
Restraint Stress and Repeated Corticosterone Administration Differentially Affect Neuronal Excitability, Synaptic Transmission and 5-HT7 Receptor Reactivity in the Dorsal Raphe Nucleus of Young Adult Male Rats

Abstract
:1. Introduction
2. Results
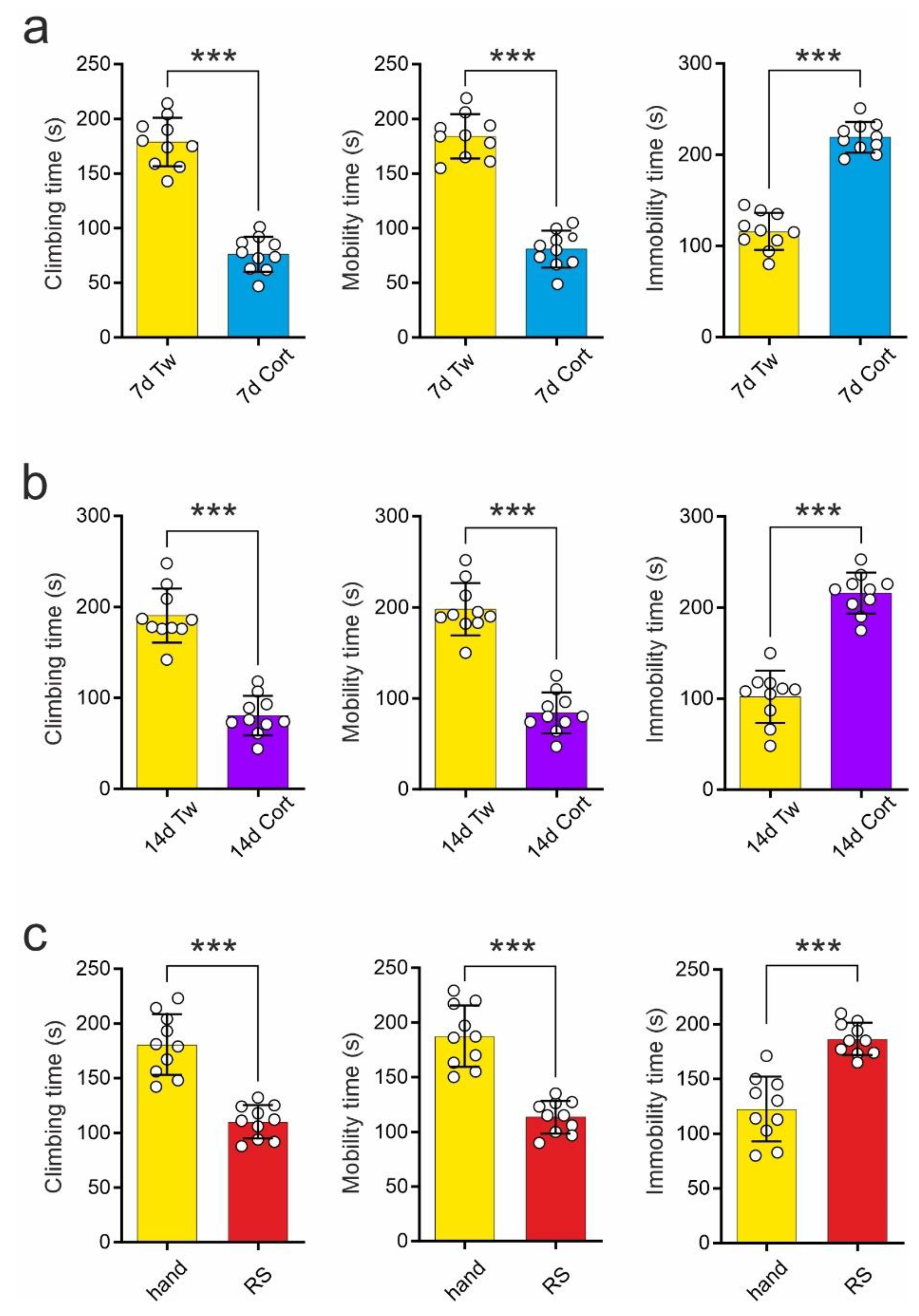
2.1. Comparison of the Effects of Repeated Corticosterone Administration and Repeated Restraint on the Forced Swim Test
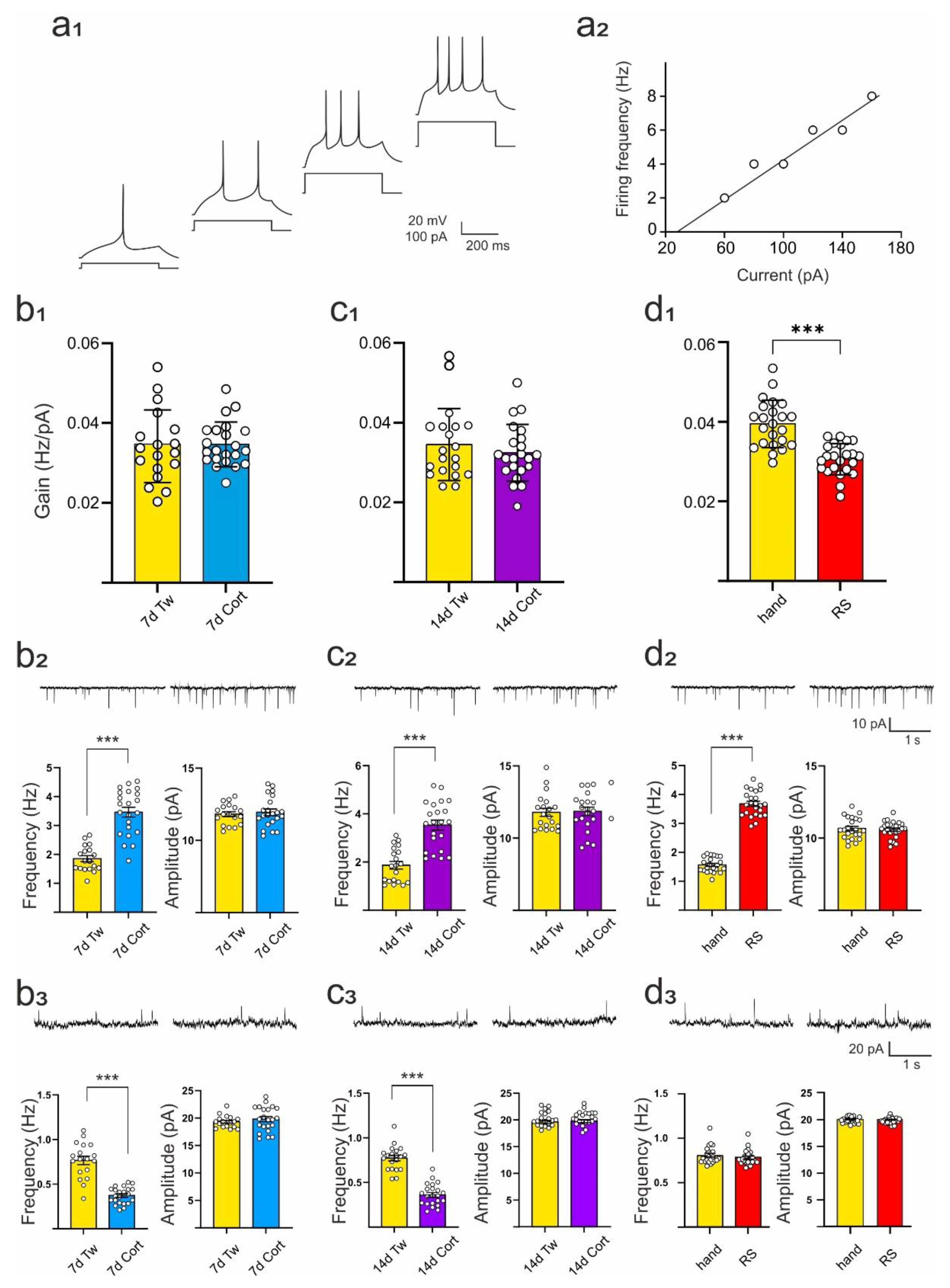
2.2. Comparison of the Effects of Repeated Corticosterone Administration and Repeated Restraint on Intrinsic Excitability of DRN Neurons and Their Synaptic Inputs
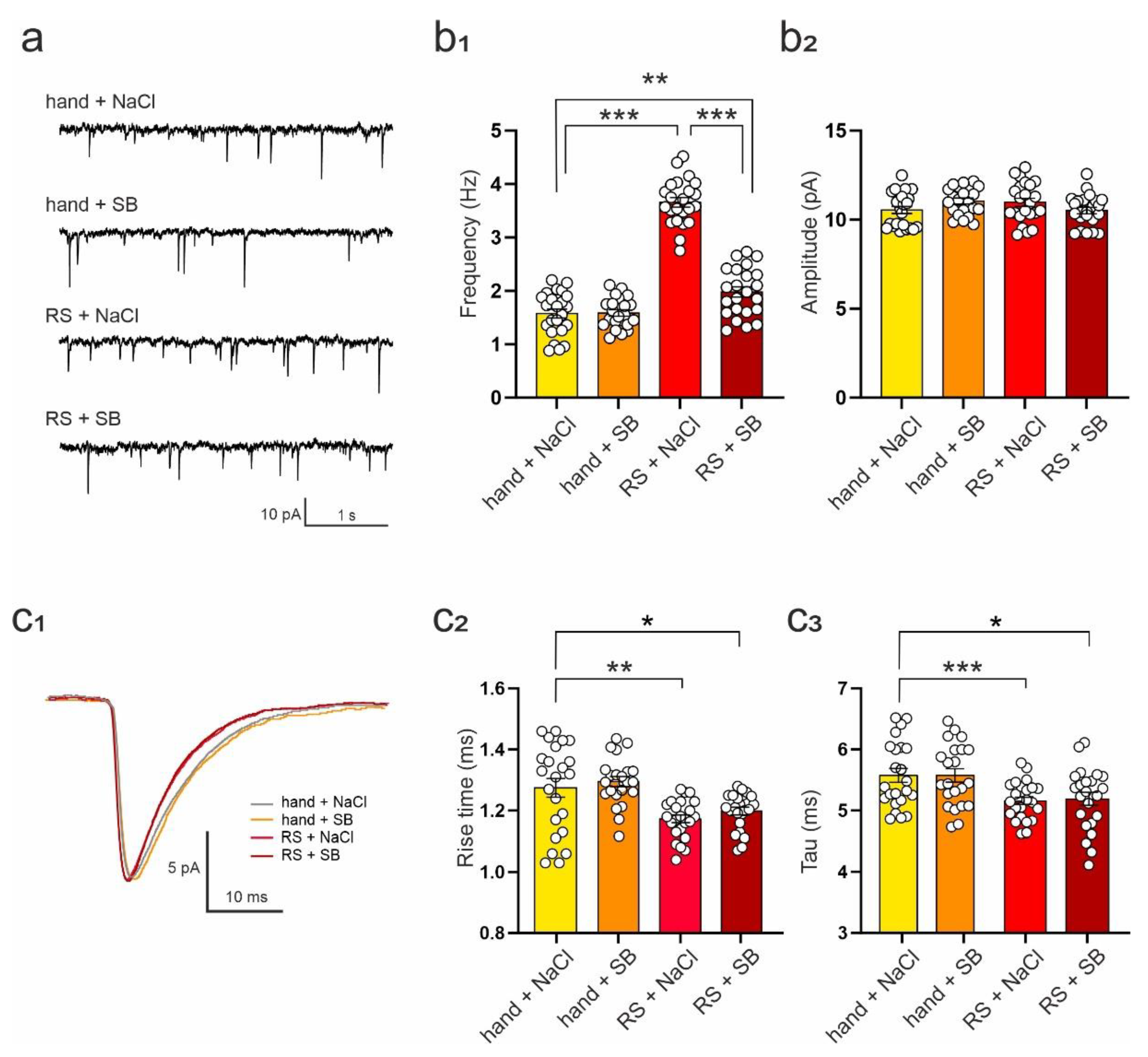
2.3. SB 269970 Attenuates the Effects of Repeated Restraint on the Excitability of DRN Neurons and Spontaneous EPSCs
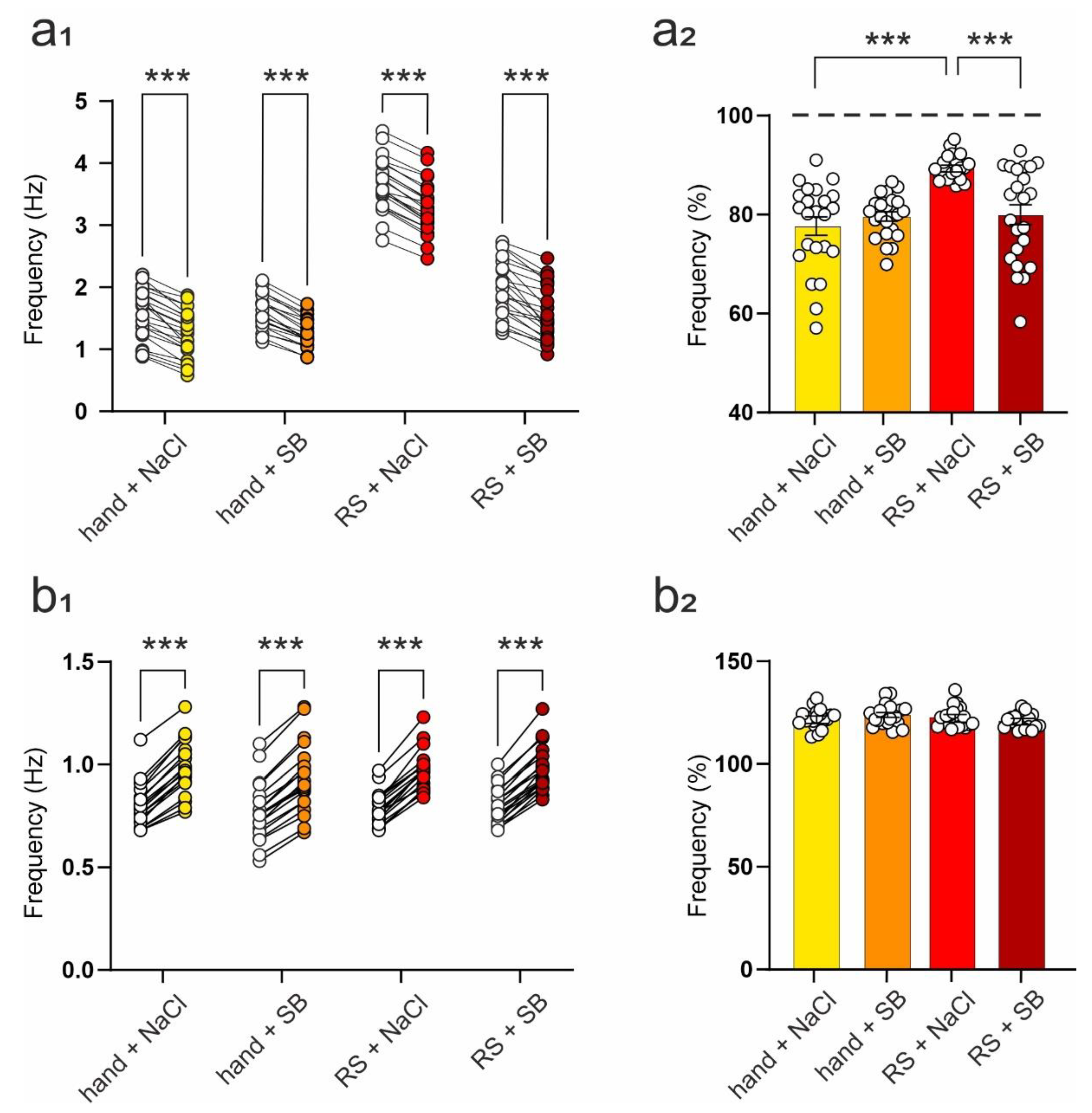
2.4. SB 269970 Counteracts the Effects of Repeated Restraint on 5-HT7 Receptor-Mediated Changes in the Frequency of Spontaneous EPSCs
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Experiment 1—The Effects of Corticosterone Administration and Restraint Stress on Performance on the Forced Swim Test and Electrophysiology of DRN Neurons
4.2.1. Treatment
4.2.2. Slice Preparation and Whole-Cell Recording
4.3. Experiment 2—Attenuation of the Effects of Repeated Restraint on the Excitability of DRN Neurons and Spontaneous EPSCs by a 5HT7R Antagonist
4.3.1. Treatment
4.3.2. Slice Preparation and Whole-Cell Recording
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reagan, L.P.; Grillo, C.A.; Piroli, G.G. The As and Ds of stress: Metabolic, morphological and behavioral consequences. Eur. J. Pharmacol. 2008, 585, 64–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammen, C.; Kim, E.Y.; Eberhart, N.K.; Brennan, P.A. Chronic and acute stress and the prediction of major depression in women. Depress. Anxiety 2009, 26, 718–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafet, G.E.; Nemeroff, C.B. The links between stress and depression: Psychoneuroendocrinological, genetic, and environmental interactions. J. Neuropsychiatry Clin. Neurosci. 2016, 28, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaouloff, F. Serotonin, stress and corticoids. J. Psychopharmacol. 2000, 14, 139–151. [Google Scholar] [CrossRef]
- Murnane, K.S. Serotonin 2A receptors are a stress response system: Implications for post-traumatic stress disorder. Behav. Pharmacol. 2019, 30, 151–162. [Google Scholar] [CrossRef]
- Lages, Y.V.M.; Rossi, A.D.; Krahe, T.E.; Landeira-Fernandez, J. Effect of chronic unpredictable mild stress on the expression profile of serotonin receptors in rats and mice: A meta-analysis. Neurosci. Biobehav. Rev. 2021, 124, 78–88. [Google Scholar] [CrossRef]
- Vincent, M.Y.; Jacobson, L. Glucocorticoid receptor deletion from the dorsal raphé nucleus of mice reduces dysphoria-like behavior and impairs hypothalamic-pituitary-adrenocortical axis feedback inhibition. Eur. J. Neurosci. 2014, 39, 1671–1681. [Google Scholar] [CrossRef] [Green Version]
- Lesiak, A.J.; Coffey, K.; Cohen, J.H.; Liang, K.J.; Chavkin, C.; Neumaier, J.F. Sequencing the serotonergic neuron translatome reveals a new role for Fkbp5 in stress. Mol. Psychiatry 2021, 26, 4742–4753. [Google Scholar] [CrossRef]
- Soiza-Reilly, M.; Commons, K.G. Glutamatergic drive of the dorsal raphe nucleus. J. Chem. Neuroanat. 2011, 41, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Liu, M.-Z.; Li, Q.; Deng, J.; Mu, D.; Sun, Y.-G. Organization of functional long-range circuits controlling the activity of serotonergic neurons in the dorsal raphe nucleus. Cell Rep. 2017, 18, 3018–3032. [Google Scholar] [CrossRef]
- Liu, R.; Jolas, T.; Aghajanian, G. Serotonin 5-HT(2) receptors activate local GABA inhibitory inputs to serotonergic neurons of the dorsal raphe nucleus. Brain Res. 2000, 873, 34–45. [Google Scholar] [CrossRef]
- Kusek, M.; Sowa, J.; Kamińska, K.; Gołembiowska, K.; Tokarski, K.; Hess, G. 5-HT7 receptor modulates GABAergic transmission in the rat dorsal raphe nucleus and controls cortical release of serotonin. Front. Cell Neurosci. 2015, 9, 324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soiza-Reilly, M.; Commons, K.G. Unraveling the architecture of the dorsal raphe synaptic neuropil using high-resolution neuroanatomy. Front. Neural. Circuits 2014, 8, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterner, E.Y.; Kalynchuk, L.E. Behavioral and neurobiological consequences of prolonged glucocorticoid exposure in rats: Relevance to depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Sowa, J.; Kusek, M.; Siwiec, M.; Sowa, J.E.; Bobula, B.; Tokarski, K.; Hess, G. The 5-HT7 receptor antagonist SB 269970 ameliorates corticosterone-induced alterations in 5-HT7 receptor-mediated modulation of GABAergic transmission in the rat dorsal raphe nucleus. Psychopharmacology 2018, 235, 3381–3390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sowa, J.; Kusek, M.; Bobula, B.; Hess, G.; Tokarski, K. Ketamine Administration Reverses Corticosterone-Induced Alterations in Excitatory and Inhibitory Transmission in the Rat Dorsal Raphe Nucleus. Neural Plast. 2019, 2019, 3219490. [Google Scholar] [CrossRef] [Green Version]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Herbert, J. Regional changes in c-fos expression in the basal forebrain and brainstem during adaptation to repeated stress: Correlations with cardiovascular, hypothermic and endocrine responses. Neuroscience 1995, 64, 675–685. [Google Scholar] [CrossRef]
- Gadek-Michalska, A.; Spyrka, J.; Rachwalska, P.; Tadeusz, J.; Bugajski, J. Influence of chronic stress on brain corticosteroid receptors and HPA axis activity. Pharmacol. Rep. 2013, 65, 1163–1175. [Google Scholar] [CrossRef]
- Sadler, A.M.; Bailey, S.J. Repeated daily restraint stress induces adaptive behavioural changes in both adult and juvenile mice. Physiol. Behav. 2016, 167, 313–323. [Google Scholar] [CrossRef]
- Shoji, H.; Miyakawa, T. Differential effects of stress exposure via two types of restraint apparatuses on behavior and plasma corticosterone level in inbred male BALB/cAJcl mice. Neuropsychopharmacol. Rep. 2020, 40, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Kusek, M.; Tokarski, K.; Hess, G. Repeated restraint stress enhances glutamatergic transmission in the paraventricular nucleus of the rat hypothalamus. J. Physiol. Pharmacol. 2013, 64, 565–570. [Google Scholar] [PubMed]
- Kusek, M.; Tokarska, A.; Siwiec, M.; Gadek-Michalska, A.; Szewczyk, B.; Hess, G.; Tokarski, K. Nitric Oxide Synthase Inhibitor Attenuates the Effects of Repeated Restraint Stress on Synaptic Transmission in the Paraventricular Nucleus of the Rat Hypothalamus. Front. Cell. Neurosci. 2017, 11, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarski, K.; Bobula, B.; Kusek, M.; Hess, G. The 5-HT(7) receptor antagonist SB 269970 counteracts restraint stress-induced attenuation of long-term potentiation in rat frontal cortex. J. Physiol. Pharmacol. 2011, 62, 663–667. [Google Scholar] [PubMed]
- Roberts, C.; Thomas, D.R.; Bate, S.T.; Kew, J.N. GABAergic modulation of 5-HT7 receptor-mediated effects on 5-HT efflux in the guinea-pig dorsal raphe nucleus. Neuropharmacology 2004, 46, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Lussier, A.L.; Lebedeva, K.; Fenton, E.Y.; Guskjolen, A.; Caruncho, H.J.; Kalynchuk, L.E. The progressive development of depression like behavior in corticosterone-treated rats is paralleled by slowed granule cell maturation and decreased reelin expression in the adult dentate gyrus. Neuropharmacology 2013, 71, 174–183. [Google Scholar] [CrossRef]
- Hill, M.N.; Brotto, L.A.; Lee, T.T.Y.; Gorzalka, B.B. Corticosterone attenuates the antidepressant-like effects elicited by melatonin in the forced swim test in both male and female rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2003, 27, 905–911. [Google Scholar] [CrossRef]
- Gregus, A.; Wintink, A.J.; Davis, A.C.; Kalynchuk, L.E. Effect of repeated corticosterone injections and restraint stress on anxiety and depression-like behavior in male rats. Behav. Brain Res. 2005, 156, 105–114. [Google Scholar] [CrossRef]
- Stamp, J.A.; Herbert, J. Multiple immediate-early gene expression during physiological and endocrine adaptation to repeated stress. Neuroscience 1999, 94, 1313–1322. [Google Scholar] [CrossRef]
- Viau, V.; Sawchenko, P.E. Hypophysiotropic neurons of the paraventricular nucleus respond in spatially, temporally, and phenotypically differentiated manners to acute vs. repeated restraint stress: Rapid publication. J. Comp. Neurol. 2002, 445, 293–307. [Google Scholar] [CrossRef]
- Grissom, N.; Bhatnagar, S. Habituation to repeated stress: Get used to it. Neurobiol. Learn. Mem. 2009, 92, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippe, T.J.; Bao, L.; Koblanski, M.E.; Viau, V. Sex differences in serotonin 5-HT 1A receptor responses to repeated restraint stress in adult male and female rats. Int. J. Neuropsychopharmacol. 2022, 25, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Nakajima, N.; Okada, S.; Yuri, K. Effects of aging on stress-related responses of serotonergic neurons in the dorsal raphe nucleus of male rats. Neurobiol. Stress 2016, 3, 43–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prouty, E.W.; Chandler, D.J.; Gao, W.-J.; Waterhouse, B.D. Selective vulnerability of dorsal raphe-medial prefrontal cortex projection neurons to corticosterone-induced hypofunction. Eur. J. Neurosci. 2019, 50, 1712–1726. [Google Scholar] [CrossRef] [PubMed]
- Matovic, S.; Ichiyama, A.; Igarashi, H.; Salter, E.W.; Sunstrum, J.K.; Wang, X.F.; Henry, M.; Kuebler, E.S.; Vernoux, N.; Martinez-Trujillo, J.; et al. Neuronal hypertrophy dampens neuronal intrinsic excitability and stress responsiveness during chronic stress. J. Physiol. 2020, 598, 2757–2773. [Google Scholar] [CrossRef]
- Goel, N.; Philippe, T.J.; Chang, J.; Koblanski, M.E.; Viau, V. Cellular and serotonergic correlates of habituated neuroendocrine responses in male and female rats. Psychoneuroendocrinology 2022, 136, 105599. [Google Scholar] [CrossRef]
- Ha, G.E.; Cheong, E. Chronic Restraint Stress Decreases the Excitability of Hypothalamic POMC Neuron and Increases Food Intake. Exp. Neurobiol. 2021, 30, 375–386. [Google Scholar] [CrossRef]
- Senst, L.; Baimoukhametova, D.; Sterley, T.-L.; Bains, J.S. Sexually dimorphic neuronal responses to social isolation. eLife 2016, 5, e18726. [Google Scholar] [CrossRef]
- Kula, J.; Gugula, A.; Blasiak, A.; Bobula, B.; Danielewicz, J.; Kania, A.; Tylko, G.; Hess, G. Diverse action of repeated corticosterone treatment on synaptic transmission, neuronal plasticity, and morphology in superficial and deep layers of the rat motor cortex. Pflug. Arch. Eur. J. Physiol. 2017, 469, 1519–1532. [Google Scholar] [CrossRef] [Green Version]
- Haj-Dahmane, S.; Shen, R.-Y. Chronic stress impairs α1-adrenoceptor-induced endocannabinoid-dependent synaptic plasticity in the dorsal raphe nucleus. J. Neurosci. 2014, 34, 14560–14570. [Google Scholar] [CrossRef]
- Marty, V.; Kuzmiski, J.B.; Baimoukhametova, D.V.; Bains, J.S. Short-term plasticity impacts information transfer at glutamate synapses onto parvocellular neuroendocrine cells in the paraventricular nucleus of the hypothalamus. J. Physiol. 2011, 589, 4259–4270. [Google Scholar] [CrossRef] [PubMed]
- Kirby, L.G.; Pan, Y.-Z.; Freeman-Daniels, E.; Rani, S.; Nunan, J.D.; Akanwa, A.; Beck, S.G. Cellular effects of swim stress in the dorsal raphe nucleus. Psychoneuroendocrinology 2007, 32, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarski, K.; Zahorodna, A.; Bobula, B.; Hess, G. 5-HT7 receptors increase the excitability of rat hippocampal CA1 pyramidal neurons. Brain Res. 2003, 993, 230–234. [Google Scholar] [CrossRef] [PubMed]
- Siwiec, M.; Kusek, M.; Sowa, J.E.; Tokarski, K.; Hess, G. 5-HT7 receptors increase the excitability of hippocampal CA1 pyramidal neurons by inhibiting the A-type potassium current. Neuropharmacology 2020, 177, 108248. [Google Scholar] [CrossRef]
- Soiza-Reilly, M.; Anderson, W.B.; Vaughan, C.W.; Commons, K.G. Presynaptic gating of excitation in the dorsal raphe nucleus by GABA. Proc. Natl. Acad. Sci. USA 2013, 110, 15800–15805. [Google Scholar] [CrossRef] [Green Version]
- Sowa, J.; Bobula, B.; Glombik, K.; Slusarczyk, J.; Basta-Kaim, A.; Hess, G. Prenatal stress enhances excitatory synaptic transmission and impairs long-term potentiation in the frontal cortex of adult offspring rats. PLoS ONE 2015, 10, e0119407. [Google Scholar] [CrossRef] [Green Version]
- Galindo-Charles, L.; Hernandez-Lopez, S.; Galarraga, E.; Tapia, D.; Bargas, J.; Garduño, J.; Frías-Dominguez, C.; Drucker-Colin, R.; Mihailescu, S. Serotoninergic dorsal raphe neurons possess functional postsynaptic nicotinic acetylcholine receptors. Synapse 2008, 62, 601–615. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Vm (mV) | Rm (MΩ) | Gain (Hz/pA) | n |
|---|---|---|---|---|
| 7d Tw | −68.78 ± 0.87 | 597.80 ± 22.33 | 0.0347 ± 0.0018 | 18 |
| 7d Cort | −69.36 ± 0.59 | 598.10 ± 16.76 | 0.0347 ± 0.0012 | 22 |
| 14d Tw | −67.10 ± 1.09 | 581.00 ± 15.79 | 0.0345 ± 0.0020 | 20 |
| 14d Cort | −67.36 ± 0.95 | 583.00 ± 13.24 | 0.0325 ± 0.0015 | 22 |
| hand | −65.91 ± 0.69 | 558.20 ± 11.30 | 0.0387 ± 0.0008 | 23 |
| RS | −65.82 ± 0.64 | 545.50 ± 11.57 | 0.0293 ± 0.0007 *** | 24 |
| Group | Mean Frequency (Hz) | Mean Amplitude (pA) | Rise Time (ms) | Decay Time Constant (τ, ms) | n |
|---|---|---|---|---|---|
| 7d Tw | 1.85 ± 0.10 | 11.82 ± 0.17 | 1.057 ± 0.022 | 4.912 ± 0.184 | 18 |
| 7d Cort | 3.46 ± 0.17 *** | 11.95 ± 0.22 | 1.046 ± 0.027 | 4.931 ± 0.157 | 22 |
| 14d Tw | 1.87 ± 0.16 | 11.79 ± 0.27 | 1.053 ± 0.023 | 4.979 ± 0.208 | 20 |
| 14d Cort | 3.53 ± 0.21 *** | 11.85 ± 0.29 | 1.056 ± 0.036 | 4.962 ± 0.170 | 22 |
| hand | 1.56 ± 0.05 | 10.67 ± 0.15 | 1.221 ± 0.013 | 5.283 ± 0.093 | 23 |
| RS | 3.67 ± 0.09 *** | 10.59 ± 0.12 | 1.146 ± 0.012 *** | 5.050 ± 0.045 * | 24 |
| Group | Mean Frequency (Hz) | Mean Amplitude (pA) | Rise Time (ms) | Decay Time Constant (τ, ms) | n |
|---|---|---|---|---|---|
| 7d Tw | 0.77 ± 0.05 | 19.33 ± 0.26 | 1.894 ± 0.092 | 6.967 ± 0.171 | 18 |
| 7d Cort | 0.37 ± 0.02 *** | 19.80 ± 0.48 | 1.873 ± 0.079 | 7.007 ± 0.159 | 22 |
| 14d Tw | 0.77 ± 0.03 | 19.61 ± 0.30 | 1.878 ± 0.078 | 6.993 ± 0.183 | 20 |
| 14d Cort | 0.36 ± 0.03 *** | 19.77 ± 0.27 | 1.810 ± 0.051 | 6.993 ± 0.174 | 22 |
| hand | 0.80 ± 0.02 | 19.82 ± 0.10 | 1.899 ± 0.018 | 7.738 ± 0.084 | 23 |
| RS | 0.78 ± 0.02 | 19.71 ± 0.09 | 1.924 ± 0.023 | 7.733 ± 0.086 | 24 |
| Group | Vm (mV) | Rm (MΩ) | Gain (Hz/pA) | n |
|---|---|---|---|---|
| hand + NaCl | −68.78 ± 0.87 | 597.80 ± 22.33 | 0.0347 ± 0.0018 | 23 |
| hand + SB | −65.05 ± 0.62 | 554.80 ± 20.92 | 0.0394 ± 0.0014 | 22 |
| RS + NaCl | −67.48 ± 0.96 | 545.30 ± 14.15 | 0.0309 ± 0.0008 *** | 23 |
| RS + SB | −67.35 ± 0.87 | 538.70 ± 16.80 | 0.0378 ± 0.0011 *** | 23 |
| Group | Mean Frequency (Hz) | Mean Amplitude (pA) | Rise Time (ms) | Decay Time Constant (τ, ms) | n |
|---|---|---|---|---|---|
| hand + NaCl | 1.58 ± 0.08 ** | 10.54 ± 0.20 | 1.274 ± 0.031 * | 5.576 ± 0.114 * | 23 |
| hand + SB | 1.59 ± 0.06 | 11.05 ± 0.17 | 1.295 ± 0.017 | 5.575 ± 0.101 | 22 |
| RS + NaCl | 3.66 ± 0.09 *** | 10.98 ± 0.23 | 1.172 ± 0.013 ** | 4.980 ± 0.066 *** | 23 |
| RS + SB | 1.98 ± 0.09 *** | 10.52 ± 0.18 | 1.198 ±0.013 | 5.190 ± 0.107 | 23 |
| Group | Mean Frequency (Hz) | Mean Amplitude (pA) | Rise Time (ms) | Decay Time Constant (τ, ms) | n |
|---|---|---|---|---|---|
| hand + NaCl | 0.80 ± 0.02 | 19.61 ± 0.17 | 1.908 ± 0.050 | 7.783 ± 0.166 | 19 |
| hand + SB | 0.77 ± 0.03 | 19.30 ± 0.24 | 1.944 ± 0.074 | 7.500 ± 0.126 | 19 |
| RS + NaCl | 0.79 ± 0.02 | 19.92 ± 0.14 | 1.944 ± 0.040 | 7.895 ± 0.158 | 19 |
| RS + SB | 0.80 ± 0.02 | 19.85 ± 0.13 | 1.933 ± 0.044 | 7.963 ± 0.128 | 20 |
| Group | Mean Frequency (Hz) | n | ||
|---|---|---|---|---|
| Before 5-CT | With 5-CT | % of Baseline | ||
| hand + NaCl | 1.58 ± 0.08 | 1.24 ± 0.08 *** | 77.68 ± 1.85 | 23 |
| hand + SB | 1.59 ± 0.06 | 1.26 ± 0.05 *** | 79.62 ± 0.94 | 22 |
| RS + NaCl | 3.66 ± 0.09 | 3.28 ± 0.09 *** | 89.44 ± 0.51 *** | 23 |
| RS + SB | 1.98 ± 0.09 | 1.59 ± 0.09 *** | 79.98 ± 2.02 *** | 23 |
| Group | Mean Frequency (Hz) | n | ||
|---|---|---|---|---|
| Before 5-CT | With 5-CT | % of Baseline | ||
| hand + NaCl | 0.80 ± 0.02 | 0.98 ± 0.03 *** | 122.20 ± 1.07 | 19 |
| hand + SB | 0.77 ± 0.03 | 0.93 ± 0.04 *** | 121.40 ± 0.83 | 19 |
| RS + NaCl | 0.79 ± 0.02 | 0.97 ± 0.02 *** | 123.90 ± 1.22 | 19 |
| RS + SB | 0.80 ± 0.02 | 0.98 ± 0.02 *** | 122.90 ± 1.11 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bąk, J.; Bobula, B.; Hess, G. Restraint Stress and Repeated Corticosterone Administration Differentially Affect Neuronal Excitability, Synaptic Transmission and 5-HT7 Receptor Reactivity in the Dorsal Raphe Nucleus of Young Adult Male Rats. Int. J. Mol. Sci. 2022, 23, 14303. https://doi.org/10.3390/ijms232214303
Bąk J, Bobula B, Hess G. Restraint Stress and Repeated Corticosterone Administration Differentially Affect Neuronal Excitability, Synaptic Transmission and 5-HT7 Receptor Reactivity in the Dorsal Raphe Nucleus of Young Adult Male Rats. International Journal of Molecular Sciences. 2022; 23(22):14303. https://doi.org/10.3390/ijms232214303
Chicago/Turabian StyleBąk, Joanna, Bartosz Bobula, and Grzegorz Hess. 2022. "Restraint Stress and Repeated Corticosterone Administration Differentially Affect Neuronal Excitability, Synaptic Transmission and 5-HT7 Receptor Reactivity in the Dorsal Raphe Nucleus of Young Adult Male Rats" International Journal of Molecular Sciences 23, no. 22: 14303. https://doi.org/10.3390/ijms232214303