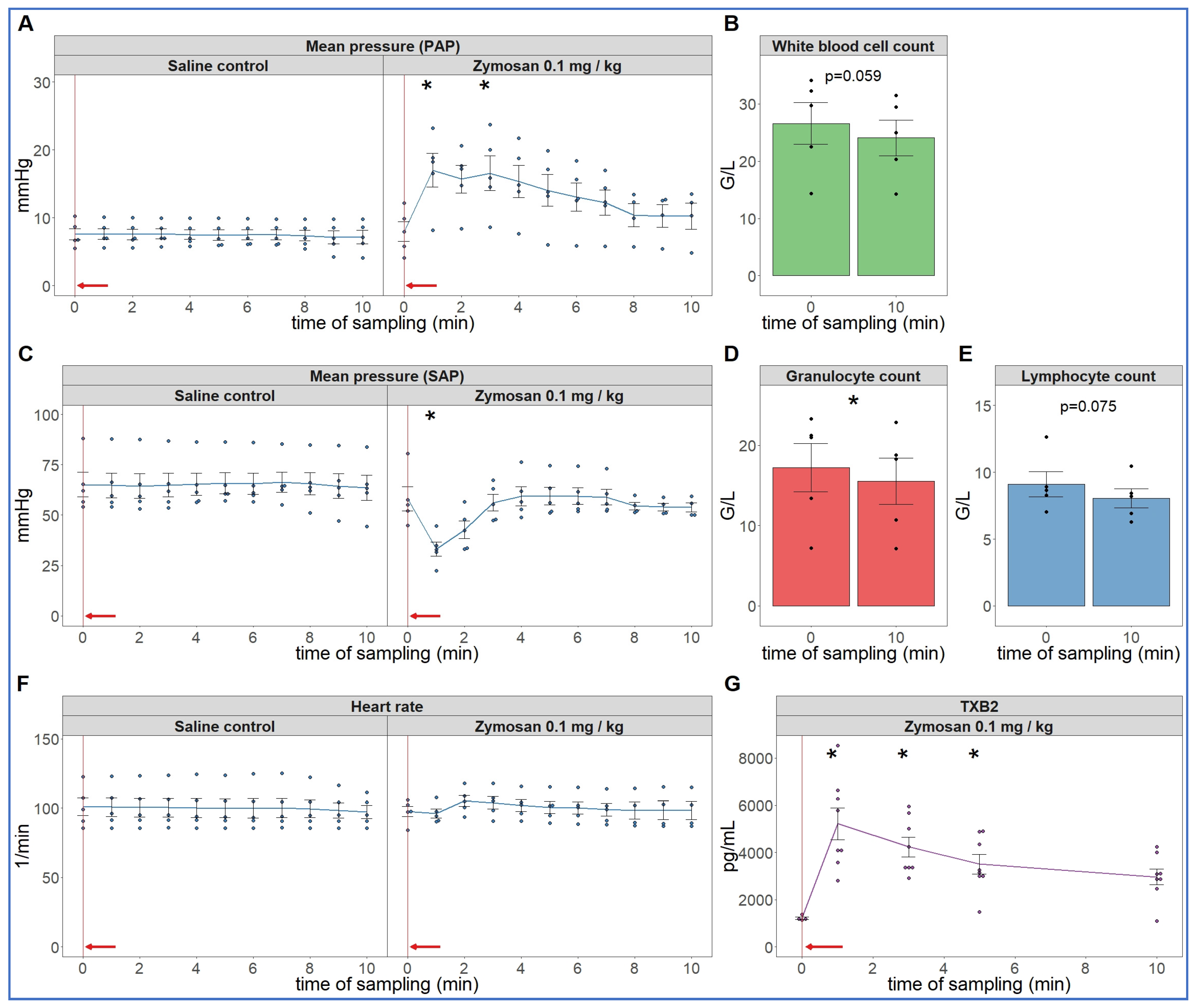
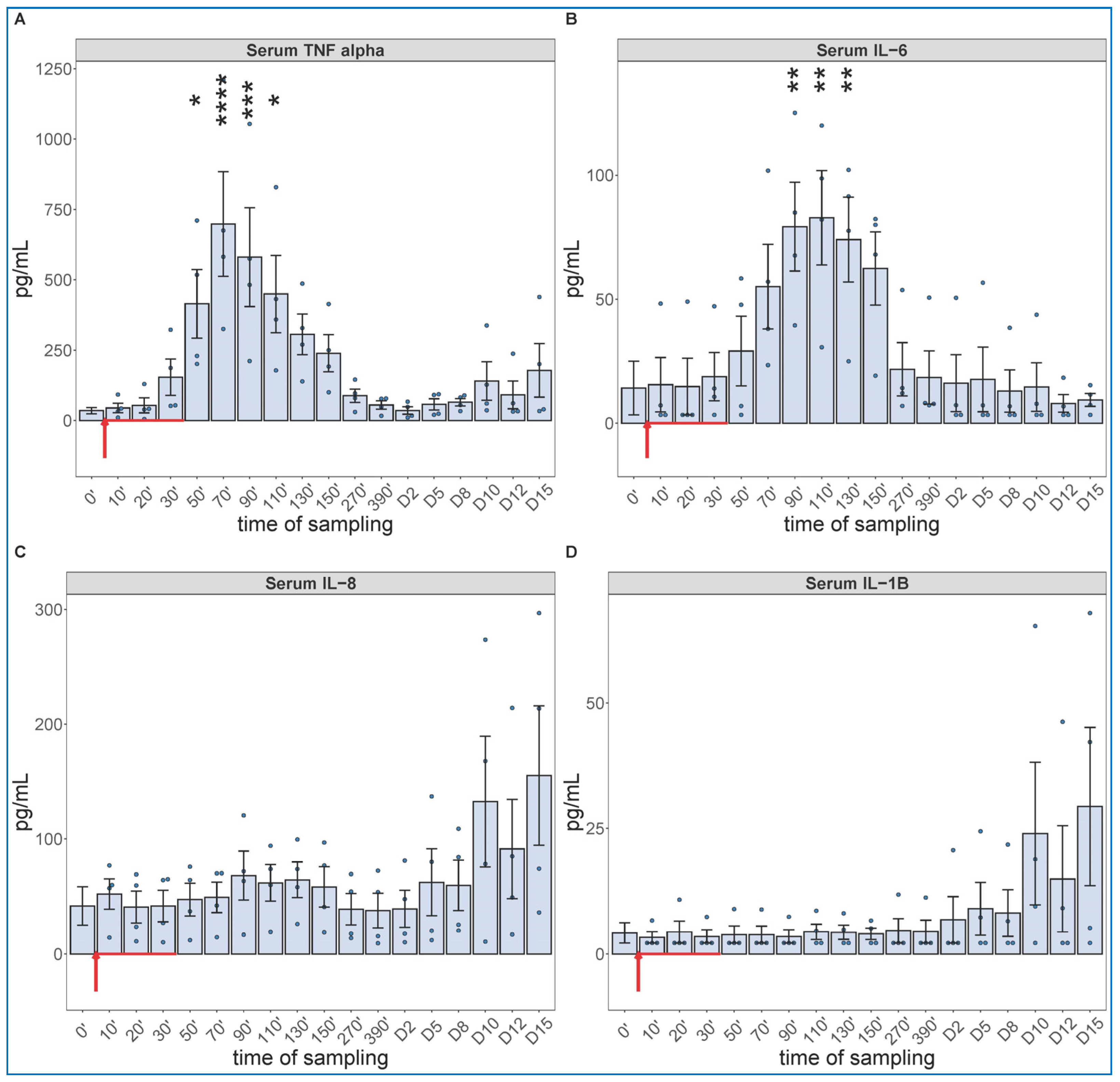
2.1. Early Hemodynamic, Hematological, and Immune Mediator Changes Caused by Low-Dose Bolus Injection of Zymosan
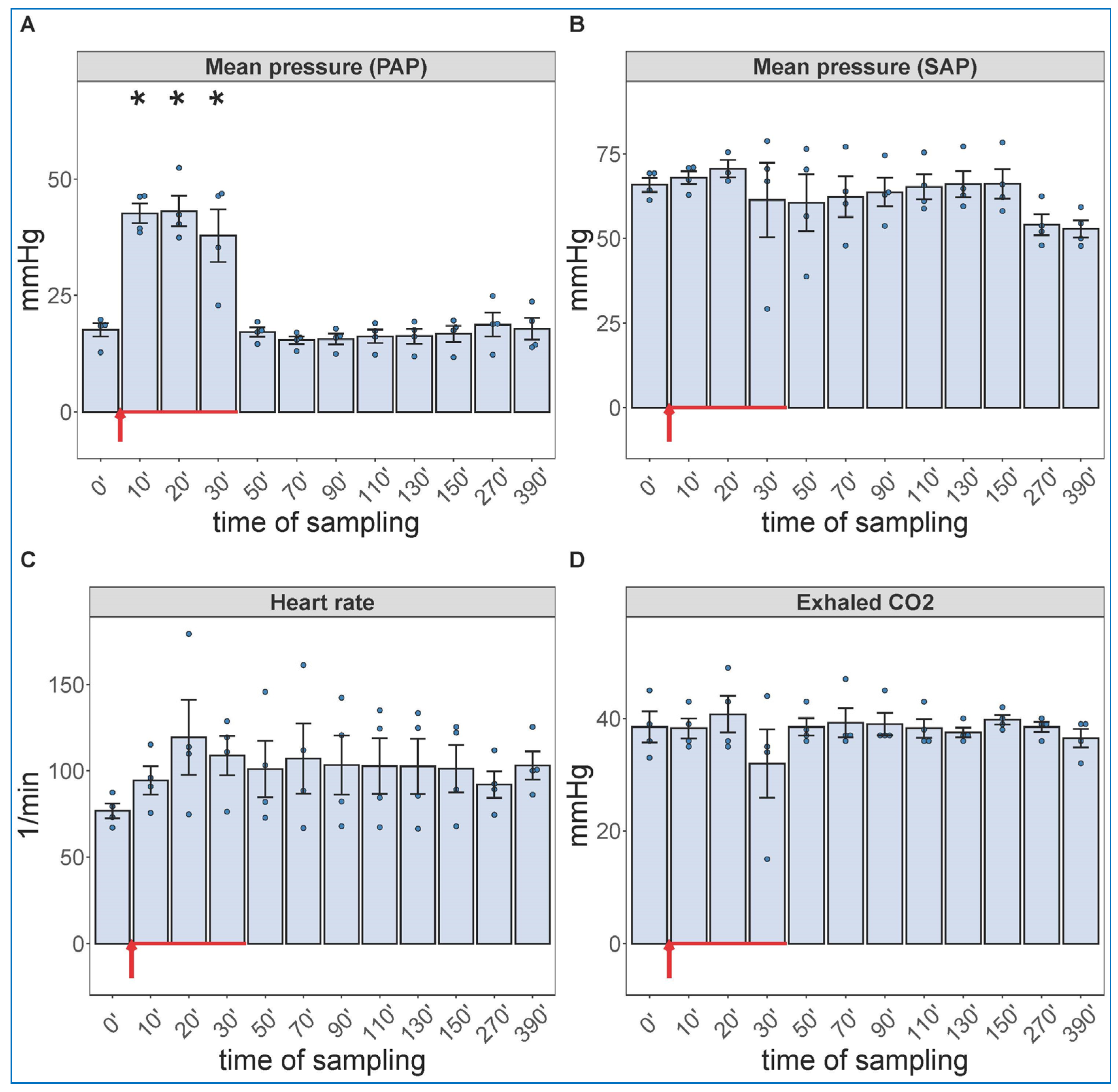
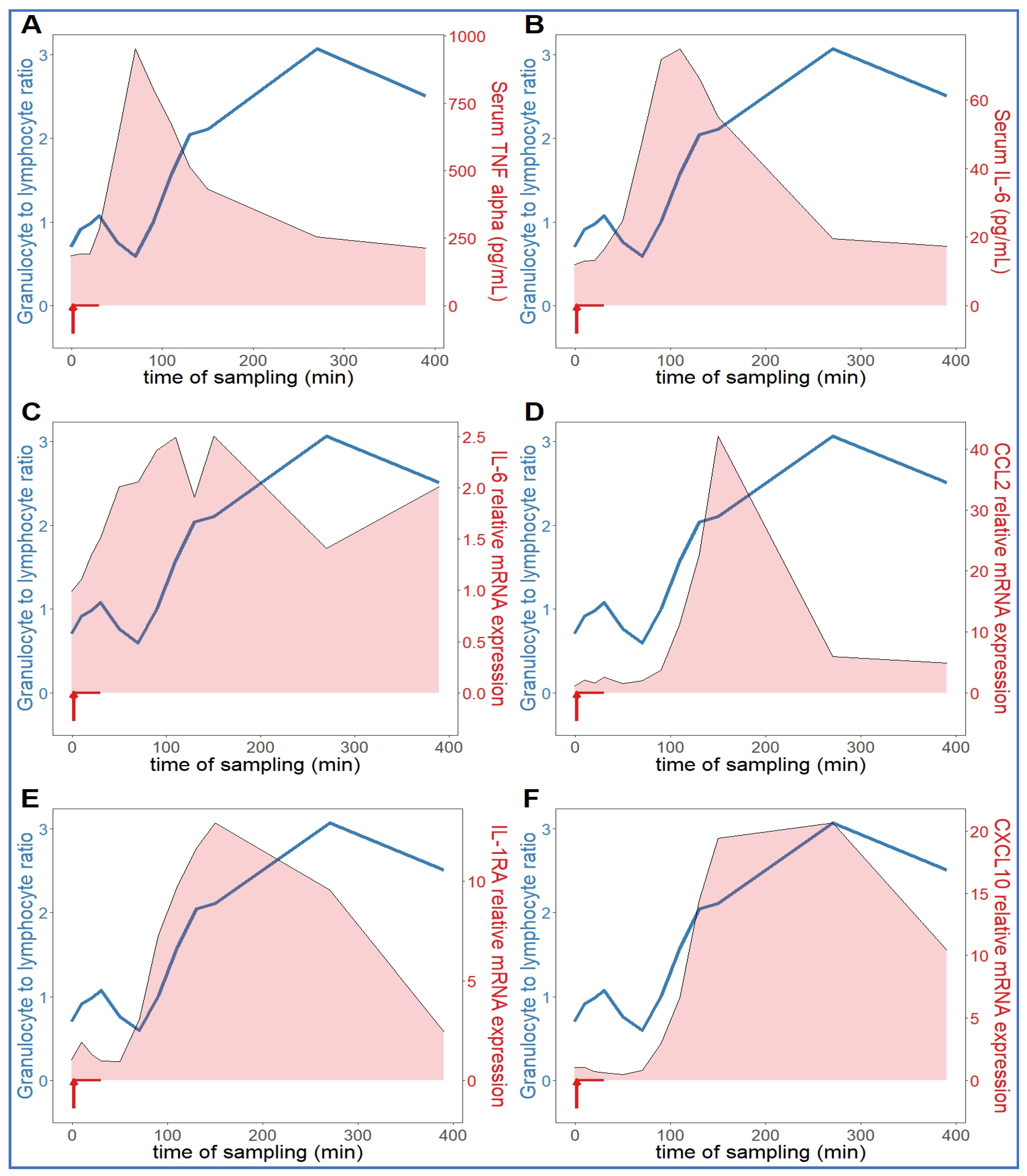
2.2. Extended Follow-Up of Hemodynamic, Hematological, and Immune Mediator Changes Caused by High Dose Zymosan Infusion
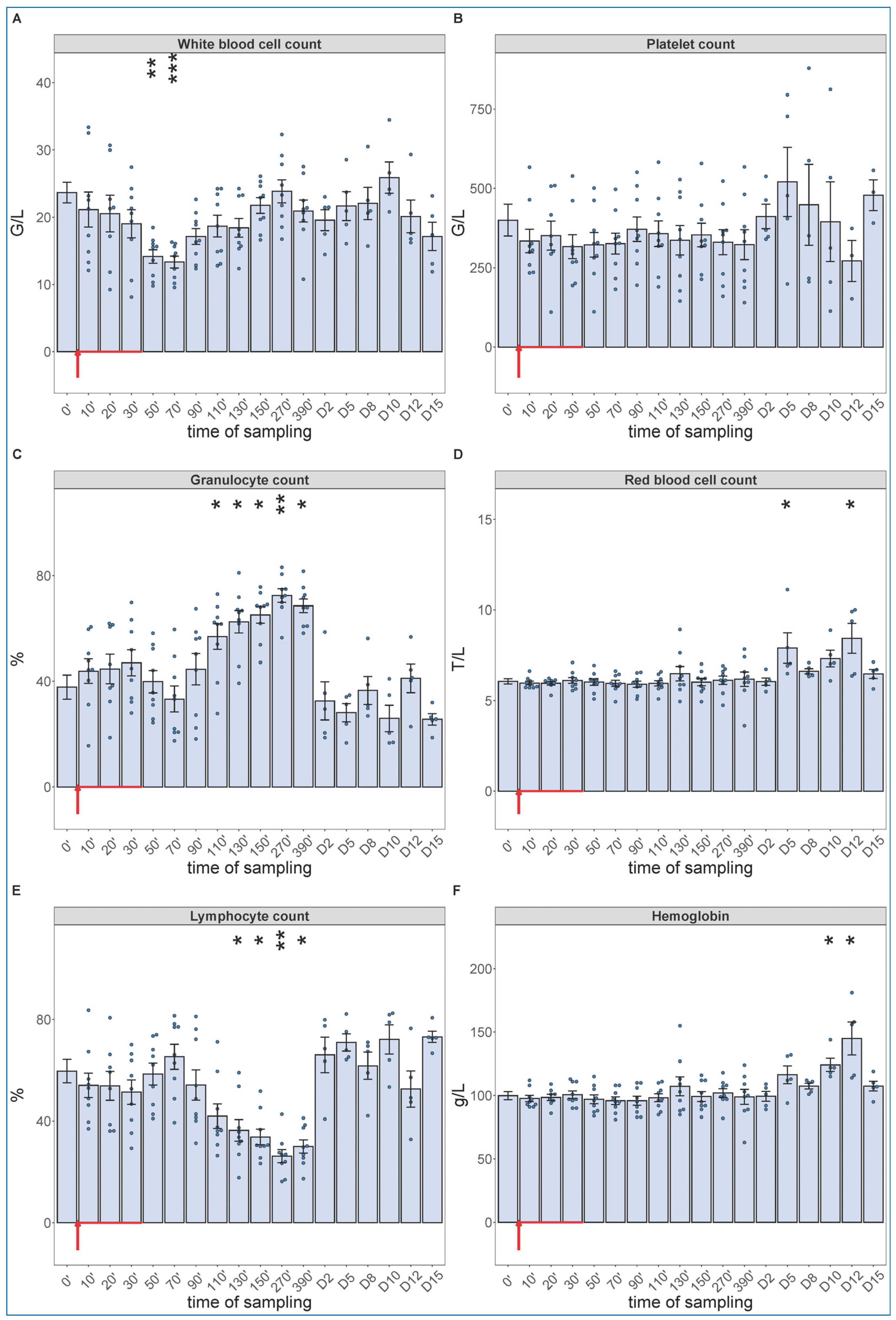
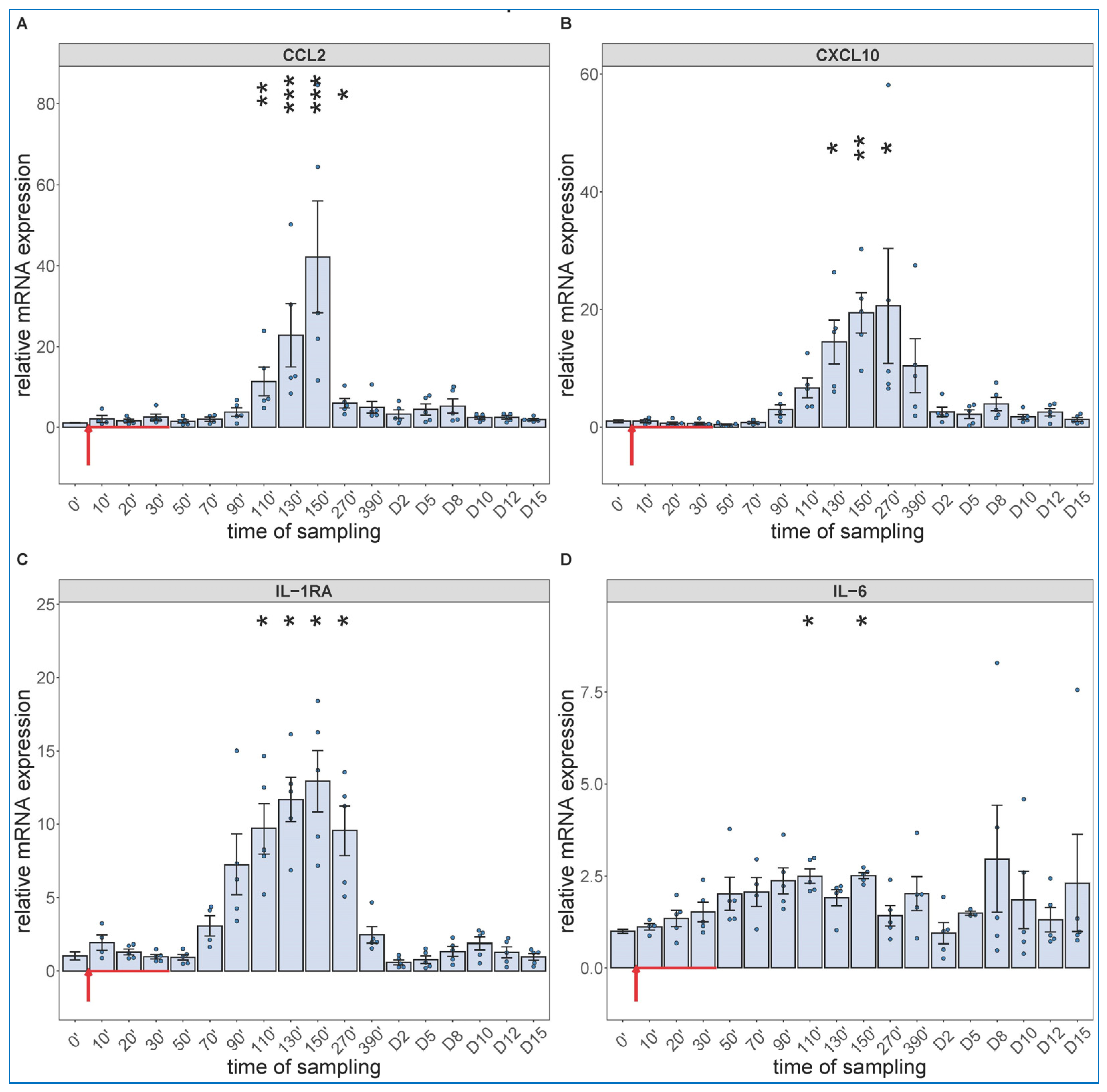
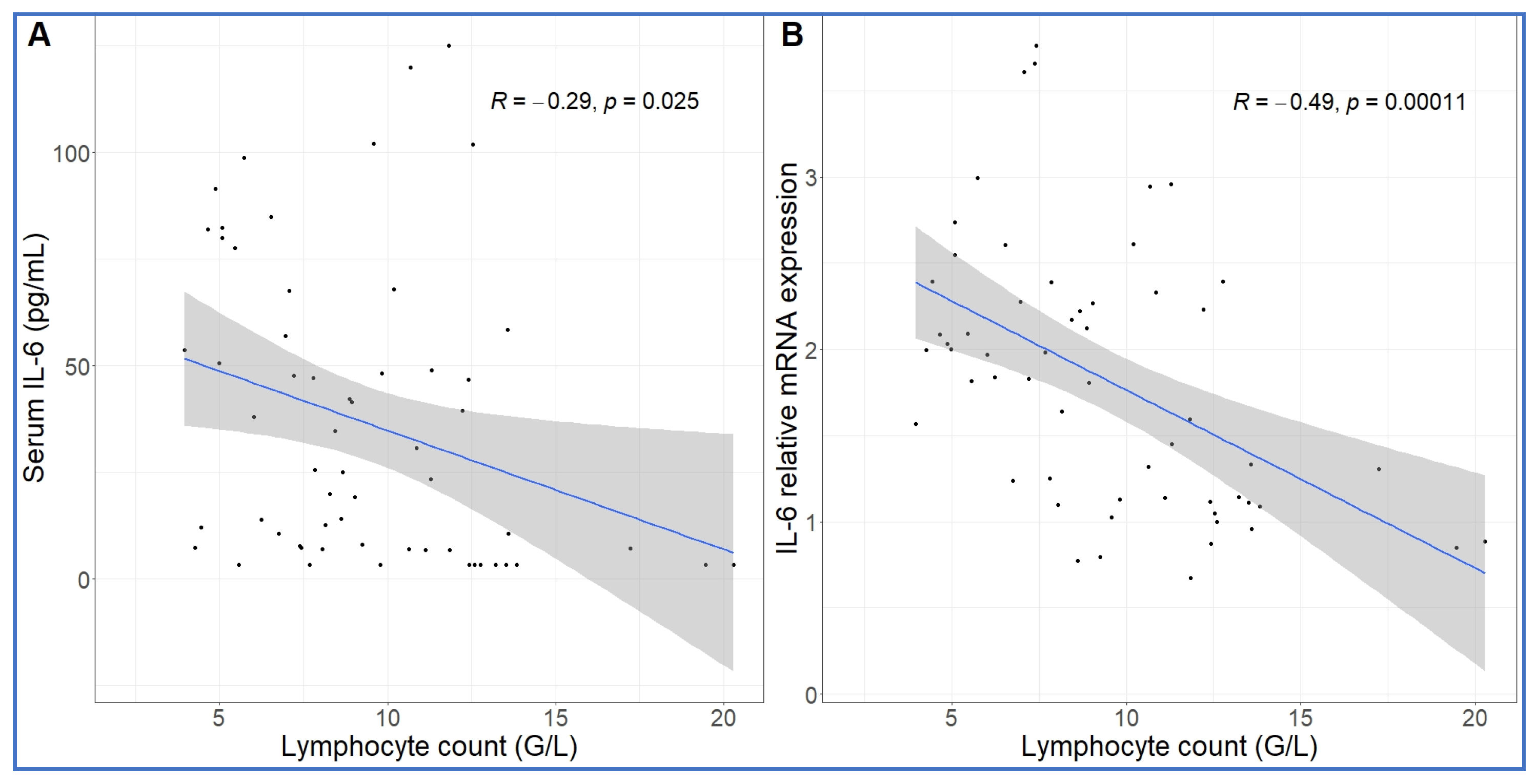
2.3. Long-Term Follow-Up of Hematological, TXB2 and Cytokine Changes Caused by High Dose Zymosan Infusion
3. Discussion
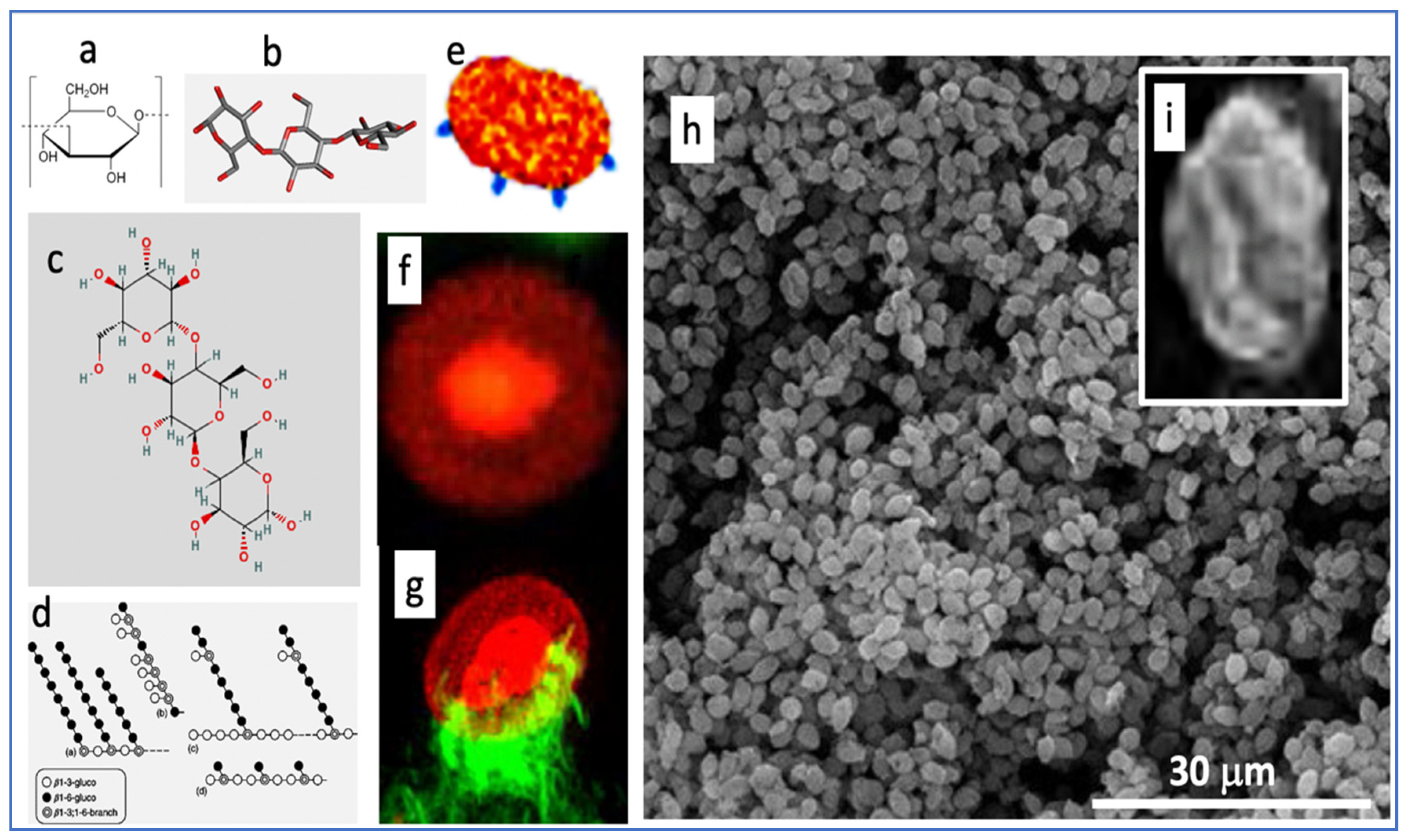
3.1. The Structure and Immune Effects of Zymosan
3.2. Current Animal Models of CSS
3.3. Molecular and Cellular Mechanisms of Zymosan’s Multiple Effects
3.4. The Utility of Porcine Zymosan-Induced CSS Model
3.5. Outlook for the Pharmaceutical Industry
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Anesthesia and Instrumentation
4.4. Experimental Protocols in Different Stages
4.5. Blood Assays
4.6. Cytokine Measurements
4.7. Isolation of PBMC and Quantitative RT-PCR
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, Y.; Liu, J.; Zhang, D.; Xu, Z.; Ji, J.; Wen, C. Cytokine Storm in COVID-19: The Current Evidence and Treatment Strategies. Front. Immunol. 2020, 11, 1708. [Google Scholar] [CrossRef]
- Pearce, L.; Davidson, S.M.; Yellon, D.M. The cytokine storm of COVID-19: A spotlight on prevention and protection. Expert Opin. Ther. Targets 2020, 24, 723–730. [Google Scholar] [CrossRef]
- Sinha, P.; Matthay, M.A.; Calfee, C.S. Is a “Cytokine Storm” Relevant to COVID-19? JAMA Intern. Med. 2020, 180, 1152–1154. [Google Scholar] [CrossRef]
- Buszko, M.; Park, J.H.; Verthelyi, D.; Sen, R.; Young, H.A.; Rosenberg, A.S. The dynamic changes in cytokine responses in COVID-19: A snapshot of the current state of knowledge. Nat. Immunol. 2020, 21, 1146–1151. [Google Scholar] [CrossRef]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Iannaccone, G.; Scacciavillani, R.; Del Buono, M.G.; Camilli, M.; Ronco, C.; Lavie, C.J.; Abbate, A.; Crea, F.; Massetti, M.; Aspromonte, N. Weathering the Cytokine Storm in COVID-19: Therapeutic Implications. Cardiorenal Med. 2020, 10, 277–287. [Google Scholar] [CrossRef]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Peter, A.E.; Sandeep, B.V.; Rao, B.G.; Kalpana, V.L. Calming the Storm: Natural Immunosuppressants as Adjuvants to Target the Cytokine Storm in COVID-19. Front. Pharmacol. 2020, 11, 583777. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Ricci, G.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: Further advances in our understanding the role of specific chemokines involved. Cytokine Growth Factor Rev. 2021, 58, 82–91. [Google Scholar] [CrossRef]
- Coperchini, F.; Chiovato, L.; Rotondi, M. Interleukin-6, CXCL10 and Infiltrating Macrophages in COVID-19-Related Cytokine Storm: Not One for All but All for One! Front. Immunol. 2021, 12, 668507. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef]
- Shen, W.X.; Luo, R.C.; Wang, J.Q.; Chen, Z.S. Features of Cytokine Storm Identified by Distinguishing Clinical Manifestations in COVID-19. Front. Public Health 2021, 9, 671788. [Google Scholar] [CrossRef]
- Wright, D.J.M. Prevention of the cytokine storm in COVID-19. Lancet Infect. Dis. 2021, 21, 25–26. [Google Scholar] [CrossRef]
- Machado, C.; Gonzalez-Quevedo, A. Hypoxemia and Cytokine Storm in COVID-19: Clinical Implications. MEDICC Rev. 2021, 23, 54–59. [Google Scholar]
- Caricchio, R.; Gallucci, M.; Dass, C.; Zhang, X.; Gallucci, S.; Fleece, D.; Bromberg, M.; Criner, G.J. Preliminary predictive criteria for COVID-19 cytokine storm. Ann. Rheum. Dis. 2021, 80, 88–95. [Google Scholar] [CrossRef]
- Cron, R.Q.; Caricchio, R.; Chatham, W.W. Calming the cytokine storm in COVID-19. Nat. Med. 2021, 27, 1674–1675. [Google Scholar] [CrossRef]
- Morgulchik, N.; Athanasopoulou, F.; Chu, E.; Lam, Y.; Kamaly, N. Potential therapeutic approaches for targeted inhibition of inflammatory cytokines following COVID-19 infection-induced cytokine storm. Interface Focus 2022, 12, 20210006. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, E.; Zhao, S.; Li, J.; Zhu, Y.; Zhang, Y.; Cao, L.; Liu, H.; Dong, Y.; Wang, F.; et al. Implications of Laboratory Tests in Disease Grading and Death Risk Stratification of COVID-19: A Retrospective Study in Wuhan, China. Front. Med. 2021, 8, 629296. [Google Scholar] [CrossRef]
- Wu, J.; Shen, J.; Han, Y.; Qiao, Q.; Dai, W.; He, B.; Pang, R.; Zhao, J.; Luo, T.; Guo, Y.; et al. Upregulated IL-6 Indicates a Poor COVID-19 Prognosis: A Call for Tocilizumab and Convalescent Plasma Treatment. Front. Immunol. 2021, 12, 598799. [Google Scholar] [CrossRef]
- Galván-Román, J.M.; Rodríguez-García, S.C.; Roy-Vallejo, E.; Marcos-Jiménez, A.; Sánchez-Alonso, S.; Fernández-Díaz, C.; Alcaraz-Serna, A.; Mateu-Albero, T.; Rodríguez-Cortes, P.; Sánchez-Cerrillo, I.; et al. IL-6 serum levels predict severity and response to tocilizumab in COVID-19: An observational study. J. Allergy Clin. Immunol. 2021, 147, 72–80.e8. [Google Scholar] [CrossRef]
- Smieszek, S.P.; Przychodzen, B.P.; Polymeropoulos, V.M.; Polymeropoulos, C.M.; Polymeropoulos, M.H. Assessing the potential correlation of polymorphisms in the IL6R with relative IL6 elevation in severely ill COVID-19 patients’. Cytokine 2021, 148, 155662. [Google Scholar] [CrossRef]
- Garcia-Gasalla, M.; Berman-Riu, M.; Pons, J.; Rodríguez, A.; Iglesias, A.; Martínez-Pomar, N.; Llompart-Alabern, I.; Riera, M.; Beltrán, A.F.; Figueras-Castilla, A.; et al. Hyperinflammatory State and Low T1 Adaptive Immune Response in Severe and Critical Acute COVID-19 Patients. Front. Med. 2022, 9, 828678. [Google Scholar] [CrossRef]
- Jakobs, K.; Reinshagen, L.; Puccini, M.; Friebel, J.; Wilde, A.-C.B.; Alsheik, A.; Rroku, A.; Landmesser, U.; Haghikia, A.; Kränkel, N.; et al. Disease Severity in Moderate-to-Severe COVID-19 Is Associated with Platelet Hyperreactivity and Innate Immune Activation. Front. Immunol. 2022, 13, 844701. [Google Scholar] [CrossRef]
- Pirsalehi, A.; Salari, S.; Baghestani, A.; Vahidi, M.; Khave, L.J.; Akbari, M.E.; Bashash, D. Neutrophil-to-lymphocyte ratio (NLR) greater than 6.5 may reflect the progression of COVID-19 towards an unfavorable clinical outcome. Iran. J. Microbiol. 2020, 12, 466–474. [Google Scholar] [CrossRef]
- Ciccullo, A.; Borghetti, A.; Verme, L.Z.D.; Tosoni, A.; Lombardi, F.; Garcovich, M.; Biscetti, F.; Montalto, M.; Cauda, R.; Di Giambenedetto, S. Neutrophil-to-lymphocyte ratio and clinical outcome in COVID-19: A report from the Italian front line. Int. J. Antimicrob. Agents 2020, 56, 106017. [Google Scholar] [CrossRef]
- Tudoran, C.; Tudoran, M.; Lazureanu, V.E.; Marinescu, A.R.; Pop, G.N.; Pescariu, A.S.; Enache, A.; Cut, T.G. Evidence of Pulmonary Hypertension after SARS-CoV-2 Infection in Subjects without Previous Significant Cardiovascular Pathology. J. Clin. Med. 2021, 10, 199. [Google Scholar] [CrossRef]
- Pagnesi, M.; Baldetti, L.; Beneduce, A.; Calvo, F.; Gramegna, M.; Pazzanese, V.; Ingallina, G.; Napolano, A.; Finazzi, R.; Ruggeri, A.; et al. Pulmonary hypertension and right ventricular involvement in hospitalised patients with COVID-19. Heart 2020, 106, 1324–1331. [Google Scholar] [CrossRef]
- Khan, A.W.; Ullah, I.; Khan, K.S.; Tahir, M.J.; Masyeni, S.; Harapan, H. Pulmonary arterial hypertension post COVID-19: A sequala of SARS-CoV-2 infection? Respir. Med. Case Rep. 2021, 33, 101429. [Google Scholar] [CrossRef]
- Sun, B.; Wang, H.; Lv, J.; Pei, H.; Bai, Z. Predictors of Mortality in Hospitalized COVID-19 Patients Complicated with Hypotension and Hypoxemia: A Retrospective Cohort Study. Front. Med. 2021, 8, 753035. [Google Scholar] [CrossRef]
- Chen, Q.; Xu, L.; Dai, Y.; Ling, Y.; Mao, J.; Qian, J.; Zhu, W.; Di, W.; Ge, J. Cardiovascular manifestations in severe and critical patients with COVID-19. Clin. Cardiol. 2020, 43, 796–802. [Google Scholar] [CrossRef]
- Szebeni, J.; Bedocs, P.; Csukas, D.; Rosivall, L.; Bunger, R.; Urbanics, R. A porcine model of complement-mediated infusion reactions to drug carrier nanosystems and other medicines. Adv. Drug Deliv. Rev. 2012, 64, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Urbanics, R.; Szebeni, J. Lessons learned from the porcine CARPA model: Constant and variable responses to different nanomedicines and administration protocols. Eur. J. Nanomed. 2015, 7, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Szebeni, J.; Bedőcs, P.; Dézsi, L.; Urbanics, R. A porcine model of complement activation-related pseudoallergy to nano-pharmaceuticals: Pros and cons of translation to a preclinical safety test. Precis. Nanomed. 2018, 1, 63–73. [Google Scholar] [CrossRef]
- Szebeni, J.; Bawa, R. Human Clinical Relevance of the Porcine Model of Pseudoallergic Infusion Reactions. Biomedicines 2020, 8, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patkó, Z.; Szebeni, J. Blood cell changes in complement activation-related pseudoallergy. Eur. J. Nanomed. 2015, 7, 233–244. [Google Scholar] [CrossRef]
- von Asmuth, E.J.; Maessen, J.G.; van der Linden, C.J.; Buurman, W.A. Tumour necrosis factor alpha (TNF-alpha) and interleukin 6 in a zymosan-induced shock model. Scand. J. Immunol. 1990, 32, 313–319. [Google Scholar] [CrossRef]
- Mogilski, S.; Kubacka, M.; Łażewska, D.; Więcek, M.; Głuch-Lutwin, M.; Tyszka-Czochara, M.; Bukowska-Strakova, K.; Filipek, B.; Kieć-Kononowicz, K. Aryl-1, 3, 5-triazine ligands of histamine H4 receptor attenuate inflammatory and nociceptive response to carrageen, zymosan and lipopolysaccharide. Inflamm. Res. 2017, 66, 79–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozma, G.T.; Mészáros, T.; Bakos, T.; Hennies, M.; Bencze, D.; Uzonyi, B.; Győrffy, B.; Cedrone, E.; Dobrovolskaia, M.A.; Józsi, M.; et al. Mini-Factor H Modulates Complement-Dependent IL-6 and IL-10 Release in an Immune Cell Culture (PBMC) Model: Potential Benefits against Cytokine Storm. Front. Immunol. 2021, 12, 642860. [Google Scholar] [CrossRef]
- Fülöp, T.; Kozma, G.T.; Vashegyi, I.; Mészáros, T.; Rosivall, L.; Urbanics, R.; Storm, G.; Metselaar, J.M.; Szebeni, J. Liposome-induced hypersensitivity reactions: Risk reduction by design of safe infusion protocols in pigs. J. Control. Release 2019, 309, 333–338. [Google Scholar] [CrossRef]
- Mészáros, T.; Kozma, G.T.; Shimizu, T.; Miyahara, K.; Turjeman, K.; Ishida, T.; Barenholz, Y.; Urbanics, R.; Szebeni, J. Involvement of complement activation in the pulmonary vasoactivity of polystyrene nanoparticles in pigs: Unique surface properties underlying alternative pathway activation and instant opsonization. Int. J. Nanomed. 2018, 13, 6345–6357. [Google Scholar] [CrossRef] [Green Version]
- Jackman, J.A.; Meszaros, T.; Fulop, T.; Urbanics, R.; Szebeni, J.; Cho, N.J. Comparison of complement activation-related pseudoallergy in miniature and domestic pigs: Foundation of a validatable immune toxicity model. Nanomedicine 2016, 12, 933–943. [Google Scholar] [CrossRef]
- Szebeni, J.; Bedőcs, P.; Rozsnyay, Z.; Weiszhár, Z.; Urbanics, R.; Rosivall, L.; Cohen, R.; Garbuzenko, O.; Báthori, G.; Tóth, M.; et al. Liposome-induced complement activation and related cardiopulmonary distress in pigs: Factors promoting reactogenicity of Doxil and AmBisome. Nanomedicine 2012, 8, 176–184. [Google Scholar] [CrossRef] [Green Version]
- Szebeni, J.; Baranyi, L.; Savay, S.; Bodo, M.; Milosevits, J.; Alving, C.R.; Bünger, R. Complement activation-related cardiac anaphylaxis in pigs: Role of C5a anaphylatoxin and adenosine in liposome-induced abnormalities in ECG and heart function. Am. J. Physiol. Heart Circ. Physiol. 2006, 290, H1050–H1058. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, J.; Fontana, J.L.; Wassef, N.M.; Mongan, P.D.; Morse, D.S.; Dobbins, D.E.; Stahl, G.L.; Bünger, R.; Alving, C.R. Hemodynamic changes induced by liposomes and liposome-encapsulated hemoglobin in pigs: A model for pseudoallergic cardiopulmonary reactions to liposomes. Role of complement and inhibition by soluble CR1 and anti-C5a antibody. Circulation 1999, 99, 2302–2309. [Google Scholar] [CrossRef] [Green Version]
- Di Carlo, F.J.; Fiore, J.V. On the composition of zymosan. Science 1958, 127, 756–757. [Google Scholar] [CrossRef]
- Underhill, D.M. Macrophage recognition of zymosan particles. J. Endotoxin Res. 2003, 9, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Horsthemke, M.; Bachg, A.C.; Groll, K.; Moyzio, S.; Müther, B.; Hemkemeyer, S.A.; Wedlich-Söldner, R.; Sixt, M.; Tacke, S.; Bähler, M.; et al. Multiple roles of filopodial dynamics in particle capture and phagocytosis and phenotypes of Cdc42 and Myo10 deletion. J. Biol. Chem. 2017, 292, 7258–7273. [Google Scholar] [CrossRef] [Green Version]
- Pillemer, L.; Ecker, E.E. Anticomplementary factor in fresh yeast. J. Biol. Chem. 1941, 137, 139–142. [Google Scholar] [CrossRef]
- Sato, M.; Sano, H.; Iwaki, D.; Kudo, K.; Konishi, M.; Takahashi, H.; Takahashi, T.; Imaizumi, H.; Asai, Y.; Kuroki, Y. Direct binding of Toll-like receptor 2 to zymosan, and zymosan-induced NF-kappa B activation and TNF-alpha secretion are down-regulated by lung collectin surfactant protein A. J. Immunol. 2003, 171, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liang, X.; Bao, X.; Xiao, W.; Chen, G. Toll-like receptor 4 (TLR4) inhibitors: Current research and prospective. Eur. J. Med. Chem. 2022, 235, 114291. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.; Brown, G.; Reid, D.M.; Willment, J.; Martinez-Pomares, L.; Gordon, S.; Wong, S.Y.C. The beta-glucan receptor, dectin-1, is predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J. Immunol. 2002, 169, 3876–3882. [Google Scholar] [CrossRef]
- Brown, G.D.; Taylor, P.R.; Reid, D.M.; Willment, J.A.; Williams, D.L.; Martinez-Pomares, L.; Wong, S.Y.C.; Gordon, S. Dectin-1 is a major beta-glucan receptor on macrophages. J. Exp. Med. 2002, 196, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; O’Callaghan, C.A.; Marshall, A.S.; Gilbert, R.J.; Siebold, C.; Gordon, S.; Brown, G.D.; Jones, E.Y. Structure of the fungal beta-glucan-binding immune receptor dectin-1: Implications for function. Protein Sci. 2007, 16, 1042–1052. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef]
- Goodridge, H.S.; Simmons, R.M.; Underhill, D.M. Dectin-1 stimulation by Candida albicans yeast or zymosan triggers NFAT activation in macrophages and dendritic cells. J. Immunol. 2007, 178, 3107–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, B.-K.; Chun, E.; Choi, J.J.; Shin, Y.; Kho, Y.T.; Oh, S.H.; Kim, S.Y.; Lee, T.H.; Kim, T.-W.; Shin, E.; et al. Administration of Wasabia koreana Ameliorates Irritable Bowel Syndrome-Like Symptoms in a Zymosan-Induced Mouse Model. J. Med. Food 2017, 20, 474–484. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, D.-Y.; Kim, J.; Woo, K.I.; Kim, Y.-D.; Han, J.; Chung, T.-Y.; Cha, H.-S.; Lim, D.H. Enhanced orbital adipogenesis in a mouse model of T-cell-mediated autoimmunity, zymosan A-treated SKG mice: Implications for Graves’ ophthalmopathy. Sci. Rep. 2020, 10, 7329. [Google Scholar] [CrossRef]
- Young, S.H.; Wolfarth, M.G.; Roberts, J.R.; Kashon, M.L.; Antonini, J.M. Adjuvant effect of zymosan after pulmonary treatment in a mouse ovalbumin allergy model. Exp. Lung Res. 2013, 39, 48–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahat, M.A.; Brod, V.; Amit-Cohen, B.C.; Henig, O.; Younis, S.; Bitterman, H. Oxygen Mitigates the Inflammatory Response in a Model of Hemorrhage and Zymosan-Induced Inflammation. Shock 2016, 45, 198–208. [Google Scholar] [CrossRef]
- Sugimoto, Y.; Endo, D.; Aratani, Y. Mice Deficient in NOX2 Display Severe Thymic Atrophy, Lymphopenia, and Reduced Lymphopoiesis in a Zymosan-Induced Model of Systemic Inflammation. Inflammation 2021, 44, 371–382. [Google Scholar] [CrossRef]
- Frasnelli, M.E.; Tarussio, D.; Chobaz-Peclat, V.; Busso, N.; So, A. TLR2 modulates inflammation in zymosan-induced arthritis in mice. Arthritis Res. Ther. 2005, 7, R370–R379. [Google Scholar] [CrossRef]
- Xie, K.; Yu, Y.; Zhang, Z.; Liu, W.; Pei, Y.; Xiong, L.; Hou, L.; Wang, G. Hydrogen gas improves survival rate and organ damage in zymosan-induced generalized inflammation model. Shock 2010, 34, 495–501. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.-Y.; Kwon, Y.-B.; Kim, H.-W.; Roh, D.-H.; Kang, S.-Y.; Kim, C.-Y.; Han, H.J.; Kim, K.-W.; Yang, I.-S.; Beitz, A.J.; et al. Intrathecal neostigmine reduces the zymosan-induced inflammatory response in a mouse air pouch model via adrenomedullary activity: Involvement of spinal muscarinic type 2 receptors. Neuropharmacology 2005, 49, 275–282. [Google Scholar] [CrossRef]
- Volman, T.J.; Goris, R.J.; Hendriks, T. Pentoxifylline does not improve outcome in a murine model for the multiple-organ dysfunction syndrome. Intensive Care Med. 2005, 31, 701–708. [Google Scholar] [CrossRef]
- Mizuno, M.; Ito, Y.; Hepburn, N.; Mizuno, T.; Noda, Y.; Yuzawa, Y.; Harris, C.L.; Morgan, B.P.; Matsuo, S. Zymosan, but not lipopolysaccharide, triggers severe and progressive peritoneal injury accompanied by complement activation in a rat peritonitis model. J. Immunol. 2009, 183, 1403–1412. [Google Scholar] [CrossRef] [Green Version]
- Chaves, H.V.; de Albuquerque Ribeiro, R.; de Souza, A.M.B.; e Silva, A.A.R.; Gomes, A.S.; Vale, M.L.; Bezerra, M.M.; de Castro Brito, G.A. Experimental model of zymosan-induced arthritis in the rat temporomandibular joint: Role of nitric oxide and neutrophils. J. Biomed. Biotechnol. 2011, 2011, 707985. [Google Scholar] [CrossRef] [PubMed]
- Dief, A.E.; Mostafa, D.K.; Sharara, G.M.; Zeitoun, T.H. Hydrogen sulfide releasing naproxen offers better anti-inflammatory and chondroprotective effect relative to naproxen in a rat model of zymosan induced arthritis. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 1537–1546. [Google Scholar] [PubMed]
- Paya, M.; Terencio, M.C.; Ferrandiz, M.L.; Alcaraz, M.J. Involvement of secretory phospholipase A2 activity in the zymosan rat air pouch model of inflammation. Br. J. Pharmacol. 1996, 117, 1773–1779. [Google Scholar] [CrossRef] [Green Version]
- Aydogdu, O.; Gocun, P.U.; Aronsson, P.; Carlsson, T.; Winder, M. Prostate-to-bladder cross-sensitization in a model of zymosan-induced chronic pelvic pain syndrome in rats. Prostate 2021, 81, 252–260. [Google Scholar] [CrossRef]
- Lu, Q.Y.; Zhou, Y.Y.; Wang, J.B.; Wang, L.; Meng, L.; Weng, J.K.; Yu, B.; Quan, S. [Preparation of rat model of systemic inflammatory response syndrome induced by zymosan]. Zhejiang Da Xue Xue Bao Yi Xue Ban 2011, 40, 641–646. [Google Scholar] [PubMed]
- Yamamoto, S.; Kubota, Y.; Tsuji, K.; Yanagitani, K.; Takaoka, M.; Kin, H.; Ogura, M.; Inoue, K. Effect of obstructive jaundice on neutrophil chemotactic activity: An in vivo assessment in zymosan-induced peritonitis model in rats. J. Gastroenterol. Hepatol. 1998, 13, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Unsal, D.; Kacan, M.; Temiz-Resitoglu, M.; Guden, D.; Korkmaz, B.; Sari, A.; Buharalioglu, C.; Yildirim-Yaroglu, H.; Tamer-Gumus, L.; Tunctan, B.; et al. The role of Syk/IkB-alpha/NF-kB pathway activation in the reversal effect of BAY 61-3606, a selective Syk inhibitor, on hypotension and inflammation in a rat model of zymosan-induced non-septic shock. Clin. Exp. Pharmacol. Physiol. 2018, 45, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Cron, R.Q. No perfect therapy for the imperfect COVID-19 cytokine storm. Lancet Rheumatol. 2022, 4, e308–e310. [Google Scholar] [CrossRef]
- Abusalah, M.A.H.; Khalifa, M.; Al-Hatamleh, M.A.I.; Jarrar, M.; Mohamud, R.; Chan, Y.Y. Nucleic Acid-Based COVID-19 Therapy Targeting Cytokine Storms: Strategies to Quell the Storm. J. Pers. Med. 2022, 12, 386. [Google Scholar] [CrossRef]
- Zanza, C.; Romenskaya, T.; Manetti, A.C.; Franceschi, F.; La Russa, R.; Bertozzi, G.; Maiese, A.; Savioli, G.; Volonnino, G.; Longhitano, Y. Cytokine Storm in COVID-19: Immunopathogenesis and Therapy. Medicina 2022, 58, 144. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Rubin, L.; Peng, T.; Liu, L.; Xing, X.; Lazarovici, P.; Zheng, W. Cytokine storm in COVID-19: From viral infection to immune responses, diagnosis and therapy. Int. J. Biol. Sci. 2022, 18, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Fan, J.; Liu, E. Animal Models for COVID-19 Therapeutic Development: Where We Are and Where We Need to Go. Front. Microbiol. 2022, 13, 907406. [Google Scholar] [CrossRef]
- Sefik, E.; Israelow, B.; Mirza, H.; Zhao, J.; Qu, R.; Kaffe, E.; Song, E.; Halene, S.; Meffre, E.; Kluger, Y.; et al. A humanized mouse model of chronic COVID-19. Nat. Biotechnol. 2022, 40, 906–920. [Google Scholar] [CrossRef]
- Fan, C.; Wu, Y.; Rui, X.; Yang, Y.; Ling, C.; Liu, S.; Liu, S.; Wang, Y. Animal models for COVID-19: Advances, gaps and perspectives. Signal Transduct. Target. Ther. 2022, 7, 220. [Google Scholar] [CrossRef]
- Park, J.G.; Pino, P.A.; Akhter, A.; Alvarez, X.; Torrelles, J.B.; Martinez-Sobrido, L. Animal Models of COVID-19: Transgenic Mouse Model. Methods Mol. Biol. 2022, 2452, 259–289. [Google Scholar]
- Singh, D.K.; Cole, J.; Escobedo, R.A.; Alfson, K.J.; Singh, B.; Lee, T.-H.; Alvarez, X.; Ganatra, S.R.; Carrion, J.R.; Kaushal, D. Animal Models of COVID-19: Nonhuman Primates. Methods Mol. Biol. 2022, 2452, 227–258. [Google Scholar] [PubMed]
- Stich, N.; Waclavicek, M.; Model, N.; Eibl, M.M. Staphylococcal superantigen (TSST-1) mutant analysis reveals that t cell activation is required for biological effects in the rabbit including the cytokine storm. Toxins 2010, 2, 2272–2288. [Google Scholar] [CrossRef] [PubMed]
- Hod, E.A.; Cadwell, C.M.; Liepkalns, J.S.; Zimring, J.C.; Sokol, S.A.; Schirmer, D.A.; Jhang, J.; Spitalnik, S.L. Cytokine storm in a mouse model of IgG-mediated hemolytic transfusion reactions. Blood 2008, 112, 891–894. [Google Scholar] [CrossRef] [PubMed]
- Rosinski, S.L.; Storb, R.; Strong, R.K.; Sale, G.E.; Stone, D.M.; Gewe, M.M.; Friend, D.J.; Abrams, V.K.; Randolph-Habecker, J.; Graves, S.S. Anti-CD28 Antibody-Initiated Cytokine Storm in Canines. Transplant. Direct 2015, 1, 1–11. [Google Scholar] [CrossRef]
- Keating, S.M.; Heitman, J.W.; Wu, S.; Deng, X.; Stacey, A.R.; Zahn, R.C.; de la Rosa, M.; Finstad, S.L.; Lifson, J.D.; Piatak, M.; et al. Magnitude and Quality of Cytokine and Chemokine Storm during Acute Infection Distinguish Nonprogressive and Progressive Simian Immunodeficiency Virus Infections of Nonhuman Primates. J. Virol. 2016, 90, 10339–10350. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, J.; Zhang, Y.; Yang, J.; Wang, L.; Qi, Y.; Han, X.; Zhou, X.; Miao, F.; Chen, T.; et al. Cytokine Storm in Domestic Pigs Induced by Infection of Virulent African Swine Fever Virus. Front. Vet. Sci. 2020, 7, 601641. [Google Scholar] [CrossRef]
- Xu, X.; Jia, C.; Luo, S.; Li, Y.; Xiao, F.; Dai, H.; Wang, C. Effect of HA330 resin-directed hemoadsorption on a porcine acute respiratory distress syndrome model. Ann. Intensive Care 2017, 7, 84. [Google Scholar] [CrossRef] [Green Version]
- Csukas, D.; Urbanics, R.; Weber, G.; Rosivall, L.; Szebeni, J. Pulmonary intravascular macrophages: Prime suspects as cellular mediators of porcine CARPA. Eur. J. Nanomed. 2015, 7, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.; Park, J.Y.; Lee, J.; Lim, J.-H.; Shin, E.-C.; Ahn, Y.S.; Kim, C.-H.; Kim, S.-J.; Kim, J.-D.; Choi, I.S.; et al. Fas ligand and Fas are expressed constitutively in human astrocytes and the expression increases with IL-1, IL-6, TNF-alpha, or IFN-gamma. J. Immunol. 1999, 162, 1889–1895. [Google Scholar] [CrossRef]
- Maeda, K.; Baba, Y.; Nagai, Y.; Miyazaki, K.; Malykhin, A.; Nakamura, K.; Kincade, P.W.; Sakaguchi, N.; Coggeshall, K.M. IL-6 blocks a discrete early step in lymphopoiesis. Blood 2005, 106, 879–885. [Google Scholar] [CrossRef] [Green Version]
- Sordillo, P.P.; Helson, L. Curcumin suppression of cytokine release and cytokine storm. A potential therapy for patients with Ebola and other severe viral infections. In Vivo 2015, 29, 1–4. [Google Scholar] [PubMed]
- Lee, D.W.; A Gardner, R.; Porter, D.L.; Louis, C.U.; Ahmed, N.; Jensen, M.C.; Grupp, S.A.; Mackall, C.L. Current concepts in the diagnosis and management of cytokine release syndrome. Blood 2014, 124, 188–195. [Google Scholar] [CrossRef]
- Kroschinsky, F.; Stölzel, F.; von Bonin, S.; Beutel, G.; Kochanek, M.; Kiehl, M.; Schellongowski, S.; on behalf of the Intensive Care in Hematological; Oncological Patients (iCHOP) Collaborative Group. New drugs, new toxicities: Severe side effects of modern targeted and immunotherapy of cancer and their management. Crit. Care 2017, 21, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuhof, H. Actions and interactions of mediator systems and mediators in the pathogenesis of ARDS and multiorgan failure. Acta Anaesthesiol. Scand. Suppl. 1991, 95, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Kollias, G.; Douni, E.; Kassiotis, G.; Kontoyiannis, D. On the role of tumor necrosis factor and receptors in models of multiorgan failure, rheumatoid arthritis, multiple sclerosis and inflammatory bowel disease. Immunol. Rev. 1999, 169, 175–194. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal | Disease | Reference |
|---|---|---|
| mouse | irritable bowel syndrome | [56] |
| T-cell-mediated autoimmune response | [57] | |
| allergy | [58] | |
| systemic inflammation | [59,60] | |
| arthritis | [61] | |
| sepsis | [62] | |
| air pouch model of inflammation | [63] | |
| multiple organ dysfunction syndrome (MODS) | [64] | |
| septic shock | [36] | |
| rat | peritonitis | [65] |
| arthritis | [66,67] | |
| air pouch model of inflammation | [68] | |
| chronic pelvic pain syndrome | [69] | |
| systemic inflammatory response syndrome (SIRS) | [70] | |
| peritonitis | [71] | |
| non-septic shock | [72] |
| Gene Symbol | Forward Primer | Reverse Primer |
|---|---|---|
| ssRN18S (NR_046261) | GACAAATCGCTCCACCAACT | CCTGCGGCTTAATTTGACTC |
| ssIL1RA (NM_214262) | CAAGCCTTCAGAATCTGGGATGTC | GGCTCAACAGGCACCACATC |
| ssCCL2 (NM_214214) | GAAGCAGTGATCTTCAAGAC | GGGCAAGTTAGAAGGAAATG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kökény, G.; Bakos, T.; Barta, B.A.; Nagy, G.V.; Mészáros, T.; Kozma, G.T.; Szabó, A.; Szebeni, J.; Merkely, B.; Radovits, T. Zymosan Particle-Induced Hemodynamic, Cytokine and Blood Cell Changes in Pigs: An Innate Immune Stimulation Model with Relevance to Cytokine Storm Syndrome and Severe COVID-19. Int. J. Mol. Sci. 2023, 24, 1138. https://doi.org/10.3390/ijms24021138
Kökény G, Bakos T, Barta BA, Nagy GV, Mészáros T, Kozma GT, Szabó A, Szebeni J, Merkely B, Radovits T. Zymosan Particle-Induced Hemodynamic, Cytokine and Blood Cell Changes in Pigs: An Innate Immune Stimulation Model with Relevance to Cytokine Storm Syndrome and Severe COVID-19. International Journal of Molecular Sciences. 2023; 24(2):1138. https://doi.org/10.3390/ijms24021138
Chicago/Turabian StyleKökény, Gábor, Tamás Bakos, Bálint András Barta, Georgina Viktória Nagy, Tamás Mészáros, Gergely T. Kozma, András Szabó, János Szebeni, Béla Merkely, and Tamás Radovits. 2023. "Zymosan Particle-Induced Hemodynamic, Cytokine and Blood Cell Changes in Pigs: An Innate Immune Stimulation Model with Relevance to Cytokine Storm Syndrome and Severe COVID-19" International Journal of Molecular Sciences 24, no. 2: 1138. https://doi.org/10.3390/ijms24021138