
Availability of Ferritin-Bound Iron to Enterobacteriaceae
 , , , , , and
, , , , , and 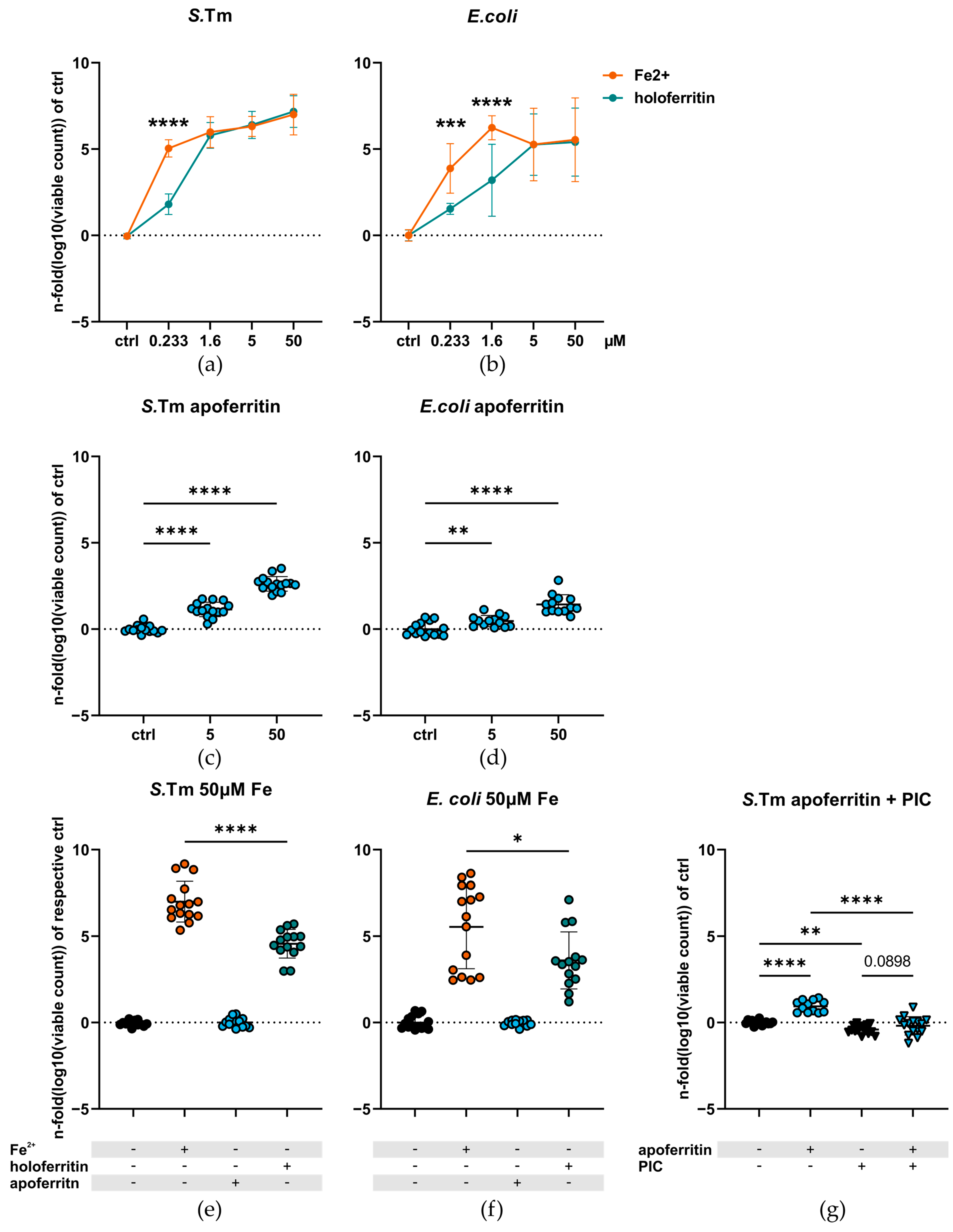{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Ferritin-Bound Iron Is Less Available for Enterobacteriaceae Than Ionic Ferrous Iron
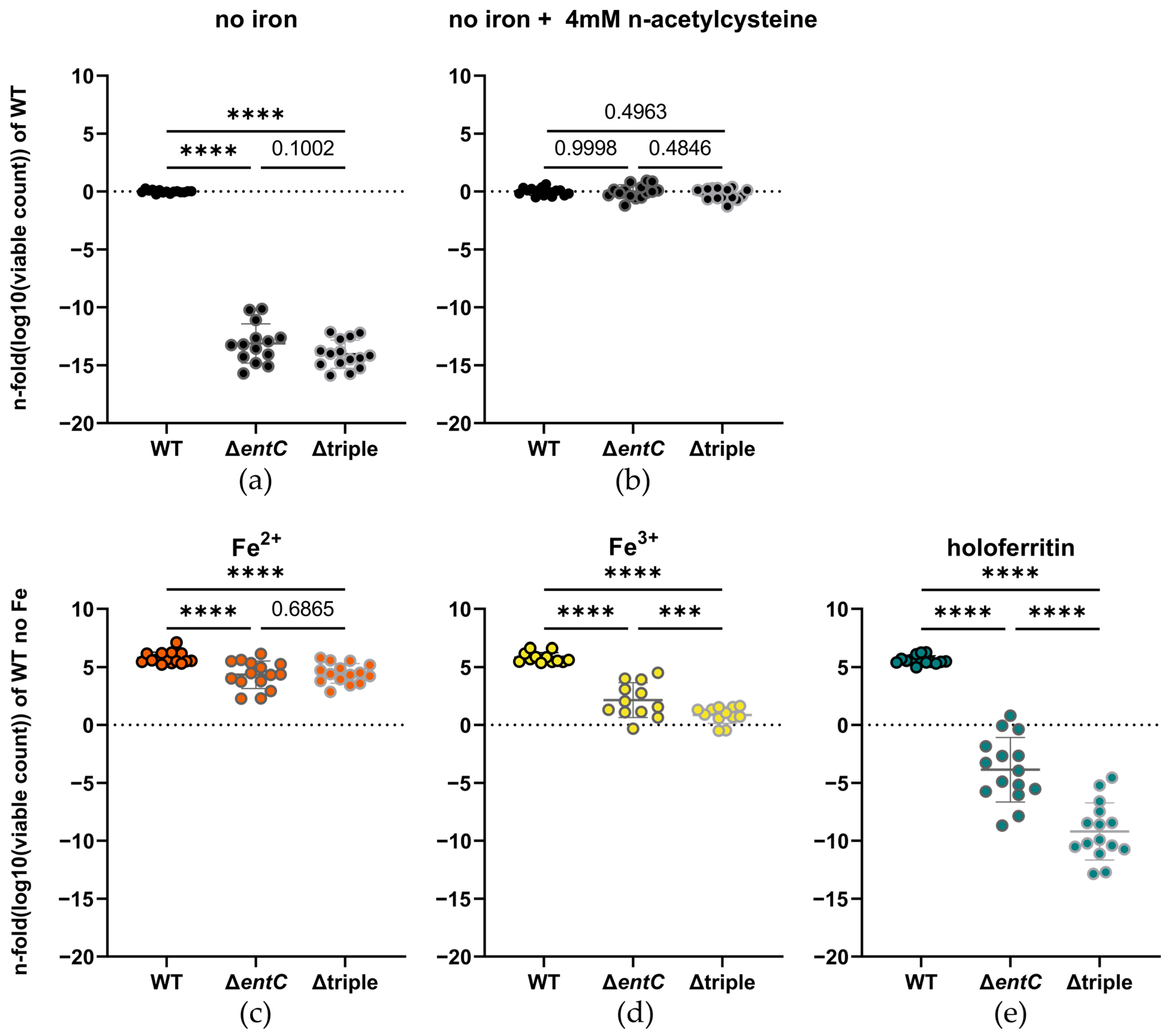
2.2. Iron Acquisition from Ferritin Is Still Possible in Absence of Enterobactin and Salmochelins
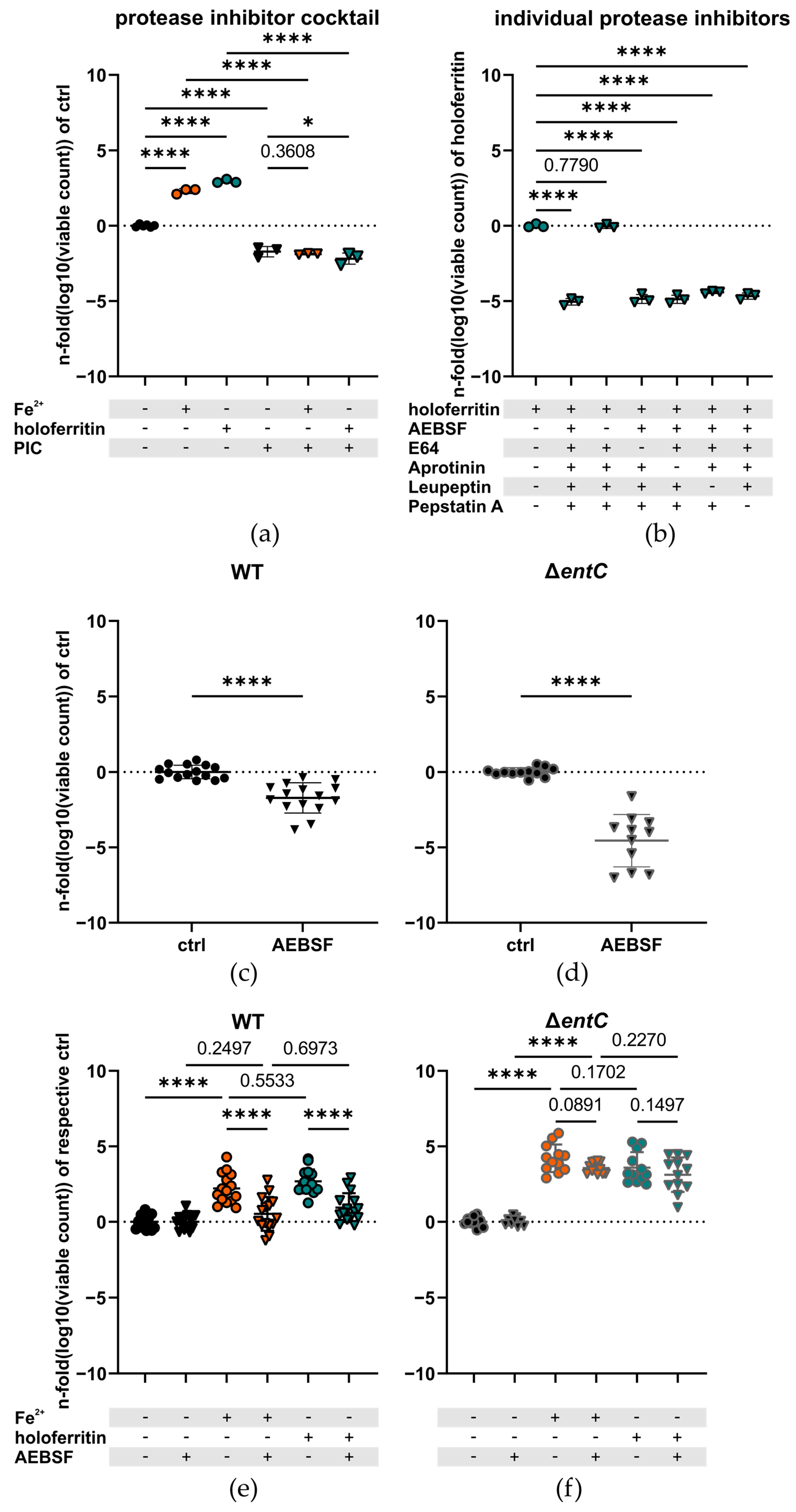
2.3. The Serine Protease Inhibitor AEBSF Impairs Iron Utilization in the WT but Not the ΔentC Mutant
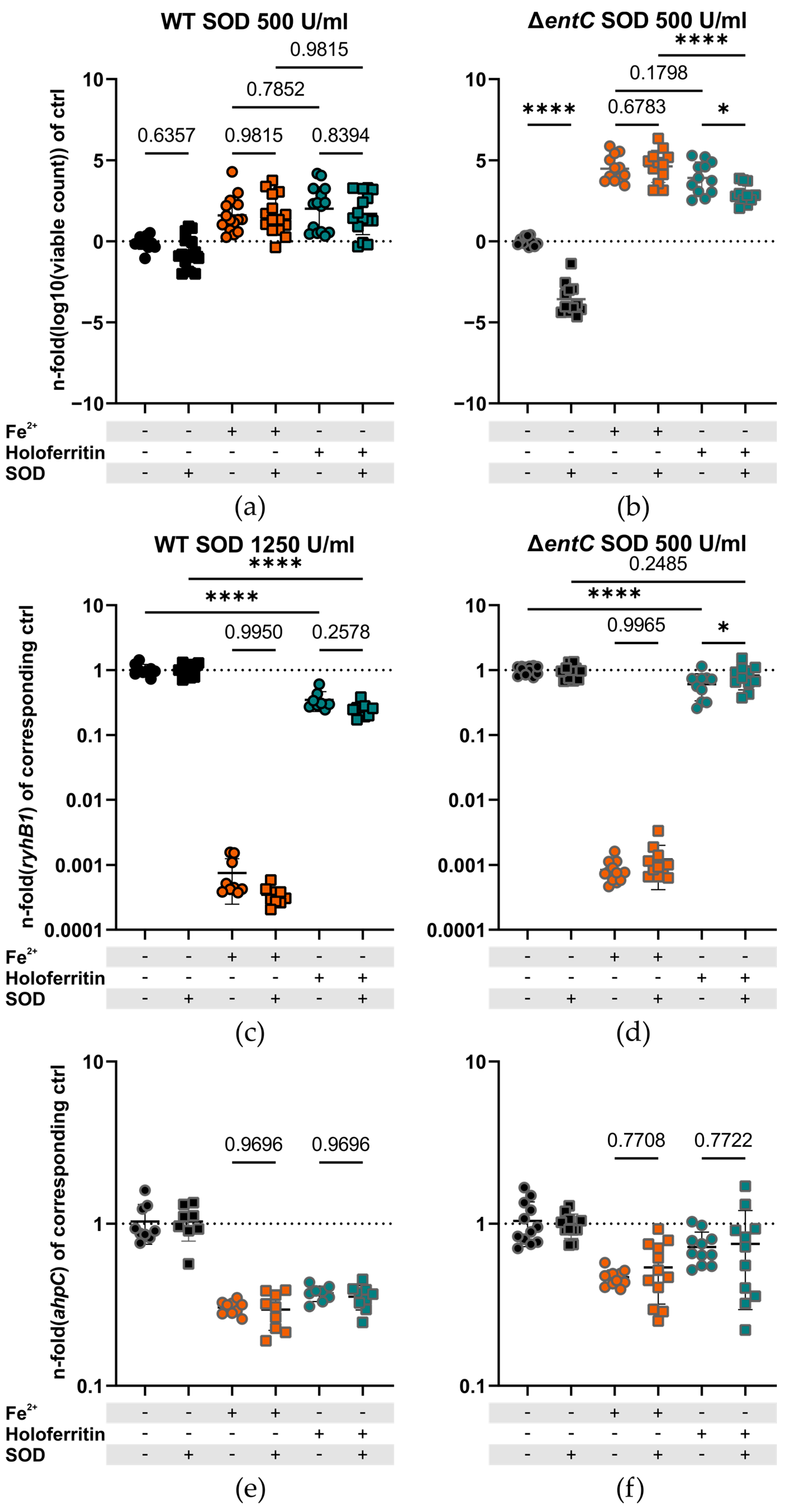
2.4. Superoxide Is Necessary for Bacterial Iron Acquisition from Ferritin in the Absence of Enterobactin and Salmochelins
3. Discussion
4. Materials and Methods
4.1. Reagents, Media and Laboratory Supplies
4.2. Bacterial Strains and General Growth Conditions
4.3. General Experimental Conditions
4.4. Specific Experimental Conditions
4.5. Quantification of Iron Released from Ferritin
4.6. Quantitative Real-Time PCR
- 16S rRNA: CGGTGAATACGTTCYCGG; GGWTACCTTGTTACGACTT; CTTGTACACACCGCCCGTC
- rpoB: GATGCGTCCCGTATCGTTATC; CTGGTTAGAGCGGGTGTATTT;
- ryhB1: TACGGAGAACCTGAAAGCAC; AATAATACTGGAAGCAATGTGAG;
4.7. Statistical Analysis and Data Handling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stanaway, J.D.; Parisi, A.; Sarkar, K.; Blacker, B.F.; Reiner, R.C.; Hay, S.I.; Nixon, M.R.; Dolecek, C.; James, S.L.; Mokdad, A.H.; et al. The global burden of non-typhoidal salmonella invasive disease: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Infect. Dis. 2019, 19, 1312–1324. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.D.t.; Bagamian, K.H.; Muhib, F.; Amaya, M.P.; Laytner, L.A.; Wierzba, T.; Rheingans, R. Burden of enterotoxigenic Escherichia coli and shigella non-fatal diarrhoeal infections in 79 low-income and lower middle-income countries: A modelling analysis. Lancet Glob. Health 2019, 7, e321–e330. [Google Scholar] [CrossRef] [Green Version]
- Tan, Z.; Chekabab, S.M.; Yu, H.; Yin, X.; Diarra, M.S.; Yang, C.; Gong, J. Growth and Virulence of Salmonella Typhimurium Mutants Deficient in Iron Uptake. ACS Omega 2019, 4, 13218–13230. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Haschka, D.; Demetz, E.; Weiss, G. Iron at the interface of immunity and infection. Front. Pharmacol. 2014, 5, 152. [Google Scholar] [CrossRef] [Green Version]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Healy, C.; Munoz-Wolf, N.; Strydom, J.; Faherty, L.; Williams, N.C.; Kenny, S.; Donnelly, S.C.; Cloonan, S.M. Nutritional immunity: The impact of metals on lung immune cells and the airway microbiome during chronic respiratory disease. Respir. Res. 2021, 22, 133. [Google Scholar] [CrossRef] [PubMed]
- Nairz, M.; Weiss, G. Iron in infection and immunity. Mol. Asp. Med. 2020, 75, 100864. [Google Scholar] [CrossRef]
- Brissot, P.; Ropert, M.; Le Lan, C.; Loréal, O. Non-transferrin bound iron: A key role in iron overload and iron toxicity. Biochim. Et Biophys. Acta 2012, 1820, 403–410. [Google Scholar] [CrossRef]
- Zhao, G.; Arosio, P.; Chasteen, N.D. Iron(II) and hydrogen peroxide detoxification by human H-chain ferritin. An EPR spin-trapping study. Biochemistry 2006, 45, 3429–3436. [Google Scholar] [CrossRef] [PubMed]
- Rosário, C.; Zandman-Goddard, G.; Meyron-Holtz, E.G.; D’Cruz, D.P.; Shoenfeld, Y. The Hyperferritinemic Syndrome: Macrophage activation syndrome, Still’s disease, septic shock and catastrophic antiphospholipid syndrome. BMC Med. 2013, 11, 185. [Google Scholar] [CrossRef] [PubMed]
- Konijn, A.M.; Hershko, C. Ferritin synthesis in inflammation. I. Pathogenesis of impaired iron release. Br. J. Haematol. 1977, 37, 7–16. [Google Scholar] [PubMed]
- Ruddell, R.G.; Hoang-Le, D.; Barwood, J.M.; Rutherford, P.S.; Piva, T.J.; Watters, D.J.; Santambrogio, P.; Arosio, P.; Ramm, G.A. Ferritin functions as a proinflammatory cytokine via iron-independent protein kinase C zeta/nuclear factor kappaB-regulated signaling in rat hepatic stellate cells. Hepatology 2009, 49, 887–900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darton, T.C.; Blohmke, C.J.; Giannoulatou, E.; Waddington, C.S.; Jones, C.; Sturges, P.; Webster, C.; Drakesmith, H.; Pollard, A.J.; Armitage, A.E. Rapidly Escalating Hepcidin and Associated Serum Iron Starvation Are Features of the Acute Response to Typhoid Infection in Humans. PLoS Negl. Trop. Dis. 2015, 9, e0004029. [Google Scholar] [CrossRef]
- Hoffmann, A.; Haschka, D.; Valente de Souza, L.; Tymoszuk, P.; Seifert, M.; von Raffay, L.; Hilbe, R.; Petzer, V.; Moser, P.L.; Nairz, M.; et al. Baseline iron status and presence of anaemia determine the course of systemic Salmonella infection following oral iron supplementation in mice. eBioMedicine 2021, 71, 103568. [Google Scholar] [CrossRef]
- Dehner, C.; Morales-Soto, N.; Behera, R.K.; Shrout, J.; Theil, E.C.; Maurice, P.A.; Dubois, J.L. Ferritin and ferrihydrite nanoparticles as iron sources for Pseudomonas aeruginosa. J. Biol. Inorg. Chem. JBIC A Publ. Soc. Biol. Inorg. Chem. 2013, 18, 371–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitby, P.W.; VanWagoner, T.M.; Springer, J.M.; Morton, D.J.; Seale, T.W.; Stull, T.L. Burkholderia cenocepacia utilizes ferritin as an iron source. J. Med. Microbiol. 2006, 55, 661–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teawtrakul, N.; Jetsrisuparb, A.; Sirijerachai, C.; Chansung, K.; Wanitpongpun, C. Severe bacterial infections in patients with non-transfusion-dependent thalassemia: Prevalence and clinical risk factors. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2015, 39, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Adamkiewicz, T.V.; Berkovitch, M.; Krishnan, C.; Polsinelli, C.; Kermack, D.; Olivieri, N.F. Infection due to Yersinia enterocolitica in a series of patients with beta-thalassemia: Incidence and predisposing factors. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 1998, 27, 1362–1366. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.C.; Lin, K.H.; Chern, J.P.; Lu, M.Y.; Jou, S.T.; Lin, D.T.; Lin, K.S. Severe bacterial infection in transfusion-dependent patients with thalassemia major. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2003, 37, 984–988. [Google Scholar] [CrossRef]
- Harrison, P.M.; Arosio, P. The ferritins: Molecular properties, iron storage function and cellular regulation. Biochim. Et Biophys. Acta BBA Bioenerg. 1996, 1275, 161–203. [Google Scholar] [CrossRef]
- Mehlenbacher, M.; Poli, M.; Arosio, P.; Santambrogio, P.; Levi, S.; Chasteen, N.D.; Bou-Abdallah, F. Iron Oxidation and Core Formation in Recombinant Heteropolymeric Human Ferritins. Biochemistry 2017, 56, 3900–3912. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.; Firlar, E.; Rasul, M.G.; Foroozan, T.; Farajpour, N.; Covnot, L.; Shahbazian-Yassar, R.; Shokuhfar, T. On the structure and chemistry of iron oxide cores in human heart and human spleen ferritins using graphene liquid cell electron microscopy. Nanoscale 2019, 11, 16868–16878. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chasteen, N.D. Iron oxidation chemistry in ferritin. Increasing Fe/O2 stoichiometry during core formation. J. Biol. Chem. 1991, 266, 19965–19970. [Google Scholar] [CrossRef]
- Li, W.; Garringer, H.J.; Goodwin, C.B.; Richine, B.; Acton, A.; VanDuyn, N.; Muhoberac, B.B.; Irimia-Dominguez, J.; Chan, R.J.; Peacock, M.; et al. Systemic and cerebral iron homeostasis in ferritin knock-out mice. PLoS ONE 2015, 10, e0117435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velayudhan, J.; Castor, M.; Richardson, A.; Main-Hester, K.L.; Fang, F.C. The role of ferritins in the physiology of Salmonella enterica sv. Typhimurium: A unique role for ferritin B in iron-sulphur cluster repair and virulence. Mol. Microbiol. 2007, 63, 1495–1507. [Google Scholar] [CrossRef]
- Harris, W.R.; Carrano, C.J.; Cooper, S.R.; Sofen, S.R.; Avdeef, A.E.; McArdle, J.V.; Raymond, K.N. Coordination chemistry of microbial iron transport compounds. 19. Stability constants and electrochemical behavior of ferric enterobactin and model complexes. J. Am. Chem. Soc. 1979, 101, 6097–6104. [Google Scholar] [CrossRef]
- Aisen, P.; Leibman, A.; Zweier, J. Stoichiometric and site characteristics of the binding of iron to human transferrin. J. Biol. Chem. 1978, 253, 1930–1937. [Google Scholar] [CrossRef]
- Carrano, C.J.; Raymond, K.N. Ferric ion sequestering agents. 2. Kinetics and mechanism of iron removal from transferrin by enterobactin and synthetic tricatechols. J. Am. Chem. Soc. 1979, 101, 5401–5404. [Google Scholar] [CrossRef]
- Buchanan, S.K.; Smith, B.S.; Venkatramani, L.; Xia, D.; Esser, L.; Palnitkar, M.; Chakraborty, R.; van der Helm, D.; Deisenhofer, J. Crystal structure of the outer membrane active transporter FepA from Escherichia coli. Nat. Struct. Biol. 1999, 6, 56–63. [Google Scholar] [CrossRef]
- Sprencel, C.; Cao, Z.; Qi, Z.; Scott, D.C.; Montague, M.A.; Ivanoff, N.; Xu, J.; Raymond, K.M.; Newton, S.M.; Klebba, P.E. Binding of ferric enterobactin by the Escherichia coli periplasmic protein FepB. J. Bacteriol. 2000, 182, 5359–5364. [Google Scholar] [CrossRef]
- Pierce, J.R.; Earhart, C.F. Escherichia coli K-12 envelope proteins specifically required for ferrienterobactin uptake. J. Bacteriol. 1986, 166, 930–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chenault, S.S.; Earhart, C.F. Identification of hydrophobic proteins FepD and FepG of the Escherichia coli ferrienterobactin permease. J. Gen. Microbiol. 1992, 138, 2167–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brickman, T.J.; McIntosh, M.A. Overexpression and purification of ferric enterobactin esterase from Escherichia coli. Demonstration of enzymatic hydrolysis of enterobactin and its iron complex. J. Biol. Chem. 1992, 267, 12350–12355. [Google Scholar] [CrossRef]
- Zhu, M.; Valdebenito, M.; Winkelmann, G.; Hantke, K. Functions of the siderophore esterases IroD and IroE in iron-salmochelin utilization. Microbiology 2005, 151, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Fischbach, M.A.; Liu, D.R.; Walsh, C.T. In vitro characterization of salmochelin and enterobactin trilactone hydrolases IroD, IroE, and Fes. J. Am. Chem. Soc. 2005, 127, 11075–11084. [Google Scholar] [CrossRef] [Green Version]
- Peralta, D.R.; Adler, C.; Corbalán, N.S.; Paz García, E.C.; Pomares, M.F.; Vincent, P.A. Enterobactin as Part of the Oxidative Stress Response Repertoire. PLoS ONE 2016, 11, e0157799. [Google Scholar] [CrossRef] [Green Version]
- Achard, M.E.; Chen, K.W.; Sweet, M.J.; Watts, R.E.; Schroder, K.; Schembri, M.A.; McEwan, A.G. An antioxidant role for catecholate siderophores in Salmonella. Biochem. J. 2013, 454, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Kehres, D.G.; Janakiraman, A.; Slauch, J.M.; Maguire, M.E. SitABCD is the alkaline Mn(2+) transporter of Salmonella enterica serovar Typhimurium. J. Bacteriol. 2002, 184, 3159–3166. [Google Scholar] [CrossRef] [Green Version]
- Sabri, M.; Léveillé, S.; Dozois, C.M. A SitABCD homologue from an avian pathogenic Escherichia coli strain mediates transport of iron and manganese and resistance to hydrogen peroxide. Microbiology 2006, 152, 745–758. [Google Scholar] [CrossRef] [Green Version]
- Janakiraman, A.; Slauch, J.M. The putative iron transport system SitABCD encoded on SPI1 is required for full virulence of Salmonella typhimurium. Mol. Microbiol. 2000, 35, 1146–1155. [Google Scholar] [CrossRef]
- Lau, C.K.; Krewulak, K.D.; Vogel, H.J. Bacterial ferrous iron transport: The Feo system. FEMS Microbiol. Rev. 2016, 40, 273–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, H.; Shin, D. The FeoA protein is necessary for the FeoB transporter to import ferrous iron. Biochem. Biophys. Res. Commun. 2012, 423, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Kammler, M.; Schön, C.; Hantke, K. Characterization of the ferrous iron uptake system of Escherichia coli. J. Bacteriol. 1993, 175, 6212–6219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Lee, H.; Shin, D. Lon-mediated proteolysis of the FeoC protein prevents Salmonella enterica from accumulating the Fe(II) transporter FeoB under high-oxygen conditions. J. Bacteriol. 2015, 197, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Vartivarian, S.E.; Cowart, R.E. Extracellular iron reductases: Identification of a new class of enzymes by siderophore-producing microorganisms. Arch. Biochem. Biophys. 1999, 364, 75–82. [Google Scholar] [CrossRef]
- Massey, V. Activation of molecular oxygen by flavins and flavoproteins. J. Biol. Chem. 1994, 269, 22459–22462. [Google Scholar] [CrossRef]
- Romero, E.; Gómez Castellanos, J.R.; Gadda, G.; Fraaije, M.W.; Mattevi, A. Same Substrate, Many Reactions: Oxygen Activation in Flavoenzymes. Chem. Rev. 2018, 118, 1742–1769. [Google Scholar] [CrossRef] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Murakami, K.; Murata, N.; Noda, Y.; Irie, K.; Shirasawa, T.; Shimizu, T. Stimulation of the amyloidogenic pathway by cytoplasmic superoxide radicals in an Alzheimer’s disease mouse model. Biosci. Biotechnol. Biochem. 2012, 76, 1098–1103. [Google Scholar] [CrossRef]
- Koskenkorva-Frank, T.S.; Weiss, G.; Koppenol, W.H.; Burckhardt, S. The complex interplay of iron metabolism, reactive oxygen species, and reactive nitrogen species: Insights into the potential of various iron therapies to induce oxidative and nitrosative stress. Free Radic. Biol. Med. 2013, 65, 1174–1194. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, T.; Omura, S.; Takahashi, Y. Generation of superoxide anions by a glycation reaction in conventional laboratory media. J. Biosci. Bioeng. 2012, 114, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Bolann, B.J.; Ulvik, R.J. Release of iron from ferritin by xanthine oxidase. Role of the superoxide radical. Biochem. J. 1987, 243, 55–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, R.F.; McCleary, C.J. Superoxide ion as a primary reductant in ascorbate-mediated ferritin iron release. Free Radic. Biol. Med. 1987, 3, 389–395. [Google Scholar] [CrossRef]
- Thomas, C.E.; Morehouse, L.A.; Aust, S.D. Ferritin and superoxide-dependent lipid peroxidation. J. Biol. Chem. 1985, 260, 3275–3280. [Google Scholar] [CrossRef]
- Thomas, C.E.; Aust, S.D. Reductive release of iron from ferritin by cation free radicals of paraquat and other bipyridyls. J. Biol. Chem. 1986, 261, 13064–13070. [Google Scholar] [CrossRef]
- Bou-Abdallah, F.; McNally, J.; Liu, X.X.; Melman, A. Oxygen catalyzed mobilization of iron from ferritin by iron(III) chelate ligands. Chem. Commun. 2011, 47, 731–733. [Google Scholar] [CrossRef]
- Claiborne, A.; Fridovich, I. Purification of the o-dianisidine peroxidase from Escherichia coli B. Physicochemical characterization and analysis of its dual catalatic and peroxidatic activities. J. Biol. Chem. 1979, 254, 4245–4252. [Google Scholar] [CrossRef]
- Bravo, J.; Verdaguer, N.; Tormo, J.; Betzel, C.; Switala, J.; Loewen, P.C.; Fita, I. Crystal structure of catalase HPII from Escherichia coli. Structure 1995, 3, 491–502. [Google Scholar] [CrossRef]
- Yost, F.J., Jr.; Fridovich, I. An iron-containing superoxide dismutase from Escherichia coli. J. Biol. Chem. 1973, 248, 4905–4908. [Google Scholar] [CrossRef]
- Gálvez, N.; Fernández, B.; Sánchez, P.; Cuesta, R.; Ceolín, M.; Clemente-León, M.; Trasobares, S.; López-Haro, M.; Calvino, J.J.; Stéphan, O.; et al. Comparative structural and chemical studies of ferritin cores with gradual removal of their iron contents. J Am Chem Soc 2008, 130, 8062–8068. [Google Scholar] [CrossRef] [PubMed]
- McKeown, S.R. Defining normoxia, physoxia and hypoxia in tumours-implications for treatment response. Br. J. Radiol. 2014, 87, 20130676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massé, E.; Gottesman, S. A small RNA regulates the expression of genes involved in iron metabolism in Escherichia coli. Proc. Natl. Acad. Sci. USA 2002, 99, 4620–4625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nairz, M.; Ferring-Appel, D.; Casarrubea, D.; Sonnweber, T.; Viatte, L.; Schroll, A.; Haschka, D.; Fang, F.C.; Hentze, M.W.; Weiss, G.; et al. Iron Regulatory Proteins Mediate Host Resistance to Salmonella Infection. Cell Host Microbe 2015, 18, 254–261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsolis, R.M.; Bäumler, A.J.; Heffron, F.; Stojiljkovic, I. Contribution of TonB- and Feo-mediated iron uptake to growth of Salmonella typhimurium in the mouse. Infect. Immun. 1996, 64, 4549–4556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyer, E.; Bergevin, I.; Malo, D.; Gros, P.; Cellier, M.F. Acquisition of Mn(II) in addition to Fe(II) is required for full virulence of Salmonella enterica serovar Typhimurium. Infect. Immun. 2002, 70, 6032–6042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patzer, S.I.; Hantke, K. Dual repression by Fe(2+)-Fur and Mn(2+)-MntR of the mntH gene, encoding an NRAMP-like Mn(2+) transporter in Escherichia coli. J. Bacteriol. 2001, 183, 4806–4813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schalk, I.J.; Guillon, L. Fate of ferrisiderophores after import across bacterial outer membranes: Different iron release strategies are observed in the cytoplasm or periplasm depending on the siderophore pathways. Amino Acids 2013, 44, 1267–1277. [Google Scholar] [CrossRef]
- Muselius, B.; Sukumaran, A.; Yeung, J.; Geddes-McAlister, J. Iron Limitation in Klebsiella pneumoniae Defines New Roles for Lon Protease in Homeostasis and Degradation by Quantitative Proteomics. Front. Microbiol. 2020, 11, 546. [Google Scholar] [CrossRef]
- Arends, J.; Griego, M.; Thomanek, N.; Lindemann, C.; Kutscher, B.; Meyer, H.E.; Narberhaus, F. An Integrated Proteomic Approach Uncovers Novel Substrates and Functions of the Lon Protease in Escherichia coli. Proteomics 2018, 18, e1800080. [Google Scholar] [CrossRef]
- Budiarti, S.R.I.; Mubarik, N.R. Extracellular Protease Activity of Enteropathogenic Escherechia coli on Mucin Substrate. HAYATI J. Biosci. 2007, 14, 36–38. [Google Scholar] [CrossRef] [Green Version]
- Grodberg, J.; Dunn, J.J. Comparison of Escherichia coli K-12 outer membrane protease OmpT and Salmonella typhimurium E protein. J. Bacteriol. 1989, 171, 2903–2905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, P.; Habouria, H.; Bessaiah, H.; Dozois, C.M. Serine Protease Autotransporters of the Enterobacteriaceae (SPATEs): Out and About and Chopping It Up. Microorganisms 2019, 7, 594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front. Cell. Infect. Microbiol. 2017, 7, 373. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, X.; Zhao, G. Ferritin Nanocage: A Versatile Nanocarrier Utilized in the Field of Food, Nutrition, and Medicine. Nanomaterials 2020, 10, 1894. [Google Scholar] [CrossRef] [PubMed]
- Berger, T.; Togawa, A.; Duncan, G.S.; Elia, A.J.; You-Ten, A.; Wakeham, A.; Fong, H.E.; Cheung, C.C.; Mak, T.W. Lipocalin 2-deficient mice exhibit increased sensitivity to Escherichia coli infection but not to ischemia-reperfusion injury. Proc. Natl. Acad. Sci. USA 2006, 103, 1834–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fritsche, G.; Nairz, M.; Libby, S.J.; Fang, F.C.; Weiss, G. Slc11a1 (Nramp1) impairs growth of Salmonella enterica serovar typhimurium in macrophages via stimulation of lipocalin-2 expression. J. Leukoc. Biol. 2012, 92, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Sulik, K.K.; Chen, S.Y. Nrf2-mediated transcriptional induction of antioxidant response in mouse embryos exposed to ethanol In Vivo: Implications for the prevention of fetal alcohol spectrum disorders. Antioxid. Redox Signal. 2008, 10, 2023–2033. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Deng, Q.; Sun, L.; Dong, K.; Li, Y.; Wu, S.; Huang, R. Salmonella effector SpvB interferes with intracellular iron homeostasis via regulation of transcription factor NRF2. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 13450–13464. [Google Scholar] [CrossRef] [Green Version]
- Nairz, M.; Schleicher, U.; Schroll, A.; Sonnweber, T.; Theurl, I.; Ludwiczek, S.; Talasz, H.; Brandacher, G.; Moser, P.L.; Muckenthaler, M.U.; et al. Nitric oxide-mediated regulation of ferroportin-1 controls macrophage iron homeostasis and immune function in Salmonella infection. J. Exp. Med. 2013, 210, 855–873. [Google Scholar] [CrossRef]
- Grubwieser, P.; Hoffmann, A.; Hilbe, R.; Seifert, M.; Sonnweber, T.; Böck, N.; Theurl, I.; Weiss, G.; Nairz, M. Airway Epithelial Cells Differentially Adapt Their Iron Metabolism to Infection With Klebsiella pneumoniae and Escherichia coli In Vitro. Front. Cell. Infect. Microbiol. 2022, 12, 875543. [Google Scholar] [CrossRef]
- Joshi, C.S.; Mora, A.; Felder, P.A.; Mysorekar, I.U. NRF2 promotes urothelial cell response to bacterial infection by regulating reactive oxygen species and RAB27B expression. Cell Rep. 2021, 37, 109856. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, T.; Yamamoto, H.; Sakamaki, Y.; Saito, C.; Mizushima, N. NCOA4 drives ferritin phase separation to facilitate macroferritinophagy and microferritinophagy. J. Cell. Biol. 2022, 221, e202203102. [Google Scholar] [CrossRef]
- Bauckman, K.A.; Mysorekar, I.U. Ferritinophagy drives uropathogenic Escherichia coli persistence in bladder epithelial cells. Autophagy 2016, 12, 850–863. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, N.; Pantopoulos, K. The IRP/IRE system in vivo: Insights from mouse models. Front. Pharmacol. 2014, 5, 176. [Google Scholar] [CrossRef] [Green Version]
- Miao, Y.; Li, G.; Zhang, X.; Xu, H.; Abraham, S.N. A TRP Channel Senses Lysosome Neutralization by Pathogens to Trigger Their Expulsion. Cell 2015, 161, 1306–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asano, T.; Komatsu, M.; Yamaguchi-Iwai, Y.; Ishikawa, F.; Mizushima, N.; Iwai, K. Distinct mechanisms of ferritin delivery to lysosomes in iron-depleted and iron-replete cells. Mol. Cell. Biol. 2011, 31, 2040–2052. [Google Scholar] [CrossRef] [Green Version]
- Liss, V.; Swart, A.L.; Kehl, A.; Hermanns, N.; Zhang, Y.; Chikkaballi, D.; Böhles, N.; Deiwick, J.; Hensel, M. Salmonella enterica Remodels the Host Cell Endosomal System for Efficient Intravacuolar Nutrition. Cell Host Microbe 2017, 21, 390–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreibich, S.; Emmenlauer, M.; Fredlund, J.; Rämö, P.; Münz, C.; Dehio, C.; Enninga, J.; Hardt, W.-D. Autophagy Proteins Promote Repair of Endosomal Membranes Damaged by the Salmonella Type Three Secretion System 1. Cell Host Microbe 2015, 18, 527–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Azad, M.B.; Gibson, S.B. Superoxide is the major reactive oxygen species regulating autophagy. Cell Death Differ. 2009, 16, 1040–1052. [Google Scholar] [CrossRef] [PubMed]
- Crouch, M.-L.V.; Castor, M.; Karlinsey, J.E.; Kalhorn, T.; Fang, F.C. Biosynthesis and IroC-dependent export of the siderophore salmochelin are essential for virulence of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2008, 67, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Dichtl, S.; Demetz, E.; Haschka, D.; Tymoszuk, P.; Petzer, V.; Nairz, M.; Seifert, M.; Hoffmann, A.; Brigo, N.; Würzner, R.; et al. Dopamine Is a Siderophore-Like Iron Chelator That Promotes Salmonella enterica Serovar Typhimurium Virulence in Mice. mBio 2019, 10, e02624-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, M.T.; Taylor, L.T.; DeLong, E.F. Quantitative analysis of small-subunit rRNA genes in mixed microbial populations via 5′-nuclease assays. Appl. Environ. Microbiol. 2000, 66, 4605–4614. [Google Scholar] [CrossRef] [PubMed]
- Tukey, J.W. (Ed.) Exploratory Data Analysis; Addision-Wesley Publishing Company: Boston, MA, USA, 1977; pp. 43–44. [Google Scholar]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gehrer, C.M.; Hoffmann, A.; Hilbe, R.; Grubwieser, P.; Mitterstiller, A.-M.; Talasz, H.; Fang, F.C.; Meyron-Holtz, E.G.; Atkinson, S.H.; Weiss, G.; et al. Availability of Ferritin-Bound Iron to Enterobacteriaceae. Int. J. Mol. Sci. 2022, 23, 13087. https://doi.org/10.3390/ijms232113087
Gehrer CM, Hoffmann A, Hilbe R, Grubwieser P, Mitterstiller A-M, Talasz H, Fang FC, Meyron-Holtz EG, Atkinson SH, Weiss G, et al. Availability of Ferritin-Bound Iron to Enterobacteriaceae. International Journal of Molecular Sciences. 2022; 23(21):13087. https://doi.org/10.3390/ijms232113087
Chicago/Turabian StyleGehrer, Clemens M., Alexander Hoffmann, Richard Hilbe, Philipp Grubwieser, Anna-Maria Mitterstiller, Heribert Talasz, Ferric C. Fang, Esther G. Meyron-Holtz, Sarah H. Atkinson, Günter Weiss, and et al. 2022. "Availability of Ferritin-Bound Iron to Enterobacteriaceae" International Journal of Molecular Sciences 23, no. 21: 13087. https://doi.org/10.3390/ijms232113087