
Optimal Intravenous Administration Procedure for Efficient Delivery of Canine Adipose-Derived Mesenchymal Stem Cells
 , , , and
, , , and 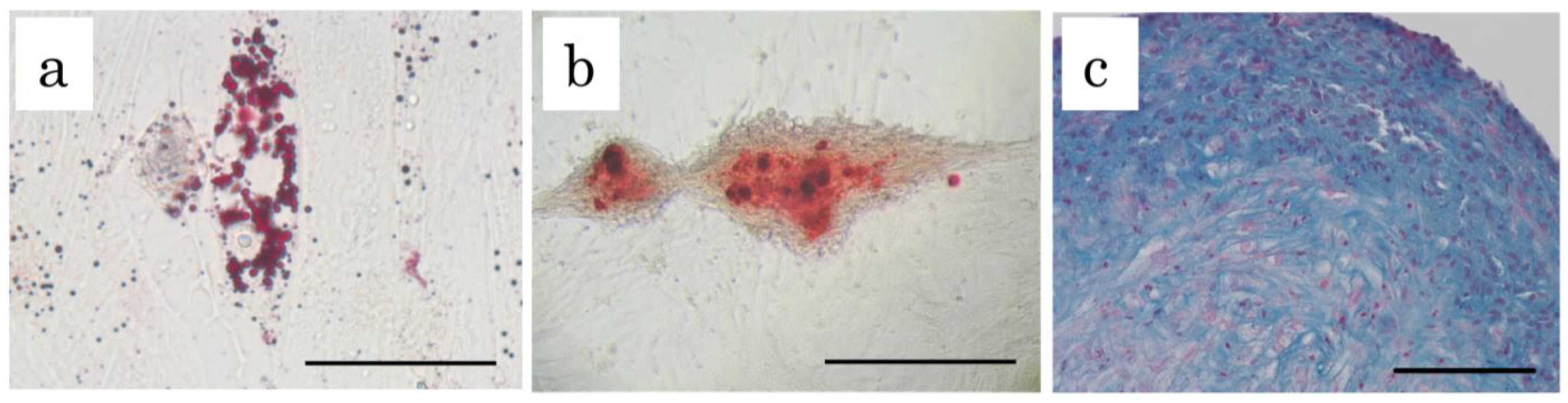{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of cADSC
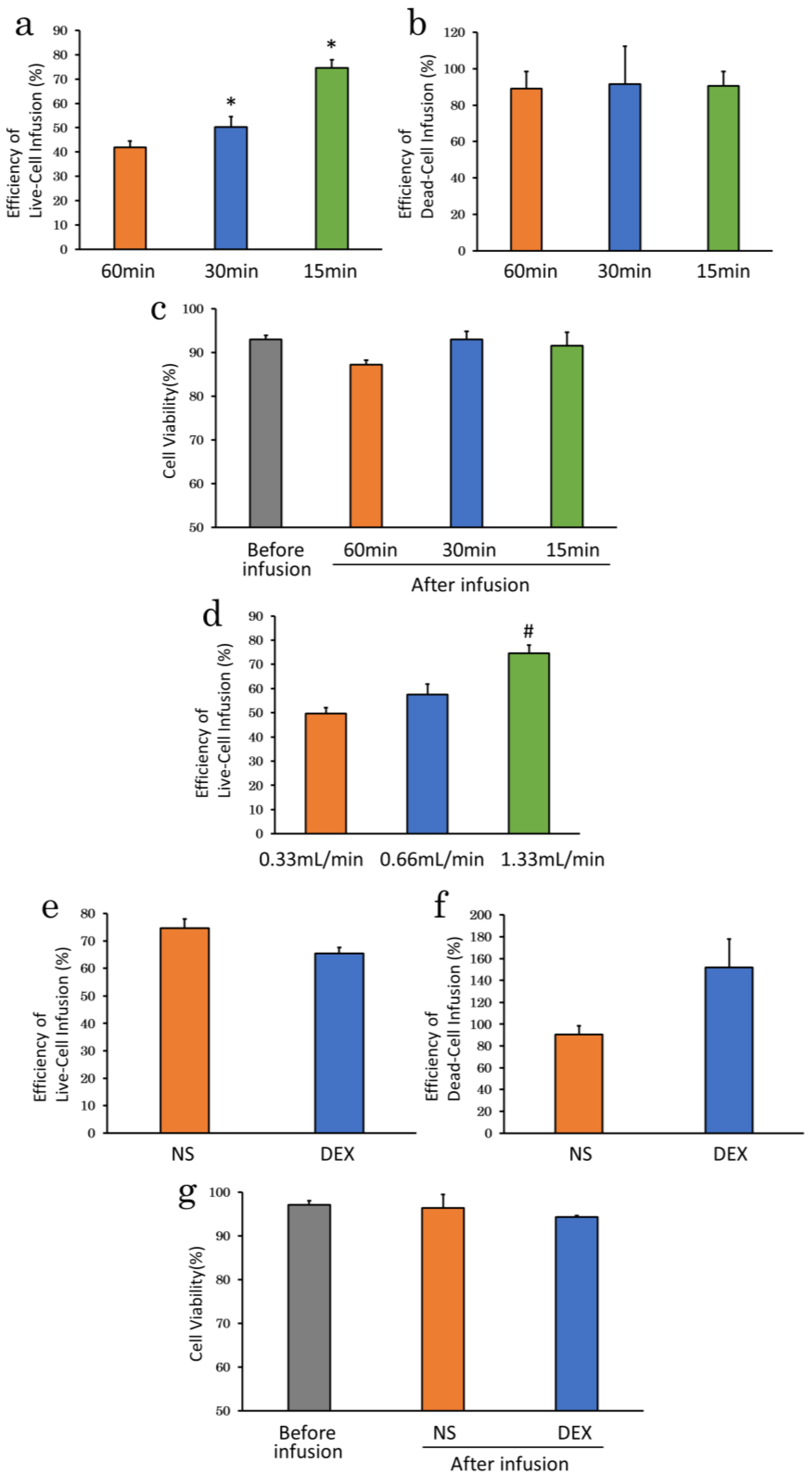
2.2. Evaluation of Cell Delivery and Cell Viability under Each Infusion Condition
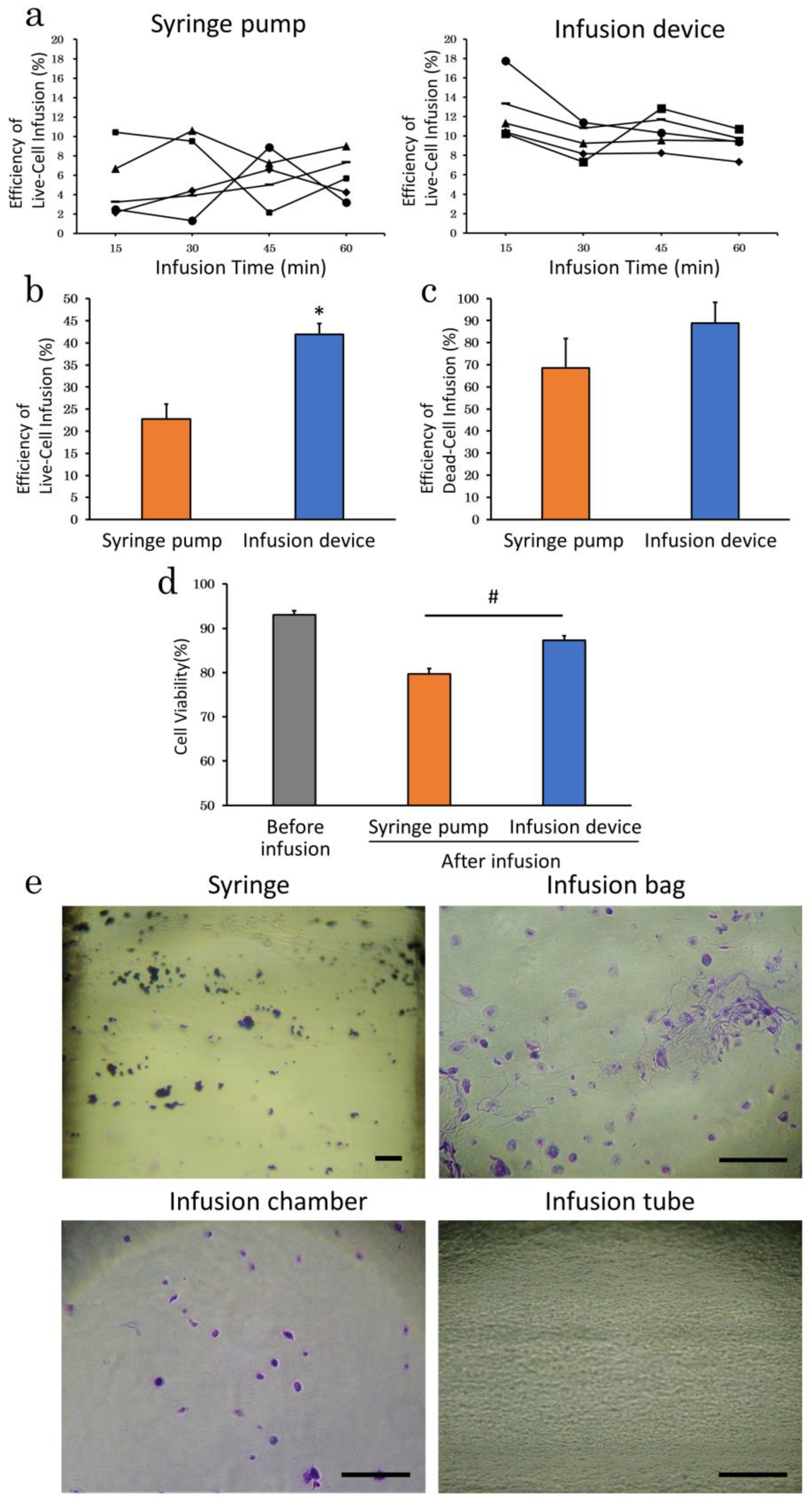
2.2.1. Infusion Apparatus
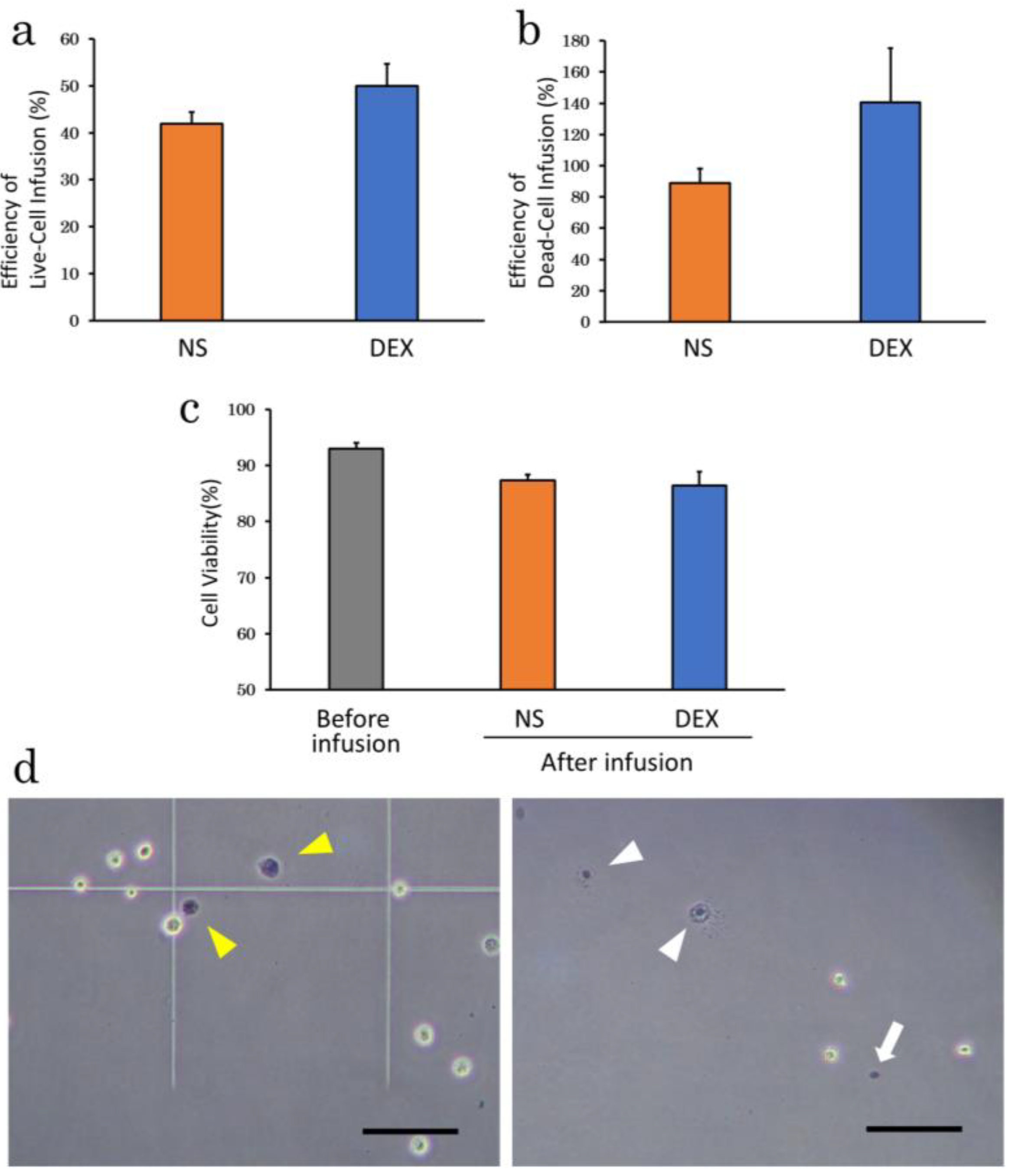
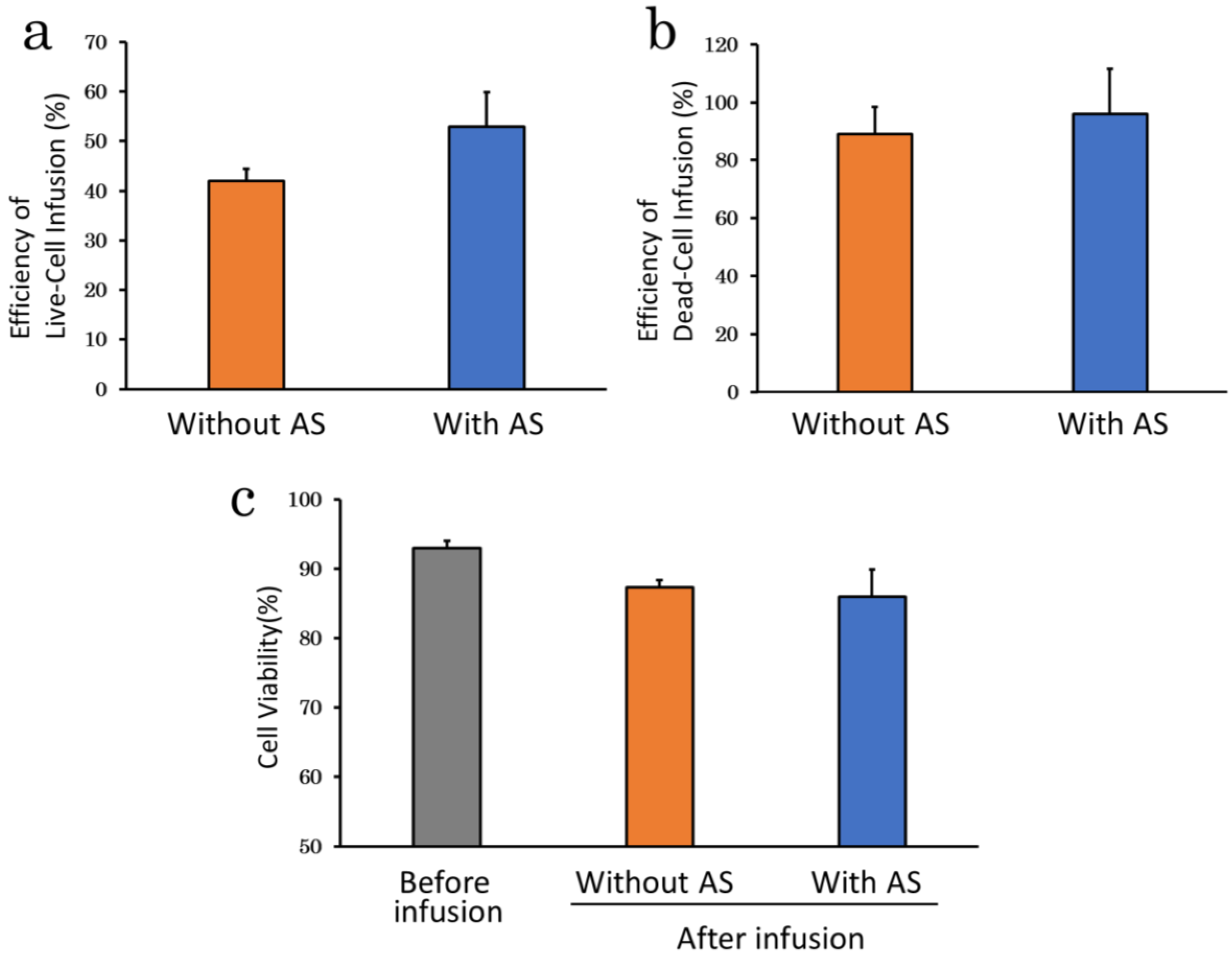
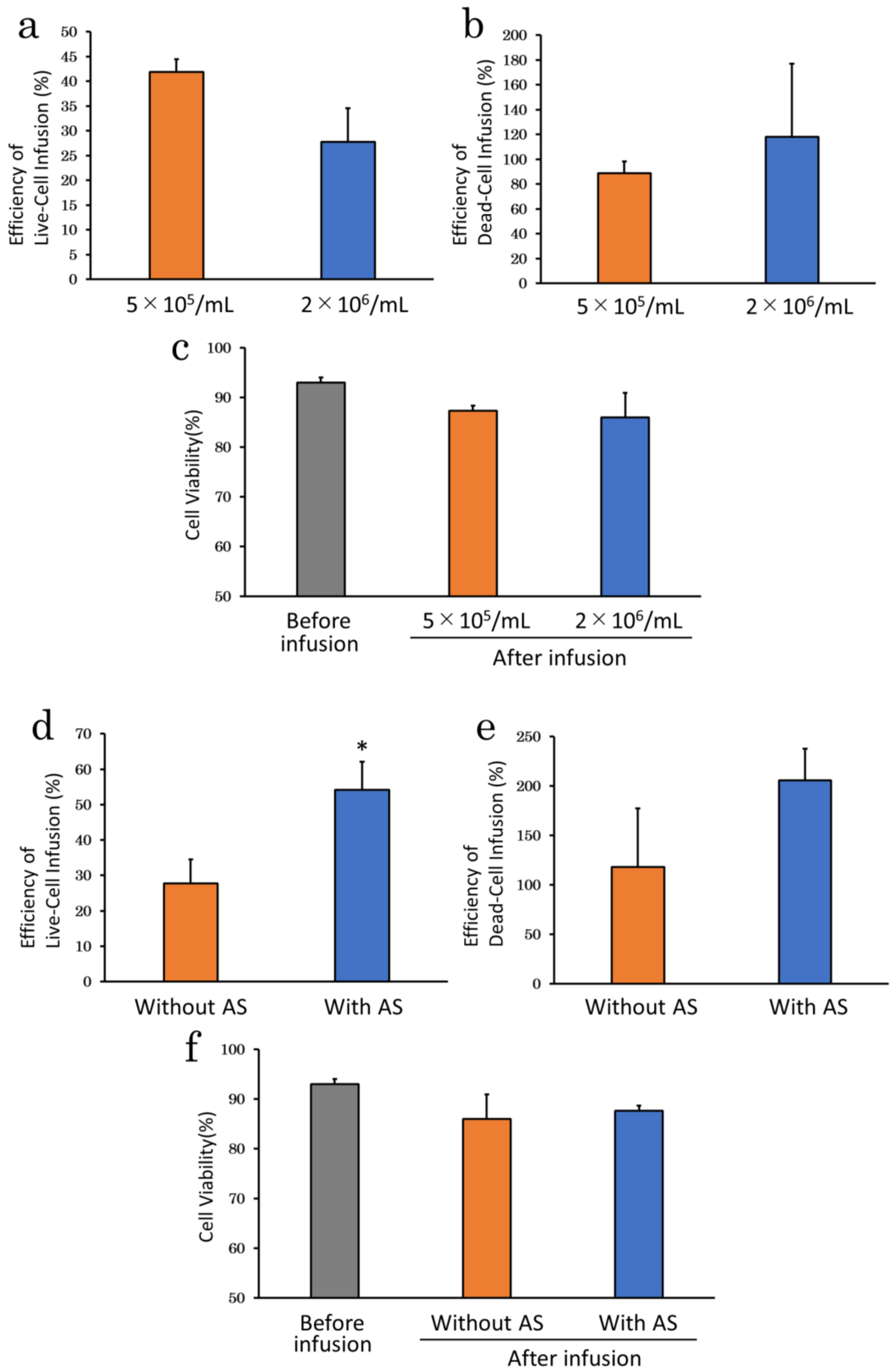
2.2.2. Suspension Solution
2.2.3. Serum Supplementation
2.2.4. Infusion Time
2.2.5. Cell Density
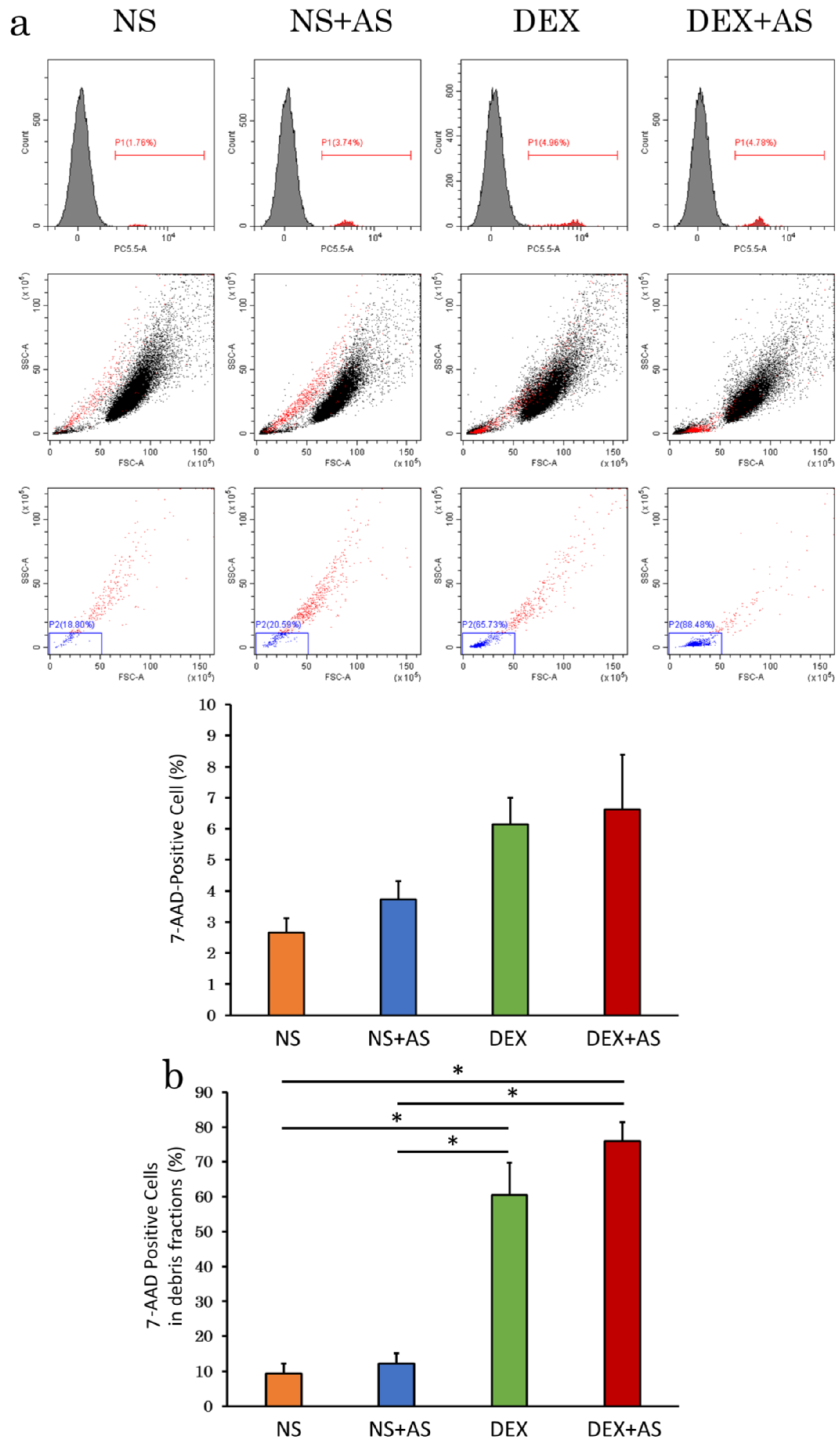
2.3. Evaluation of Cell Viability by Flow Cytometry
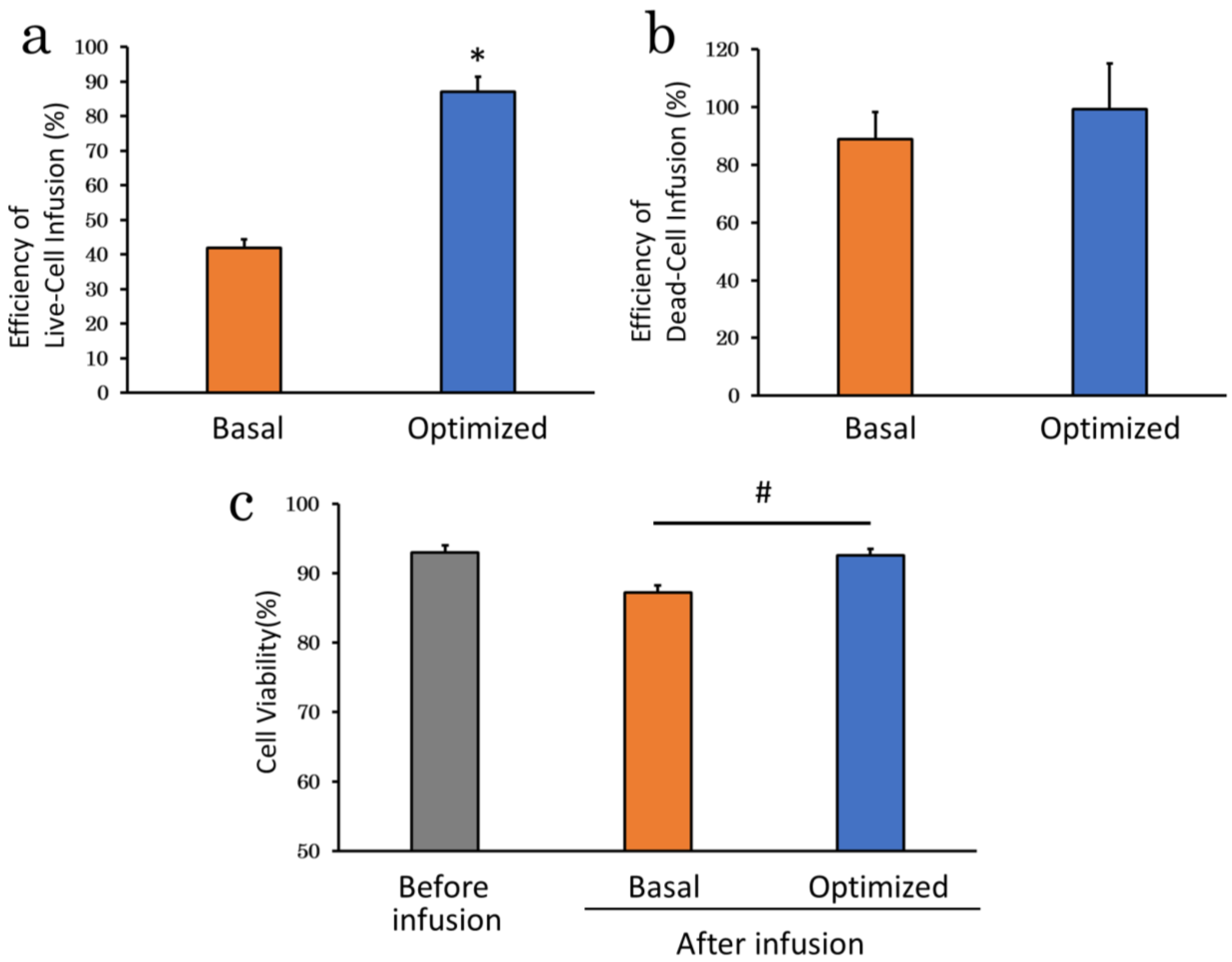
2.4. Assessment of Optimized Infusion Procedures
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Isolation, Culture and Cryopreservation of cADSC
4.3. Characterization of cADSC Surface Markers
4.4. Differentiation Assay
4.5. Collection of Canine AS
4.6. Preparation of cADSC
4.7. Efficiency of Cell Infusion and Cell Viability
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andrzejewska, A.; Dabrowska, S.; Lukomska, B.; Janowski, M. Mesenchymal stem cells for neurological disorders. Adv. Sci. 2021, 8, 2002944. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, C.; Noël, D. Mesenchymal stem cells in osteoarticular diseases. Regen. Med. 2011, 6, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Bagno, L.; Hatzistergos, K.E.; Balkan, W.; Hare, J.M. Mesenchymal stem cell-based therapy for cardiovascular disease: Progress and challenges. Mol. Ther. 2018, 26, 1610–1623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markov, A.; Thangavelu, L.; Aravindhan, S.; Zekiy, A.O.; Jarahian, M.; Chartrand, M.S.; Pathak, Y.; Marofi, F.; Shamlou, S.; Hassanzadeh, A. Mesenchymal stem/stromal cells as a valuable source for the treatment of immune-mediated disorders. Stem. Cell Res. Ther. 2021, 12, 192. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Blanc, K.L.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Friedenstein, A.J.; Piatetzky-Shapiro, I.I.; Petrakova, K.V. Osteogenesis in transplants of bone marrow cells. J. Embryol. Exp. Morphol. 1966, 16, 381–390. [Google Scholar] [CrossRef]
- Tavakoli, S.; Ghaderi, H.R.J.; Shariati, A.; Jahangiryan, A.; Jadidi, F.; Kouhbanani, M.A.J.; Hassanzadeh, A.; Zamani, M.; Javidi, K.; Naimi, A. Mesenchymal stromal cells; A new horizon in regenerative medicine. J. Cell Physiol. 2020, 235, 9185–9210. [Google Scholar] [CrossRef]
- Iwashima, S.; Ozaki, T.; Maruyama, S.; Saka, Y.; Kobori, M.; Omae, K.; Yamaguchi, H.; Niimi, T.; Toriyama, K.; Kamei, Y.; et al. Novel culture system of mesenchymal stromal cells from human subcutaneous adipose tissue. Stem Cells Dev. 2009, 18, 533–543. [Google Scholar] [CrossRef]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. A 2018, 93, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal stem cells for regenerative medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef]
- Mattar, P.; Bieback, K. Comparing the immunomodulatory properties of bone marrow, adipose tissue, and birth-associated tissue mesenchymal stromal cells. Front. Immunol. 2015, 6, 560. [Google Scholar] [CrossRef] [Green Version]
- Chu, D.T.; Phuong, T.N.T.; Tien, N.L.B.; Tran, D.K.; Thanh, V.V.; Quang, T.L.; Truong, D.T.; Pham, V.H.; Ngoc, V.T.N.; Chu-Dinh, T.; et al. An update on the progress of isolation, culture, storage, and clinical application of human bone marrow mesenchymal stem/stromal cells. Int. J. Mol. Sci. 2020, 21, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, X.L.; Zhang, Y.; Li, X.; Fu, Q.L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell Mol. Life Sci. 2020, 77, 2771–2794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, Y.; Koh, Y.G.; Choi, Y.J.; Kim, S.H.; Yoon, D.S.; Lee, M.; Lee, J.W. Characterization of adipose tissue-derived stromal vascular fraction for clinical application to cartilage regeneration. In Vitro Cell Dev. Biol. Anim. 2015, 51, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Neri, S. Genetic stability of mesenchymal stromal cells for regenerative medicine applications: A fundamental biosafety aspect. Int. J. Mol. Sci. 2019, 20, 2406. [Google Scholar] [CrossRef] [Green Version]
- Moseley, T.A.; Zhu, M.; Hedrick, M.H. Adipose-derived stem and progenitor cells as fillers in plastic and reconstructive surgery. Plast. Reconstr. Surg. 2006, 118, 121S–128S. [Google Scholar] [CrossRef] [Green Version]
- Laloze, J.; Fiévet, L.; Desmoulière, A. Adipose-derived mesenchymal stromal cells in regenerative medicine: State of play, current clinical trials, and future prospects. Adv. Wound Care (New Rochelle) 2021, 10, 24–48. [Google Scholar] [CrossRef]
- Furuhashi, K.; Tsuboi, N.; Shimizu, A.; Katsuno, T.; Kim, H.; Saka, Y.; Ozaki, T.; Sado, Y.; Imai, E.; Matsuo, S.; et al. Serum-starved adipose-derived stromal cells ameliorate crescentic GN by promoting immunoregulatory macrophages. J. Am. Soc. Nephrol. 2013, 24, 587–603. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef]
- Miguel, M.P.D.; Fuentes-Julián, S.; Blázquez-Martínez, A.; Pascual, C.Y.; Aller, M.A.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive properties of mesenchymal stem cells: Advances and applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef]
- Omar, R.E.I.; Beroud, J.; Stoltz, J.F.; Menu, P.; Velot, E.; Decot, V. Umbilical cord mesenchymal stem cells: The new gold standard for mesenchymal stem cell-based therapies? Tissue Eng. Part B Rev. 2014, 20, 523–544. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Nagamura-Inoue, T.; Takahashi, A.; Mori, Y.; Yamamoto, Y.; Shimazu, T.; Tsunoda, H.; Tojo, A. Immunosuppressive properties of Wharton’s jelly-derived mesenchymal stromal cells in vitro. Int. J. Hematol. 2015, 102, 368–378. [Google Scholar] [CrossRef]
- Hsieh, J.Y.; Wang, H.W.; Chang, S.J.; Liao, K.H.; Lee, I.H.; Lin, W.S.; Wu, C.H.; Lin, W.Y.; Cheng, S.M. Mesenchymal stem cells from human umbilical cord express preferentially secreted factors related to neuroprotection, neurogenesis, and angiogenesis. PLoS ONE 2013, 8, e72604. [Google Scholar] [CrossRef] [Green Version]
- Mukai, T.; Nagamura-Inoue, T.; Shimazu, T.; Mori, Y.; Takahashi, A.; Tsunoda, H.; Yamaguchi, S.; Tojo, A. Neurosphere formation enhances the neurogenic differentiation potential and migratory ability of umbilical cord-mesenchymal stromal cells. Cytotherapy 2016, 18, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Liu, R.; Jiang, J.; Peng, J.; Yang, C.; Zhang, W.; Wang, S.; Song, J. What is the impact of human umbilical cord mesenchymal stem cell transplantation on clinical treatment? Stem Cell Res. Ther. 2020, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Racz, G.Z.; Kadar, K.; Foldes, A.; Kallo, K.; Perczel-Kovach, K.; Keremi, B.; Nagy, A.; Varga, G. Immunomodulatory and potential therapeutic role of mesenchymal stem cells in periodontitis. J. Physiol. Pharmacol. 2014, 65, 327–339. [Google Scholar]
- Ledesma-Martínez, E.; Mendoza-Núñez, V.M.; Santiago-Osorio, E. Mesenchymal stem cells derived from dental pulp: A review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef] [Green Version]
- Mattei, V.; Santacroce, C.; Tasciotti, V.; Martellucci, S.; Santilli, F.; Manganelli, V.; Piccoli, L.; Misasi, R.; Sorice, M.; Garofalo, T. Role of lipid rafts in neuronal differentiation of dental pulp-derived stem cells. Exp. Cell Res. 2015, 339, 231–240. [Google Scholar] [CrossRef]
- Muroi, K.; Miyamura, K.; Okada, M.; Yamashita, T.; Murata, M.; Ishikawa, T.; Uike, N.; Hidaka, M.; Kobayashi, R.; Imamura, M.; et al. Bone marrow-derived mesenchymal stem cells (JR-031) for steroid-refractory grade III or IV acute graft-versus-host disease: A phase II/III study. Int. J. Hematol. 2016, 103, 243–250. [Google Scholar] [CrossRef]
- Jurado, M.; Mata, C.D.L.; Ruiz-García, A.; López-Fernández, E.; Espinosa, O.; Remigia, M.J.; Moratalla, L.; Goterris, R.; García-Martín, P.; Ruiz-Cabello, F. Adipose tissue-derived mesenchymal stromal cells as part of therapy for chronic graft-versus-host disease: A phase I/II study. Cytotherapy 2017, 19, 927–936. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Y.; Hu, B.; Liu, J.; Kong, P.; Lou, S.; Su, Y.; Yang, T.; Li, H.; Liu, Y.; et al. Phase II multicenter, randomized, double-blind controlled study of efficacy and safety of umbilical cord-derived mesenchymal stromal cells in the prophylaxis of chronic graft-versus-host disease after HLA-haploidentical stem-cell transplantation. J. Clin. Oncol. 2016, 34, 2843–2850. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhu, C.Y.; Ma, D.X.; Gu, Z.Y.; Xu, C.C.; Wang, F.Y.; Chen, J.G.; Liu, C.J.; Guan, L.X.; Gao, R.; et al. Efficacy and safety of mesenchymal stromal cells for the prophylaxis of chronic graft-versus-host disease after allogeneic hematopoietic stem cell transplantation: A meta-analysis of randomized controlled trials. Ann. Hematol. 2018, 97, 1941–1950. [Google Scholar] [CrossRef] [PubMed]
- Grégoire, C.; Ritacco, C.; Hannon, M.; Seidel, L.; Delens, L.; Belle, L.; Dubois, S.; Vériter, S.; Lechanteur, C.; Briquet, A.; et al. Comparison of mesenchymal stromal cells from different origins for the treatment of graft-vs.-host-disease in a humanized mouse model. Front. Immunol. 2019, 10, 619. [Google Scholar] [CrossRef] [PubMed]
- Voga, M.; Adamic, N.; Vengust, M.; Majdic, G. Stem cells in veterinary medicine-current state and treatment options. Front. Vet. Sci. 2020, 7, 278. [Google Scholar] [CrossRef] [PubMed]
- Bearden, R.N.; Huggins, S.S.; Cummings, K.J.; Smith, R.; Gregory, C.A.; Saunders, W.B. In-vitro characterization of canine multipotent stromal cells isolated from synovium, bone marrow, and adipose tissue: A donor-matched comparative study. Stem Cell Res. Ther. 2017, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Kang, B.J.; Ryu, H.H.; Park, S.S.; Koyama, Y.; Kikuchi, M.; Woo, H.M.; Kim, W.H.; Kweon, O.K. Comparing the osteogenic potential of canine mesenchymal stem cells derived from adipose tissues, bone marrow, umbilical cord blood, and Wharton’s jelly for treating bone defects. J. Vet. Sci. 2012, 13, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.M.; Dow, S.W. Concise review: Stem cell trials using companion animal disease models. Stem Cells 2016, 34, 1709–1729. [Google Scholar] [CrossRef]
- Kabat, M.; Bobkov, I.; Kumar, S.; Grumet, M. Trends in mesenchymal stem cell clinical trials 2004-2018: Is efficacy optimal in a narrow dose range? Stem. Cells Transl. Med. 2020, 9, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Rajendran, R.L.; Jogalekar, M.P.; Gangadaran, P.; Ahn, B.C. Noninvasive in vivo cell tracking using molecular imaging: A useful tool for developing mesenchymal stem cell-based cancer treatment. World J. Stem Cells 2020, 12, 1492–1510. [Google Scholar] [CrossRef]
- Ikebe, C.; Suzuki, K. Mesenchymal stem cells for regenerative therapy: Optimization of cell preparation protocols. Biomed. Res. Int. 2014, 2014, 951512. [Google Scholar] [CrossRef] [Green Version]
- Wiese, D.M.; Wood, C.A.; Braid, L.R. From vial to vein: Crucial gaps in mesenchymal stromal cell clinical trial reporting. Front. Cell Dev. Biol. 2022, 10, 867426. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, M.; Liao, X.; Li, S.; Yan, J.; Fan, L.; She, W.; Song, J.; Liu, H. Effects of storage culture media, temperature and duration on human adipose-derived stem cell viability for clinical use. Mol. Med. Rep. 2019, 19, 2189–2201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngo, A.T.L.; Le, H.M.; Trinh, N.T.H.; Jun, A.P.G.; Bach, T.Q.; Bui, H.T.H.; Hoang, V.T.; Bui, A.V.; Nguyen, L.T.; Hoang, D.M. Clinically relevant preservation conditions for mesenchymal stem/stromal cells derived from perinatal and adult tissue sources. J. Cell Mol. Med. 2021, 25, 10747–10760. [Google Scholar] [CrossRef]
- Kubrova, E.; Qu, W.; Galvan, M.L.; Paradise, C.R.; Yang, J.; Dietz, A.B.; Dudakovic, A.; Smith, J.; Wijnen, A.J. Hypothermia and nutrient deprivation alter viability of human adipose-derived mesenchymal stem cells. Gene 2020, 722, 144058. [Google Scholar] [CrossRef] [PubMed]
- Pal, R.; Hanwate, M.; Totey, S.M. Effect of holding time, temperature and different parenteral solutions on viability and functionality of adult bone marrow-derived mesenchymal stem cells before transplantation. J. Tissue Eng. Regen. Med. 2008, 2, 436–444. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, B.; Xue, G.; Zhao, J.; Li, R.; Liu, Z.; Niu, B. Effects of storage solutions on the viability of human umbilical cord mesenchymal stem cells for transplantation. Cell Transpl. 2013, 22, 1075–1086. [Google Scholar] [CrossRef]
- Zhang, F.; Ren, H.; Shao, X.; Zhuang, C.; Chen, Y.; Qi, N. Preservation media, durations and cell concentrations of short-term storage affect key features of human adipose-derived mesenchymal stem cells for therapeutic application. PeerJ 2017, 17, e3301. [Google Scholar] [CrossRef] [Green Version]
- Sultana, T.; Dayem, A.A.; Lee, S.B.; Cho, S.G.; Lee, J.I. Effects of carrier solutions on the viability and efficacy of canine adipose-derived mesenchymal stem cells. BMC Vet. Res. 2022, 18, 26. [Google Scholar] [CrossRef]
- Heng, B.C.; Cowan, C.M.; Basu, A.S. Temperature and calcium ions affect aggregation of mesenchymal stem cells in phosphate buffered saline. Cytotechnology 2008, 58, 69–75. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.L.; Kinnunen, T.; Boltze, J.; Nystedt, J.; Jolkkonen, J. Clumping and viability of bone marrow derived mesenchymal stromal cells under different preparation procedures: A flow cytometry-based in vitro study. Stem Cells Int. 2016, 2016, 1764938. [Google Scholar] [CrossRef] [Green Version]
- Parsha, K.; Mir, O.; Satani, N.; Yang, B.; Guerrero, W.; Mei, Z.; Cai, C.; Chen, P.R.; Gee, A.; Hanley, P.J.; et al. Mesenchymal stromal cell secretomes are modulated by suspension time, delivery solution, passage through catheter, and exposure to adjuvants. Cytotherapy 2017, 19, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Wahlberg, B.; Ghuman, H.; Liu, J.R.; Modo, M. Ex vivo biomechanical characterization of syringe-needle ejections for intracerebral cell delivery. Sci. Rep. 2018, 8, 9194. [Google Scholar] [CrossRef] [PubMed]
- Onishi, K.; Jones, D.L.; Riester, S.M.; Lewallen, E.A.; Lewallen, D.G.; Sellon, D.L.; Dietz, A.B.; Qu, W.; Wijnen, A.J.; Smith, J. Human adipose-derived mesenchymal stromal/stem cells remain viable and metabolically active following needle passage. PM. R. 2016, 8, 844–854. [Google Scholar] [CrossRef] [PubMed]
- Mamidi, M.K.; Singh, G.; Husin, J.M.; Nathan, K.G.; Sasidharan, G.; Zakaria, Z.; Bhonde, R.; Majumdar, A.S.; Das, A.K. Impact of passing mesenchymal stem cells through smaller bore size needles for subsequent use in patients for clinical or cosmetic indications. J. Transl. Med. 2012, 10, 229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoury, R.E.; Misra, V.; Sharma, S.; Cox, C.S.; Walker, P.; Grotta, J.C.; Gee, A.; Suzuki, S.; Savitz, S.I. The effect of transcatheter injections on cell viability and cytokine release of mononuclear cells. AJNR Am. J. Neuroradiol. 2010, 31, 1488–1492. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.A.; Jimenez, F.; Gerber, M.H.; Aroom, K.R.; Shah, S.K.; Harting, M.T.; Gill, B.S.; Savitz, S.I.; Cox, C.S., Jr. Effect of needle diameter and flow rate on rat and human mesenchymal stromal cell characterization and viability. Tissue Eng. Part. C Methods 2010, 16, 989–997. [Google Scholar] [CrossRef] [Green Version]
- Heng, B.C.; Hsu, S.H.; Cowan, C.M.; Liu, A.; Tai, J.; Chan, Y.; Sherman, W.; Basu, S. Transcatheter injection-induced changes in human bone marrow-derived mesenchymal stem cells. Cell Transplant. 2009, 18, 1111–1121. [Google Scholar] [CrossRef] [Green Version]
- Quimby, J.M.; Webb, T.L.; Habenicht, L.M.; Dow, S.W. Safety and efficacy of intravenous infusion of allogeneic cryopreserved mesenchymal stem cells for treatment of chronic kidney disease in cats: Results of three sequential pilot studies. Stem Cell Res. Ther. 2013, 4, 48. [Google Scholar] [CrossRef] [Green Version]
- Rashidghamat, E.; Kadiyirire, T.; Ayis, S.; Petrof, G.; Liu, L.; Pullabhatla, V.; Ainali, C.; Guy, A.; Aristodemou, S.; McMillan, J.R.; et al. Phase I/II open-label trial of intravenous allogeneic mesenchymal stromal cell therapy in adults with recessive dystrophic epidermolysis bullosa. J. Am. Acad. Dermatol. 2020, 83, 447–454. [Google Scholar] [CrossRef]
- Sánchez-Guijo, F.; Caballero-Velázquez, T.; López-Villar, O.; Redondo, A.; Parody, R.; Martínez, C.; Olavarría, E.; Andreu, E.; Prósper, F.; Díez-Campelo, M.; et al. Sequential third-party mesenchymal stromal cell therapy for refractory acute graft-versus-host disease. Biol. Blood Marrow Transplant. 2014, 20, 1580–1585. [Google Scholar] [CrossRef] [Green Version]
- Bang, O.Y.; Lee, J.S.; Lee, P.H.; Lee, G. Autologous mesenchymal stem cell transplantation in stroke patients. Ann. Neurol. 2005, 57, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Huang, L.; Zhang, C.; Wang, Y.; Zhang, R.; Tu, Z.; Wang, Y.; Zhou, X.; Xiao, Z.; Liu, Z.; et al. Therapeutic evidence of umbilical cord-derived mesenchymal stem cell transplantation for cerebral palsy: A randomized, controlled trial. Stem Cell Res. Ther. 2020, 11, 43. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Niu, C.; Li, R.; Huang, T.; Wang, Y.; Huang, M.; Ji, N.; Zheng, Y.; Chen, X.; Shi, L.; et al. Treatment of severe COVID-19 with human umbilical cord mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, M.F.; Andrade, C.M.; Mello, M.J.E.; Crispi, C.P. Effect of temperature on fluidity of irrigation fluids. Br. J. Anaesth. 2011, 106, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Jarocha, D.; Milczarek, O.; Wedrychowicz, A.; Kwiatkowski, S.; Majka, M. Continuous improvement after multiple mesenchymal stem cell transplantations in a patient with complete spinal cord injury. Cell Transpl. 2015, 24, 661–672. [Google Scholar] [CrossRef]
- Riordan, N.H.; Morales, I.; Fernández, G.; Allen, N.; Fearnot, N.E.; Leckrone, M.E.; Markovich, D.J.; Mansfield, D.; Avila, D.; Patel, A.N.; et al. Clinical feasibility of umbilical cord tissue-derived mesenchymal stem cells in the treatment of multiple sclerosis. J. Transl. Med. 2018, 16, 57. [Google Scholar] [CrossRef] [Green Version]
- Glassberg, M.K.; Minkiewicz, J.; Toonkel, R.L.; Simonet, E.S.; Rubio, G.A.; DiFede, D.; Shafazand, S.; Khan, A.; Pujol, M.V.; LaRussa, V.F.; et al. Allogeneic human mesenchymal stem cells in patients with idiopathic pulmonary fibrosis via intravenous delivery (AETHER): A phase I safety clinical trial. Chest 2017, 151, 971–981. [Google Scholar] [CrossRef]
- Florea, V.; Rieger, A.C.; DiFede, D.L.; El-Khorazaty, J.; Natsumeda, M.; Banerjee, M.N.; Tompkins, B.A.; Khan, A.; Schulman, I.H.; Landin, A.M.; et al. Dose comparison study of allogeneic mesenchymal stem cells in patients with ischemic cardiomyopathy (the TRIDENT study). Circ. Res. 2017, 121, 1279–1290. [Google Scholar] [CrossRef]
- Hare, J.M.; DiFede, D.L.; Rieger, A.C.; Florea, V.; Landin, A.M.; El-Khorazaty, J.; Khan, A.; Mushtaq, M.; Lowery, M.H.; Byrnes, J.J.; et al. Randomized comparison of allogeneic versus autologous mesenchymal stem cells for nonischemic dilated cardiomyopathy: POSEIDON-DCM trial. J. Am. Coll. Cardiol. 2017, 69, 526–537. [Google Scholar] [CrossRef]
- Yamazoe, H.; Tanabe, T. Preparation of water-insoluble albumin film possessing nonadherent surface for cells and ligand binding ability. J. Biomed. Mater. Res. A 2008, 86, 228–234. [Google Scholar] [CrossRef]
- Verdanova, M.; Sauerova, P.; Hempel, U.; Kalbacova, M.H. Initial cell adhesion of three cell types in the presence and absence of serum proteins. Histochem. Cell Biol. 2017, 148, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E.; Pierschbacher, M.E. New perspectives in cell adhesion: RGD and integrins. Science 1987, 238, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lin, F. Mesenchymal stem cells are injured by complement after their contact with serum. Blood 2012, 120, 3436–3443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Jiang, W.; Chen, L.; Xu, Z.; Zhang, Q.; Zhu, M.; Ye, P.; Li, H.; Yu, L.; Zhou, X.; et al. Evaluation of the safety and efficacy of using human menstrual blood-derived mesenchymal stromal cells in treating severe and critically ill COVID-19 patients: An exploratory clinical trial. Clin. Transl. Med. 2021, 11, e297. [Google Scholar] [CrossRef]
- Levy, O.; Kuai, R.; Siren, E.M.J.; Bhere, J.; Milton, Y.; Nissar, N.; Biasio, M.D.; Heinelt, M.; Reeve, B.; Abdi, R.; et al. Shattering barriers toward clinically meaningful MSC therapies. Sci. Adv. 2020, 6, eaba6884. [Google Scholar] [CrossRef] [PubMed]
- Espina, M.; Jülke, H.; Brehm, W.; Ribitsch, I.; Winter, K.; Delling, U. Evaluation of transport conditions for autologous bone marrow-derived mesenchymal stromal cells for therapeutic application in horses. PeerJ 2016, 4, e1773. [Google Scholar] [CrossRef] [Green Version]
- Teshima, T.; Matsumoto, H.; Michishita, M.; Matsuoka, A.; Shiba, M.; Nagashima, T.; Koyama, H. Allogenic adipose tissue-derived mesenchymal stem cells ameliorate acute hepatic injury in dogs. Stem Cells Int. 2017, 2017, 3892514. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yasumura, Y.; Teshima, T.; Taira, Y.; Saito, T.; Yuchi, Y.; Suzuki, R.; Matsumoto, H. Optimal Intravenous Administration Procedure for Efficient Delivery of Canine Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 14681. https://doi.org/10.3390/ijms232314681
Yasumura Y, Teshima T, Taira Y, Saito T, Yuchi Y, Suzuki R, Matsumoto H. Optimal Intravenous Administration Procedure for Efficient Delivery of Canine Adipose-Derived Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2022; 23(23):14681. https://doi.org/10.3390/ijms232314681
Chicago/Turabian StyleYasumura, Yuyo, Takahiro Teshima, Yoshiaki Taira, Takahiro Saito, Yunosuke Yuchi, Ryohei Suzuki, and Hirotaka Matsumoto. 2022. "Optimal Intravenous Administration Procedure for Efficient Delivery of Canine Adipose-Derived Mesenchymal Stem Cells" International Journal of Molecular Sciences 23, no. 23: 14681. https://doi.org/10.3390/ijms232314681