

JUL1, Ring-Type E3 Ubiquitin Ligase, Is Involved in Transcriptional Reprogramming for ERF15-Mediated Gene Regulation
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
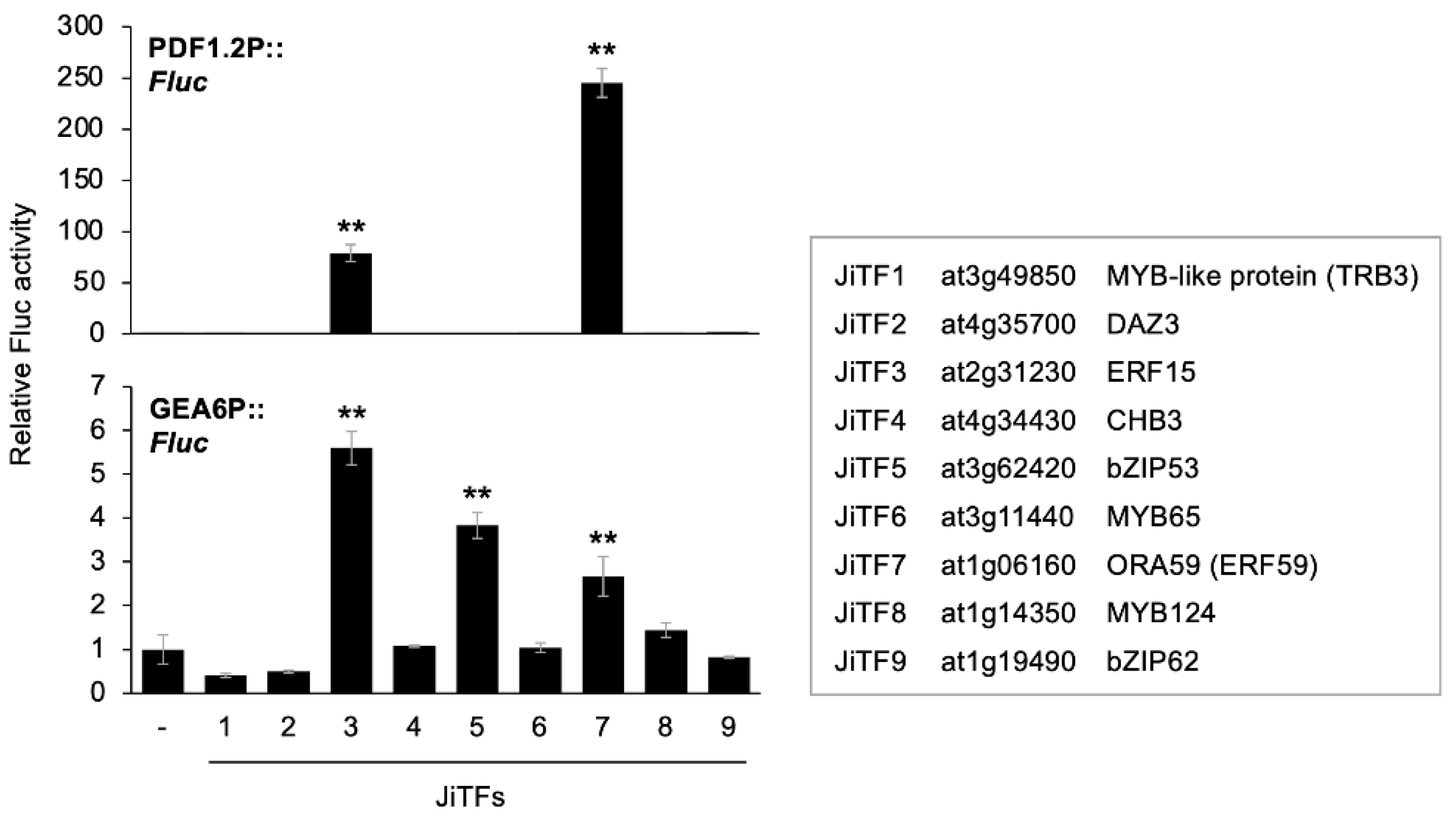
2.1. Transcriptional Activation of PDF1.2 and GEA6 Promoters through JiTFs
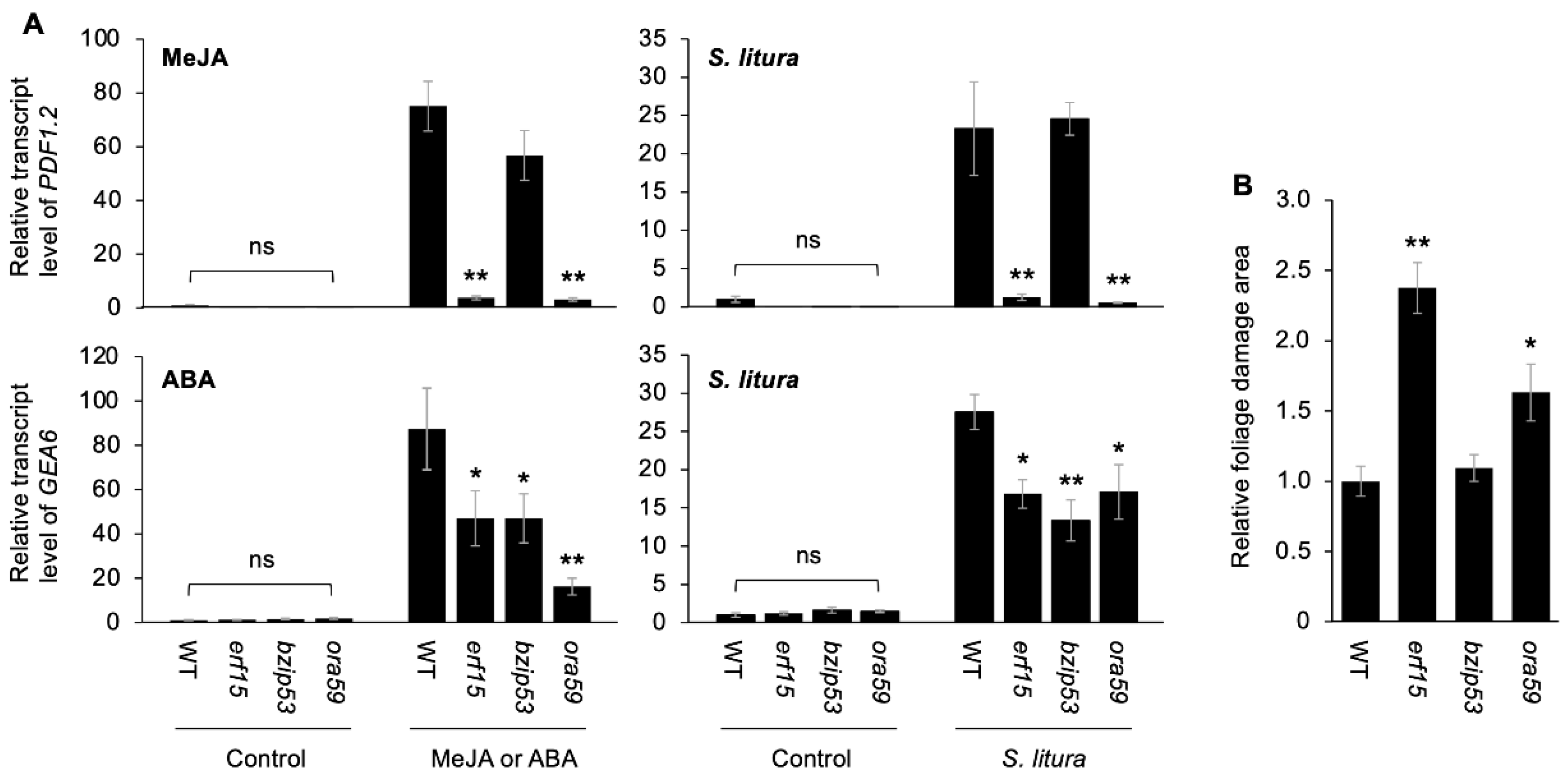
2.2. Anti-Herbivore and JA/ABA Responses Mediated by ERF15, bZIP53 and ORA59
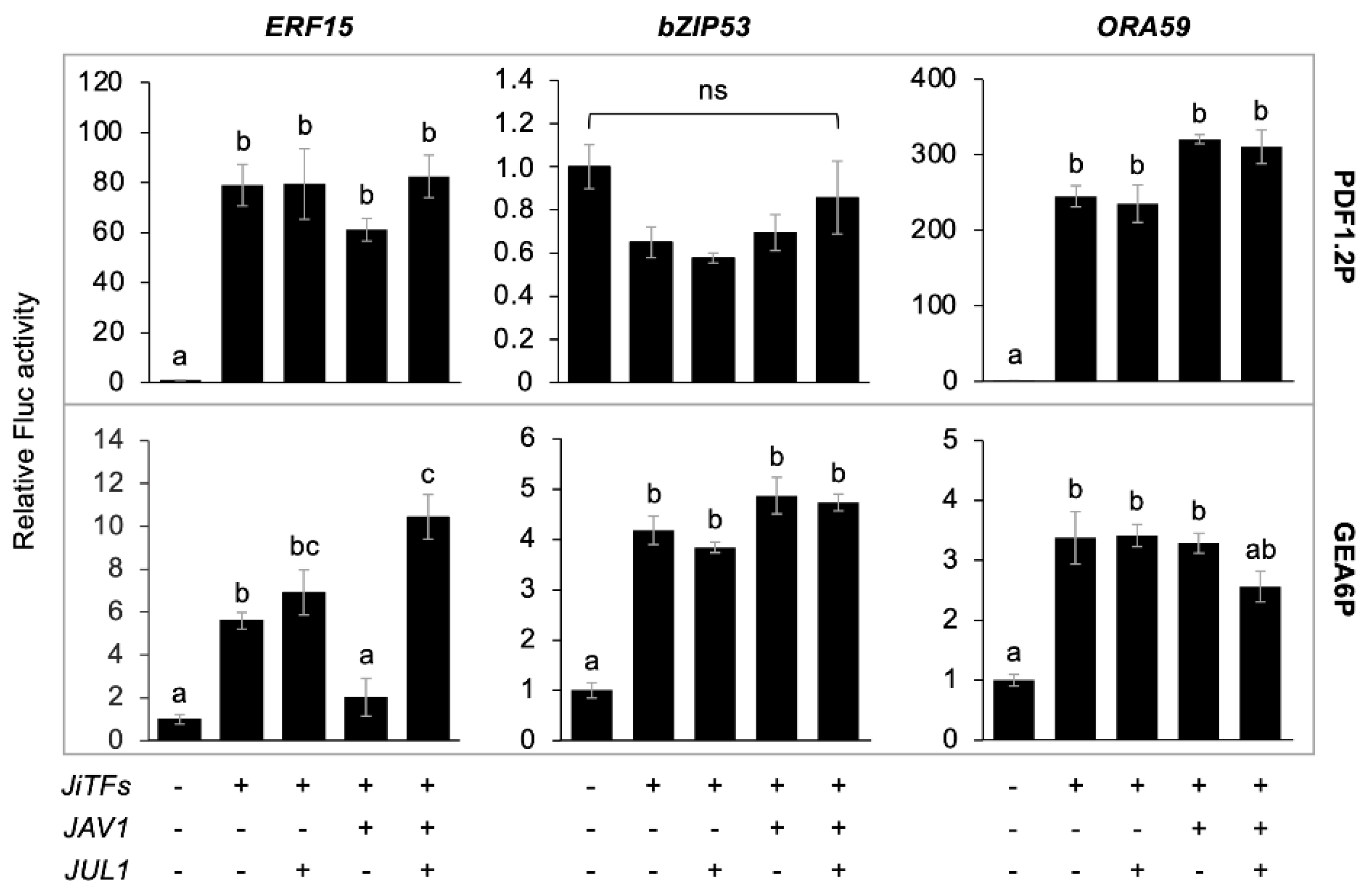
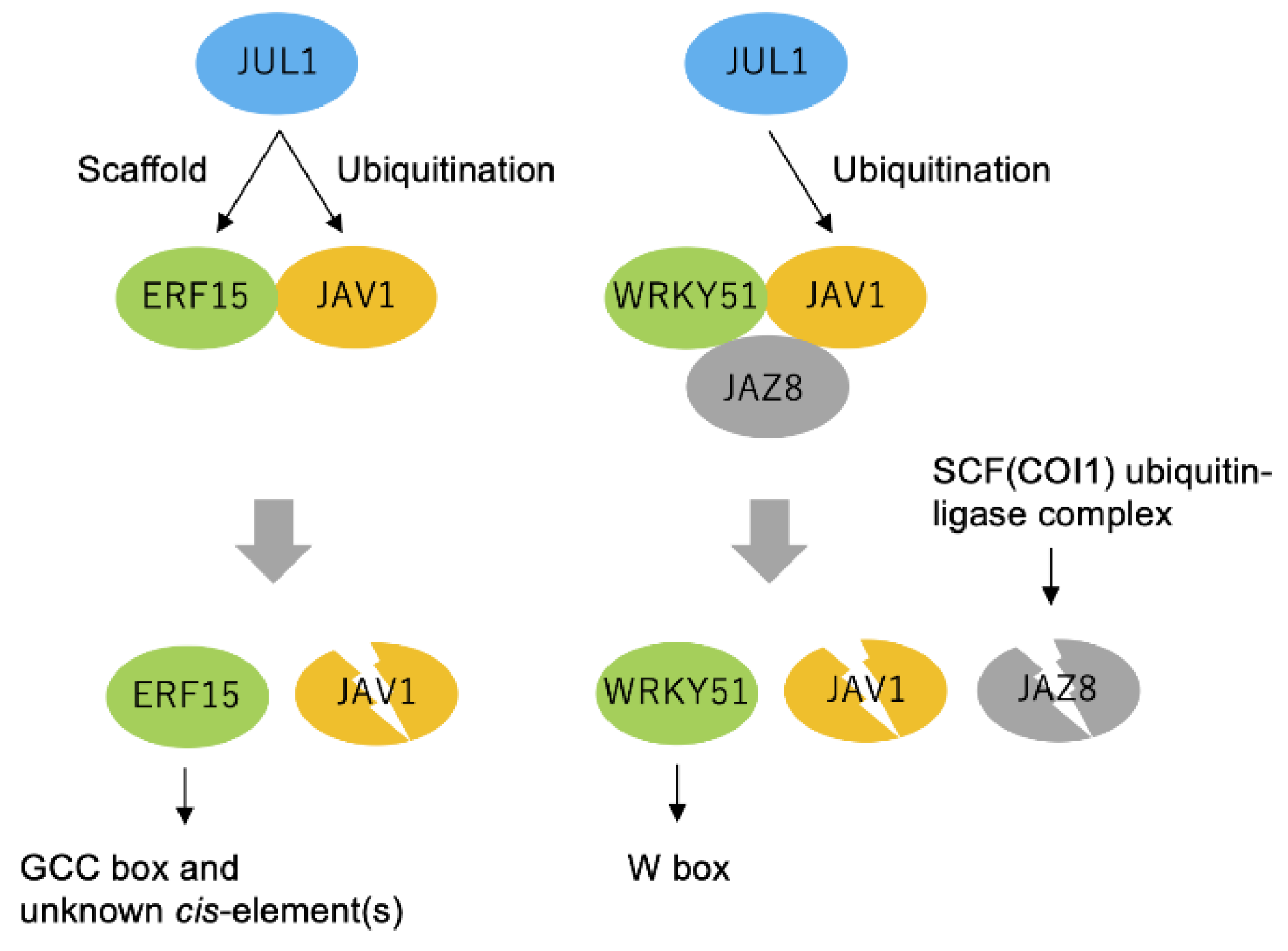
2.3. Transcriptional Reprogramming with JAV1, JUL1 and JiTFs
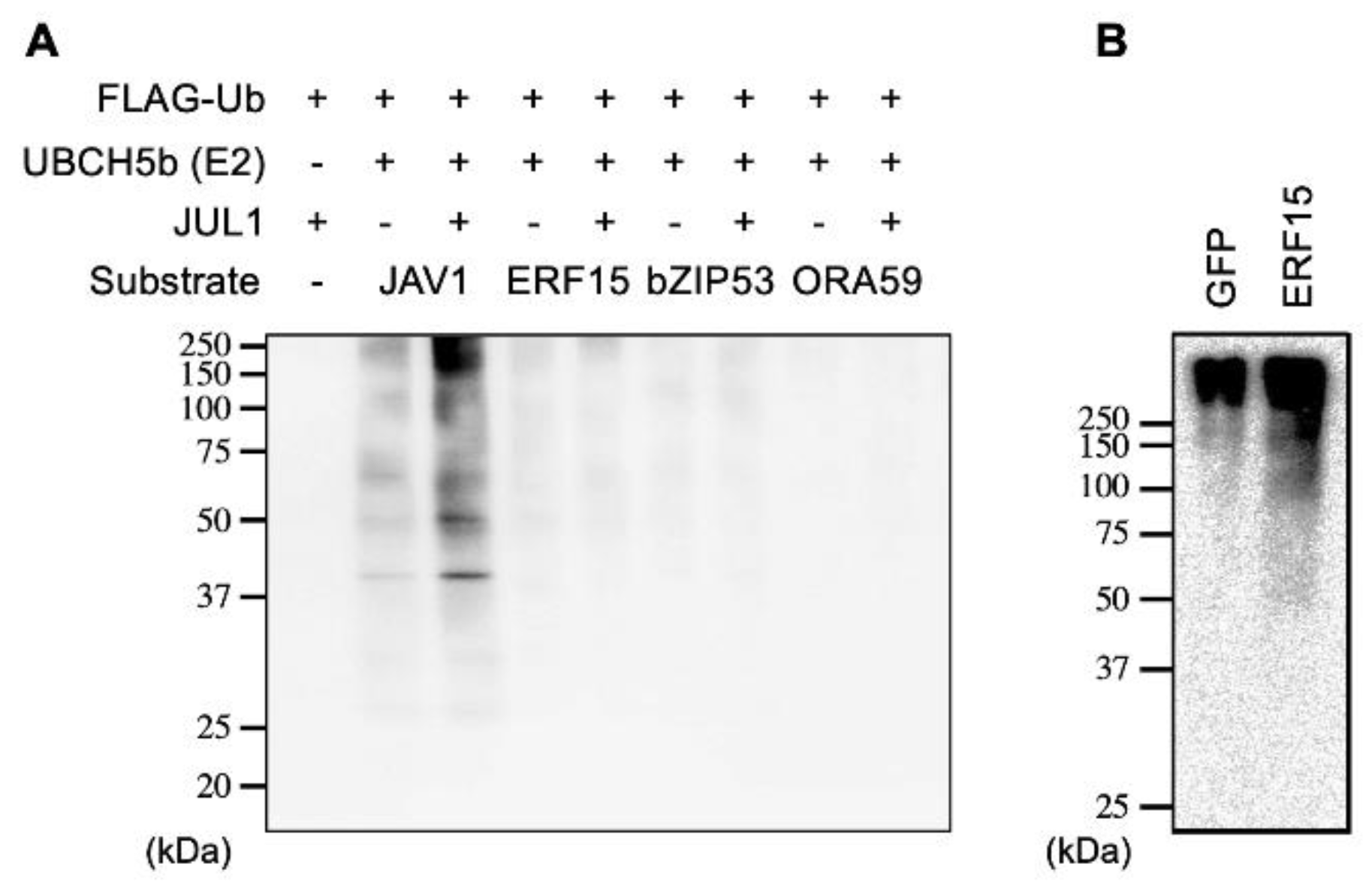
2.4. Not Enhanced Ubiquitination of JAV1 by JUL1
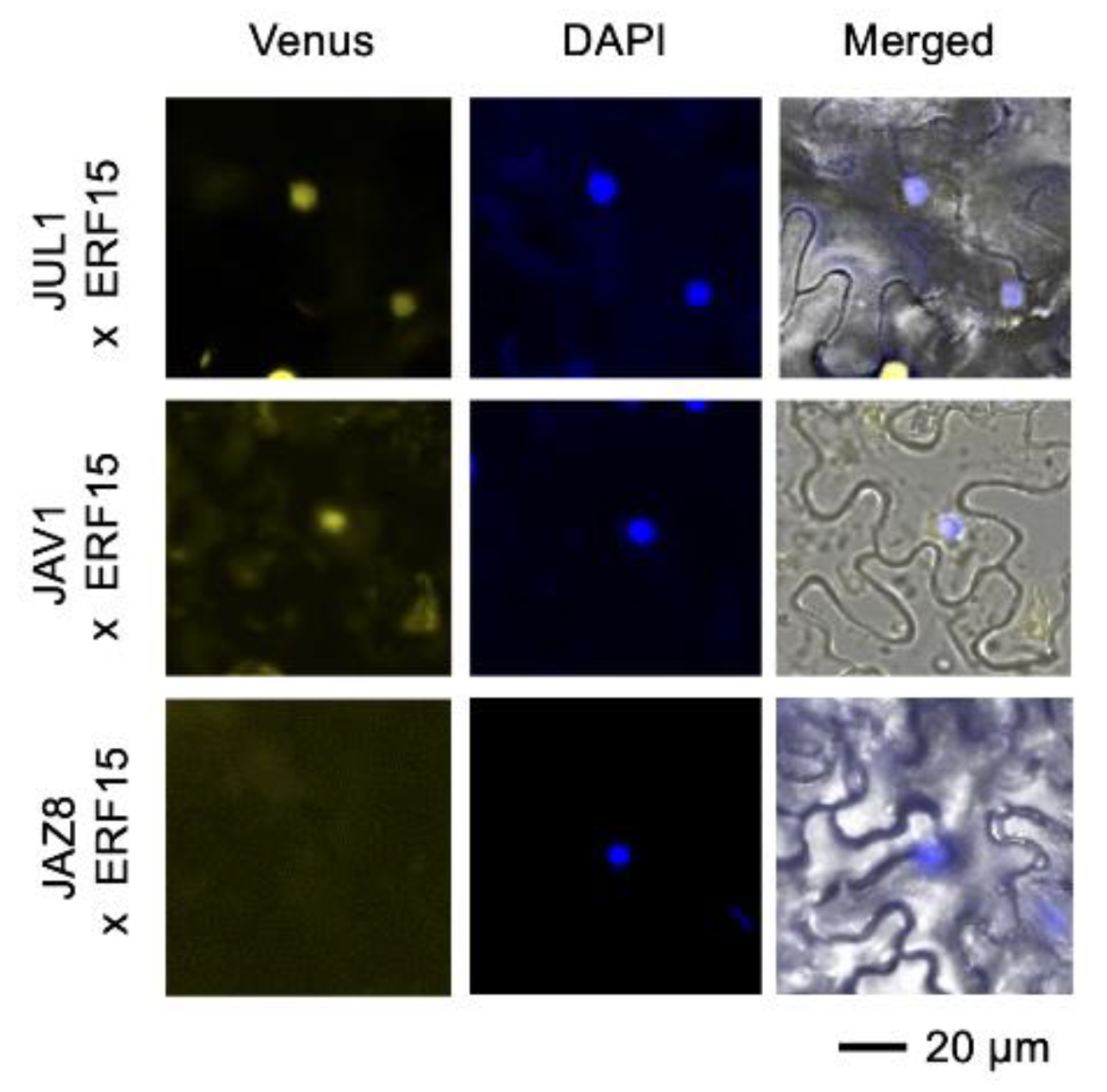
2.5. In Planta Complex Composed of JAV1, JUL1 and ERF15
3. Discussion
4. Materials and Methods
4.1. Primers
4.2. Cell-Free Protein Synthesis, AlphaScreen Assay and Immunoblotting
4.3. Plants
4.4. Chemical and Herbivore Treatment
4.5. Protoplast Preparation and Transfection
4.6. Luciferase (LUC) Assay
4.7. RNA Extraction, cDNA Synthesis and qPCR
4.8. BiFC
4.9. In Vitro Ubiquitination Assay
4.10. Statistical Analyses and Data Reproducibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campos, M.L.; Yoshida, Y.; Major, I.T.; de Oliveira Ferreira, D.; Weraduwage, S.M.; Froehlich, J.E.; Johnson, B.F.; Kramer, D.M.; Jander, G.; Sharkey, T.D.; et al. Rewiring of jasmonate and phytochrome B signalling uncouples plant growth-defense tradeoffs. Nat. Commun. 2016, 7, 12570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, F.; Melotto, M.; Yao, J.; He, S.Y. Jasmonate signaling and manipulation by pathogens and insects. J. Exp. Bot. 2017, 68, 1371–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhou, W.; Cheng, Z.; Fan, M.; Wang, L.; Xie, D. JAV1 controls jasmonate-regulated plant defense. Mol. Cell 2013, 50, 504–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.; Fan, M.; Yang, M.; Zhao, J.; Zhang, W.; Su, Y.; Xiao, L.; Deng, H.; Xie, D. Injury activates Ca(2+)/calmodulin-dependent phosphorylation of JAV1-JAZ8-WRKY51 complex for jasmonate biosynthesis. Mol. Cell 2018, 70, 136–149.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.R.M.; Uemura, T.; Ramadan, A.; Adachi, K.; Nemoto, K.; Nozawa, A.; Hoshino, R.; Abe, H.; Sawasaki, T.; Arimura, G. The ring-type E3 ubiquitin ligase JUL1 targets the VQ-motif protein JAV1 to coordinate jasmonate signaling. Plant Physiol. 2019, 179, 1273–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.G.; Kim, J.H.; Cho, N.H.; Oh, T.R.; Kim, W.T. Arabidopsis RING E3 ubiquitin ligase JUL1 participates in ABA-mediated microtubule depolymerization, stomatal closure, and tolerance response to drought stress. Plant J. 2020, 103, 824–842. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, Y.; Yang, Y.; Chi, Y.J.; Zhou, J.; Chen, J.Y.; Wang, F.; Fan, B.; Shi, K.; Zhou, Y.H.; et al. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012, 159, 810–825. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Brambila, J.; Meagher, R.L. Use of DNA barcodes to identify invasive armyworm Spodoptera species in Florida. J. Insect Sci. 2011, 11, 154. [Google Scholar] [CrossRef] [Green Version]
- Manners, J.M.; Penninckx, I.A.M.A.; Vermaere, K.; Kazan, K.; Brown, R.L.; Morgan, A.; Maclean, D.J.; Curtis, M.D.; Cammue, B.P.A.; Broekaert, W.F. The promoter of the plant defensin gene PDF1.2 from Arabidopsis is systemically activated by fungal pathogens and responds to methyl jasmonate but not to salicylic acid. Plant Mol. Biol. 1998, 38, 1071–1080. [Google Scholar] [CrossRef]
- Manfre, A.J.; LaHatte, G.A.; Climer, C.R.; Marcotte, W.R., Jr. Seed dehydration and the establishment of desiccation tolerance during seed maturation is altered in the Arabidopsis thaliana mutant atem6-1. Plant Cell Physiol. 2009, 50, 243–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, K.; Abe, H.; Arimura, G. Jasmonates induce both defense responses and communication in monocotyledonous and dicotyledonous plants. Plant Cell Physiol. 2015, 56, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Sumioka, H.; Takiguchi, M.; Uemura, T.; Kihara, Y.; Shinya, T.; Galis, I.; Arimura, G.I. Phytohormone-dependent plant defense signaling orchestrated by oral bacteria of the herbivore Spodoptera litura. New Phytol. 2021, 231, 2029–2038. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Lin, R. The VQ motif-containing protein family of plant-specific transcriptional regulators. Plant Physiol. 2015, 169, 371–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Lal, N.K.; Lin, Z.D.; Ma, S.; Liu, J.; Castro, B.; Toruno, T.; Dinesh-Kumar, S.P.; Coaker, G. Regulation of reactive oxygen species during plant immunity through phosphorylation and ubiquitination of RBOHD. Nat. Commun. 2020, 11, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spoel, S.H.; Mou, Z.; Tada, Y.; Spivey, N.W.; Genschik, P.; Dong, X. Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 2009, 137, 860–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yahya, G.; Wu, Y.; Peplowska, K.; Röhrl, J.; Soh, Y.M.; Bürmann, F.; Gruber, S.; Storchova, Z. Phospho-regulation of the Shugoshin—Condensin interaction at the centromere in budding yeast. PLoS Genet. 2020, 16, e1008569. [Google Scholar] [CrossRef]
- Parisi, E.; Yahya, G.; Flores, A.; Aldea, M. Cdc48/p97 segregase is modulated by cyclin-dependent kinase to determine cyclin fate during G1 progression. EMBO J. 2018, 37, e98724. [Google Scholar] [CrossRef]
- Fujimoto, S.Y.; Ohta, M.; Usui, A.; Shinshi, H.; Ohme-Takagi, M. Arabidopsis ethylene-responsive element binding factors act as transcriptional activators or repressors of GCC box-mediated gene expression. Plant Cell 2000, 12, 393–404. [Google Scholar]
- Chen, H.Y.; Hsieh, E.J.; Cheng, M.C.; Chen, C.Y.; Hwang, S.Y.; Lin, T.P. ORA47 (octadecanoid-responsive AP2/ERF-domain transcription factor 47) regulates jasmonic acid and abscisic acid biosynthesis and signaling through binding to a novel cis-element. New Phytol. 2016, 211, 599–613. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.Y.; Lin, P.H.; Chen, K.H.; Cheng, Y.S. Structural insights into Arabidopsis ethylene response factor 96 with an extended N-terminal binding to GCC box. Plant Mol. Biol. 2020, 104, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Nozawa, A.; Matsubara, Y.; Tanaka, Y.; Takahashi, H.; Akagi, T.; Seki, M.; Shinozaki, K.; Endo, Y.; Sawasaki, T. Construction of a protein library of Arabidopsis transcription factors using a wheat cell-free protein production system and its application for DNA binding analysis. Biosci. Biotechnol. Biochem. 2009, 73, 1661–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, T.; Takeda, H.; Uematsu, A.; Yamanaka, S.; Nomura, S.; Nemoto, K.; Iwasaki, T.; Takahashi, H.; Sawasaki, T. AGIA tag system based on a high affinity rabbit monoclonal antibody against human dopamine receptor D1 for protein analysis. PLoS ONE 2016, 11, e0156716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Han, X.; Yang, M.; Zhang, M.; Pan, J.; Yu, D. The transcription factor Inducer of cbf expression1 interacts with abscisic acid insensitive5 and della proteins to fine-tune abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2019, 31, 1520–1538. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, T.; Uemura, T.; Nemoto, K.; Daito, M.; Nozawa, A.; Sawasaki, T.; Arimura, G. Tyrosine kinase-dependent defense responses against herbivory in Arabidopsis. Front. Plant Sci. 2019, 10, 776. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, K.; Suzuki, H.; Uemura, T.; Nozawa, A.; Desaki, Y.; Hoshino, R.; Yoshida, A.; Abe, H.; Nishiyama, M.; Nishiyama, C.; et al. Immune gene activation by NPR and TGA transcriptional regulators in the model monocot Brachypodium distachyon. Plant J. 2022, 110, 470–481. [Google Scholar] [CrossRef]
- Yoo, S.D.; Cho, Y.H.; Sheen, J. Arabidopsis mesophyll protoplasts: A versatile cell system for transient gene expression analysis. Nat. Protoc. 2007, 2, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Uemura, T.; Yashiro, T.; Oda, R.; Shioya, N.; Nakajima, T.; Hachisu, M.; Kobayashi, S.; Nishiyama, C.; Arimura, G. Intestinal Anti-Inflammatory Activity of Perillaldehyde. J. Agric. Food Chem. 2018, 66, 3443–3448. [Google Scholar] [CrossRef]
- Kuai, X.; MacLeod, B.J.; Despres, C. Integrating data on the Arabidopsis NPR1/NPR3/NPR4 salicylic acid receptors; a differentiating argument. Front. Plant Sci. 2015, 6, 235. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, A.; Nemoto, K.; Seki, M.; Shinozaki, K.; Takeda, H.; Takahashi, H.; Sawasaki, T. Wheat germ-based protein libraries for the functional characterisation of the Arabidopsis E2 ubiquitin conjugating enzymes and the RING-type E3 ubiquitin ligase enzymes. BMC Plant Biol. 2015, 15, 275. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawaguchi, J.; Hayashi, K.; Desaki, Y.; Ramadan, A.; Nozawa, A.; Nemoto, K.; Sawasaki, T.; Arimura, G.-i. JUL1, Ring-Type E3 Ubiquitin Ligase, Is Involved in Transcriptional Reprogramming for ERF15-Mediated Gene Regulation. Int. J. Mol. Sci. 2023, 24, 987. https://doi.org/10.3390/ijms24020987
Kawaguchi J, Hayashi K, Desaki Y, Ramadan A, Nozawa A, Nemoto K, Sawasaki T, Arimura G-i. JUL1, Ring-Type E3 Ubiquitin Ligase, Is Involved in Transcriptional Reprogramming for ERF15-Mediated Gene Regulation. International Journal of Molecular Sciences. 2023; 24(2):987. https://doi.org/10.3390/ijms24020987
Chicago/Turabian StyleKawaguchi, Junna, Kaito Hayashi, Yoshitake Desaki, Abdelaziz Ramadan, Akira Nozawa, Keiichirou Nemoto, Tatsuya Sawasaki, and Gen-ichiro Arimura. 2023. "JUL1, Ring-Type E3 Ubiquitin Ligase, Is Involved in Transcriptional Reprogramming for ERF15-Mediated Gene Regulation" International Journal of Molecular Sciences 24, no. 2: 987. https://doi.org/10.3390/ijms24020987