
Enhanced Succinate Oxidation with Mitochondrial Complex II Reactive Oxygen Species Generation in Human Prostate Cancer
 ,
,
Abstract
:1. Introduction
2. Results
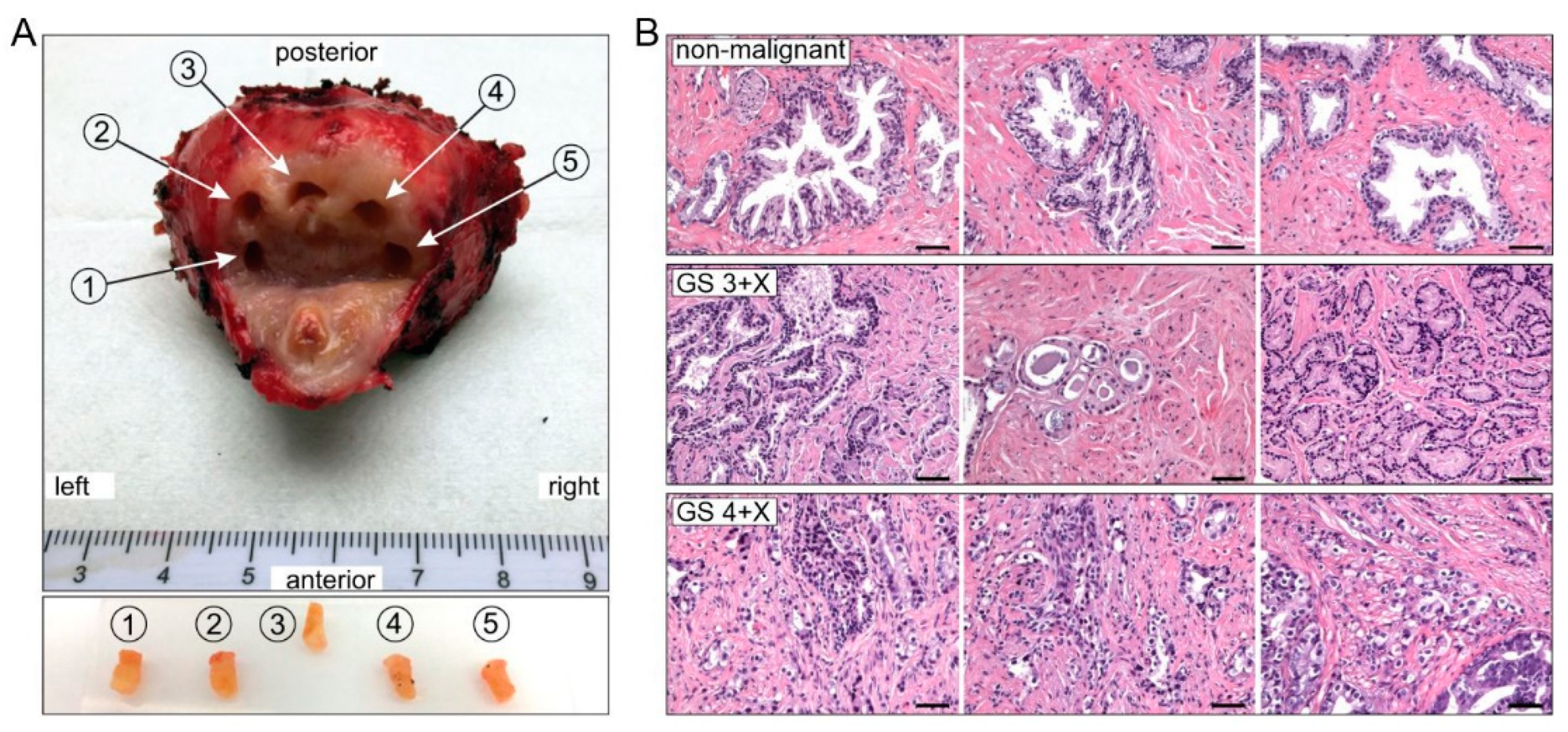
2.1. Patient Demographics and Biopsy Pathology
2.2. The Oxidative Phosphorylation (OXPHOS) Rate of CII (PCII) Is Higher Than That of CI (PCI) in Non-Malignant Tissue
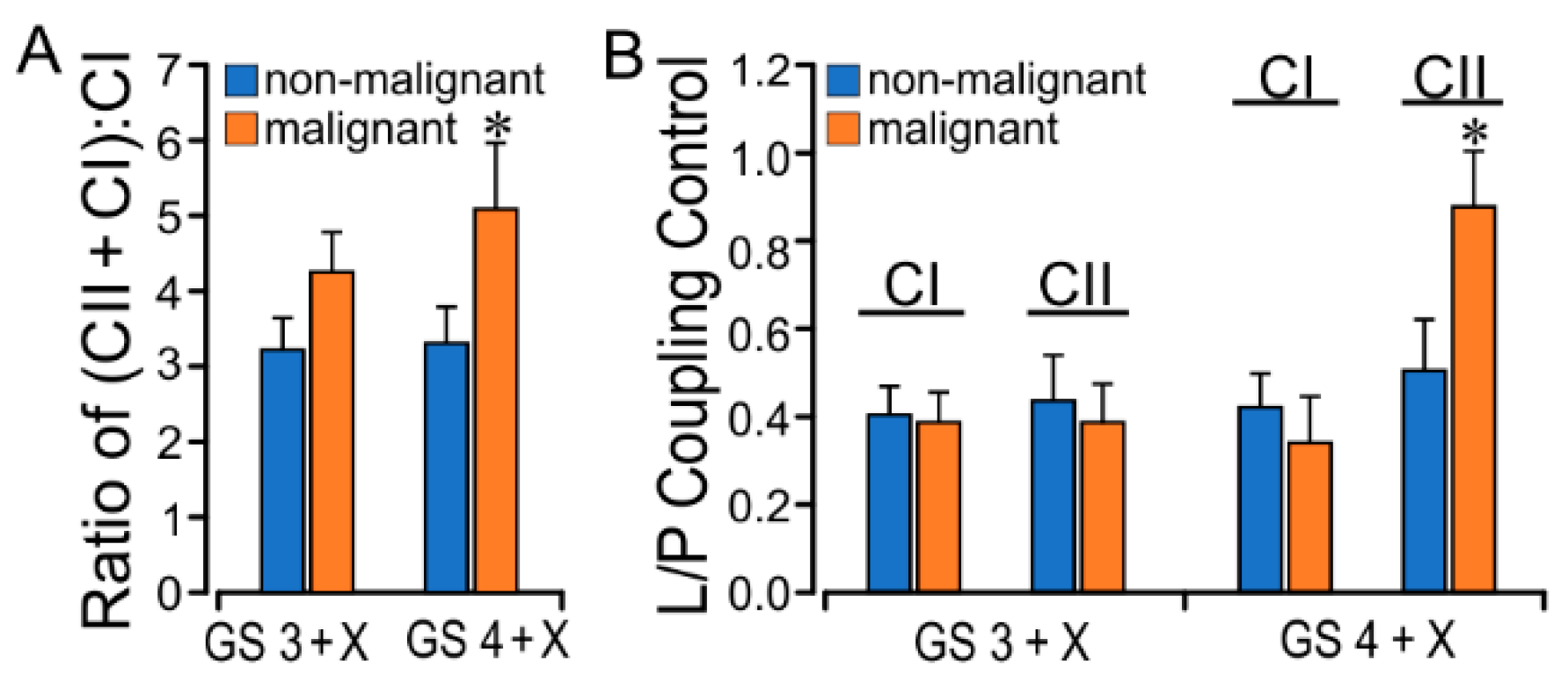
2.3. PCa GS Grade Enhances CII Flux with Increased L/P Coupling Ratio
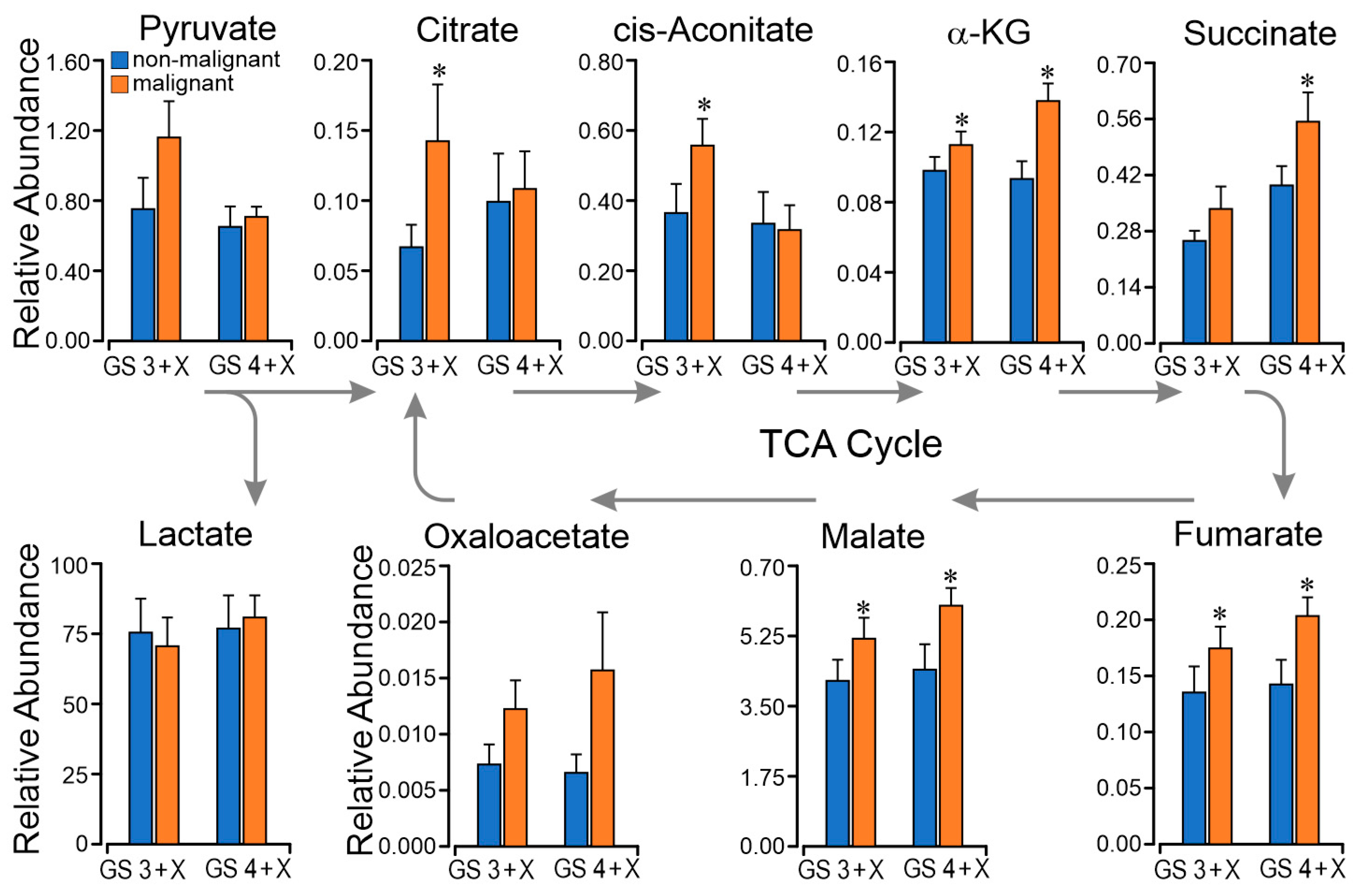
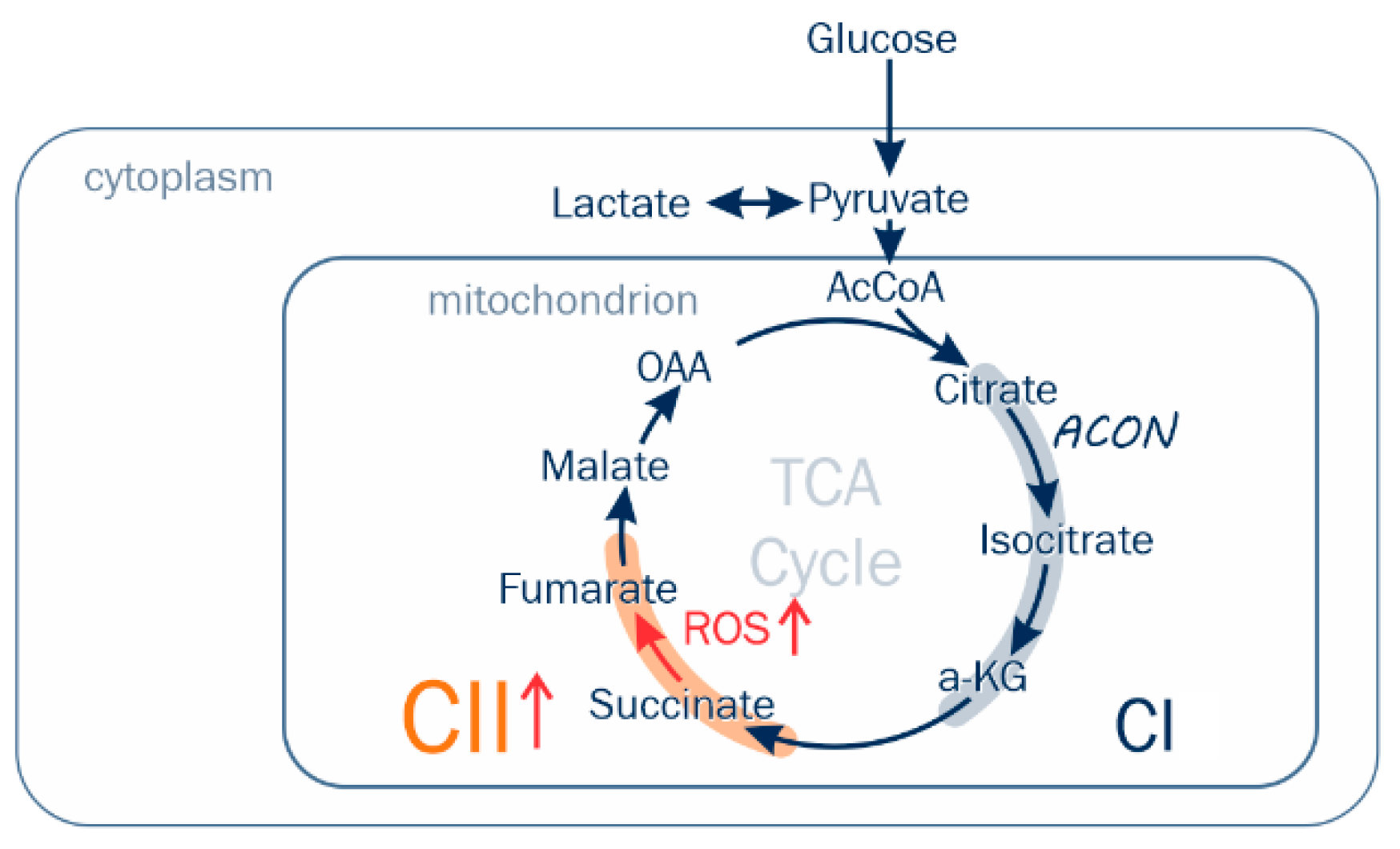
2.4. TCA Metabolomics Support Respiratory Finding of Succinate CII Dominant Flux
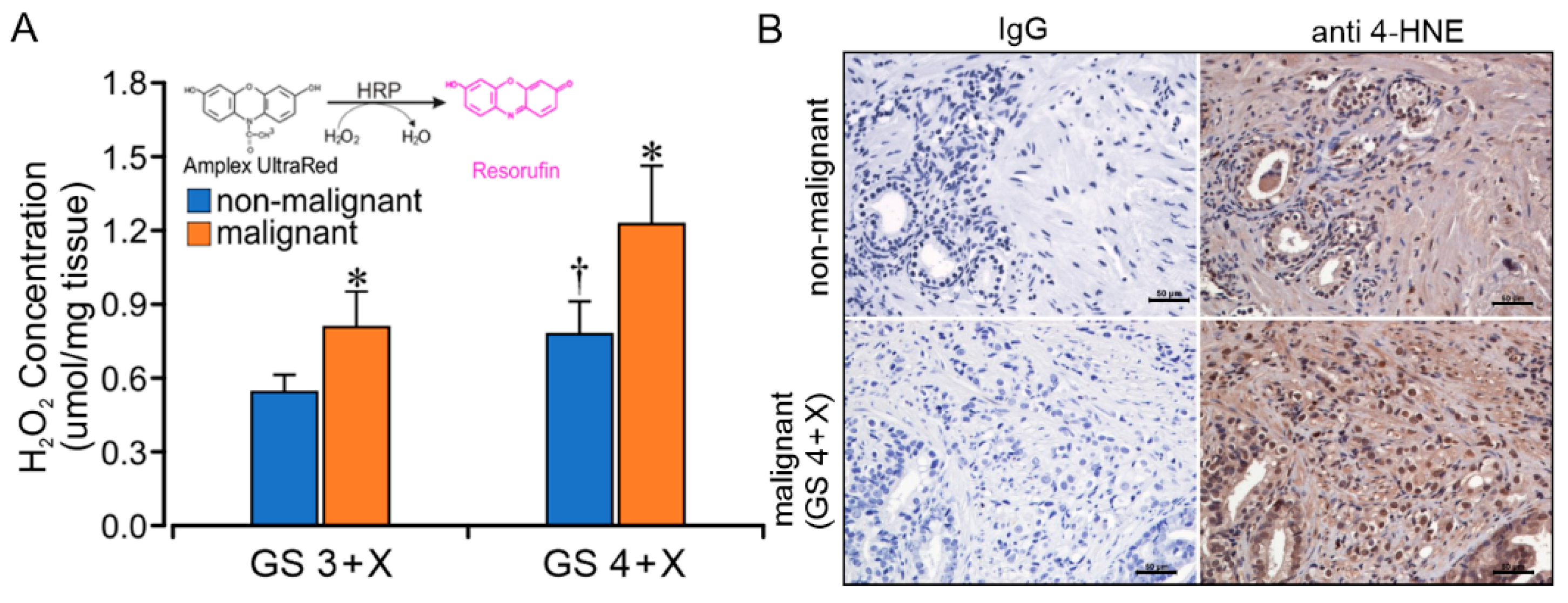
2.5. GS Grade Is Related to CII Forward ROS Production Rate
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Tissue Procurement and Processing
4.3. Histopathology and Immunohistochemistry
4.4. Respirometry Protocols
4.5. ROS Analysis
4.6. Targeted Metabolomics
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cronin, K.A.; Lake, A.J.; Scott, S.; Sherman, R.L.; Noone, A.M.; Howlader, N.; Henley, S.J.; Anderson, R.N.; Firth, A.U.; Ma, J.; et al. Annual Report to the Nation on the Status of Cancer, part I: National cancer statistics. Cancer 2018, 124, 2785–2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, A.B.; Matulewicz, R.S.; Eggener, S.E.; Schaeffer, E.M. Increasing incidence of metastatic prostate cancer in the United States (2004–2013). Prostate Cancer Prostatic Dis. 2016, 19, 395–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosoian, J.J.; Carter, H.B.; Lepor, A.; Loeb, S. Active surveillance for prostate cancer: Current evidence and contemporary state of practice. Nat. Rev. Urol. 2016, 13, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Epstein, J.I.; Walsh, P.C.; Carter, H.B. Dedifferentiation of prostate cancer grade with time in men followed expectantly for stage T1c disease. J. Urol. 2001, 166, 1688–1691. [Google Scholar] [CrossRef]
- Huggins, C. The prostate secretion. Harvey Lect. 1946, 52, 148–193. [Google Scholar]
- Cooper, J.F.; Imfeld, H. The role of citric acid in the physiology of the prostate: A preliminary report. J. Urol. 1959, 81, 157–164. [Google Scholar] [CrossRef]
- Costello, L.C.; Franklin, R.B. The clinical relevance of the metabolism of prostate cancer; zinc and tumor suppression: Connecting the dots. Mol. Cancer 2006, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Costello, L.C.; Franklin, R.B. Aconitase activity, citrate oxidation, and zinc inhibition in rat ventral prostate. Enzyme 1981, 26, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.K.; Desouki, M.M.; Franklin, R.B.; Costello, L.C. Mitochondrial aconitase and citrate metabolism in malignant and nonmalignant human prostate tissues. Mol. Cancer 2006, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Dakubo, G.D.; Parr, R.L.; Costello, L.C.; Franklin, R.B.; Thayer, R.E. Altered metabolism and mitochondrial genome in prostate cancer. J. Clin. Pathol. 2006, 59, 10–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, R.B.; Milon, B.; Feng, P.; Costello, L.C. Zinc and zinc transporters in normal prostate and the pathogenesis of prostate cancer. Front. Biosci. 2005, 10, 2230–2239. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Gokden, N.; Greene, G.F.; Mukunyadzi, P.; Kadlubar, F.F. Extensive somatic mitochondrial mutations in primary prostate cancer using laser capture microdissection. Cancer Res. 2002, 62, 6470–6474. [Google Scholar] [PubMed]
- Gomez-Zaera, M.; Abril, J.; Gonzalez, L.; Aguilo, F.; Condom, E.; Nadal, M.; Nunes, V. Identification of somatic and germline mitochondrial DNA sequence variants in prostate cancer patients. Mutat. Res. 2006, 595, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Petros, J.A.; Baumann, A.K.; Ruiz-Pesini, E.; Amin, M.B.; Sun, C.Q.; Hall, J.; Lim, S.; Issa, M.M.; Flanders, W.D.; Hosseini, S.H.; et al. mtDNA mutations increase tumorigenicity in prostate cancer. Proc. Natl. Acad. Sci. USA 2005, 102, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Kloss-Brandstatter, A.; Schafer, G.; Erhart, G.; Huttenhofer, A.; Coassin, S.; Seifarth, C.; Summerer, M.; Bektic, J.; Klocker, H.; Kronenberg, F. Somatic mutations throughout the entire mitochondrial genome are associated with elevated PSA levels in prostate cancer patients. Am. J. Hum. Genet. 2010, 87, 802–812. [Google Scholar] [CrossRef] [Green Version]
- Hopkins, J.F.; Sabelnykova, V.Y.; Weischenfeldt, J.; Simon, R.; Aguiar, J.A.; Alkallas, R.; Heisler, L.E.; Zhang, J.; Watson, J.D.; Chua, M.L.K.; et al. Mitochondrial mutations drive prostate cancer aggression. Nat. Commun. 2017, 8, 656. [Google Scholar] [CrossRef]
- Mehra, N.; Penning, M.; Maas, J.; van Daal, N.; Giles, R.H.; Voest, E.E. Circulating mitochondrial nucleic acids have prognostic value for survival in patients with advanced prostate cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2007, 13, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Tu, H.; Gu, J.; Meng, Q.H.; Kim, J.; Davis, J.W.; He, Y.; Wagar, E.A.; Thompson, T.C.; Logothetis, C.J.; Wu, X. Mitochondrial DNA copy number in peripheral blood leukocytes and the aggressiveness of localized prostate cancer. Oncotarget 2015, 6, 41988–41996. [Google Scholar] [CrossRef] [Green Version]
- Philley, J.V.; Kannan, A.; Qin, W.; Sauter, E.R.; Ikebe, M.; Hertweck, K.L.; Troyer, D.A.; Semmes, O.J.; Dasgupta, S. Complex-I Alteration and Enhanced Mitochondrial Fusion Are Associated With Prostate Cancer Progression. J. Cell. Physiol. 2016, 231, 1364–1374. [Google Scholar] [CrossRef] [Green Version]
- Weber, A.; Klocker, H.; Oberacher, H.; Gnaiger, E.; Neuwirt, H.; Sampson, N.; Eder, I.E. Succinate Accumulation Is Associated with a Shift of Mitochondrial Respiratory Control and HIF-1alpha Upregulation in PTEN Negative Prostate Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2129. [Google Scholar] [CrossRef] [Green Version]
- Dueregger, A.; Schopf, B.; Eder, T.; Hofer, J.; Gnaiger, E.; Aufinger, A.; Kenner, L.; Perktold, B.; Ramoner, R.; Klocker, H.; et al. Differential Utilization of Dietary Fatty Acids in Benign and Malignant Cells of the Prostate. PLoS ONE 2015, 10, e0135704. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.D.; Sun, C.; Lambeth, J.D.; Marshall, F.; Amin, M.; Chung, L.; Petros, J.A.; Arnold, R.S. Increased Nox1 and hydrogen peroxide in prostate cancer. Prostate 2005, 62, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Len, J.S.; Koh, W.S.D.; Tan, S.X. The roles of reactive oxygen species and antioxidants in cryopreservation. Biosci Rep. 2019, 39, BSR20191601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grupp, K.; Jedrzejewska, K.; Tsourlakis, M.C.; Koop, C.; Wilczak, W.; Adam, M.; Quaas, A.; Sauter, G.; Simon, R.; Izbicki, J.R.; et al. High mitochondria content is associated with prostate cancer disease progression. Mol. Cancer 2013, 12, 145. [Google Scholar] [CrossRef] [Green Version]
- Rosario, E.; Rosario, D.J. Localized Prostate Cancer; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Cheng, L.; MacLennan, G.T.; Lopez-Beltran, A.; Montironi, R. Anatomic, morphologic and genetic heterogeneity of prostate cancer: Implications for clinical practice. Expert Rev. Anticancer Ther. 2012, 12, 1371–1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schopf, B.; Schafer, G.; Weber, A.; Talasz, H.; Eder, I.E.; Klocker, H.; Gnaiger, E. Oxidative phosphorylation and mitochondrial function differ between human prostate tissue and cultured cells. FEBS J. 2016, 283, 2181–2196. [Google Scholar] [CrossRef] [PubMed]
- Dudka, I.; Thysell, E.; Lundquist, K.; Antti, H.; Iglesias-Gato, D.; Flores-Morales, A.; Bergh, A.; Wikstrom, P.; Grobner, G. Comprehensive metabolomics analysis of prostate cancer tissue in relation to tumor aggressiveness and TMPRSS2-ERG fusion status. BMC Cancer 2020, 20, 437. [Google Scholar] [CrossRef]
- Schopf, B.; Weissensteiner, H.; Schafer, G.; Fazzini, F.; Charoentong, P.; Naschberger, A.; Rupp, B.; Fendt, L.; Bukur, V.; Giese, I.; et al. OXPHOS remodeling in high-grade prostate cancer involves mtDNA mutations and increased succinate oxidation. Nat. Commun. 2020, 11, 1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaikhibrahim, Z.; Lindstrot, A.; Ellinger, J.; Rogenhofer, S.; Buettner, R.; Perner, S.; Wernert, N. The peripheral zone of the prostate is more prone to tumor development than the transitional zone: Is the ETS family the key? Mol. Med. Rep. 2012, 5, 313–316. [Google Scholar]
- Cardoso, H.J.; Carvalho, T.M.A.; Fonseca, L.R.S.; Figueira, M.I.; Vaz, C.V.; Socorro, S. Revisiting prostate cancer metabolism: From metabolites to disease and therapy. Med. Res. Rev. 2021, 41, 1499–1538. [Google Scholar] [CrossRef]
- Chen, C.L.; Lin, C.Y.; Kung, H.J. Targeting Mitochondrial OXPHOS and Their Regulatory Signals in Prostate Cancers. Int. J. Mol. Sci. 2021, 22, 13435. [Google Scholar] [CrossRef]
- Nogueira, V.; Rigoulet, M.; Piquet, M.A.; Devin, A.; Fontaine, E.; Leverve, X.M. Mitochondrial respiratory chain adjustment to cellular energy demand. J. Biol. Chem. 2001, 276, 46104–46110. [Google Scholar] [CrossRef]
- Wojtczak, L.; Bogucka, K.; Duszynski, J.; Zablocka, B.; Zolkiewska, A. Regulation of mitochondrial resting state respiration: Slip, leak, heterogeneity? Biochim. Et Biophys. Acta 1990, 1018, 177–181. [Google Scholar] [CrossRef]
- Quinlan, C.L.; Orr, A.L.; Perevoshchikova, I.V.; Treberg, J.R.; Ackrell, B.A.; Brand, M.D. Mitochondrial complex II can generate reactive oxygen species at high rates in both the forward and reverse reactions. J. Biol. Chem. 2012, 287, 27255–27264. [Google Scholar] [CrossRef] [Green Version]
- Grivennikova, V.G.; Kozlovsky, V.S.; Vinogradov, A.D. Respiratory complex II: ROS production and the kinetics of ubiquinone reduction. Biochim. Biophys Acta Bioenerg 2017, 1858, 109–117. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijevic, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef] [Green Version]
- Scialo, F.; Fernandez-Ayala, D.J.; Sanz, A. Role of Mitochondrial Reverse Electron Transport in ROS Signaling: Potential Roles in Health and Disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Sanchez, R.; Hernandez-Esquivel, L.; Rivero-Segura, N.A.; Marin-Hernandez, A.; Neuzil, J.; Ralph, S.J.; Rodriguez-Enriquez, S. Reactive oxygen species are generated by the respiratory complex II--evidence for lack of contribution of the reverse electron flow in complex I. FEBS J. 2013, 280, 927–938. [Google Scholar] [CrossRef]
- Shen, K.; Liu, B.; Zhou, X.; Ji, Y.; Chen, L.; Wang, Q.; Xue, W. The Evolving Role of (18)F-FDG PET/CT in Diagnosis and Prognosis Prediction in Progressive Prostate Cancer. Front. Oncol. 2021, 11, 683793. [Google Scholar] [CrossRef]
- Usenik, A.; Legisa, M. Evolution of allosteric citrate binding sites on 6-phosphofructo-1-kinase. PLoS ONE 2010, 5, e15447. [Google Scholar] [CrossRef]
- Dalla Pozza, E.; Dando, I.; Pacchiana, R.; Liboi, E.; Scupoli, M.T.; Donadelli, M.; Palmieri, M. Regulation of succinate dehydrogenase and role of succinate in cancer. Semin. Cell Dev. Biol. 2020, 98, 4–14. [Google Scholar] [CrossRef]
- Ahmad, F.; Cherukuri, M.K.; Choyke, P.L. Metabolic reprogramming in prostate cancer. Br. J. Cancer 2021, 125, 1185–1196. [Google Scholar] [CrossRef]
- Xin, M.; Qiao, Z.; Li, J.; Liu, J.; Song, S.; Zhao, X.; Miao, P.; Tang, T.; Wang, L.; Liu, W.; et al. miR-22 inhibits tumor growth and metastasis by targeting ATP citrate lyase: Evidence in osteosarcoma, prostate cancer, cervical cancer and lung cancer. Oncotarget 2016, 7, 44252–44265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, N.; Swinnen, J.V.; Smans, K. ATP-citrate lyase: A key player in cancer metabolism. Cancer Res. 2012, 72, 3709–3714. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, D.J.; Zhang, A.; Li, S.; Cao, T.N.; Smith, J.A.; Vedula, I.; Cordero-Reyes, A.M.; Youker, K.A.; Torre-Amione, G.; Gupte, A.A. Combination of angiotensin II and l-NG-nitroarginine methyl ester exacerbates mitochondrial dysfunction and oxidative stress to cause heart failure. Am. J. Physiology Heart Circ. Physiol. 2016, 310, H667-80. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Respiratory State (pmol/(s·mg)) | Biopsy Paired Source | |||
|---|---|---|---|---|
| GS 3 + X (n = 9) | GS 4 + X (n = 6) | |||
| Non-Malignant | Malignant | Non-Malignant | Malignant | |
| LEAK (L) | ||||
| CI (LCI) | 2.14 ± 0.32 | 2.02 ± 0.52 | 2.09 ± 0.34 | 2.16 ± 0.98 |
| CII (LCII) | 4.61 ± 0.79 | 5.88 ± 1.43 | 5.27 ± 0.85 | 8.70 ± 2.58 * |
| CI + CII (LCI+CII) | ||||
| OXPHOS (P) | ||||
| CI (PCI) | 5.49 ± 0.82 (33.19%) 1 | 5.14 ± 0.90 (24.68%) | 5.06 ± 0.62 (32.29%) | 5.34 ± 0.75 (25.05%) |
| CII (PCII) | 11.26 ± 1.31 (68.08%) | 12.43 ± 1.31 (59.67%) | 10.79 ± 1.74 (68.86%) | 13.48 ± 2.85 (63.23%) |
| CI + CII (PCI+CII) | 16.54 ± 1.31 | 20.83 ± 3.79 | 15.67 ± 1.61 | 21.32 ± 2.25 * |
| ETS (E) | ||||
| CI (ECI) | ||||
| CII (ECII) | 11.98 ± 1.15 | 13.21 ± 1.43 | 11.86 ± 1.38 | 17.44 ± 3.14 |
| CI + CII (ECI+CII) | 19.07 ± 1.40 | 23.72 ± 4.19 | 17.81 ± 1.38 | 23.98 ± 2.31 * |
| CI + CII Protocol | CII + Rotenone Protocol | ||||
|---|---|---|---|---|---|
| Injection | Abbreviation | Concentration | Injection | Abbreviation | Concentration |
| glutamate | G | 10 mM | rotenone | R | 0.5 µM |
| malate | M | 2 mM | succinate | S | 10 mM |
| ADP | D | 2.5 mM | ADP | D | 2.5 mM |
| cytochrome c | Cyt C | 10 µM | cytochrome c | Cyt C | 10 µM |
| pyruvate | P | 5 mM | FCCP | F | 1.5 µM |
| succinate | S | 10 mM | malonate | Mna | 5 mM |
| FCCP | F | 1.5 µM | Antimycin A | AA | 2.5 µM |
| rotenone | R | 0.5 µM | |||
| malonate | Mna | 5 mM | |||
| Antimycin A | AA | 2.5 µM | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.; Gupte, A.A.; Chatterjee, S.; Li, S.; Ayala, A.G.; Miles, B.J.; Hamilton, D.J. Enhanced Succinate Oxidation with Mitochondrial Complex II Reactive Oxygen Species Generation in Human Prostate Cancer. Int. J. Mol. Sci. 2022, 23, 12168. https://doi.org/10.3390/ijms232012168
Zhang A, Gupte AA, Chatterjee S, Li S, Ayala AG, Miles BJ, Hamilton DJ. Enhanced Succinate Oxidation with Mitochondrial Complex II Reactive Oxygen Species Generation in Human Prostate Cancer. International Journal of Molecular Sciences. 2022; 23(20):12168. https://doi.org/10.3390/ijms232012168
Chicago/Turabian StyleZhang, Aijun, Anisha A. Gupte, Somik Chatterjee, Shumin Li, Alberto G. Ayala, Brian J. Miles, and Dale J. Hamilton. 2022. "Enhanced Succinate Oxidation with Mitochondrial Complex II Reactive Oxygen Species Generation in Human Prostate Cancer" International Journal of Molecular Sciences 23, no. 20: 12168. https://doi.org/10.3390/ijms232012168