
Influence of Two Widely Used Solvents, Ethanol and Dimethyl Sulfoxide, on Human Sperm Parameters

Abstract
:1. Introduction
2. Results
2.1. Parameters of Sperm Samples Included in This Study
2.2. Influence of EtOH and DMSO on Sperm Parameters
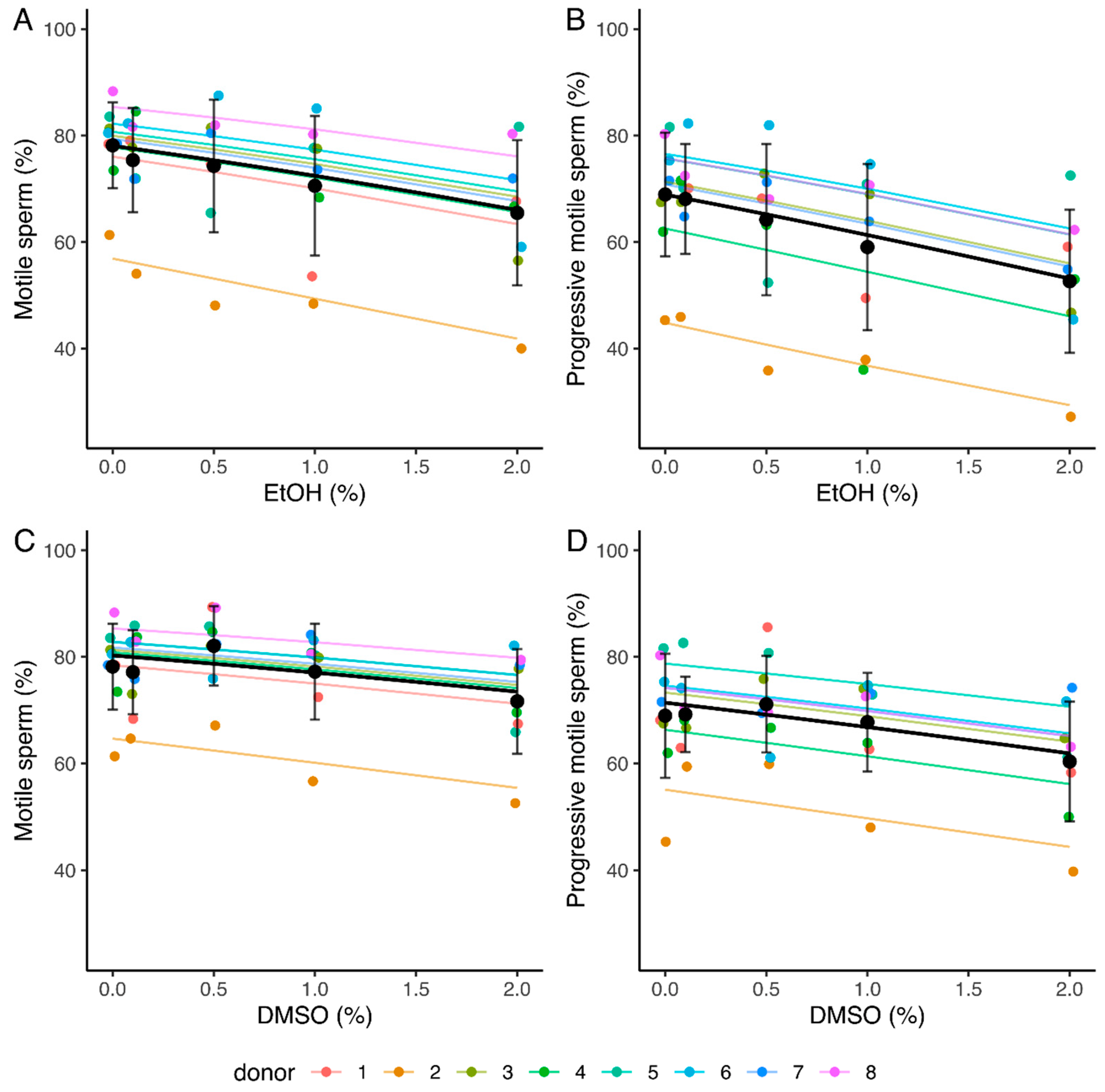
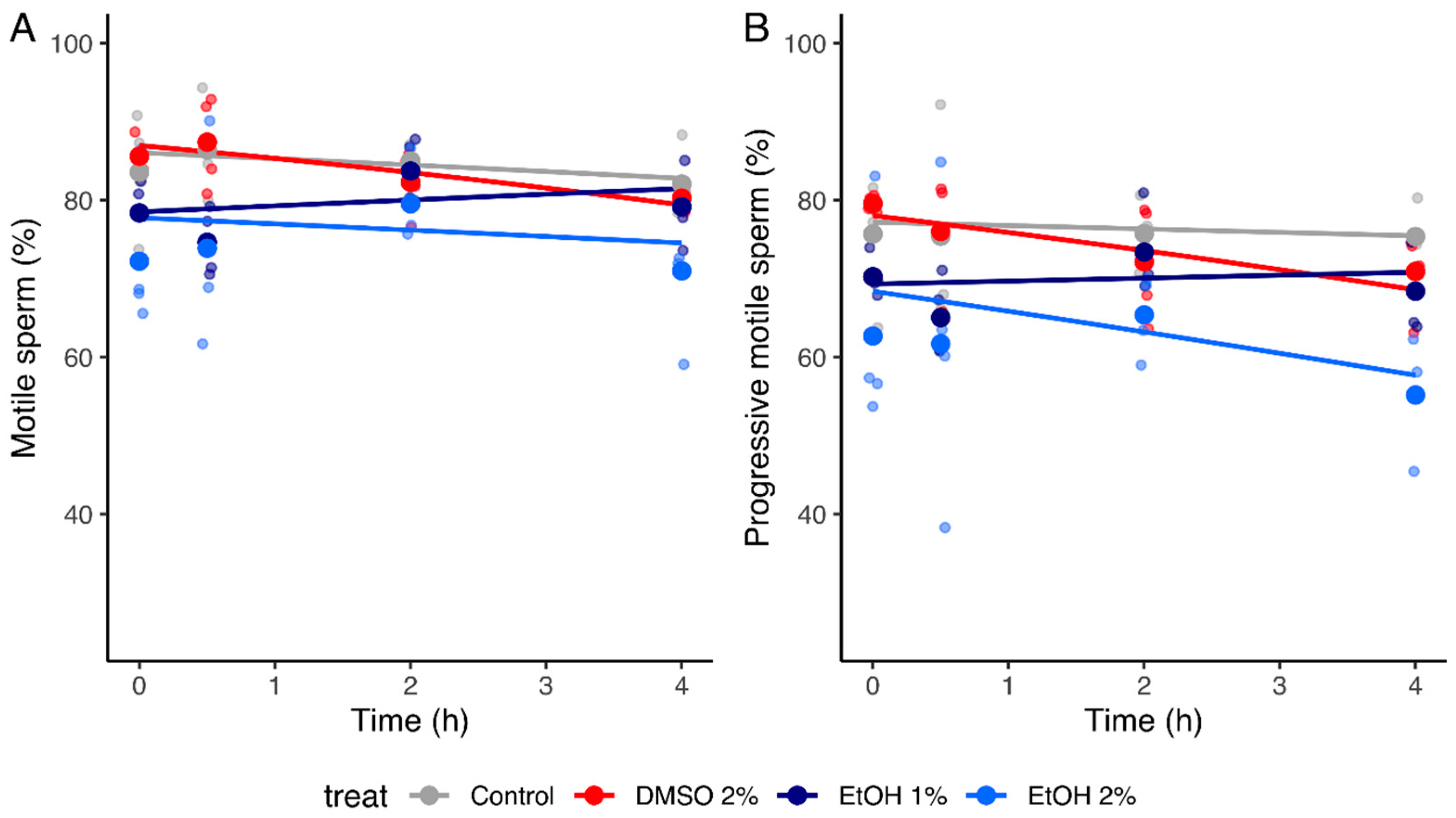
2.2.1. Sperm Motility
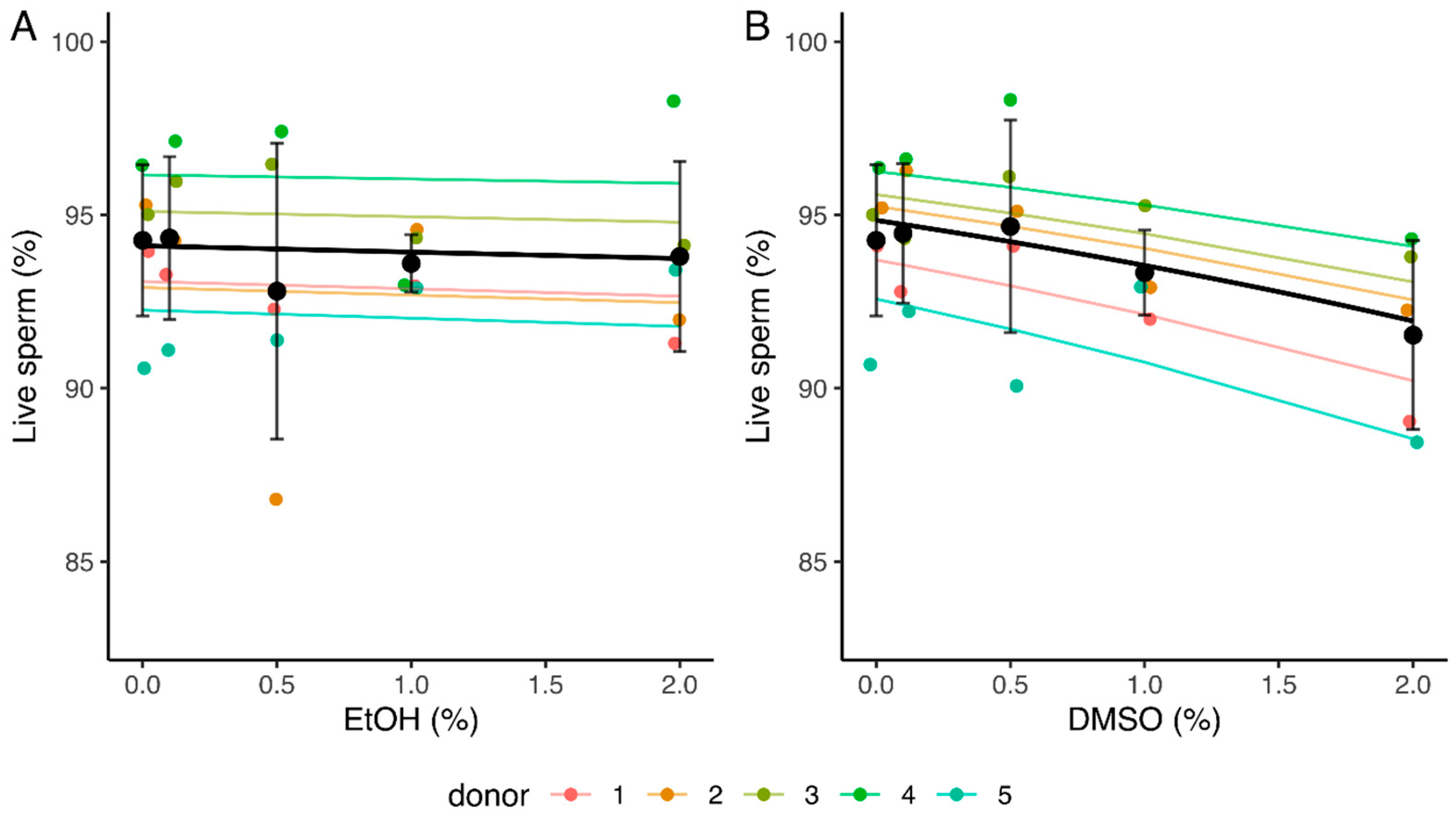
2.2.2. Sperm Vitality
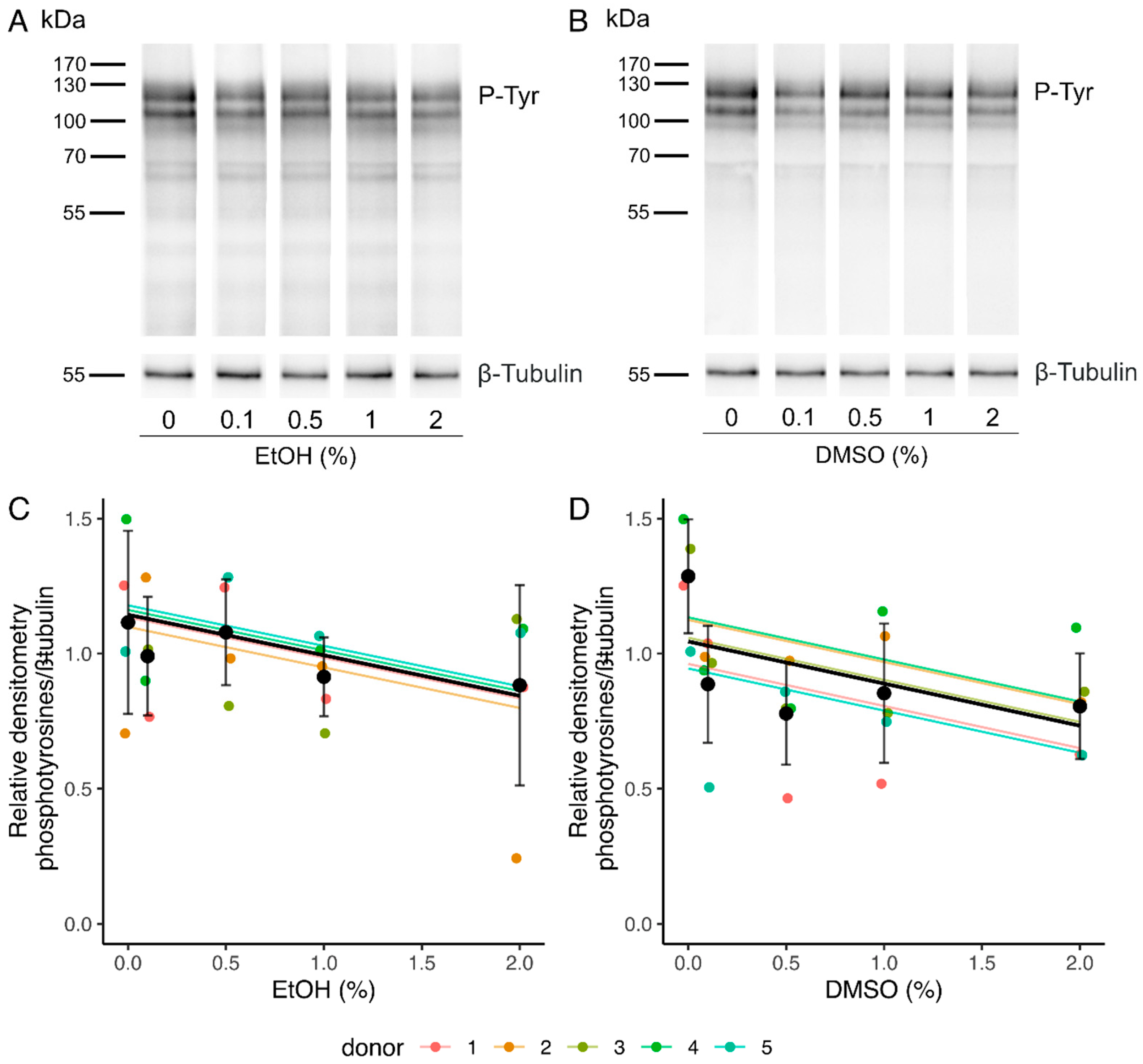
2.2.3. Sperm Capacitation
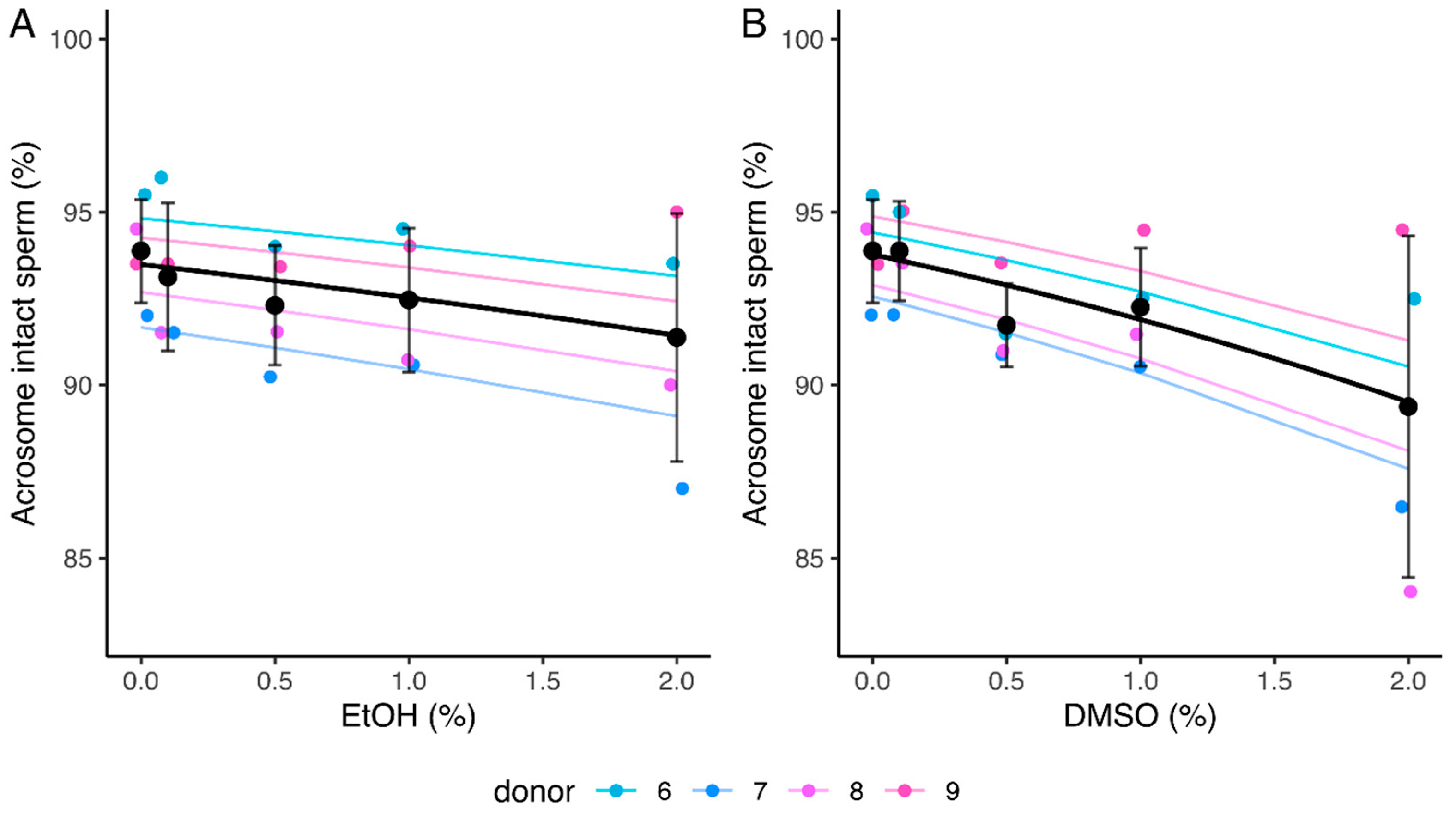
2.2.4. Acrosome Integrity
3. Discussion
4. Materials and Methods
4.1. Subjects and Ethics
4.2. Sperm Preparation
4.3. Assessment of Sperm Motility
4.4. Assessment of Sperm Vitality
4.5. Assessment of Sperm Capacitation
4.6. Evaluation of Acrosome Integrity
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Xiao, F.; Lu, S.; Song, J.; Zhang, C.; Li, J.; Gu, K.; Lan, A.; Lv, B.; Zhang, R.; et al. Research trends and perspectives of male infertility: A bibliometric analysis of 20 years of scientific literature. Andrology 2016, 4, 990–1001. [Google Scholar] [CrossRef] [PubMed]
- Kent, K.; Johnston, M.; Strump, N.; Garcia, T.X. Toward Development of the Male Pill: A Decade of Potential Non-hormonal Contraceptive Targets. Front. Cell Dev. Biol. 2020, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, J.E.; Lee, M.S.; Blithe, D.L. Update on Novel Hormonal and Nonhormonal Male Contraceptive Development. J. Clin. Endocrinol. Metab. 2021, 106, e2381–e2392. [Google Scholar] [CrossRef] [PubMed]
- Hess, K.C.; Jones, B.H.; Marquez, B.; Chen, Y.; Ord, T.S.; Kamenetsky, M.; Miyamoto, C.; Zippin, J.H.; Kopf, G.S.; Suarez, S.S.; et al. The “Soluble” Adenylyl Cyclase in Sperm Mediates Multiple Signaling Events Required for Fertilization. Dev. Cell 2005, 9, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujinoki, M. Melatonin-enhanced hyperactivation of hamster sperm. Reproduction 2008, 136, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Aquila, S.; Guido, C.; Santoro, A.; Perrotta, I.; Laezza, C.; Bifulco, M.; Sebastiano, A. Human Sperm Anatomy: Ultrastructural Localization of the Cannabinoid1 Receptor and a Potential Role of Anandamide in Sperm Survival and Acrosome Reaction. Anat. Rec. 2010, 293, 298–309. [Google Scholar] [CrossRef]
- Nixon, B.; Bielanowicz, A.; Anderson, A.L.; Walsh, A.; Hall, T.; Mccloghry, A.; Aitken, R.J. Elucidation of the signaling pathways that underpin capacitation-associated surface phosphotyrosine expression in mouse spermatozoa. J. Cell. Physiol. 2010, 224, 71–83. [Google Scholar] [CrossRef]
- Rahman, M.B.; Vandaele, L.; Rijsselaere, T.; El-Deen, M.S.; Maes, D.; Shamsuddin, M.; Van Soom, A. Bovine spermatozoa react to in vitro heat stress by activating the mitogen-activated protein kinase 14 signalling pathway. Reprod. Fertil. Dev. 2014, 26, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Roy, D.; Levi, K.; Kiss, V.; Nevo, R.; Eisenbach, M. Rhodopsin and melanopsin coexist in mammalian sperm cells and activate different signaling pathways for thermotaxis. Sci. Rep. 2020, 10, 112. [Google Scholar] [CrossRef] [Green Version]
- Giaccagli, M.M.; Gómez-Elías, M.D.; Herzfeld, J.D.; Marín-Briggiler, C.I.; Cuasnicú, P.S.; Cohen, D.J.; Da Ros, V.G. Capacitation-Induced Mitochondrial Activity Is Required for Sperm Fertilizing Ability in Mice by Modulating Hyperactivation. Front. Cell Dev. Biol. 2021, 9, 767161. [Google Scholar] [CrossRef]
- Sun, P.; Wang, Y.; Gao, T.; Li, K.; Zheng, D.; Liu, A.; Ni, Y. Hsp90 modulates human sperm capacitation via the Erk1/2 and p38 MAPK signaling pathways. Reprod. Biol. Endocrinol. 2021, 19, 39. [Google Scholar] [CrossRef] [PubMed]
- Hunnicutt, G.R.; Primakoff, P.; Myles, D.G. Sperm Surface Protein PH-20 is Bifunctional: One Activity is a Hyaluronidase and a Second, Distinct Activity is Required in Secondary Sperm-Zona Binding. Biol. Reprod. 1996, 55, 80–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montfort, L.; Frenette, G.; Sullivan, R. Sperm-zona pellucida interaction involves a carbonyl reductase activity in the hamster. Mol. Reprod. Dev. 2002, 61, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Sumigama, S.; Mansell, S.; Miller, M.; Lishko, P.V.; Cherr, G.N.; Meyers, S.A.; Tollner, T. Progesterone Accelerates the Completion of Sperm Capacitation and Activates CatSper Channel in Spermatozoa from the Rhesus Macaque1. Biol. Reprod. 2015, 93, 130. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, C.; Gao, S.-Q.; Kong, T.-T.; Chen, L.; Li, X.-F.; Song, L.; Wang, Y.-B. Effects of permethrin, cypermethrin and 3-phenoxybenzoic acid on rat sperm motility in vitro evaluated with computer-assisted sperm analysis. Toxicol. Vitr. 2010, 24, 382–386. [Google Scholar] [CrossRef] [PubMed]
- Lukáčová, A.; Kňažická, Z.; Tvrdá, E.; Greń, A.; Lukáč, N.; Massányi, P. The impact of nonylphenol (NP) on the spermatozoa motility in vitro. J. Microbiol. Biotechnol. Food Sci. 2012, 1, 1551–1560. [Google Scholar]
- Rahman, M.S.; Kwon, W.-S.; Lee, J.-S.; Yoon, S.-J.; Ryu, B.-Y.; Pang, M.-G. Bisphenol-A Affects Male Fertility via Fertility-related Proteins in Spermatozoa. Sci. Rep. 2015, 5, 9169. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.; Kwon, W.-S.; Yoon, S.-J.; Park, Y.-J.; Ryu, B.-Y.; Pang, M.-G. A novel approach to assessing bisphenol-A hazards using an in vitro model system. BMC Genom. 2016, 17, 577. [Google Scholar] [CrossRef] [Green Version]
- Barbonetti, A.; Castellini, C.; Di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In vitro exposure of human spermatozoa to bisphenol A induces pro-oxidative/apoptotic mitochondrial dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef]
- Khasin, L.G.; Della Rosa, J.; Petersen, N.; Moeller, J.; Kriegsfeld, L.J.; Lishko, P.V. The Impact of Di-2-Ethylhexyl Phthalate on Sperm Fertility. Front. Cell Dev. Biol. 2020, 8, 426. [Google Scholar] [CrossRef]
- Amjad, S.; Rahman, S.; Pang, W.-K.; Ryu, D.-Y.; Adegoke, E.O.; Park, Y.-J.; Pang, M.-G. Effects of phthalates on the functions and fertility of mouse spermatozoa. Toxicology 2021, 454, 152746. [Google Scholar] [CrossRef] [PubMed]
- Ferramosca, A.; Lorenzetti, S.; Di Giacomo, M.; Murrieri, F.; Coppola, L.; Zara, V. Herbicides glyphosate and glufosinate ammonium negatively affect human sperm mitochondria respiration efficiency. Reprod. Toxicol. 2021, 99, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.; Ball, B.A. Effect of α-tocopherol and tocopherol succinate on lipid peroxidation in equine spermatozoa. Anim. Reprod. Sci. 2005, 87, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Giacone, F.; Condorelli, R.A.; Mongioì, L.M.; Bullara, V.; La Vignera, S.; Calogero, A.E. In vitro effects of zinc, D-aspartic acid, and coenzyme-Q10 on sperm function. Endocrine 2017, 56, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Llavanera, M.; Delgado-Bermúdez, A.; Olives, S.; Mateo-Otero, Y.; Recuero, S.; Bonet, S.; Fernández-Fuertes, B.; Yeste, M.; Barranco, I. Glutathione S-Transferases Play a Crucial Role in Mitochondrial Function, Plasma Membrane Stability and Oxidative Regulation of Mammalian Sperm. Antioxidants 2020, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Knuth, U.A.; Yeung, C.-H.; Nieschlag, E. Combination of 19-nortestosterone-hexyloxyphenylpropionate (Anadur) and depot-medroxyprogesterone-acetate (Clinovir) for male contraception. Fertil. Steril. 1989, 51, 1011–1018. [Google Scholar] [CrossRef]
- Chaudhury, K.; Bhattacharyya, A.; Guha, S. Studies on the membrane integrity of human sperm treated with a new injectable male contraceptive. Hum. Reprod. 2004, 19, 1826–1830. [Google Scholar] [CrossRef] [Green Version]
- Naz, R.K. Can curcumin provide an ideal contraceptive? Mol. Reprod. Dev. 2011, 78, 116–123. [Google Scholar] [CrossRef]
- Gruber, F.S.; Johnston, Z.C.; Barratt, C.L.; Andrews, P.D. A phenotypic screening platform utilising human spermatozoa identifies compounds with contraceptive activity. eLife 2020, 9, e51739. [Google Scholar] [CrossRef]
- Cummins, J.M.; Pember, S.M.; Jequier, A.M.; Yovich, J.; Hartmann, P.E. A test of the human sperm acrosome reaction following ionophore challenge. Relationship to fertility and other seminal parameters. J. Androl. 1991, 12, 98–103. [Google Scholar]
- Cafe, S.L.; Anderson, A.L.; Nixon, B. In vitro Induction and Detection of Acrosomal Exocytosis in Human Spermatozoa. BIO-PROTOCOL 2020, 10, e3689. [Google Scholar] [CrossRef]
- Srivastava, N.; Pande, M. Determination of Mitochondrial Function in Sperm Cells. In Protocols in Semen Biology (Comparing Assays); Srivastava, N., Pande, M., Eds.; Springer: Singapore, 2017. [Google Scholar] [CrossRef]
- Mocé, E.; Vicente, J.S. Rabbit sperm cryopreservation: A review. Anim. Reprod. Sci. 2009, 110, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sieme, H.; Oldenhof, H.; Wolkers, W.F. Mode of action of cryoprotectants for sperm preservation. Anim. Reprod. Sci. 2016, 169, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Öztürk, A.E.; Bucak, M.N.; Bodu, M.; Başpınar, N.; Çelik, İ.; Shu, Z.; Keskin, N.; Gao, D. Cryobiology and Cryopreservation of Sperm. In Cryopreservation—Current Advances and Evaluations; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Timm, M.; Saaby, L.; Moesby, L.; Hansen, E.W. Considerations regarding use of solvents in in vitro cell based assays. Cytotechnology 2013, 65, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, R.Y.-K.; Lee, K.K.-H.; Wong, W.-Y.; Tang, J.C.-O.; Lam, K.-H.; Cheng, C.-H.; Lau, F.-Y.; Kok, S.H.-L.; Gambari, R.; Chui, C.-H. The Puzzling Issue of ‘Vehicle-Treated Control’ when Using Ethanol as Drug Carrier for MCF-7 Cells. Phytotherapy Res. 2014, 28, 1735–1736. [Google Scholar] [CrossRef] [PubMed]
- Galvao, J.; Davis, B.; Tilley, M.; Normando, E.; Duchen, M.R.; Cordeiro, M.F. Unexpected low-dose toxicity of the universal solvent DMSO. FASEB J. 2014, 28, 1317–1330. [Google Scholar] [CrossRef]
- Yuan, C.; Gao, J.; Guo, J.; Bai, L.; Marshall, C.; Cai, Z.; Wang, L.; Xiao, M. Dimethyl Sulfoxide Damages Mitochondrial Integrity and Membrane Potential in Cultured Astrocytes. PLoS ONE 2014, 9, e107447. [Google Scholar] [CrossRef] [Green Version]
- Jamalzadeh, L.; Ghafoori, H.; Sariri, R.; Rabuti, H.; Nasirzade, J.; Hasani, H.; Aghamaali, M.R. Cytotoxic Effects of Some Common Organic Solvents on MCF-7, RAW-264.7 and Human Umbilical Vein Endothelial Cells. J. Med. Biochem. 2016, 4, e33453. [Google Scholar] [CrossRef] [Green Version]
- Verheijen, M.; Lienhard, M.; Schrooders, Y.; Clayton, O.; Nudischer, R.; Boerno, S.; Timmermann, B.; Selevsek, N.; Schlapbach, R.; Gmuender, H.; et al. DMSO induces drastic changes in human cellular processes and epigenetic landscape in vitro. Sci. Rep. 2019, 9, 4641. [Google Scholar] [CrossRef] [Green Version]
- Sherman, J.K. Dimethyl Sulfoxide As a Protective Agent during Freezing and Thawing of Human Spermatozoa. Exp. Biol. Med. 1964, 117, 261–264. [Google Scholar] [CrossRef]
- Serafini, P.C.; Hauser, D.; Moyer, D.; Marrs, R.P. Cryopreservation of human spermatozoa: Correlations of ultrastructural sperm head configuration with sperm motility and ability to penetrate zona-free hamster ova. Fertil. Steril. 1986, 46, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Si, W.; Zheng, P.; Li, Y.; Dinnyes, A.; Ji, W. Effect of glycerol and dimethyl sulfoxide on cryopreservation of rhesus monkey (Macaca mulatta) sperm. Am. J. Primatol. 2004, 62, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Farshad, A.; Khalili, B.; Fazeli, P. The Effect of Different Concentrations of Glycerol and DMSO on Viability of Markhoz Goat Spermatozoa during Different Freezing Temperatures Steps. Pak. J. Biol. Sci. 2009, 12, 239–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Lee, Y.-J.; Ji, D.-B.; Kim, Y.-J. Evaluation of Different Cryoprotectants (CPAs) in Boar Semen Cryopreservation. J. Vet. Med. Sci. 2011, 73, 961–963. [Google Scholar] [CrossRef] [Green Version]
- Najafi, A.; Daghigh-Kia, H.; Dodaran, H.V.; Mehdipour, M.; Alvarez-Rodriguez, M. Ethylene glycol, but not DMSO, could replace glycerol inclusion in soybean lecithin-based extenders in ram sperm cryopreservation. Anim. Reprod. Sci. 2017, 177, 35–41. [Google Scholar] [CrossRef]
- Oldenhof, H.; Bigalk, J.; Hettel, C.; Barros, L.D.O.; Sydykov, B.; Bajcsy, C.; Sieme, H.; Wolkers, W.F. Stallion Sperm Cryopreservation Using Various Permeating Agents: Interplay between Concentration and Cooling Rate. Biopreservation Biobanking 2017, 15, 422–431. [Google Scholar] [CrossRef]
- Karakus, F.N.; Kuran, S.B.; Solakoglu, S. Effect of curcumin on sperm parameters after the cryopreservation. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 267, 161–166. [Google Scholar] [CrossRef]
- Lewin, A.; Lavon, H. The effect of coenzyme Q10 on sperm motility and function. Mol. Asp. Med. 1997, 18, 213–219. [Google Scholar] [CrossRef]
- Naz, R.K. The Effect of Curcumin on Intracellular pH (pHi), Membrane Hyperpolarization and Sperm Motility. J. Reprod. Infertil. 2014, 15, 62–70. [Google Scholar]
- Salonen, I. Exposure to ethanol during capacitation impairs the fertilizing ability of human spermatozoa in vitro. Int. J. Androl. 1986, 9, 259–270. [Google Scholar] [CrossRef]
- Rogers, B.J.; Cash, M.K.M.; Vaughn, W.K. Ethanol inhibits human and hamster sperm penetration of eggs. Gamete Res. 1987, 16, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A.; Reddy, J.M.; Joyce, C.; Willis, B.R.; Van der Ven, H.; Zaneveld, L.J.D. Inhibition of Mouse Sperm Capacitation by Ethanol. Biol. Reprod. 1982, 27, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Chian, R.-C.; Lapointe, S. Capacitation in vitro of bovine spermatozoa by oviduct epithelial cell monolayer conditioned medium. Mol. Reprod. Dev. 1995, 42, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.G.; Lee, M.A.; Iozzo, R.V.; Lopez, I.; Touchstone, J.C.; Storey, B.T. Ethanol Accelerates Acrosomal Loss in Human Spermatozoa. J. Androl. 1988, 9, 357–366. [Google Scholar] [CrossRef]
- Donnelly, G.P.; McClure, N.; Kennedy, M.S.; Lewis, S.E. Direct effect of alcohol on the motility and morphology of human spermatozoa. Andrologia 1999, 31, 43–47. [Google Scholar] [CrossRef]
- Si, Y.; Olds-Clarke, P. Evidence for the Involvement of Calmodulin in Mouse Sperm Capacitation. Biol. Reprod. 2000, 62, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Aitken, R.J.; Buckingham, D.W.; Harkiss, D.; Paterson, M.; Fisher, H.; Irvine, D. The extragenomic action of progesterone on human spermatozoa is influenced by redox regulated changes in tyrosine phosphorylation during capacitation. Mol. Cell. Endocrinol. 1996, 117, 83–93. [Google Scholar] [CrossRef]
- Naz, R.K.; Rajesh, P.B. Role of tyrosine phosphorylation in sperm capacitation / acrosome reaction. Reprod. Biol. Endocrinol. 2004, 2, 75. [Google Scholar] [CrossRef] [Green Version]
- Uguz, C.; Varisli, O.; Agca, C.; Agca, Y. Effects of nonylphenol on motility and subcellular elements of epididymal rat sperm. Reprod. Toxicol. 2009, 28, 542–549. [Google Scholar] [CrossRef]
- Uribe, P.; Boguen, R.; Treulen, F.; Sánchez, R.; Villegas, J. Peroxynitrite-mediated nitrosative stress decreases motility and mitochondrial membrane potential in human spermatozoa. Mol. Hum. Reprod. 2015, 21, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Wen, Z.; Lei, Z.; Tian, E.; Wang, Y.; Zhong, Y.; Ge, R.-S. Inhibition of human sperm motility and capacitation by ziram is mediated by decreasing tyrosine protein kinase. Ecotoxicol. Environ. Saf. 2021, 218, 112281. [Google Scholar] [CrossRef]
- Treulen, F.; Uribe, P.; Boguen, R.; Villegas, J. Mitochondrial outer membrane permeabilization increases reactive oxygen species production and decreases mean sperm velocity but is not associated with DNA fragmentation in human sperm. Mol. Hum. Reprod. 2016, 22, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naz, R.K. Effect of Actinomycin D and Cycloheximide on Human Sperm Function. Arch. Androl. 1998, 41, 135–142. [Google Scholar] [CrossRef]
- Bureau, M.; Bailey, J.L.; Sirard, M.A. Binding regulation of porcine spermatozoa to oviductal vesicles in vitro. J. Androl. 2002, 23, 188–193. [Google Scholar] [PubMed]
- Carretera, M.; Fumuso, F.; Neild, D.; Giuliano, S.; Cetica, P.; Miragaya, M. Evaluation of the acrosomal status in Lama glama sperm incubated with acrosome reaction inducers. Anim. Reprod. Sci. 2015, 160, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Brenker, C.; Goodwin, N.; Weyand, I.; Kashikar, N.D.; Naruse, M.; Krähling, M.; Müller, A.; Kaupp, U.B.; Strünker, T. The CatSper channel: A polymodal chemosensor in human sperm. EMBO J. 2012, 31, 1654–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.-H.; Zhu, Y.-Y.; Wang, L.; Liu, H.-L.; Ling, Y.; Li, Z.-L.; Sun, L.-B. The Catsper channel and its roles in male fertility: A systematic review. Reprod. Biol. Endocrinol. 2017, 15, 65. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Young, S.; Krenz, H.; Tüttelmann, F.; Röpke, A.; Krallmann, C.; Kliesch, S.; Zeng, X.-H.; Brenker, C.; Strünker, T. The Ca2+ channel CatSper is not activated by cAMP/PKA signaling but directly affected by chemicals used to probe the action of cAMP and PKA. J. Biol. Chem. 2020, 295, 13181–13193. [Google Scholar] [CrossRef]
- Ren, D.; Navarro, B.; Perez, G.; Jackson, A.C.; Hsu, S.; Shi, Q.; Tilly, J.L.; Clapham, D.E. A sperm ion channel required for sperm motility and male fertility. Nature 2001, 413, 603–609. [Google Scholar] [CrossRef]
- Tamburrino, L.; Marchiani, S.; Minetti, F.; Forti, G.; Muratori, M.; Baldi, E. The CatSper calcium channel in human sperm: Relation with motility and involvement in progesterone-induced acrosome reaction. Hum. Reprod. 2014, 29, 418–428. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, P.; Durairajanayagam, D.; Agarwal, A. Fuel/Energy Sources of Spermatozoa. In Male Infertility; Parekattil, S., Esteves, S., Agarwal, A., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Nicholson, C.; Abramsson, L.; Holm, S.; Bjurulf, E. Bacterial contamination and sperm recovery after semen preparation by density gradient centrifugation using silane-coated silica particles at different g forces. Hum. Reprod. 2000, 15, 662–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jonge, C. Biological basis for human capacitation. Hum. Reprod. Updat. 2005, 11, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sati, L.; Cayli, S.; Delpiano, E.; Sakkas, D.; Huszar, G. The Pattern of Tyrosine Phosphorylation in Human Sperm in Response to Binding to Zona Pellucida or Hyaluronic Acid. Reprod. Sci. 2014, 21, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sáez-Espinosa, P.; Huerta-Retamal, N.; Robles-Gómez, L.; Avilés, M.; Aizpurua, J.; Velasco, I.; Romero, A.; Gómez-Torres, M.J. Influence of in vitro capacitation time on structural and functional human sperm parameters. Asian J. Androl. 2020, 22, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Alquézar-Baeta, C.; Gimeno-Martos, S.; Jiménez, S.M.; Santolaria, P.; Yániz, J.; Palacín, I.; Casao, A.; Cebrián-Pérez, J.; Muiño-Blanco, T.; Pérez-Pé, R. OpenCASA: A new open-source and scalable tool for sperm quality analysis. PLoS Comput. Biol. 2019, 15, e1006691. [Google Scholar] [CrossRef]
- Elia, J.; Imbrogno, N.; Delfino, M.; Mazzilli, R.; Rossi, T.; Mazzilli, F. The importance of the sperm motility classes—Future directions. Open Androl. J. 2010, 2, 42–43. [Google Scholar]
- Bisconti, M.; Leroy, B.; Gallagher, M.T.; Senet, C.; Martinet, B.; Arcolia, V.; Wattiez, R.; Kirkman-Brown, J.C.; Simon, J.-F.; Hennebert, E. The ribosome inhibitor chloramphenicol induces motility deficits in human spermatozoa: A proteomic approach identifies potentially involved proteins. Front. Cell Dev. Biol. 2022, 10, 965076. [Google Scholar] [CrossRef]
- Larson, J.L.; Miller, D. Simple histochemical stain for acrosomes on sperm from several species. Mol. Reprod. Dev. 1999, 52, 445–449. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Age (years) | Volume (mL) | Viscosity | pH | Sperm Count (×106 Sperm/mL) | Total Motility (%) | Progressive Motility (%) | Normal Morphology (%) | Experiment |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 30 | 2.8 | normal | 8.1 | 38.5 | 79.3 | 63.2 | 4 | C, M, V |
| 2 | 31 | 2.9 | normal | 7.9 | 114.07 | 56.1 | 30 | 5.79 | C, M, V |
| 3 | 29 | 6 | normal | 8.5 | 110.3 | 80.3 | 53.8 | <1 | C, M, V |
| 4 | 21 | 6 | increased | 8.1 | 62.93 | 85.14 | 54.16 | 4 | C, M, V |
| 5 | 30 | 3 | normal | 8.1 | 51.5 | 70.9 | 45.7 | 4.1 | C, M, V |
| 6 | 25 | 2 | increased | 8.3 | 146.9 | 91.8 | 69.5 | 0.8 | AI, M, MT |
| 7 | 37 | 3 | increased | 8.1 | 121.2 | 88.7 | 67.7 | 0.81 | AI, M, MT |
| 8 | 30 | 3.9 | increased | 8.1 | 104.1 | 79.14 | 58.32 | 7.5 | AI, M, MT |
| 9 | 34 | 1.9 | normal | 7.9 | 80.78 | 74.5 | 59.9 | 1.7 | AI, M, MT |
| Parameter | EtOH | DMSO | ||
|---|---|---|---|---|
| Coefficient (95% CI) | n | Coefficient (95% CI) | n | |
| Motile sperm (%) | −0.30 [−0.39, −0.22] * | 8 | −0.19 [−0.28, −0.10] * | 8 |
| Progressively motile sperm (%) | −0.34 [−0.41, −0.26] * | 8 | −0.22 [−0.30, −0.13] * | 8 |
| Live sperm (%) | −0.032 [−0.16, 0.10] | 5 | −0.24 [−0.37, −0.11] * | 5 |
| Relative phosphotyrosine content | −0.15 [−0.28, −0.0048] * | 5 | −0.16 [−0.29, −0.021] * | 5 |
| Acrosome-intact sperm (%) | −0.15 [−0.32, 0.022] | 4 | −0.28 [−0.43, −0.12] * | 4 |
| Parameter | Control | 1% EtOH | 2% EtOH | 2% DMSO |
|---|---|---|---|---|
| Time (95% CI) | Difference of Time (95% CI) | Difference of Time (95% CI) | Difference of Time (95% CI) | |
| Motile sperm (%) | −0.062 [−0.15, 0.023] | 0.11 [−0.0072, 0.23] | 0.018 [−0.099, 0.14] | −0.075 [−0.19, 0.037] |
| Parameter | EtOH (%) | DMSO (%) |
|---|---|---|
| Motile sperm (%) | 0.9 | 1.5 |
| Progressively motile sperm (%) | 0.7 | 1.1 |
| Live sperm (%) | † | >2 * |
| Relative phosphotyrosine content | 0.3 | 0.3 |
| Acrosome-intact sperm (%) | † | >2 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bisconti, M.; Grosjean, P.; Arcolia, V.; Simon, J.-F.; Hennebert, E. Influence of Two Widely Used Solvents, Ethanol and Dimethyl Sulfoxide, on Human Sperm Parameters. Int. J. Mol. Sci. 2023, 24, 505. https://doi.org/10.3390/ijms24010505
Bisconti M, Grosjean P, Arcolia V, Simon J-F, Hennebert E. Influence of Two Widely Used Solvents, Ethanol and Dimethyl Sulfoxide, on Human Sperm Parameters. International Journal of Molecular Sciences. 2023; 24(1):505. https://doi.org/10.3390/ijms24010505
Chicago/Turabian StyleBisconti, Marie, Philippe Grosjean, Vanessa Arcolia, Jean-François Simon, and Elise Hennebert. 2023. "Influence of Two Widely Used Solvents, Ethanol and Dimethyl Sulfoxide, on Human Sperm Parameters" International Journal of Molecular Sciences 24, no. 1: 505. https://doi.org/10.3390/ijms24010505