
Tomato Sterol 22-desaturase Gene CYP710A11: Its Roles in Meloidogyne incognita Infection and Plant Stigmasterol Alteration




Abstract
:1. Introduction
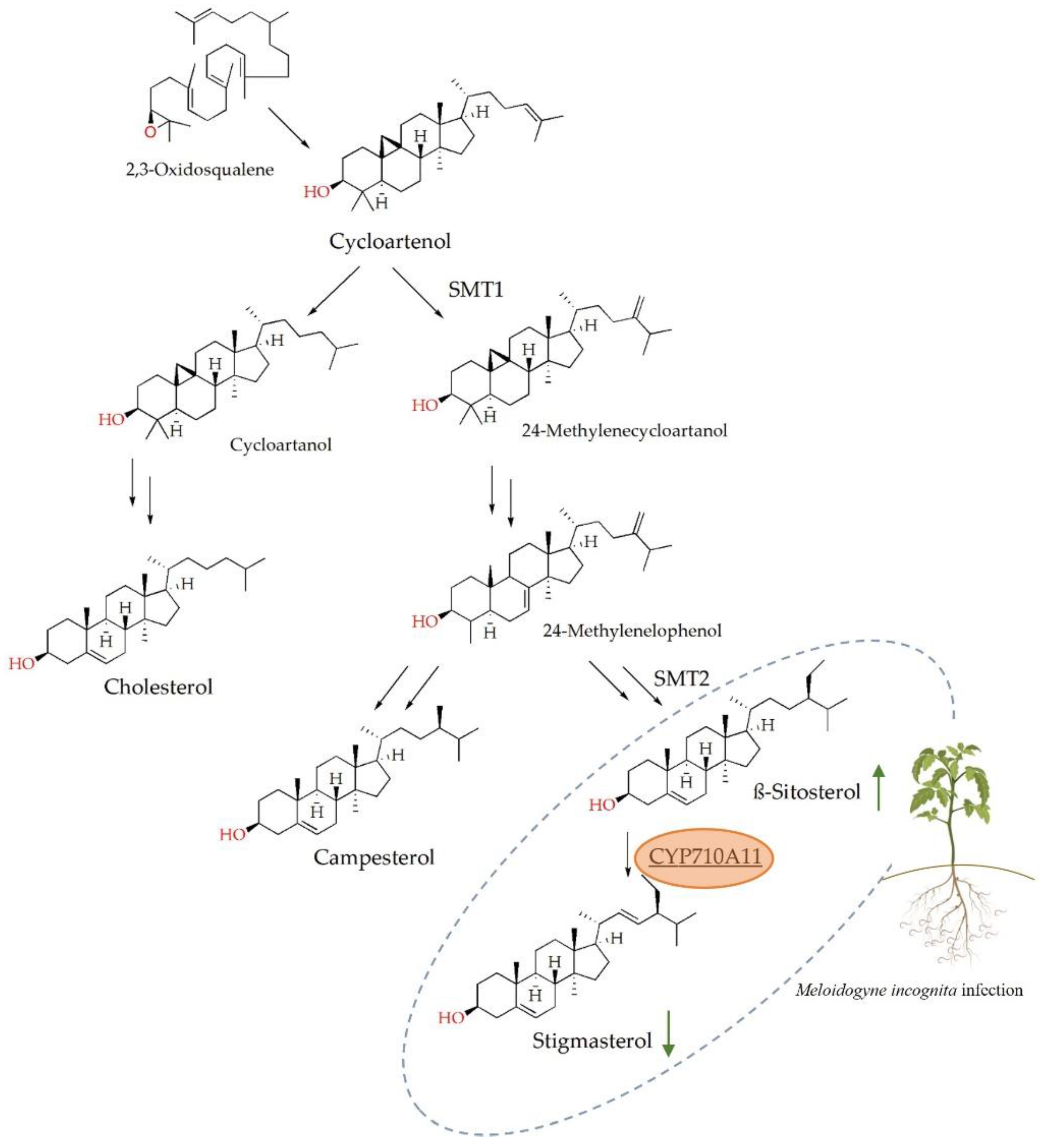
2. Results
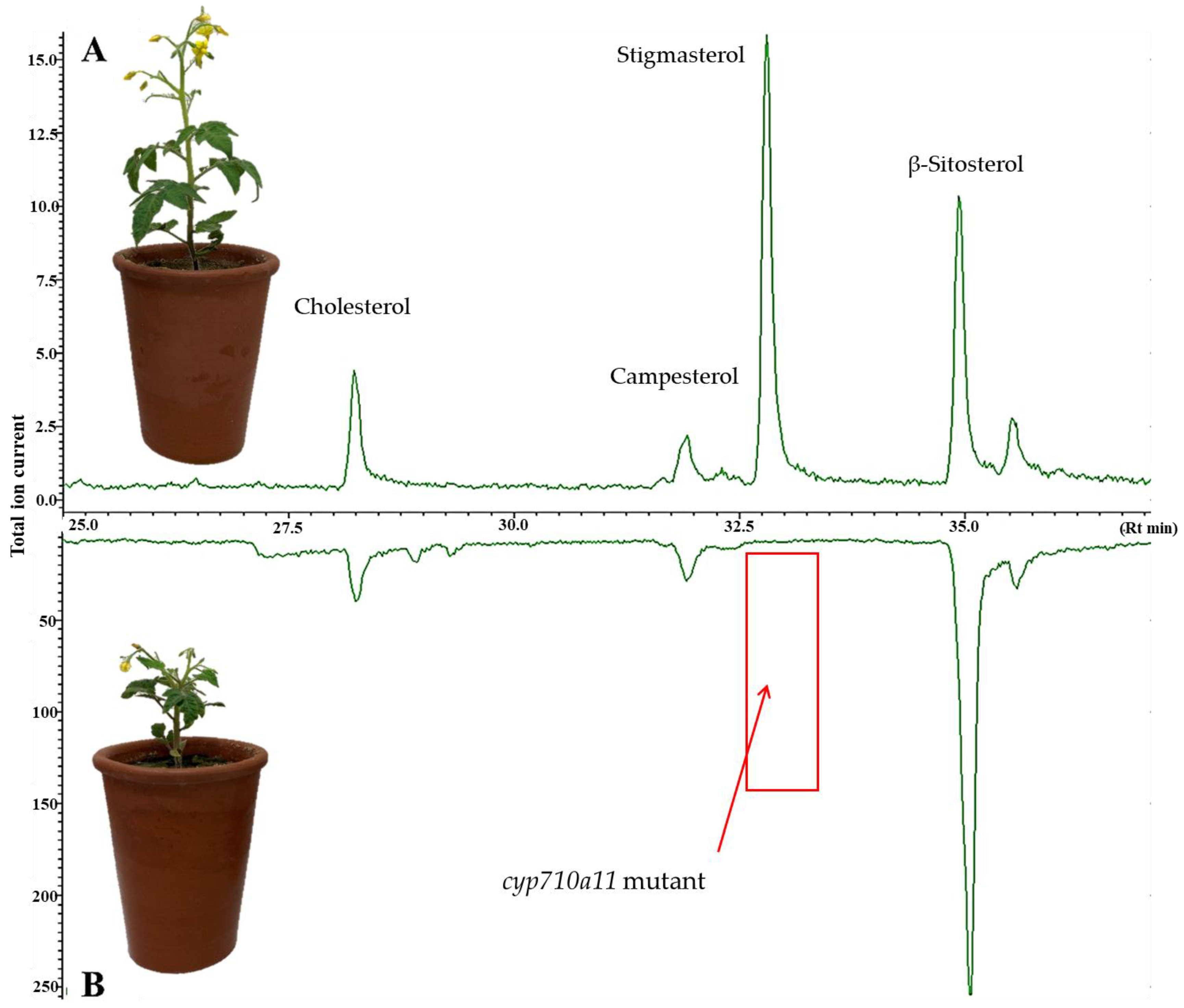
2.1. Characterization of the Micro-Tom cyp710a11 Mutant Line
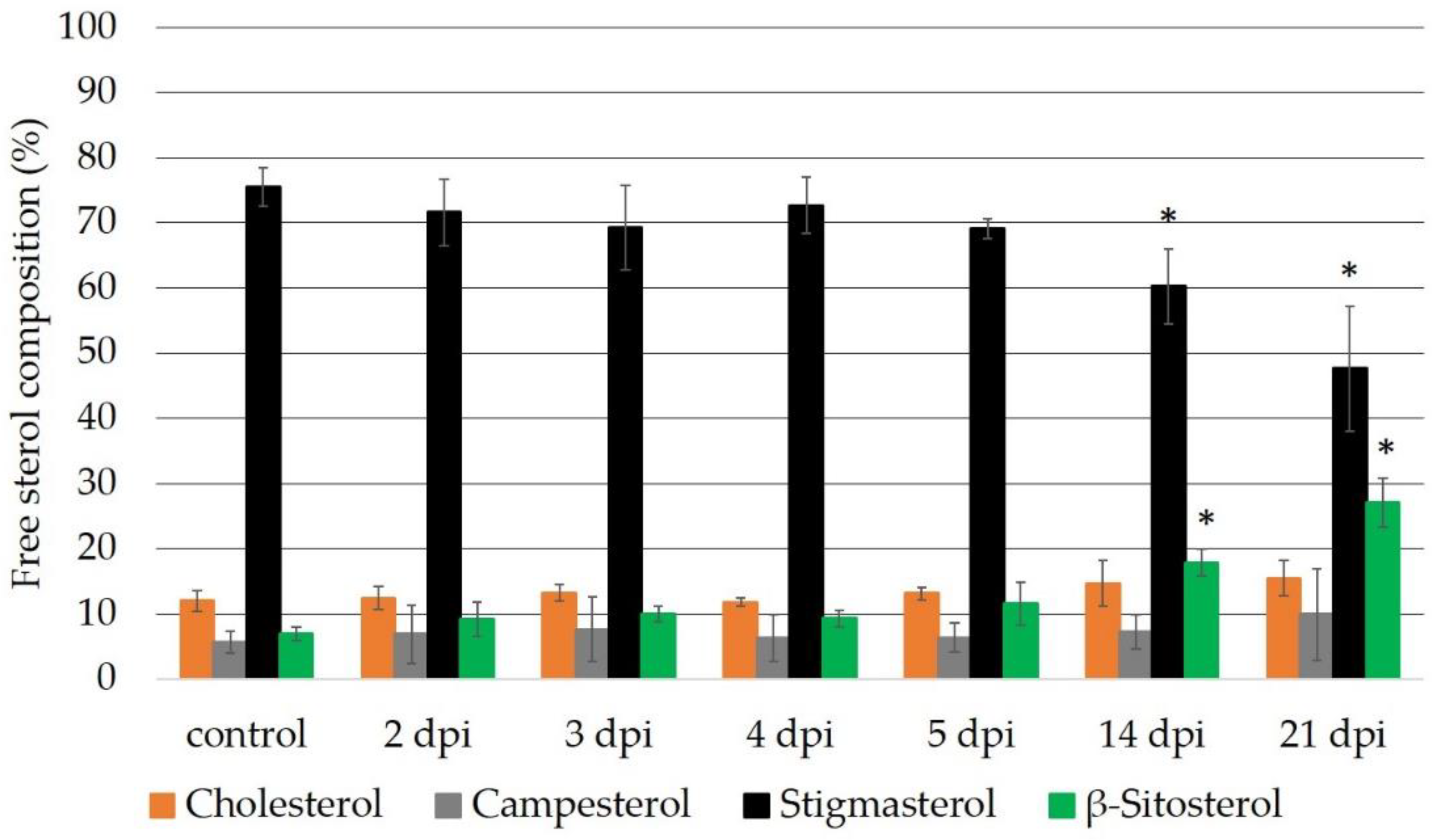
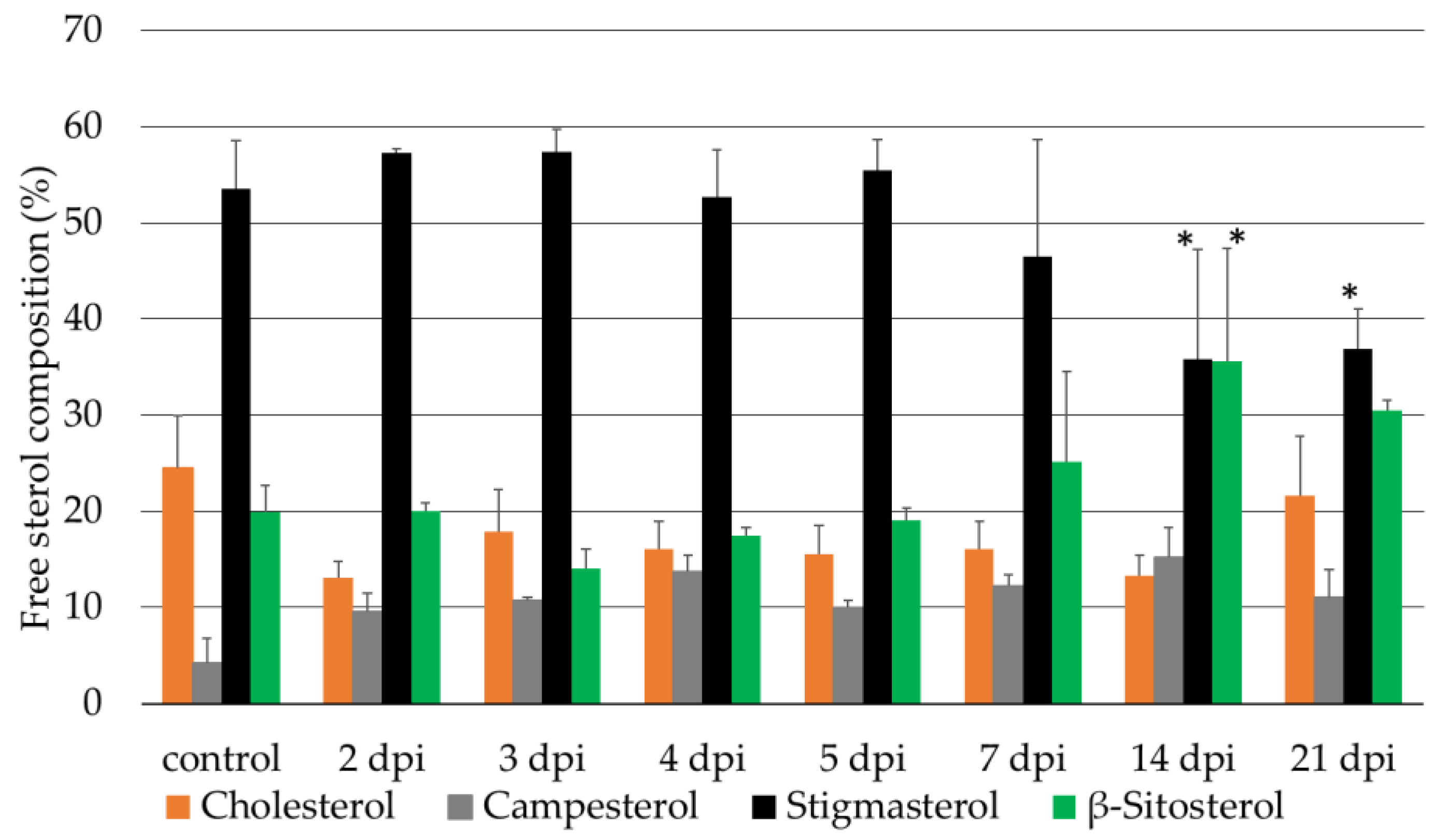
2.2. Sterol Composition of the Micro-Tom wt and cyp710a11 Mutant Plants after M. incognita Infection
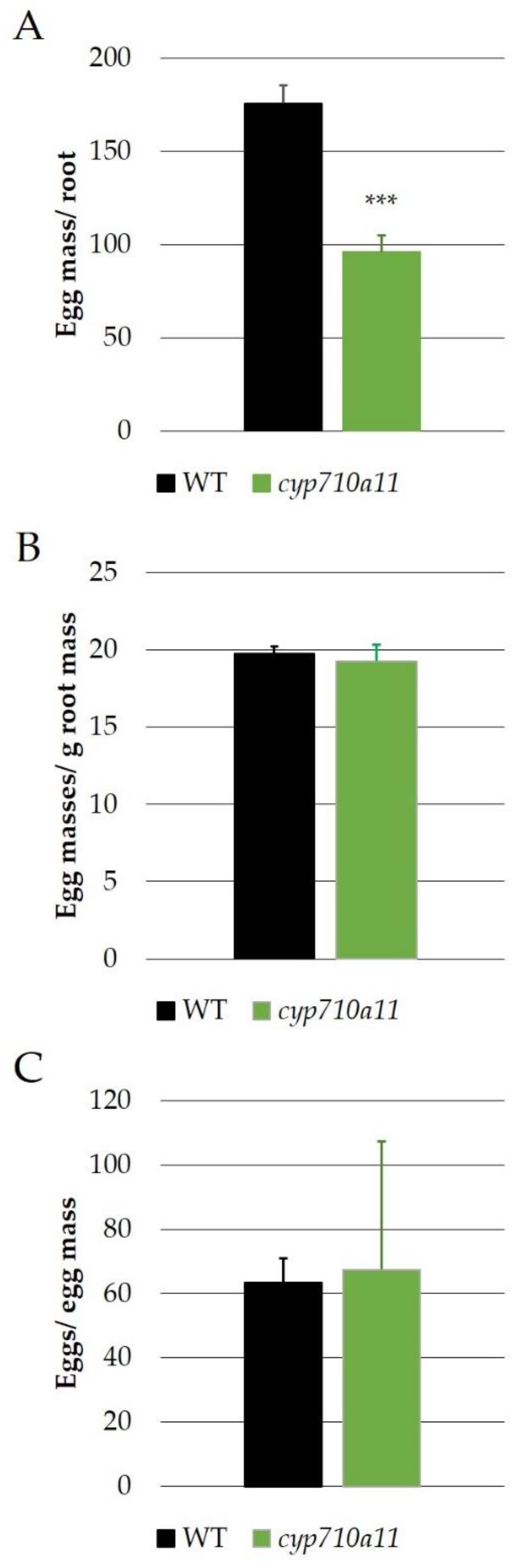
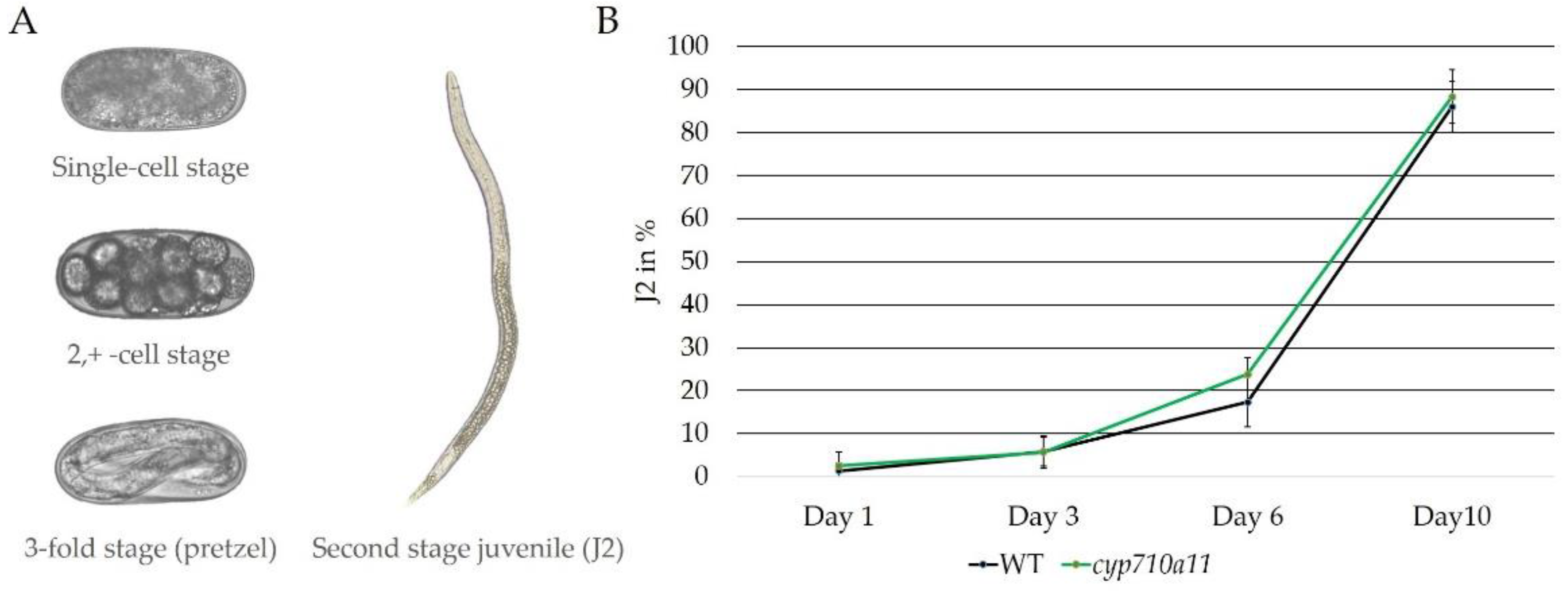
2.3. Effect of M. incognita on Micro-Tom cyp710a11 Mutant Plants
3. Discussion
4. Materials and Methods
4.1. Nematode Inoculum
4.2. Tomato cv. Micro-Tom cyp710a11 Mutant Line
4.3. CYP710A11 Gene Expression Analysis
4.4. Sterol Extraction and Gas Chromatography–Mass Spectrometry Analysis
4.5. M. incognita Growth on the Tomato cv. Micro-Tom cyp710a11 Mutant Line
4.6. Comparison of the Root Development of the Tomato cv. Micro-Tom wt and cyp710a11 Mutant Lines Infected with M. incognita
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dufourc, E.J. Sterols and membrane dynamics. J. Chem. Biol. 2008, 1, 63–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skubic, C.; Rozman, D. Sterols from the post-lanosterol part of cholesterol synthesis: Novel signaling players. In Mammalian Sterols; Rozman, D., Gebhardt, R., Eds.; Springer: Cham, Switzerland, 2020; pp. 1–22. [Google Scholar]
- Summons, R.E.; Bradley, A.S.; Jahnke, L.L.; Waldbauer, J.R. Steroids, triterpenoids and molecular oxygen. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006, 361, 951–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, N.K.; Coates, H.W.; Brown, A.J. Squalene monooxygenase: A journey to the heart of cholesterol synthesis. Prog. Lipid Res. 2020, 79, 101033. [Google Scholar] [CrossRef] [PubMed]
- Desmond, E.; Gribaldo, S. Phylogenomics of sterol synthesis: Insights into the origin, evolution, and diversity of a key eukaryotic feature. Genome Biol. Evol. 2009, 1, 364–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.; Behmer, S.T. Insect sterol nutrition: Physiological mechanisms, ecology, and applications. Annu. Rev. Entomol. 2020, 65, 251–271. [Google Scholar] [CrossRef] [PubMed]
- Shamsuzzama; Lebedev, R.; Trabelcy, B.; Goncalves, I.L.; Gerchman, Y.; Sapir, A. Metabolic reconfiguration in C. elegans suggests a pathway for widespread sterol auxotrophy in the animal kingdom. Curr. Biol. 2020, 30, 3031–3038. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.; Govers, F. The mysterious route of sterols in oomycetes. PLoS Pathog. 2021, 17, e1009591. [Google Scholar] [CrossRef]
- Benveniste, P. Biosynthesis and accumulation of sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T.; Grebenok, R.J.; Douglas, A.E. Plant sterols and host plant suitability for a phloem-feeding insect. Funct. Ecol. 2011, 25, 484–491. [Google Scholar] [CrossRef]
- Hedin, P.A.; Callahan, F.E.; Dollar, D.A.; Greech, R.G. Total sterols in root-knot nematode Meloidogyne incognita infected cotton Gossypium hirsutum (L.) plant roots. Comp. Biochem. Physiol. 1995, 111, 447–452. [Google Scholar] [CrossRef]
- Cabianca, A.; Müller, L.; Pawlowski, K.; Dahlin, P. Changes in the plant β-Sitosterol/Stigmasterol ratio caused by the plant parasitic nematode Meloidogyne incognita. Plants 2021, 10, 292. [Google Scholar] [CrossRef]
- Morikawa, T.; Mizutani, M.; Aoki, N.; Watanabe, B.; Saga, H.; Saito, S.; Oikawa, A.; Suzuki, H.; Sakurai, N.; Shibata, D.; et al. Cytochrome P450 CYP710A encodes the sterol C-22 desaturase in Arabidopsis and tomato. Plant Cell 2006, 18, 1008–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Y.; Fu, X.; Chu, Y.; Wu, P.; Liu, Y.; Ma, L.; Tian, H.; Zhu, B. Biosynthesis and the Roles of Plant Sterols in Development and Stress Responses. Int. J. Mol. Sci. 2022, 23, 2332. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-Lòpez, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozbuga, R.; Lilley, C.J.; Knox, J.P.; Urwin, P.E. Host-specific signatures of the cell wall changes induced by the plant parasitic nematode, Meloidogyne incognita. Sci. Rep. 2018, 8, 17302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meidani, C.; Ntalli, N.G.; Giannoutsou, E.; Adamakis, I.D.S. Cell wall modifications in giant cells induced by the plant parasitic nematode Meloidogyne incognita in wild-type (col-0) and the fra2 Arabidopsis thaliana katanin mutant. Int. J. Mol. Sci. 2019, 20, 5465. [Google Scholar] [CrossRef] [Green Version]
- Zeck, W.M. Rating scheme for field evaluation of root-knot nematode infestations. Pflanzenschutz Nachr. 1971, 24, 141–144. [Google Scholar]
- Seethepalli, A.; York, L.M. RhizoVision Explorer—Interactive software for generalized root image analysis designed for everyone (Version 2.0.3). Zenodo 2020. [Google Scholar] [CrossRef]
- Aboobucker, S.I.; Suza, W.P. Why do plants convert sitosterol to stigmasterol? Front. Plant Sci. 2019, 10, 354. [Google Scholar] [CrossRef] [Green Version]
- Tang, J.; Ohyama, K.; Kawaura, K.; Hashinokuchi, H.; Kamiya, Y.; Suzuki, M.; Muranaka, T.; Ogihara, Y. A new insight into application for barley chromosome addition lines of common wheat: Achievement of stigmasterol accumulation. Plant Physiol. 2011, 157, 1555–1567. [Google Scholar] [CrossRef] [Green Version]
- He, J.-X.; Fujioka, S.; Li, T.-C.; Kang, S.G.; Seto, H.; Takatsuto, S.; Yosshida, S.; Jang, J.-C. Sterols regulate development and gene expression in Arabidopsis. Plant Physiol. 2003, 131, 1258–1269. [Google Scholar] [CrossRef] [PubMed]
- El-Tantawy, A.A.; Azoz, S.N. Enhancement of growth and increased productivity of fresh herb and aromatic oil in basil plant by foliar application with stigmasterol. JMB 2019, 3, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Hussein, H.A.A.; Alshammari, S.O.; Elkady, F.M.; Ramadan, A.A.; Kenawy, S.K.; Abdelkawy, A.M. Radio-Protective Effects of Stigmasterol on Wheat (Triticum aestivum L.) Plants. Antioxidants 2022, 11, 1144. [Google Scholar] [CrossRef]
- Willemsen, V.; Friml, J.; Grebe, M.; Van Den Toorn, A.; Palme, K.; Scheres, B. Cell polarity and PIN protein positioning in Arabidopsis require STEROL METHYLTRANSFERASE1 function. Plant Cell 2003, 15, 612–625. [Google Scholar] [CrossRef] [Green Version]
- Batista, B.D.; Dourado, M.N.; Figueredo, E.F.; Hortencio, R.O.; Marques, J.P.R.; Piotto, F.A.; Bonatelli, M.L.; Settles, M.L.; Azevedo, J.L.; Quecine, M.C. The auxin-producing Bacillus thuringiensis RZ2MS9 promotes the growth and modifies the root architecture of tomato (Solanum lycopersicum cv. Micro-Tom). Arch. Microbiol. 2021, 203, 3869–3882. [Google Scholar] [CrossRef] [PubMed]
- Arnqvist, L.; Persson, M.; Jonsson, L.; Dutta, P.C.; Sitbon, F. Overexpression of CYP710A1 and CYP710A4 in transgenic Arabidopsis plants increases the level of stigmasterol at the expense of sitosterol. Planta 2008, 227, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Ming, R.; Alam, M.; Schuler, M.A. Comparison of cytochrome P450 genes from six plant genomes. Trop. Plant Biol. 2008, 1, 216–235. [Google Scholar] [CrossRef]
- Griebel, T.; Zeier, J. A role for β-sitosterol to stigmasterol conversion in plant–pathogen interactions. Plant J. 2010, 63, 254–268. [Google Scholar] [CrossRef]
- Fabro, G.; Di Rienzo, J.A.; Voigt, C.A.; Savchenko, T.; Dehesh, K.; Somerville, S.; Alvarez, M.E. Genome-wide expression profiling Arabidopsis at the stage of Golovinomyces cichoracearum haustorium formation. Plant Physiol. 2008, 146, 1421–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Senthil-Kumar, M.; Ryu, C.M.; Kang, L.; Mysore, K.S. Phytosterols play a key role in plant innate immunity against bacterial pathogens by regulating nutrient efflux into the apoplast. Plant Physiol. 2012, 158, 1789–1802. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Li, G.; Wang, Q.; Meng, Q.; Xu, F.; Chen, Q.; Liu, F.; Hu, Y.; Luo, M. GhCYP710A1 Participates in Cotton Resistance to Verticillium Wilt by Regulating Stigmasterol Synthesis and Plasma Membrane Stability. Int. J. Mol. Sci. 2022, 23, 8437. [Google Scholar] [CrossRef] [PubMed]
- Chitwood, D.J.; Lusby, W.R. Metabolism of plant sterols by nematodes. Lipids 1991, 26, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Chitwood, D.J. Biochemistry and functions of nematode steroids. Crit. Rev. Biochem. Mol. Biol. 1999, 34, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Matyash, V.; Entchev, E.V.; Mende, F.; Wilsch-Bräuninger, M.; Thiele, C.; Schmidt, A.W.; Knölker, H.J.; Ward, S.; Kurzchalia, T.V.; Ahringer, J. Sterol-derived hormone (s) controls entry into diapause in Caenorhabditis elegans by consecutive activation of DAF-12 and DAF-16. PLoS Biol. 2004, 2, e280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersson, E.V.; Nahar, N.; Dahlin, P.; Broberg, A.; Tröger, R.; Dutta, P.C.; Jonsson, L.; Sitbon, F. Conversion of exogenous cholesterol into glycoalkaloids in potato shoots, using two methods for sterol solubilisation. PLoS ONE 2013, 8, e82955. [Google Scholar] [CrossRef]
- Itkin, M.; Rogachev, I.; Alkan, N.; Rosenberg, T.; Malitsky, S.; Masini, L.; Meir, S.; Iijima, Y.; Aoki, K.; de Vos, R.; et al. GLYCOALKALOID METABOLISM1 is required for steroidal alkaloid glycosylation and prevention of phytotoxicity in tomato. Plant Cell 2011, 23, 4507–4525. [Google Scholar] [CrossRef] [Green Version]
- Petersson, E.V.; Arif, U.; Schulzova, V.; Krtková, V.; Hajšlová, J.; Meijer, J.; Andersson, H.C.; Jonsson, L.; Sitbon, F. Glycoalkaloid and calystegine levels in table potato cultivars subjected to wounding, light, and heat treatments. J. Agric. Food Chem. 2013, 61, 5893–5902. [Google Scholar] [CrossRef]
- Zhang, W.; Zuo, C.; Chen, Z.; Kang, Y.; Qin, S. RNA sequencing reveals that both abiotic and biotic stress-responsive genes are induced during expression of steroidal glycoalkaloid in potato tuber subjected to light exposure. Genes 2019, 10, 920. [Google Scholar] [CrossRef] [Green Version]
- Fabian, M.L.; Zhang, C.; Sun, J.; Price, N.P.; Chen, P.; Clarke, C.R.; Jones, R.W.; Stommel, J.R. Steroidal glycoalkaloids contribute to anthracnose resistance in Solanum lycopersicum. bioRxiv 2022, 503224. [Google Scholar] [CrossRef]
- Jing, X.; Grebenok, R.J.; Brehmer, S.T. Plant sterols and host suitability for generalist and specialist caterpillars. J. Insect Physiol. 2012, 58, 235–244. [Google Scholar] [CrossRef]
- Hallmann, J.; Subbotin, S.A. Methods for extraction, processing and detection of plant and soil nematodes. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 3rd ed.; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI: Wallingford, UK, 2018; pp. 87–119. [Google Scholar]
- Blight, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Azadmard-Damirchi, S.; Dutta, P.C. Novel solid-phase extraction method to separate 4-desmethyl-, 4-monomethyl- and 4,4′-dimethylsterols in vegetable oils. J. Chromatogr. A 2006, 1108, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, J.; Daub, M.; Wesemael, W. Estimating numbers. In Techniques for Work with Plant and Soil Nematodes; Perry, R.N., Hunt, D.J., Subbotin, S.A., Eds.; CABI: Oxdordshire, UK, 2020; p. 56. [Google Scholar]
- Seethepalli, A.; Dhakal, K.; Griffiths, M.; Guo, H.; Freschet, G.T.; York, L.M. RhizoVision Explorer: Open-source software for root image analysis and measurement standardization. AoB Plants 2021, 13, plab056. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Micro-Tom | Root Fresh Weight (g) | Shoot Fresh Weight (g) | Shoot Dry Weight in % | Shoot Length (cm) |
|---|---|---|---|---|
| Wt | 3.50 ± 0.50 a | 4.32 ± 0.53 a | 11.57 a | 18.62 ± 1.07 a |
| cyp710a11 | 2.22 ± 0.24 b | 2.82 ± 0.39 b | 10.92 a | 10.58 ± 1.08 b |
| Sterols | Micro-Tom Roots | Micro-Tom Leaves | ||
|---|---|---|---|---|
| wt | cyp710a11 | wt | cyp710a11 | |
| Cholesterol | 14.78 ± 1.38 a | 14.69 ± 0.74 a | 8.99 ± 1.13 a | 9.69 ± 0.44 a |
| Campesterol | 13.85 ± 0.46 a | 13.09 ± 1.77 a | 14.18 ± 1.99 a | 16.91 ± 3.13 a |
| Stigmasterol | 43.36 ± 1.37 a | 0 b | 69.75 ± 2.87 a | 0 b |
| β-Sitosterol | 28.01 ± 2.12 a | 72.22 ± 2.04 b | 7.20 ± 2.76 a | 73.39 ± 3.52 b |
| Micro-Tom | GI Experiment 1 | GI Experiment 2 |
|---|---|---|
| Wt | 6.67 ± 0.48 a | 6.43 ± 0.51 a |
| cyp710a11 | 6.13 ± 0.67 a | 7.00 ± 0.60 a |
| Micro-Tom | Median Number of Roots | Max. Number of Roots | Number of Root Tips | Total Root Length (mm) | Egg mass/mm Root |
|---|---|---|---|---|---|
| wt | 10.80 ± 2.86 a | 34.40 ± 6.35 a | 2967.20 ± 756.11 a | 95,996.17 ± 25,993.51 a | 545.62 ± 117.76 a |
| cyp710a11 | 10.00 ±2.17 a | 28.00 ± 3.81 a | 2124.83 ± 324.30 b | 64,667.00 ± 12,144.3 b | 670.41 ± 83.39 a |
| p-value | 0.61 | 0.07 | 0.03 | 0.03 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabianca, A.; Ruthes, A.C.; Pawlowski, K.; Dahlin, P. Tomato Sterol 22-desaturase Gene CYP710A11: Its Roles in Meloidogyne incognita Infection and Plant Stigmasterol Alteration. Int. J. Mol. Sci. 2022, 23, 15111. https://doi.org/10.3390/ijms232315111
Cabianca A, Ruthes AC, Pawlowski K, Dahlin P. Tomato Sterol 22-desaturase Gene CYP710A11: Its Roles in Meloidogyne incognita Infection and Plant Stigmasterol Alteration. International Journal of Molecular Sciences. 2022; 23(23):15111. https://doi.org/10.3390/ijms232315111
Chicago/Turabian StyleCabianca, Alessandro, Andrea Caroline Ruthes, Katharina Pawlowski, and Paul Dahlin. 2022. "Tomato Sterol 22-desaturase Gene CYP710A11: Its Roles in Meloidogyne incognita Infection and Plant Stigmasterol Alteration" International Journal of Molecular Sciences 23, no. 23: 15111. https://doi.org/10.3390/ijms232315111