
Minocycline Attenuates Lipopolysaccharide-Induced Locomotor Deficit and Anxiety-like Behavior and Related Expression of the BDNF/CREB Protein in the Rat Medial Prefrontal Cortex (mPFC)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
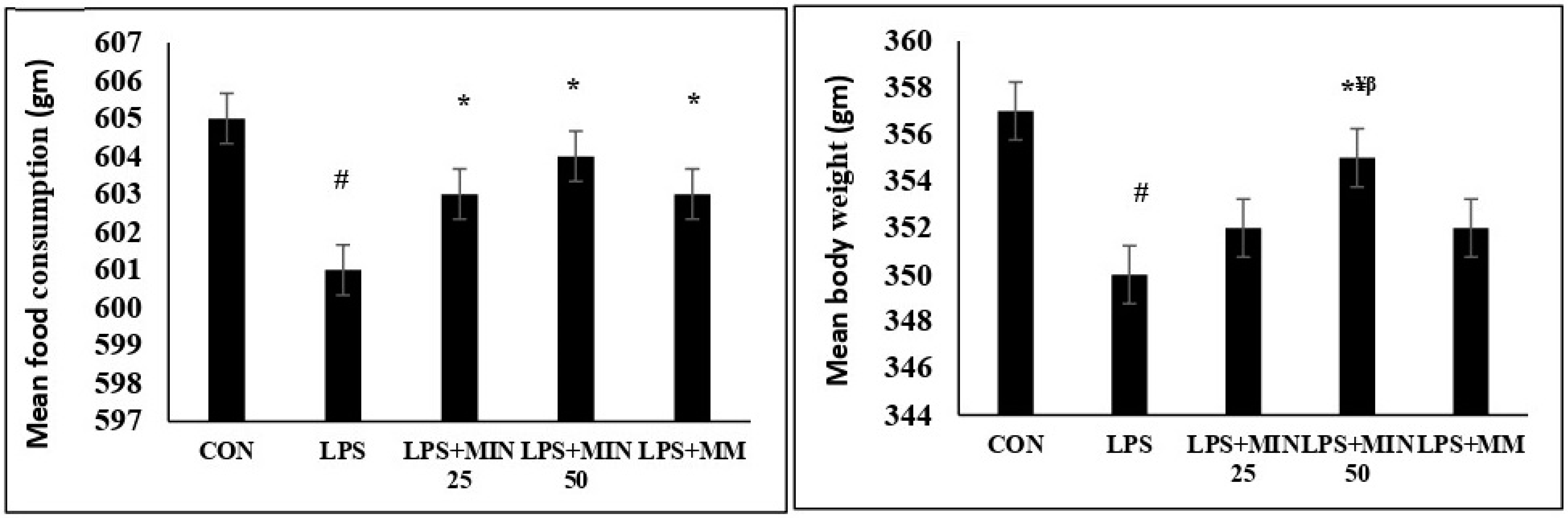
2.1. Effects of Minocycline on Food Intake and Body Weight in LPS-Injected Rats
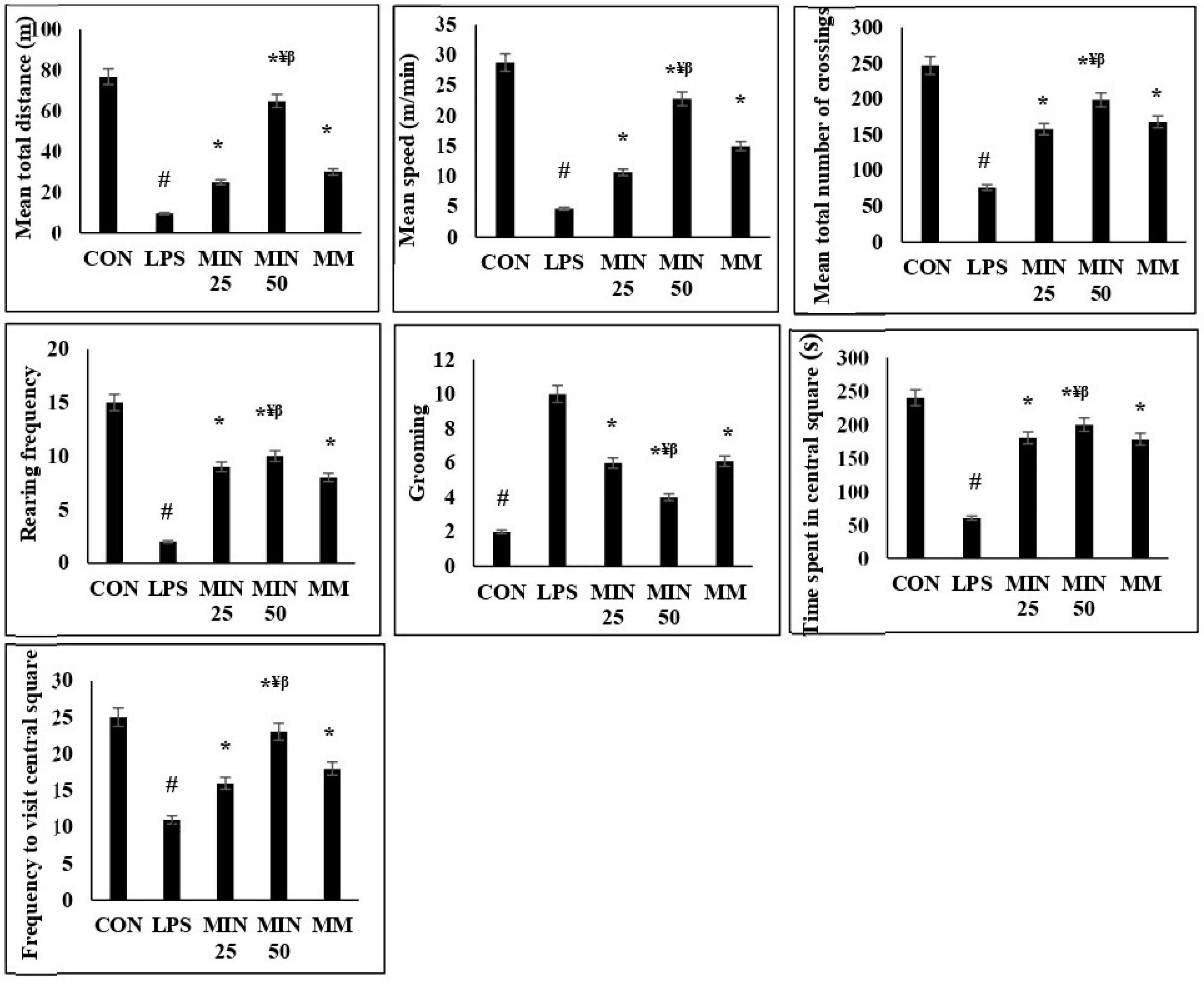
2.2. Effects of Minocycline on the Locomotor Deficits and Anxiety-like Behaviour in LPS-Injected Rats
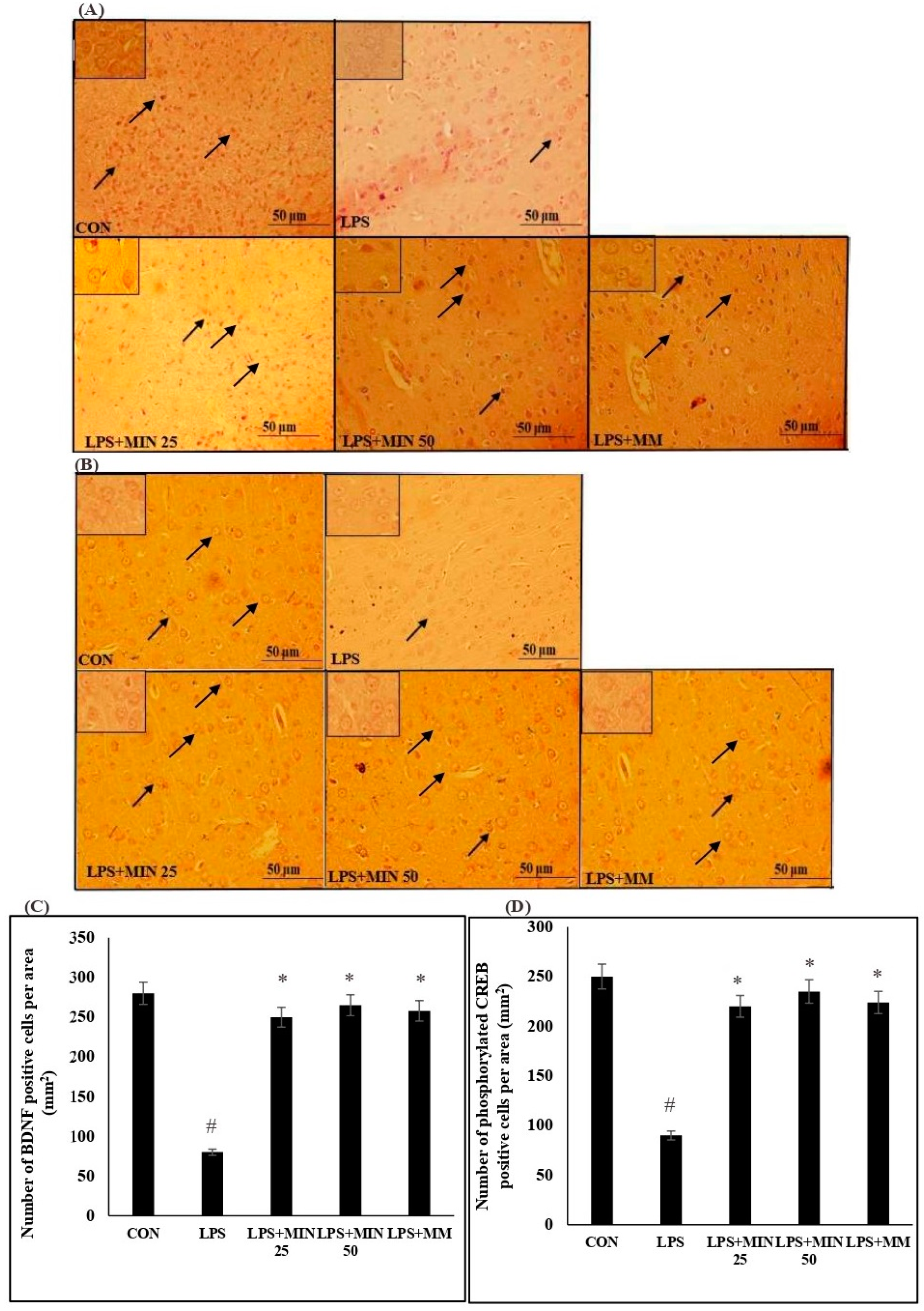
2.3. Effects of Minocycline on the Expression of BDNF- and Phosphorylated CREB-Positive Cells in LPS-Injected Rats
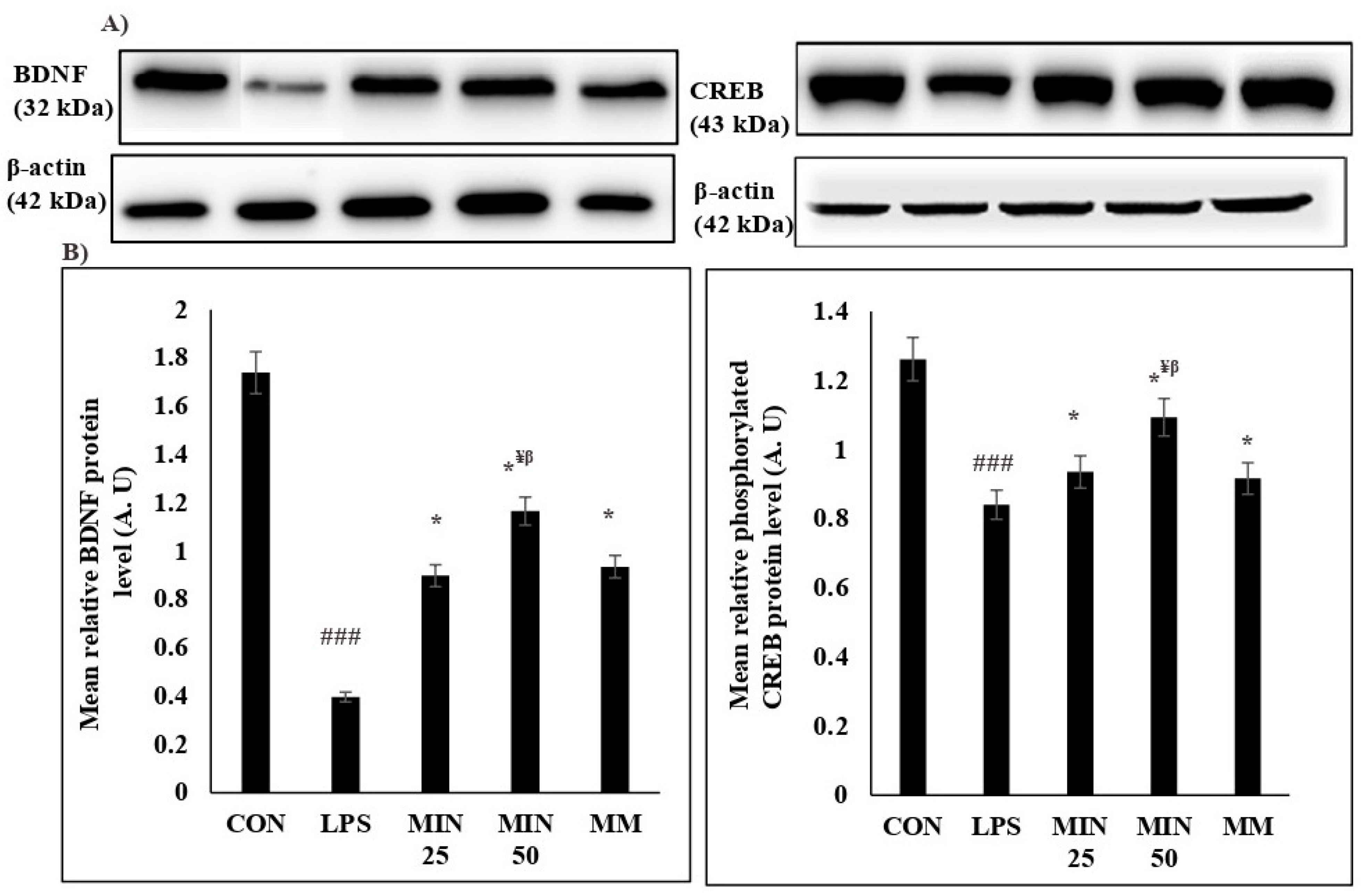
2.4. Effects of Minocycline on the Expression Levels of BDNF and Phosphorylated CREB Proteins in LPS-Injected Rats
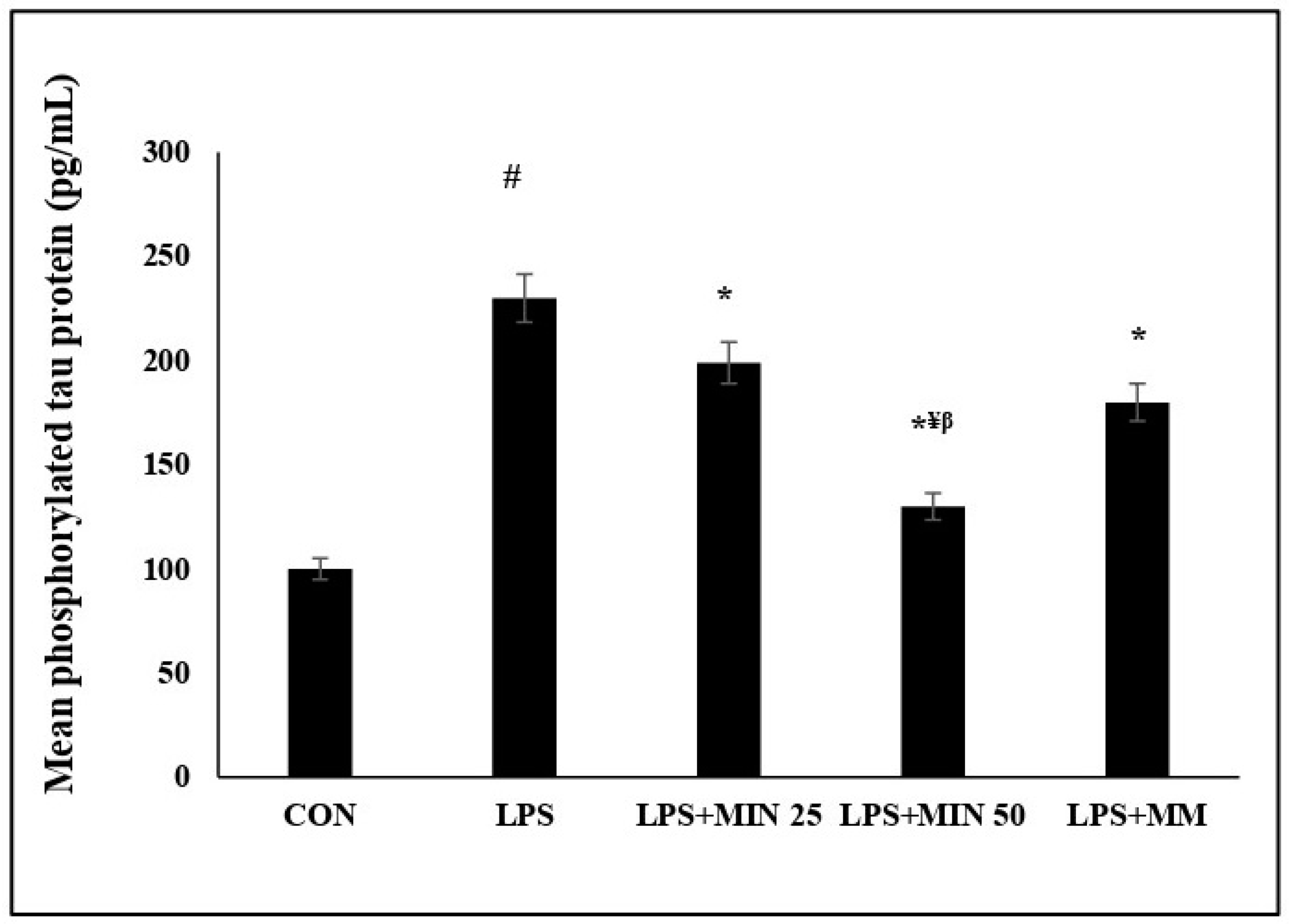
2.5. Effects of Minocycline on Phosphorylated Tau Protein Expression Level on LPS-Injected Rats
3. Discussion
4. Materials and Methods
4.1. Animals
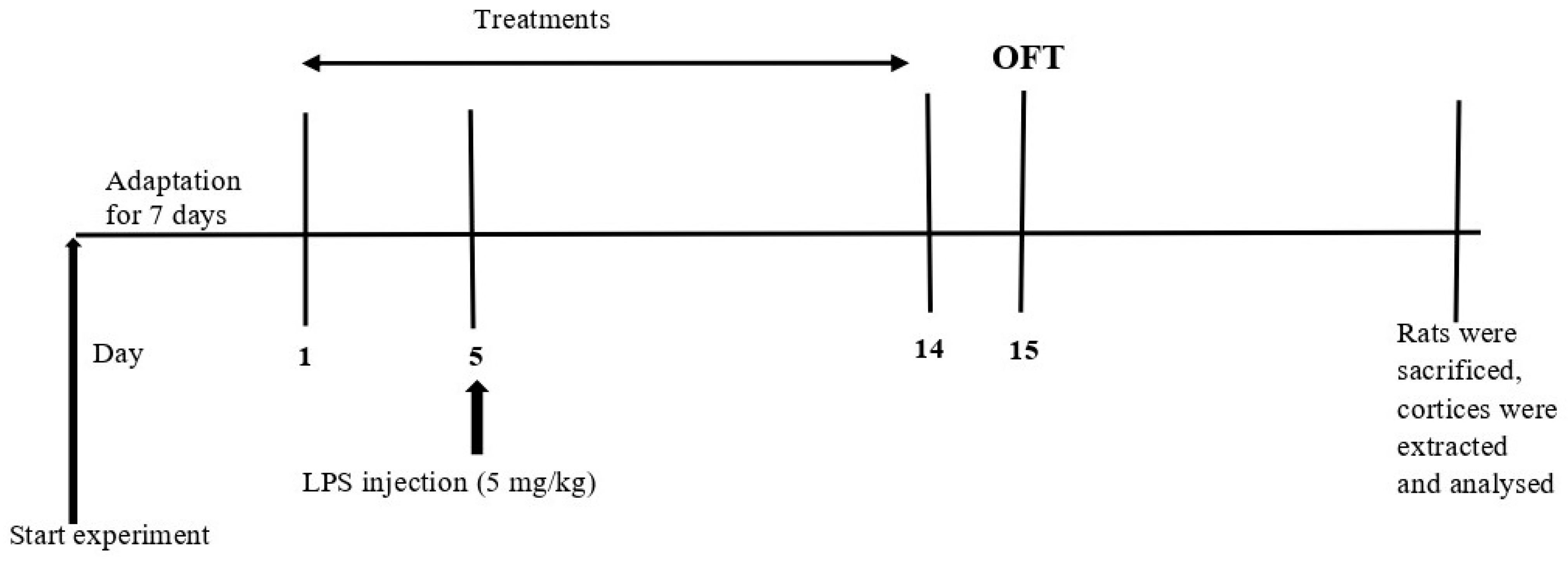
4.2. Experimental Design
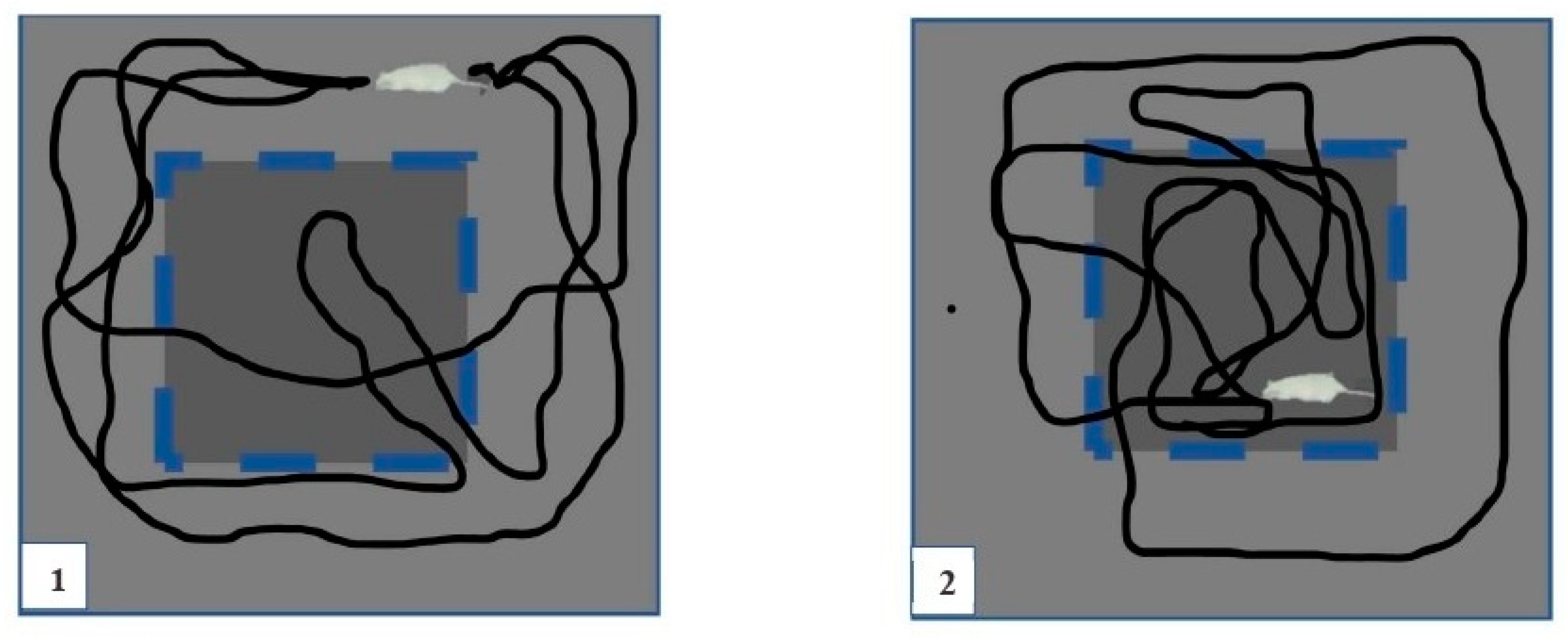
4.3. Open-Field Test (OFT)
4.4. Immunohistochemistry for BDNF- and Phosphorylated CREB-Positive Cell Expressions
4.5. Western Blotting for BDNF and Phosphorylated CREB Protein Expression Levels
4.6. Phosphorylated Tau Protein Assay by Enzyme-Linked Immunosorbent Assay (ELISA)
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shishkina, G.T.; Bannova, A.V.; Komysheva, N.P.; Dygalo, N.N. Anxiogenic-like effect of chronic lipopolysaccharide is associated with increased expression of matrix metalloproteinase 9 in the rat amygdala. Stress 2020, 23, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Belzung, C.; Griebel, G. Measuring normal and pathological anxiety-like behaviour in mice: A review. Behav. Brain Res. 2001, 125, 141–149. [Google Scholar] [CrossRef]
- Tufvesson-Alm, M.; Imbeault, S.; Liu, X.C.; Zheng, Y.; Faka, A.; Choi, D.S.; Schwieler, L.; Engberg, G.; Erhardt, S. Repeated administration of LPS exaggerates amphetamine-induced locomotor response and causes learning deficits in mice. J. Neuroimmunol. 2020, 349, 577401. [Google Scholar] [CrossRef] [PubMed]
- Ge, L.; Liu, L.; Liu, H.; Liu, S.; Xue, H.; Wang, X.; Yuan, L.; Wang, Z.; Liu, D. Resveratrol abrogates lipopolysaccharide-induced depressive-like behavior, neuroinflammatory response, and CREB/BDNF signaling in mice. Eur. J. Pharmacol. 2015, 768, 49–57. [Google Scholar] [CrossRef]
- Zhang, L.; Previn, R.; Lu, L.; Liao, R.-F.; Jin, Y.; Wang, R.-L. Crocin a natural product attenuates lipopolysaccharide-induced anxiety and depressive-like behaviors through suppressing NF-кβ and NLRP3 signaling pathway. Brain Research Bulletin. 2018, 142, 352–359. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, L.; Chen, J.; Liu, C.; Li, S.; Hua, W.; Qu, D.; Shao, Z.; Sun, Y. Ginsenoside RK1 alleviates LPS-induced depression-like behavior in mice by promoting BDNF and suppressing the neuroinflammatory response. Biochem. Biophys. Res. Commun. 2020, 530, 658–664. [Google Scholar] [CrossRef]
- Liu, J.; Wang, D.; Li, S.-Q.; Yu, Y.; Ye, R.D. Suppression of LPS-induced tau hyperphosphorylation by serum amyloid A. J. Neuroinflamm. 2016, 13, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Badshah, H.; Ali, T.; Kim, M.O. Osmotin attenuates LPS-induced neuroinflammation and memory impairment via the TLR4/NFкβ signaling pathway. Sci. Reports 2016, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Gibney, S.M.; McGuinness, B.; Prendergast, C.; Harkin, A.; Connor, T.J. Poly I:C-induced activation of the immune response is accompanied by depression and anxiety-like behaviours, kynurenine pathway activation and reduced BDNF expression. Brain Behav. Immun. 2013, 28, 170–181. [Google Scholar] [CrossRef]
- Rosa, E.; Mahendram, S.; Ke, Y.D.; Ittner, L.M.; Ginsberg, S.D.; Fahnestock, M. Tau downregulates BDNF expression in animal and cellular models of Alzheimer’s disease. Neurobiol. Aging 2016, 48, 135–142. [Google Scholar] [CrossRef]
- Jiang, C.; Salton, S.R. The role of neurotrophins in major depressive disorder. Transl. Neurosci. 2013, 4, 46–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Tang, B.; Liao, X.; Su, Z.; Lee, S.M.-Y.; Cai, Y.; Li, C. Suppressive effects of the supercritical-carbon dioxide fluid extract of Chrysanthemum indicum on chronic unpredictable mild stress-induced depressive-like behavior in mice. Food Funct. 2019, 10, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Valverde, O.; Mantamadiotis, T.; Torrecilla, M.; Ugedo, L.; Pineda, J.; Bleckmann, S.; Gass, P.; Kretz, O.; Mitchell, J.M.; Schütz, G.; et al. Modulation of anxiety-like behavior and morphine dependence in CREB-deficient mice. Neuropsychopharmacology 2004, 29, 1122–1133. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.; Yao, H.; Li, Y.; Ding, R.; Ren, X.; Tan, Y.; Ren, W.; Yu, H.; Zhan, X.; Wang, X.; et al. Regulation of tau protein on the antidepressant effects of ketamine in the chronic unpredictable mild stress model. Front. Psychiatry 2019, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Onogi, H.; Ishigaki, S.; Nakagawasai, O.; Arai-Kato, Y.; Arai, Y.; Watanabe, H.; Miyamoto, A.; Tan-No, K.; Tadano, T. Influence of memantine on brain monoaminergic neurotransmission parameters in mice: Neurochemical and behavioral study. Biol. Pharm. Bull. 2009, 32, 850–855. [Google Scholar] [CrossRef] [Green Version]
- Yaacob, W.M.H.W.; Long, I.; Zakaria, R.; Othman, Z. Tualang Honey and its Methanolic Fraction Improve LPS-induced Learning and Memory Impairment in Male Rats: Comparison with Memantine. Curr. Nutr. Food Sci. 2018, 16, 333–342. [Google Scholar] [CrossRef]
- Kos, T.; Popik, P. A comparison of the predictive therapeutic and undesired side-effects of the NMDA receptor antagonist, memantine, in mice. Behav. Pharmacol. 2005, 16, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Shuvaev, A.N.; Belozor, O.S.; Mozhei, O.I.; Mileiko, A.G.; Mosina, L.D.; Laletina, I.V.; Mikhailov, I.G.; Fritsler, Y.V.; Shuvaev, A.N.; Teschemacher, A.G.; et al. Memantine Disrupts Motor Coordination through Anxiety-like Behavior in CD1 Mice. Brain Sci. 2022, 12, 495. [Google Scholar] [CrossRef]
- Yong, V.W.; Wells, J.; Giuliani, F.; Casha, S.; Power, C.; Metz, L.M. The promise of minocycline in neurology. Lancet Neurol. 2004, 3, 744–751. [Google Scholar] [CrossRef]
- Plane, J.M.; Shen, Y.; Pleasure, D.E.; Deng, W. Prospects for minocycline neuroprotection. Arch. Neurol. 2010, 67, 1442–1448. [Google Scholar] [CrossRef]
- Cruz, S.L.; Armenta-Reséndiz, M.; Carranza-Aguilar, C.J.; Galván, E.J. Minocycline prevents neuronal hyperexcitability and neuroinflammation in medial prefrontal cortex, as well as memory impairment caused by repeated toluene inhalation in adolescent rats. Toxicol. Appl. Pharmacol. 2020, 395, 114980. [Google Scholar] [CrossRef]
- Rooney, S.; Sah, A.; Unger, M.S.; Kharitonova, M.; Sartori, S.B.; Schwarzer, C.; Aigner, L.; Kettenmann, H.; Wolf, S.A.; Singewald, N. Neuroinflammatory alterations in trait anxiety: Modulatory effects of minocycline. Transl. Psychiatry 2020, 10, 256. [Google Scholar] [CrossRef]
- Salehi, P.; Shahmirzadi, Z.Y.; Mirrezaei, F.S.; Shirvani Boushehri, F.; Mayahi, F.; Songhori, M.; Abofazeli, M.; Motaghinejad, M.; Safari, S. A hypothetic role of minocycline as a neuroprotective agent against methylphenidate-induced neuronal mitochondrial dysfunction and tau protein hyper-phosphorylation: Possible role of PI3/Akt/GSK3β signaling pathway. Med. Hypotheses 2019, 128, 6–10. [Google Scholar] [CrossRef]
- Motaghinejad, M.; Mashayekh, R.; Motevalian, M.; Safari, S. The possible role of CREB-BDNF signaling pathway in neuroprotective effects of minocycline against alcohol-induced neurodegeneration: Molecular and behavioral evidences. Fundam. Clin. Pharmacol. 2021, 35, 113–130. [Google Scholar] [CrossRef]
- Molina-Hernández, M.; Tellez-Alcántara, N.P.; Pérez-García, J.; Olivera-Lopez, J.I.; Jaramillo-Jaimes, M.T. Antidepressant-like actions of minocycline combined with several glutamate antagonists. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 380–386. [Google Scholar] [CrossRef]
- Arakawa, S.; Shirayama, Y.; Fujita, Y.; Ishima, T.; Horio, M.; Muneoka, K.; Iyo, M.; Hashimoto, K. Minocycline produced antidepressant-like effects on the learned helplessness rats with alterations in levels of monoamine in the amygdala and no changes in BDNF levels in the hippocampus at baseline. Pharmacol. Biochem. Behav. 2012, 100, 601–606. [Google Scholar] [CrossRef]
- Fan, L.W.; Pang, Y.; Lin, S.; Tien, L.T.; Ma, T.; Rhodes, P.G.; Cai, Z. Minocycline reduces lipopolysaccharide-induced neurological dysfunction and brain injury in the neonatal rat. J. Neurosci. Res. 2005, 82, 71–82. [Google Scholar] [CrossRef]
- Castanon, N.; Bluthé, R.M.; Dantzer, R. Chronic treatment with the atypical antidepressant tianeptine attenuates sickness behavior induced by peripheral but not central lipopolysaccharide and interleukin-1β in the rat. Psychopharmacology 2001, 154, 50–60. [Google Scholar] [CrossRef]
- Qaid, E.Y.A.; Abdullah, Z.; Zakaria, R.; Long, I. Minocycline Ameliorates LPS-Induced Learning and Memory Impairment By Inhibiting Microglia and Astrocytes Activation in the Hippocampus. IIUM Med. J. Malays. 2022, 21, 53–59. [Google Scholar] [CrossRef]
- Rosi, S.; Vazdarjanova, A.; Ramirez-Amaya, V.; Worley, P.F.; Barnes, C.A.; Wenk, G.L. Memantine protects against LPS-induced neuroinflammation, restores behaviorally-induced gene expression and spatial learning in the rat. Neuroscience 2006, 142, 1303–1315. [Google Scholar] [CrossRef]
- Kinoshita, D.; Cohn, D.W.H.; Costa-Pinto, F.A.; de Sá-Rocha, L.C. Behavioral effects of LPS in adult, middle-aged and aged mice. Physiol. Behav. 2009, 96, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Murray, E.; Smith, K.B.; Stoby, K.S.; Thomas, B.J.; Swenson, M.J.; Arber, L.A.; Frenette, E.; Ismail, N. Pubertal probiotic blocks LPS-induced anxiety and the associated neurochemical and microbial outcomes, in a sex dependent manner. Psychoneuroendocrinology 2020, 112, 104481. [Google Scholar] [CrossRef] [PubMed]
- Afshary, K.; Chamanara, M.; Talari, B.; Rezaei, P.; Nassireslami, E. Therapeutic Effects of Minocycline Pretreatment in the Locomotor and Sensory Complications of Spinal Cord Injury in an Animal Model. J. Mol. Neurosci. 2020, 70, 1064–1072. [Google Scholar] [CrossRef]
- Motaghinejad, M.; Motevalian, M. Neuroprotective Properties of Minocycline Against Methylphenidate-Induced Neurodegeneration: Possible Role of CREB/BDNF and Akt/GSK3 Signaling Pathways in Rat Hippocampus. Neurotox. Res. 2022, 40, 689–713. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Hidau, M.K.; Rai, S. Memantine treatment exerts an antidepressant-like effect by preventing hippocampal mitochondrial dysfunction and memory impairment via upregulation of CREB/BDNF signaling in the rat model of chronic unpredictable stress-induced depression. Neurochem. Int. 2021, 142, 104932. [Google Scholar] [CrossRef]
- Adzic, M.; Djordjevic, J.; Mitic, M.; Brkic, Z.; Lukic, I.; Radojcic, M. The contribution of hypothalamic neuroendocrine, neuroplastic and neuroinflammatory processes to lipopolysaccharide-induced depressive-like behaviour in female and male rats: Involvement of glucocorticoid receptor and C/EBP-β. Behav. Brain Res. 2015, 291, 130–139. [Google Scholar] [CrossRef] [PubMed]
- Cappoli, N.; Tabolacci, E.; Aceto, P.; Dello Russo, C. The emerging role of the BDNF-TrkB signaling pathway in the modulation of pain perception. J. Neuroimmunol. 2020, 349, 577406. [Google Scholar] [CrossRef]
- Schnydrig, S.; Korner, L.; Landweer, S.; Ernst, B.; Walker, G.; Otten, U.; Kunz, D. Peripheral lipopolysaccharide administration transiently affects expression of brain-derived neurotrophic factor, corticotropin and proopiomelanocortin in mouse brain. Neurosci. Lett. 2007, 429, 69–73. [Google Scholar] [CrossRef]
- Yang, S.; Gong, Q.; Wu, Q.; Li, F.; Lu, Y.; Shi, J. Alkaloids enriched extract from Dendrobium nobile Lindl. attenuates tau protein hyperphosphorylation and apoptosis induced by lipopolysaccharide in rat brain. Phytomedicine 2014, 21, 712–716. [Google Scholar] [CrossRef]
- Beheshti Nasr, S.M.; Moghimi, A.; Mohammad-Zadeh, M.; Shamsizadeh, A.; Noorbakhsh, S.M. The effect of minocycline on seizures induced by amygdala kindling in rats. Seizure 2013, 22, 670–674. [Google Scholar] [CrossRef]
- Hemmati, F.; Dargahi, L.; Nasoohi, S.; Omidbakhsh, R. Neurorestorative effect of FTY720 in a rat model of Alzheimer’s disease: Comparison with memantine. Behav. Brain Res. 2013, 252, 415–421. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdo Qaid, E.Y.; Abdullah, Z.; Zakaria, R.; Long, I. Minocycline Attenuates Lipopolysaccharide-Induced Locomotor Deficit and Anxiety-like Behavior and Related Expression of the BDNF/CREB Protein in the Rat Medial Prefrontal Cortex (mPFC). Int. J. Mol. Sci. 2022, 23, 13474. https://doi.org/10.3390/ijms232113474
Abdo Qaid EY, Abdullah Z, Zakaria R, Long I. Minocycline Attenuates Lipopolysaccharide-Induced Locomotor Deficit and Anxiety-like Behavior and Related Expression of the BDNF/CREB Protein in the Rat Medial Prefrontal Cortex (mPFC). International Journal of Molecular Sciences. 2022; 23(21):13474. https://doi.org/10.3390/ijms232113474
Chicago/Turabian StyleAbdo Qaid, Entesar Yaseen, Zuraidah Abdullah, Rahimah Zakaria, and Idris Long. 2022. "Minocycline Attenuates Lipopolysaccharide-Induced Locomotor Deficit and Anxiety-like Behavior and Related Expression of the BDNF/CREB Protein in the Rat Medial Prefrontal Cortex (mPFC)" International Journal of Molecular Sciences 23, no. 21: 13474. https://doi.org/10.3390/ijms232113474