
The Intersection of Human and Veterinary Medicine—A Possible Direction towards the Improvement of Cell Therapy Protocols in the Treatment of Perianal Fistulas
 ,
,  and
and 
Abstract
:1. Introduction
2. Local Cell Injections as Treatment of Fistulizing Crohn’s Disease
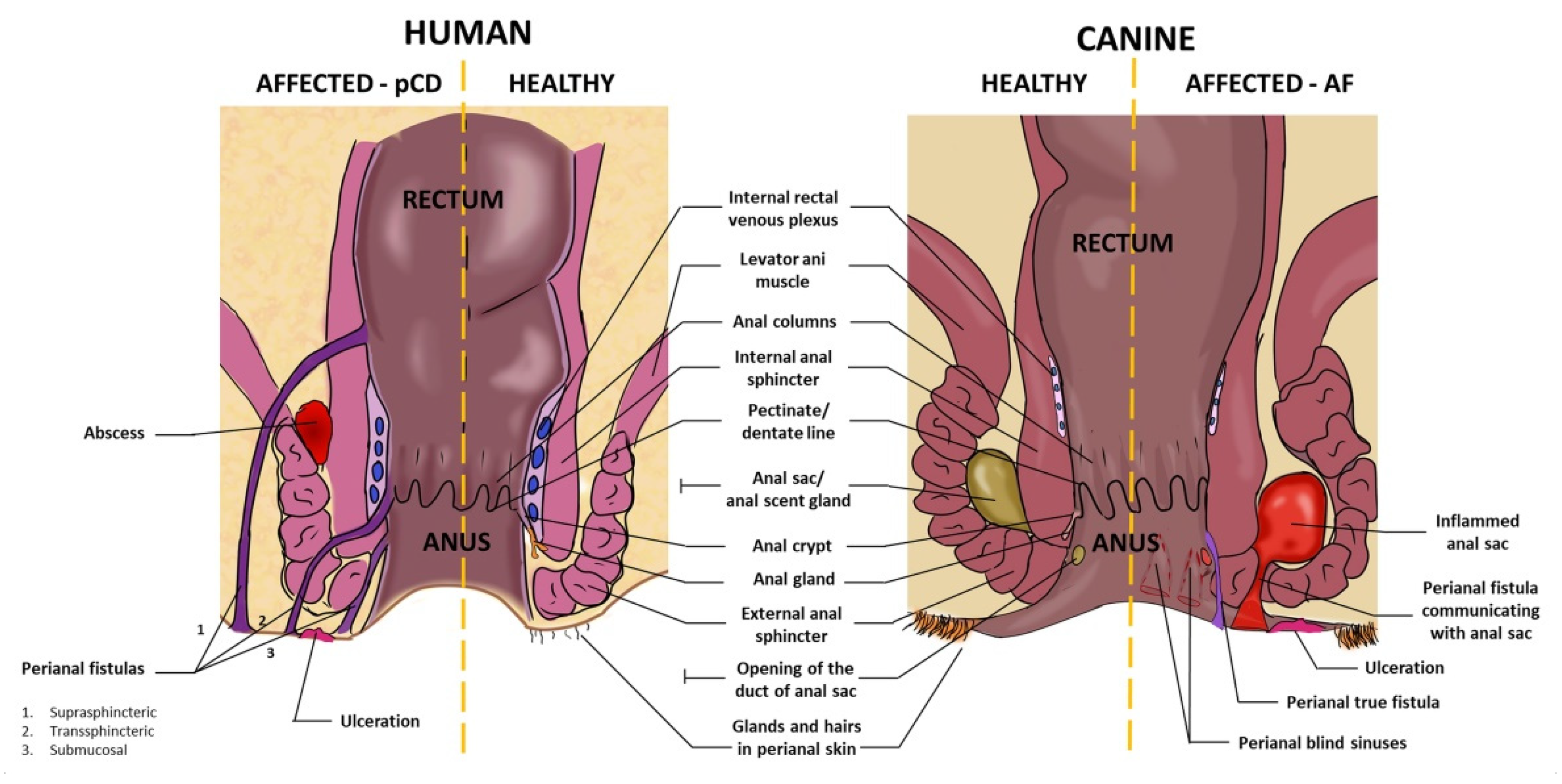
3. Anatomy of Canine and Human Perianal Region
4. Prevalence, Predispositions and Association with the Inflammation of the Intestine
5. Local Clinical Manifestations of Perianal CD in Humans and Anal Furunculosis in Dogs
6. Clinical Response to Pharmacological Treatment
7. Pathogenesis of Human Perianal CD and Canine AF
7.1. Intestinal Microbiota
7.2. Dysregulation of Immune Mechanisms
7.3. Genetic Factors
8. Human Mesenchymal Stem/Stromal Cells (hMSCs)—The Rationale behind Their Use in Fistulizing Crohn’s Disease—Potential Mechanisms of Action
9. Immunomodulatory Properties of Canine Mesenchymal Stem/Stromal Cells
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pogacnik, J.S.; Salgado, G. Perianal Crohn’s Disease. Clin. Colon Rectal Surg. 2019, 32, 377–385. [Google Scholar] [CrossRef]
- Gecse, K.B.; Bemelman, W.; Kamm, M.A.; Stoker, J.; Khanna, R.; Ng, S.C.; Panés, J.; van Assche, G.; Liu, Z.; Hart, A.; et al. A global consensus on the classification, diagnosis and multidisciplinary treatment of perianal fistulising Crohn’s disease. Gut 2014, 63, 1381–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adegbola, S.O.; Pisani, A.; Sahnan, K.; Tozer, P.; Ellul, P.; Warusavitarne, J. Medical and surgical management of perianal Crohn’s disease. Ann. Gastroenterol. 2018, 31, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Panes, J.; Reinisch, W.; Rupniewska, E.; Khan, S.; Forns, J.; Khalid, J.M.; Bojic, D.; Patel, H. Burden and outcomes for complex perianal fistulas in Crohn’s disease: Systematic review. World J. Gastroenterol. 2018, 24, 4821–4834. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Su, Q.; Zhang, B.; Shen, F.; Li, S. Efficacy of stem cells therapy for Crohn’s fistula: A meta-analysis and systematic review. Stem Cell Res. Ther. 2021, 12, 32. [Google Scholar] [CrossRef]
- Panés, J.; García-Olmo, D.; Van Assche, G.; Colombel, J.F.; Reinisch, W.; Baumgart, D.C.; Dignass, A.; Nachury, M.; Ferrante, M.; Kazemi-Shirazi, L.; et al. Expanded allogeneic adipose-derived mesenchymal stem cells (Cx601) for complex perianal fistulas in Crohn’s disease: A phase 3 randomised, double-blind controlled trial. Lancet 2016, 388, 1281–1290. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Klersy, C.; Leffler, D.A.; Rogers, R.; Bennett, D.; Corazza, G.R. Systematic review with meta-analysis: Safety and efficacy of local injections of mesenchymal stem cells in perianal fistulas. JGH Open 2019, 3, 249–260. [Google Scholar] [CrossRef]
- Johnson, S.; Hoch, J.S.; Halabi, W.J.; Ko, J.; Nolta, J.; Dave, M. Mesenchymal Stem/Stromal Cell Therapy Is More Cost-Effective Than Fecal Diversion for Treatment of Perianal Crohn’s Disease Fistulas. Front. Immunol. 2022, 13, 859954. [Google Scholar] [CrossRef]
- Lu, S.; Zhu, K.; Guo, Y.; Wang, E.; Huang, J. Evaluation of animal models of Crohn’s disease with anal fistula (Review). Exp. Ther. Med. 2021, 22, 974. [Google Scholar] [CrossRef]
- Ryska, O.; Serclova, Z.; Mestak, O.; Matouskova, E.; Vesely, P.; Mrazova, I. Local application of adipose-derived mesenchymal stem cells supports the healing of fistula: Prospective randomised study on rat model of fistulising Crohn’s disease. Scand. J. Gastroenterol. 2017, 52, 543–550. [Google Scholar] [CrossRef]
- Rivera-Nieves, J.; Bamias, G.; Vidrich, A.; Marini, M.; Pizarro, T.T.; McDuffie, M.J.; Moskaluk, C.A.; Cohn, S.M.; Cominelli, F. Emergence of perianal fistulizing disease in the SAMP1/YitFc mouse, a spontaneous model of chronic ileitis. Gastroenterology 2003, 124, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Flacs, M.; Collard, M.; Doblas, S.; Zappa, M.; Cazals-Hatem, D.; Maggiori, L.; Panis, Y.; Treton, X.; Ogier-Denis, E. Preclinical Model of Perianal Fistulizing Crohn’s Disease. Inflamm. Bowel Dis. 2020, 26, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, R.S.; Nissim-Eliraz, E.; Marsiano, N.; Nir, E.; Shemesh, H.; Leutenegger, M.; Gottier, C.; Lang, S.; Spalinger, M.R.; Leibl, S.; et al. Transplantation of Human Intestine Into the Mouse: A Novel Platform for Study of Inflammatory Enterocutaneous Fistulas. J. Crohn’s Colitis 2019, 13, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Galipeau, J.; Sensébé, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozawa, S.; Sagawa, F.; Endo, S.; De Almeida, G.M.; Mitsuishi, Y.; Sato, T.N. Predicting Human Clinical Outcomes Using Mouse Multi-Organ Transcriptome. iScience 2020, 23, 100791. [Google Scholar] [CrossRef] [Green Version]
- Kol, A.; Arzi, B.; Athanasiou, K.A.; Farmer, D.L.; Nolta, J.A.; Rebhun, R.B.; Chen, X.; Griffiths, L.G.; Verstraete, F.J.M.; Murphy, C.J.; et al. Companion animals: Translational scientist’s new best friends. Sci. Transl. Med. 2015, 7, 308ps321. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.M.; Dow, S.W. Concise Review: Stem Cell Trials Using Companion Animal Disease Models. Stem Cells 2016, 34, 1709–1729. [Google Scholar] [CrossRef]
- Ferrer, L.; Kimbrel, E.A.; Lam, A.; Falk, E.B.; Zewe, C.; Juopperi, T.; Lanza, R.; Hoffman, A. Treatment of perianal fistulas with human embryonic stem cell-derived mesenchymal stem cells: A canine model of human fistulizing Crohn’s disease. Regen. Med. 2016, 11, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Arzi, B.; Webb, T.L.; Koch, T.G.; Volk, S.W.; Betts, D.H.; Watts, A.; Goodrich, L.; Kallos, M.S.; Kol, A. Cell Therapy in Veterinary Medicine as a Proof-of-Concept for Human Therapies: Perspectives From the North American Veterinary Regenerative Medicine Association. Front. Vet. Sci. 2021, 8, 779109. [Google Scholar] [CrossRef]
- Cain, C.L. Canine Perianal Fistulas: Clinical Presentation, Pathogenesis, and Management. Vet. Clin. N. Am. Small Anim. Pract. 2019, 49, 53–65. [Google Scholar] [CrossRef]
- Hermanson, J.W.; De Lahunta, A. Miller and Evans’ Anatomy of the Dog; American Veterinary Medical Association: Schaumburg, IL, USA, 2020. [Google Scholar]
- Cohen, R.; Windsor, A. Anus: Surgical Treatment and Pathology; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Seow-Choen, F.; Ho, J.M.S. Histoanatomy of anal glands. Dis. Colon Rectum 1994, 37, 1215–1218. [Google Scholar] [CrossRef] [PubMed]
- McColl, I. The comparative anatomy and pathology of anal glands. Arris and Gale lecture delivered at the Royal College of Surgeons of England on 25th February 1965. Ann. R. Coll. Surg. Engl. 1967, 40, 36–67. [Google Scholar] [PubMed]
- Shabadash, S.A.; Zelikina, T.I. [Once more about hepatoid circumanal glands of dogs. History of their discovery and reasons for revision the structural and functional data]. Izv. Akad. Nauk. Seriia Biol. 2002, 2, 176–185. [Google Scholar]
- Janssenswillen, S.; Roelants, K.; Carpentier, S.; de Rooster, H.; Metzemaekers, M.; Vanschoenwinkel, B.; Proost, P.; Bossuyt, F. Odorant-binding proteins in canine anal sac glands indicate an evolutionarily conserved role in mammalian chemical communication. BMC Ecol. Evol. 2021, 21, 182. [Google Scholar] [CrossRef] [PubMed]
- Budsberg, S.C.; Spurgeon, T.L.; Liggitt, H.D. Anatomic predisposition to perianal fistulae formation in the German shepherd dog. Am. J. Vet. Res. 1985, 46, 1468–1472. [Google Scholar]
- GBD 2017 Inflammatory Bowel Disease Collaborators. The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Killingsworth, C.R.; Walshaw, R.; Dunstan, R.W.; Rosser, E.J., Jr. Bacterial population and histologic changes in dogs with perianal fistula. Am. J. Vet. Res. 1988, 49, 1736–1741. [Google Scholar]
- Jamieson, P.M.; Simpson, J.W.; Kirbyand, B.M.; Else, R.W. Association between anal furunculosis and colitis in the dog: Preliminary observations. J. Small Anim. Pract. 2002, 43, 109–114. [Google Scholar] [CrossRef]
- House, A.; Gregory, S.P.; Catchpole, B. Expression of cytokine mRNA in canine anal furunculosis lesions. Vet. Rec. 2003, 153, 354–358. [Google Scholar] [CrossRef]
- Day, M.J.; Weaver, B.M.Q. Pathology of surgically resected tissue from 305 cases of anal furunculosis in the dog. J. Small Anim. Pract. 1992, 33, 583–589. [Google Scholar] [CrossRef]
- Kaur, M.; Panikkath, D.; Yan, X.; Liu, Z.; Berel, D.; Li, D.; Vasiliauskas, E.A.; Ippoliti, A.; Dubinsky, M.; Shih, D.Q.; et al. Perianal Crohnʼs Disease is Associated with Distal Colonic Disease, Stricturing Disease Behavior, IBD-Associated Serologies and Genetic Variation in the JAK-STAT Pathway. Inflamm. Bowel Dis. 2016, 22, 862–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, L.; McCurdy, J.D.; Ma, C.; Jairath, V.; Singh, S. Epidemiology and Natural History of Perianal Crohn’s Disease: A Systematic Review and Meta-Analysis of Population-Based Cohorts. Inflamm. Bowel Dis. 2021, 28, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- de Zoeten, E.; Pasternak, B.A.; Mattei, P.; Kramer, R.E.; Kader, H. Diagnosis and treatment of perianal Crohn disease: NASPGHAN clinical report and consensus statement. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 401–412. [Google Scholar] [CrossRef] [PubMed]
- House, A.; Catchpole, B.; Gregory, S. Matrix metalloproteinase mRNA expression in canine anal furunculosis lesions. Vet. Immunol. Immunopathol. 2007, 115, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Panés, J.; Rimola, J. Perianal fistulizing Crohn’s disease: Pathogenesis, diagnosis and therapy. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 652–664. [Google Scholar] [CrossRef]
- Mathews, K.A.; Sukhiani, H.R. Randomized controlled trial of cyclosporine for treatment of perianal fistulas in dogs. J. Am. Vet. Med. Assoc. 1997, 211, 1249–1253. [Google Scholar]
- Dejaco, C.; Harrer, M.; Waldhoer, T.; Miehsler, W.; Vogelsang, H.; Reinisch, W. Antibiotics and azathioprine for the treatment of perianal fistulas in Crohn’s disease. Aliment. Pharmacol. Ther. 2003, 18, 1113–1120. [Google Scholar] [CrossRef]
- Harkin, K.R.; Phillips, D.; Wilkerson, M. Evaluation of Azathioprine on Lesion Severity and Lymphocyte Blastogenesis in Dogs With Perianal Fistulas. J. Am. Anim. Hosp. Assoc. 2007, 43, 21–26. [Google Scholar] [CrossRef]
- Shehab, M.; Alrashed, F.; Heron, V.; Restellini, S.; Bessissow, T. Comparative Efficacy of Biologic Therapies for Inducing Response and Remission in Fistulizing Crohn’s Disease: Systematic Review and Network Meta-Analysis of Randomized Controlled Trials. Inflamm. Bowel Dis. 2022. [Google Scholar] [CrossRef]
- Mahdi, B.M. Role of HLA typing on Crohn’s disease pathogenesis. Ann. Med. Surg. 2015, 4, 248–253. [Google Scholar] [CrossRef]
- Kennedy, L.J.; O’Neill, T.; House, A.; Barnes, A.; Kyöstilä, K.; Innes, J.; Fretwell, N.; Day, M.J.; Catchpole, B.; Lohi, H.; et al. Risk of anal furunculosis in German Shepherd dogs is associated with the major histocompatibility complex. Tissue Antigens 2008, 71, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Schnitzler, F.; Friedrich, M.; Wolf, C.; Stallhofer, J.; Angelberger, M.; Diegelmann, J.; Olszak, T.; Tillack, C.; Beigel, F.; Göke, B.; et al. The NOD2 Single Nucleotide Polymorphism rs72796353 (IVS4+10 A>C) Is a Predictor for Perianal Fistulas in Patients with Crohn’s Disease in the Absence of Other NOD2 Mutations. PLoS ONE 2015, 10, e0116044. [Google Scholar] [CrossRef] [PubMed]
- House, A.K.; Gregory, S.P.; Catchpole, B. Pattern-recognition receptor mRNA expression and function in canine monocyte/macrophages and relevance to canine anal furunuclosis. Vet. Immunol. Immunopathol. 2008, 124, 230–240. [Google Scholar] [CrossRef]
- Massey, J.; Short, A.D.; Catchpole, B.; House, A.; Day, M.J.; Lohi, H.; Ollier, W.E.R.; Kennedy, L.J. Genetics of canine anal furunculosis in the German shepherd dog. Immunogenetics 2014, 66, 311–324. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Maldonado-Contreras, A.; Ferrer, L.; Cawley, C.; Crain, S.; Bhattarai, S.; Toscano, J.; Ward, D.V.; Hoffman, A. Dysbiosis in a canine model of human fistulizing Crohn’s disease. Gut Microbes 2020, 12, 1785246. [Google Scholar] [CrossRef]
- Maggi, L.; Capone, M.; Giudici, F.; Santarlasci, V.; Querci, V.; Liotta, F.; Ficari, F.; Maggi, E.; Tonelli, F.; Annunziato, F.; et al. CD4+CD161+ T Lymphocytes Infiltrate Crohn’s Disease-Associated Perianal Fistulas and Are Reduced by Anti-TNF-α Local Therapy. Int. Arch. Allergy Immunol. 2013, 161, 81–86. [Google Scholar] [CrossRef]
- Altadill, A.; Eiró, N.; González, L.O.; Junquera, S.; González-Quintana, J.M.; Sánchez, M.R.; Andicoechea, A.; Saro, C.; Rodrigo, L.; Vizoso, F.J. Comparative analysis of the expression of metalloproteases and their inhibitors in resected crohnʼs disease and complicated diverticular disease. Inflamm. Bowel Dis. 2012, 18, 120–130. [Google Scholar] [CrossRef]
- Siegmund, B.; Feakins, R.M.; Barmias, G.; Ludvig, J.C.; Teixeira, F.V.; Rogler, G.; Scharl, M. Results of the Fifth Scientific Workshop of the ECCO (II): Pathophysiology of Perianal Fistulizing Disease. J. Crohn’s Colitis 2016, 10, 377–386. [Google Scholar] [CrossRef] [Green Version]
- Ellison, G.W. Treatment of perianal fistulas in dogs. J. Am. Vet. Med. Assoc. 1995, 206, 1680–1682. [Google Scholar]
- Mathews, K.A.; Ayres, S.A.; Tano, C.A.; Riley, S.M.; Sukhiani, H.R.; Adams, C. Cyclosporin treatment of perianal fistulas in dogs. Can. Vet. J. 1997, 38, 39–41. [Google Scholar] [PubMed]
- Hanauer, S.B.; Smith, M.B. Rapid closure of Crohn’s disease fistulas with continuous intravenous cyclosporin A. Am. J. Gastroenterol. 1993, 88, 646–649. [Google Scholar] [PubMed]
- Present, D.H.; Lichtiger, S. Efficacy of cyclosporine in treatment of fistula of crohn’s disease. Am. J. Dig. Dis. 1994, 39, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Hardie, R.J.; Gregory, S.P.; Tomlin, J.; Sturgeon, C.; Lipscomb, V.; Ladlow, J. Cyclosporine treatment of anal furunculosis in 26 dogs. J. Small Anim. Pract. 2005, 46, 3–9. [Google Scholar] [CrossRef] [PubMed]
- House, A.K.; Guitian, J.; Gregory, S.P.; Hardie, R.J. Evaluation of the Effect of Two Dose Rates of Cyclosporine on the Severity of Perianal Fistulae Lesions and Associated Clinical Signs in Dogs. Vet. Surg. 2006, 35, 543–549. [Google Scholar] [CrossRef]
- Doust, R.; Griffiths, L.G.; Sullivan, M. Evaluation of once daily treatment with cyclosporine for anal furunculosis in dogs. Vet. Rec. 2003, 152, 225–229. [Google Scholar] [CrossRef]
- Wetwittayakhlang, P.; Al Khoury, A.; Hahn, G.D.; Lakatos, P.L. The Optimal Management of Fistulizing Crohn’s Disease: Evidence beyond Randomized Clinical Trials. J. Clin. Med. 2022, 11, 3045. [Google Scholar] [CrossRef]
- Rovira, P.; Mascarell, L.; Bachi, P.T. The Impact of Immunosuppressive Drugs on the Analysis of T-Cell Activation. Curr. Med. Chem. 2000, 7, 673–692. [Google Scholar] [CrossRef]
- Archer, T.; Boothe, D.; Langston, V.; Fellman, C.; Lunsford, K.; Mackin, A. Oral Cyclosporine Treatment in Dogs: A Review of the Literature. J. Vet. Intern. Med. 2013, 28, 1–20. [Google Scholar] [CrossRef]
- Khan, K.J.; Ullman, T.A.; Ford, A.; Abreu, M.T.; Abadir, A.; Marshall, J.; Talley, N.J.; Moayyedi, P. Antibiotic Therapy in Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. Am. J. Gastroenterol. 2011, 106, 661–673. [Google Scholar] [CrossRef]
- West, R.L.; Van Der Woude, C.J.; Hansen, B.; Felt-Bersma, R.J.F.; Van Tilburg, A.J.P.; Drapers, J.A.G.; Kuipers, E.J. Clinical and endosonographic effect of ciprofloxacin on the treatment of perianal fistulae in Crohn’s disease with infliximab: A double-blind placebo-controlled study. Aliment. Pharmacol. Ther. 2004, 20, 1329–1336. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Ng, S.C.; Durdey, P.; Burt, C.; Torkington, J.; Rao, P.K.D.; Mayberry, J.; Moshkovska, T.; Stone, C.D.; Carapeti, E.; et al. Randomized clinical trial of metronidazole ointment versus placebo in perianal Crohn’s disease. Br. J. Surg. 2010, 97, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Thia, K.T.; Mahadevan, U.; Feagan, B.G.; Wong, C.; Cockeram, A.; Bitton, A.; Bernstein, C.N.; Sandborn, W.J. Ciprofloxacin or metronidazole for the treatment of perianal fistulas in patients with Crohnʼs disease: A randomized, double-blind, placebo-controlled pilot study. Inflamm. Bowel Dis. 2009, 15, 17–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.Z.; Chua, E.W. Revisiting the Role of Thiopurines in Inflammatory Bowel Disease Through Pharmacogenomics and Use of Novel Methods for Therapeutic Drug Monitoring. Front. Pharmacol. 2018, 9, 1107. [Google Scholar] [CrossRef]
- Tisdall, P.; Hunt, G.B.; Beck, J.A.; Malik, R. Management of perianal fistulae in five dogs using azathioprine and metronidazole prior to surgery. Aust. Vet. J. 1999, 77, 374–378. [Google Scholar] [CrossRef]
- Kalliolias, G.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef]
- Kobayashi, T.; Momoi, Y.; Iwasaki, T. Cyclosporine A Inhibits the mRNA Expressions of IL-2, IL-4 and IFN-.GAMMA., but not TNF-.ALPHA., in Canine Mononuclear Cells. J. Vet. Med. Sci. 2007, 69, 887–892. [Google Scholar] [CrossRef] [Green Version]
- Guan, Q. A Comprehensive Review and Update on the Pathogenesis of Inflammatory Bowel Disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [Green Version]
- Kopper, J.J.; Iennarella-Servantez, C.; Jergens, A.E.; Sahoo, D.K.; Guillot, E.; Bourgois-Mochel, A.; Martinez, M.N.; Allenspach, K.; Mochel, J.P. Harnessing the Biology of Canine Intestinal Organoids to Heighten Understanding of Inflammatory Bowel Disease Pathogenesis and Accelerate Drug Discovery: A One Health Approach. Front. Toxicol. 2021, 3, 773953. [Google Scholar] [CrossRef]
- Coelho, L.P.; Kultima, J.R.; Costea, P.I.; Fournier, C.; Pan, Y.; Czarnecki-Maulden, G.; Hayward, M.R.; Forslund, S.K.; Schmidt, T.S.B.; Descombes, P.; et al. Similarity of the dog and human gut microbiomes in gene content and response to diet. Microbiome 2018, 6, 72. [Google Scholar] [CrossRef]
- Day, M.J. Immunopathology of analfurunculosis in the dog. J. Small Anim. Pract. 1993, 34, 381–388. [Google Scholar] [CrossRef]
- Bataille, F.; Klebl, F.; Rümmele, P.; Schroeder, J.; Farkas, S.; Wild, P.-J.; Fürst, A.; Hofstädter, F.; Schölmerich, J.; Herfarth, H.; et al. Morphological characterisation of Crohn’s disease fistulae. Gut 2004, 53, 1314–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruffolo, C.; Scarpa, M.; Faggian, D.; Pozza, A.; Navaglia, F.; Dʼincà, R.; Hoxha, P.; Romanato, G.; Polese, L.; Sturniolo, G.C.; et al. Cytokine network in rectal mucosa in perianal Crohnʼs disease: Relations with inflammatory parameters and need for surgery. Inflamm. Bowel Dis. 2008, 14, 1406–1412. [Google Scholar] [CrossRef] [PubMed]
- Lauro, M.L.; Burch, J.M.; Grimes, C.L. The effect of NOD2 on the microbiota in Crohn’s disease. Curr. Opin. Biotechnol. 2016, 40, 97–102. [Google Scholar] [CrossRef] [Green Version]
- House, A.K.; Binns, M.M.; Gregory, S.P.; Catchpole, B. Analysis of NOD1, NOD2, TLR1, TLR2, TLR4, TLR5, TLR6 and TLR9 genes in anal furunculosis of German shepherd dogs. Tissue Antigens 2009, 73, 250–254. [Google Scholar] [CrossRef]
- O’Sullivan, S.; Gilmer, J.F.; Medina, C. Matrix Metalloproteinases in Inflammatory Bowel Disease: An Update. Mediat. Inflamm. 2015, 2015, 964131. [Google Scholar] [CrossRef]
- Kirkegaard, T.; Hansen, A.; Bruun, E.; Brynskov, J. Expression and localisation of matrix metalloproteinases and their natural inhibitors in fistulae of patients with Crohn’s disease. Gut 2004, 53, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Lavini-Ramos, C.; Silva, H.M.; Soares-Schanoski, A.; Monteiro, S.M.; Ferreira, L.R.P.; Pacanaro, A.P.; Gomes, S.; Batista, J.; Faé, K.; Kalil, J.; et al. MMP9 integrates multiple immunoregulatory pathways that discriminate high suppressive activity of human mesenchymal stem cells. Sci. Rep. 2017, 7, 874. [Google Scholar] [CrossRef] [Green Version]
- Gu, J.J.; Hoj, J.; Rouse, C.; Pendergast, A.M. Mesenchymal stem cells promote metastasis through activation of an ABL-MMP9 signaling axis in lung cancer cells. PLoS ONE 2020, 15, e0241423. [Google Scholar] [CrossRef]
- Barnes, A.; O’Neill, T.; Kennedy, L.J.; Short, A.D.; Catchpole, B.; House, A.; Binns, M.; Fretwell, N.; Day, M.J.; Ollier, W.E.R. Association of canine anal furunculosis with TNFA is secondary to linkage disequilibrium with DLA-DRB1*. Tissue Antigens 2009, 73, 218–224. [Google Scholar] [CrossRef]
- Hugot, J.-P.; Chamaillard, M.; Zouali, H.; Lesage, S.; Cézard, J.-P.; Belaiche, J.; Almer, S.; Tysk, C.; O’Morain, C.A.; Gassull, M.; et al. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn’s disease. Nature 2001, 411, 599–603. [Google Scholar] [CrossRef] [PubMed]
- Garza-Hernandez, D.; Sepulveda-Villegas, M.; Garcia-Pelaez, J.; Aguirre-Gamboa, R.; Lakatos, P.L.; Estrada, K.; Martinez-Vazquez, M.; Trevino, V. A systematic review and functional bioinformatics analysis of genes associated with Crohn’s disease identify more than 120 related genes. BMC Genom. 2022, 23, 302. [Google Scholar] [CrossRef] [PubMed]
- Franchimont, D.; Vermeire, S.; El Housni, H.; Pierik, M.; Van Steen, K.; Gustot, T.; Quertinmont, E.; Abramowicz, M.; Van Gossum, A.; Devière, J.; et al. Deficient host-bacteria interactions in inflammatory bowel disease? The toll-like receptor (TLR)-4 Asp299gly polymorphism is associated with Crohn’s disease and ulcerative colitis. Gut 2004, 53, 987–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Włodarczyk, M.; Czerwińska, K.; Włodarczyk, J.; Fichna, J.; Dziki, A.; Dziki, L. Current Overview on the Use of Mesenchymal Stem Cells for Perianal Fistula Treatment in Patients with Crohn’s Disease. Life 2021, 11, 1133. [Google Scholar] [CrossRef]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef]
- Jiang, W.; Xu, J. Immune modulation by mesenchymal stem cells. Cell Prolif. 2020, 53, e12712. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Jiang, J.; Gu, Z.; Zhang, J.; Chen, Y.; Liu, X. Mesenchymal stromal cell therapies: Immunomodulatory properties and clinical progress. Stem Cell Res. Ther. 2020, 11, 345. [Google Scholar] [CrossRef]
- Dabrowski, F.A.; Burdzinska, A.; Kulesza, A.; Chlebus, M.; Kaleta, B.; Borysowski, J.; Zolocinska, A.; Paczek, L.; Wielgos, M. Mesenchymal Stem Cells from Human Amniotic Membrane and Umbilical Cord Can Diminish Immunological Response in an in vitro Allograft Model. Gynecol. Obstet. Investig. 2016, 82, 267–275. [Google Scholar] [CrossRef]
- Le Blanc, K.; Tammik, L.; Sundberg, B.; Haynesworth, S.E.; Ringden, O. Mesenchymal Stem Cells Inhibit and Stimulate Mixed Lymphocyte Cultures and Mitogenic Responses Independently of the Major Histocompatibility Complex. Scand. J. Immunol. 2003, 57, 11–20. [Google Scholar] [CrossRef]
- Duffy, M.M.; Pindjakova, J.; Hanley, S.A.; McCarthy, C.; Weidhofer, G.A.; Sweeney, E.M.; English, K.; Shaw, G.; Murphy, J.M.; Barry, F.P.; et al. Mesenchymal stem cell inhibition of T-helper 17 cell- differentiation is triggered by cell-cell contact and mediated by prostaglandin E2 via the EP4 receptor. Eur. J. Immunol. 2011, 41, 2840–2851. [Google Scholar] [CrossRef]
- Luz-Crawford, P.; Kurte, M.; Bravo-Alegría, J.; Contreras, R.; Nova-Lamperti, E.; Tejedor, G.; Noël, D.; Jorgensen, C.; Figueroa, F.; Djouad, F.; et al. Mesenchymal stem cells generate a CD4+CD25+Foxp3+ regulatory T cell population during the differentiation process of Th1 and Th17 cells. Stem Cell Res. Ther. 2013, 4, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Hematti, P. Mesenchymal stem cell–educated macrophages: A novel type of alternatively activated macrophages. Exp. Hematol. 2009, 37, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Dymowska, M.; Aksamit, A.; Zielniok, K.; Kniotek, M.; Kaleta, B.; Roszczyk, A.; Zych, M.; Dabrowski, F.; Paczek, L.; Burdzinska, A. Interaction between Macrophages and Human Mesenchymal Stromal Cells Derived from Bone Marrow and Wharton’s Jelly—A Comparative Study. Pharmaceutics 2021, 13, 1822. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Hisamatsu, T.; Okamoto, S.; Chinen, H.; Kobayashi, T.; Sato, T.; Sakuraba, A.; Kitazume, M.T.; Sugita, A.; Koganei, K.; et al. Unique CD14+ intestinal macrophages contribute to the pathogenesis of Crohn disease via IL-23/IFN-γ axis. J. Clin. Investig. 2008, 118, 2269–2280. [Google Scholar] [CrossRef] [Green Version]
- Sakuraba, A.; Sato, T.; Kamada, N.; Kitazume, M.; Sugita, A.; Hibi, T. Th1/Th17 Immune Response Is Induced by Mesenteric Lymph Node Dendritic Cells in Crohn’s Disease. Gastroenterology 2009, 137, 1736–1745. [Google Scholar] [CrossRef]
- Li, Y.-P.; Paczesny, S.; Lauret, E.; Poirault, S.; Bordigoni, P.; Mekhloufi, F.; Hequet, O.; Bertrand, Y.; Ou-Yang, J.-P.; Stoltz, J.-F.; et al. Human Mesenchymal Stem Cells License Adult CD34+ Hemopoietic Progenitor Cells to Differentiate into Regulatory Dendritic Cells through Activation of the Notch Pathway. J. Immunol. 2008, 180, 1598–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuerquis, J.; Romieu-Mourez, R.; François, M.; Routy, J.-P.; Young, Y.K.; Zhao, J.; Eliopoulos, N. Human mesenchymal stromal cells transiently increase cytokine production by activated T cells before suppressing T-cell proliferation: Effect of interferon-γ and tumor necrosis factor-α stimulation. Cytotherapy 2014, 16, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hass, R. Role of MSC in the Tumor Microenvironment. Cancers 2020, 12, 2107. [Google Scholar] [CrossRef]
- Liang, W.; Chen, X.; Zhang, S.; Fang, J.; Chen, M.; Xu, Y.; Chen, X. Mesenchymal stem cells as a double-edged sword in tumor growth: Focusing on MSC-derived cytokines. Cell. Mol. Biol. Lett. 2021, 26, 3. [Google Scholar] [CrossRef]
- Scharl, M.; Frei, S.; Pesch, T.; Kellermeier, S.; Arikkat, J.; Frei, P.; Fried, M.; Weber, A.; Jehle, E.; Rühl, A.; et al. Interleukin-13 and transforming growth factor β synergise in the pathogenesis of human intestinal fistulae. Gut 2013, 62, 63–72. [Google Scholar] [CrossRef]
- Carrade, D.D.; Borjesson, D.L. Immunomodulation by mesenchymal stem cells in veterinary species. Comp. Med. 2013, 63, 207–217. [Google Scholar] [PubMed]
- Luo, H.; Li, D.; Chen, Z.; Wang, B.; Chen, S. Manufacturing and banking canine adipose-derived mesenchymal stem cells for veterinary clinical application. BMC Vet. Res. 2021, 17, 96. [Google Scholar] [CrossRef] [PubMed]
- Delfi, I.; Wood, C.; Johnson, L.; Snow, M.; Innes, J.; Myint, P.; Johnson, W. An In Vitro Comparison of the Neurotrophic and Angiogenic Activity of Human and Canine Adipose-Derived Mesenchymal Stem Cells (MSCs): Translating MSC-Based Therapies for Spinal Cord Injury. Biomolecules 2020, 10, 1301. [Google Scholar] [CrossRef] [PubMed]
- Teshima, T.; Yuchi, Y.; Suzuki, R.; Matsumoto, H.; Koyama, H. Immunomodulatory Effects of Canine Adipose Tissue Mesenchymal Stem Cell-Derived Extracellular Vesicles on Stimulated CD4+ T Cells Isolated from Peripheral Blood Mononuclear Cells. J. Immunol. Res. 2021, 2021, 2993043. [Google Scholar] [CrossRef] [PubMed]
- An, J.-H.; Li, Q.; Bhang, D.-H.; Song, W.-J.; Youn, H.-Y. TNF-α and INF-γ primed canine stem cell-derived extracellular vesicles alleviate experimental murine colitis. Sci. Rep. 2020, 10, 2115. [Google Scholar] [CrossRef]
- Shahsavari, A.; Weeratunga, P.; Ovchinnikov, D.A.; Whitworth, D.J. Pluripotency and immunomodulatory signatures of canine induced pluripotent stem cell-derived mesenchymal stromal cells are similar to harvested mesenchymal stromal cells. Sci. Rep. 2021, 11, 3486. [Google Scholar] [CrossRef]


{kind=link}
| Humans—Perianal Crohn’s Disease | Ref. | Dogs—Anal Furunculosis | Ref. | ||
|---|---|---|---|---|---|
| Symptoms | Association of perianal disease with bowel inflammation | Established. Only about 5% of patients with perianal CD do not have concurrent luminal form of CD | [1] | Probable. 39% of dogs with diagnosed AF do not present histological symptoms of colitis or endoscopic signs of proctitis | [30] |
| Perianal symptoms | Fissures, skin tags, ulcers, abscesses, true fistulas with two openings, anal strictures | [37] | Perianal blind sinuses, ulcerations, fistulas communicating with anal sac or anorectum (the incidence of true fistulas with two openings not established) | [20] | |
| Response to pharmacologic therapy | Antibiotics | Proven to reduce drainage of perianal fistulas, not to induce healing, recommended as adjunctive therapy | [37] | Efficacy not proven, sometimes used as adjunctive therapy | [20] |
| Cyclosporine (CsA) | Efficacy not proven—not recommended | [2] | Proven efficacy—recommended | [38] | |
| Thiopurine derivatives (i.e., azathioprin) | Moderate efficacy—recommended | [39] | Moderate efficacy based on scant data, sometimes used if CsA therapy is not possible (due to economic reasons or availability problems) | [40] | |
| Anti-TNF | Proven efficacy—recommended | [41] | Not tested, caninized anti-TNF not available | ||
| Pathogenesis | Genetic factors | The role of genes encoding MHC molecules (generally for CD)—confirmed | [42] | the role of genes encoding MHC molecules (DLA-DRB1)—confirmed | [43] |
| The role of SNPs in NOD2 (one of PPRs) (specifically in pCD) - confirmed | [44] | The role of SNPs in chosen PPRs (including NOD2) failed to be confirmed | [45] | ||
| The role of TNF pathway genes (specifically for pCD)—confirmed | [33] | The role of TNF pathway not confirmed | [46] | ||
| The role of other genes associated with cell adhesion, extracelullar matrix, scaffolding proteins or autophagy indicated (specifically for pCD) | [33] | 6 non-synonymous SNPs identified to be related to AF. The most significantly associated genes were ADAMTS16 and CTNND2 | [46] | ||
| Intestinal microbiota | Association of gut microbiota with pCD confirmed. Dichotomy of microbiome into dysbiotic and non-dysbiotic reported | [47] | Association of gut microbiota with AF confirmed. Dichotomy of microbiome into dysbiotic and non-dysbiotic reported | [48] | |
| Disturbances in T cell mediated immunity | Association of abnormal T cell response with pCD confirmed. Excessive Th1 response; Excessive Th17 response | [49] | Association of abnormal T cell response with AF confirmed. Excessive Th1 response; role of Th17 response not studied | [31] | |
| Metallo-proteinases (MMPs) | Elevated expression of MMP-2, MMP-9, and MMP-13 in pCD fistulas | [50] | Elevated expression of MMP9 and MMP13 but not MMP2 in perianal tissue of dogs with AF | [36] | |
| Epithelial-to-mesenchymal transition | The EMT process confirmed to be involved in the formation of CD-related fistulas | [51] | The association of EMT process and AF not studied |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burdzinska, A.; Galanty, M.; Więcek, S.; Dabrowski, F.A.; Lotfy, A.; Sadkowski, T. The Intersection of Human and Veterinary Medicine—A Possible Direction towards the Improvement of Cell Therapy Protocols in the Treatment of Perianal Fistulas. Int. J. Mol. Sci. 2022, 23, 13917. https://doi.org/10.3390/ijms232213917
Burdzinska A, Galanty M, Więcek S, Dabrowski FA, Lotfy A, Sadkowski T. The Intersection of Human and Veterinary Medicine—A Possible Direction towards the Improvement of Cell Therapy Protocols in the Treatment of Perianal Fistulas. International Journal of Molecular Sciences. 2022; 23(22):13917. https://doi.org/10.3390/ijms232213917
Chicago/Turabian StyleBurdzinska, Anna, Marek Galanty, Sabina Więcek, Filip A. Dabrowski, Ahmed Lotfy, and Tomasz Sadkowski. 2022. "The Intersection of Human and Veterinary Medicine—A Possible Direction towards the Improvement of Cell Therapy Protocols in the Treatment of Perianal Fistulas" International Journal of Molecular Sciences 23, no. 22: 13917. https://doi.org/10.3390/ijms232213917