Molecular Characterization of Vitellogenin and Its Receptor in Spodoptera frugiperda (J. E. Smith, 1797), and Their Function in Reproduction of Female

Abstract
:1. Introduction
2. Results
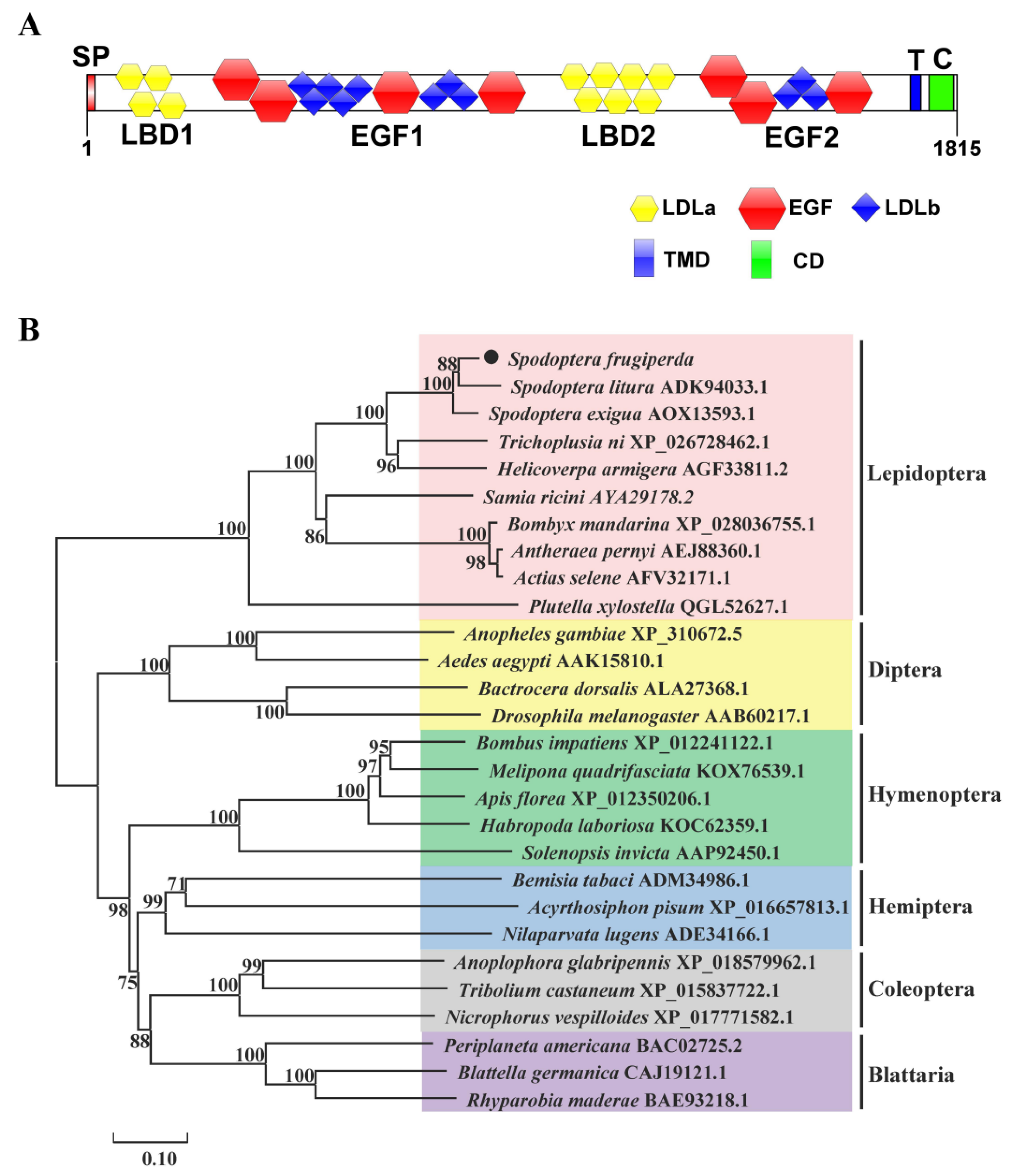
2.1. Cloning and Sequence Analysis of SfVg and SfVgR
2.2. Expression Profiling of SfVg and SfVgR
2.3. Functional Analysis of SfVg and SfVgR by RNA Interference Experiment
3. Discussion
4. Materials and Methods
4.1. Insect Collection and Rearing
4.2. RNA Isolation and cDNA Synthesis
4.3. Molecular Cloning of SfVg and SfVgR
4.4. Bioinformatics Analysis of SfVg and SfVgR
4.5. Expression Profiling Analysis of SfVg and SfVgR
4.6. Functional Verification of SfVg and SfVgR by RNA Interference Experiment
4.7. Western Blot Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roy, S.; Saha, T.T.; Zou, Z.; Raikhel, A.S. Regulatory pathways controlling female insect reproduction. Annu. Rev. Entomol. 2018, 63, 489–511. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, L.; He, Q.; Zhou, S. Regulatory mechanisms of vitellogenesis in insects. Front. Cell Dev. Biol. 2020, 8, 593613. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Tian, P.; Tang, Y.; Yang, L.; Qiu, L.; He, H.; Ding, W.; Li, Z.; Li, Y. Molecular characterization of vitellogenin and its receptor in Sogatella furcifera, and their function in oocyte maturation. Front. Physiol. 2019, 10, 1532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef]
- Tufail, M.; Takeda, M. Insect vitellogenin/lipophorin receptors: Molecular structures, role in oogenesis, and regulatory mechanisms. J. Insect Physiol. 2009, 55, 87–103. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Y.; Liu, Y.; Zhao, M.; Jin, J.; Zhou, Z.; Guo, J. Identification and expression patterns of three vitellogenin genes and their roles in reproduction of the alligatorweed flea beetle Agasicles hygrophila (Coleoptera: Chrysomelidae). Front. Physiol. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.R.; Ko, Y.G.; Mayer, R.T. Purification, characterization, and synthesis of vitellin from the cabbage butterfly, Pieris rapae L. Arch. Insect Biochem. Physiol. 1988, 9, 67–79. [Google Scholar] [CrossRef]
- Guo, J.; Wang, L.; Wu, H.; Cao, Y.; Xiao, R.; Lai, X.; Liu, J.; Yi, J.; Zhang, G. Molecular characterization and expression of vitellogenin genes from the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). Physiol. Entomol. 2018, 43, 295–305. [Google Scholar] [CrossRef]
- Husain, M.; Rasool, K.G.; Tufail, M.; Aldawood, A.S. Molecular characterization, expression pattern, and RNAi ediated silencing of vitellogenin receptor gene in almond moth, Cadra cautella. Insect Mol. Biol. 2020, 29, 417–430. [Google Scholar] [CrossRef]
- Miao, L.U.; Zhang, N.; Jiang, H.; Dong, F.; Wang, J.U. Molecular characterization and functional analysis of the vitellogenin receptor in the rice stem borer, Chilo suppressalis. Arch. Insect Biochem. Physiol. 2019, 103, e21636. [Google Scholar] [CrossRef]
- Tufail, M.; Elmogy, M.; Fouda, M.M.A.; Elgendy, A.M.; Bembenek, J.; Trang, L.T.D.; Shao, Q.; Takeda, M. Molecular cloning, characterization, expression pattern and cellular distribution of an ovarian lipophorin receptor in the cockroach, Leucophaea maderae. Insect Mol. Biol. 2010, 18, 281–294. [Google Scholar] [CrossRef]
- Pan, M.L.; Bell, W.J.; Telfer, W.H. Vitellogenic blood protein synthesis by insect fat body. Science 1969, 165, 393–394. [Google Scholar] [CrossRef]
- Ferenz, H. Yolk protein accumulation in Locusta migratoria (R. & F.) (Orthoptera: Acrididae) oocytes. Int. J. Insect Morphol. 1993, 22, 295–314. [Google Scholar]
- Jiang, J.; Zhang, Y.; Ma, L.; Niu, T.; Dong, T.; Sheng, R.; Li, L.; Xu, Y.; Xi, L.; Li, G. Molecular characterization of Neoseiulus barkeri vitellogenin genes and vitellogenin receptor during reproductive diapause. Insects 2020, 11, 203. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Jiang, F.; Guo, J.; Yi, J.; Liu, J.; Cao, Y.; Lai, X.; Zhang, G. Molecular characterization and expression of vitellogenin and vitellogenin receptor of Thitarodes pui (Lepidoptera: Hepialidae), an insect on the tibetan plateau. J. Insect Sci. 2018, 18, 23. [Google Scholar] [CrossRef]
- Yao, Q.; Xu, S.; Dong, Y.; Que, Y.; Chen, B. Characterization of vitellogenin and vitellogenin receptor of Conopomorpha sinensis Bradley and their responses to sublethal concentrations of insecticide. Front. Physiol. 2018, 9, 1250. [Google Scholar] [CrossRef]
- Garcia, J.; Munro, E.S.; Monte, M.M.; Fourrier, M.C.S.; Whitelaw, J.; Smail, D.A.; Ellis, A.E. Atlantic salmon (Salmo salar L.) serum vitellogenin neutralises infectivity of infectious pancreatic necrosis virus (IPNV). Fish Shellfish Immunol. 2010, 29, 293–297. [Google Scholar] [CrossRef]
- Salmela, H.; Amdam, G.V.; Freitak, D. Transfer of immunity from mother to offspring is mediated via egg-yolk protein vitellogenin. PLoS Pathog. 2015, 11, e1005015. [Google Scholar] [CrossRef] [Green Version]
- Amdam, G.V.; Norberg, K.; Hagen, A.; Omholt, S.W. Social exploitation of vitellogenin. Proc. Natl. Acad. Sci. USA 2003, 100, 1799–1802. [Google Scholar] [CrossRef] [Green Version]
- Huo, Y.; Yu, Y.; Chen, L.; Li, Q.; Zhang, M.; Song, Z.; Chen, X.; Fang, R.; Zhang, L. Insect tissue-specific vitellogenin facilitates transmission of plant virus. PLoS Pathog. 2018, 14, e1006909. [Google Scholar] [CrossRef] [PubMed]
- Todd, E.L.; Poole, R.W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the western hemisphere. Ann. Entomol. Soc. Am. 1980, 6, 722–738. [Google Scholar] [CrossRef]
- Wan, J.; Huang, C.; LI, C.Y.; Zhou, H.X.; Wan, F.H. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agr. 2021, 20, 646–663. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Hunt, T. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.Y.; Zhang, Y.Y.; Zhou, X.Y.; Hong, X.Y.; Chen, L. Population genetics reveal multiple independent invasions of Spodoptera frugiperda (Lepidoptera: Noctuidae) in China. Bull. Entomol. Res. 2022, 1–11. [Google Scholar] [CrossRef]
- Tepa-Yotto, G.T.; Chinwada, P.; Rwomushana, I.; Goergen, G.; Subramanian, S. Integrated management of Spodoptera frugiperda six years post-detection in Africa: A review. Curr. Opin. Insect Sci. 2022, 52, 100928. [Google Scholar] [CrossRef]
- Tufail, M.; Naeemullah, M.; Elmogy, M.; Sharma, P.N.; Takeda, M.; Nakamura, C. Molecular cloning, transcriptional regulation, and differential expression profiling of vitellogenin in two wing-morphs of the brown planthopper, Nilaparvata lugens Stl (Hemiptera: Delphacidae). Insect Mol. Biol. 2010, 19, 787–798. [Google Scholar] [CrossRef]
- Lee, K.Y.; Yoon, H.J.; Jin, B.R. Osmia cornifrons vitellogenin: cDNA cloning, structural analysis and developmental expression. Entomol. Res. 2015, 45, 94–101. [Google Scholar] [CrossRef]
- Giorgi, F.; Bradley, J.T.; Nordin, J.H. Differential vitellin polypeptide processing in insect embryos. Micron 1999, 30, 579–596. [Google Scholar] [CrossRef]
- Tufail, M.; Lee, J.M.; Hatakeyama, M.; Oishi, K.; Takeda, M. Cloning of vitellogenin cDNA of the American cockroach, Periplaneta americana (Dictyoptera), and its structural and expression analyses. Arch. Insect Biochem. Physiol. 2000, 45, 37–46. [Google Scholar] [CrossRef]
- Liang, C.; Liu, T.H.; Han, S.P.; He, Y.Z. Molecular cloning, expression profiling and RNA interference of a vitellogenin gene from Harmonia axyridis (Coleoptera: Coccinellidae). J. Integr. Agric. 2019, 18, 133–142. [Google Scholar] [CrossRef]
- Liu, C.; Mao, J.; Zeng, F. Chrysopa septempunctata (Neuroptera: Chrysopidae) vitellogenin functions through effects on egg production and hatching. J. Econ. Entomol. 2015, 108, 2779–2788. [Google Scholar] [CrossRef] [PubMed]
- Schonbaum, C.P.; Lee, S.; Mahowald, A.P. The Drosophila yolkless gene encodes a vitellogenin receptor belonging to the low density lipoprotein receptor superfamily. Proc. Natl. Acad. Sci. USA 1995, 92, 1485–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.M.; Lu, T.Y.; Wu, Y.; Yu, N.; Xu, G.M.; Han, Q.Q.; Liu, Z.W. The importance of vitellogenin receptors in the oviposition of the pond wolf spider, Pardosa pseudoannulata. Insect Sci. 2022, 29, 443–452. [Google Scholar] [CrossRef]
- Shu, Y.H.; Wang, J.W.; Lu, K.; Zhou, J.L.; Zhou, Q.; Zhang, G.R. The first vitellogenin receptor from a Lepidopteran insect: Molecular characterization, expression patterns and RNA interference analysis. Insect Mol. Biol. 2010, 20, 61–73. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Xiao, H.; Xie, B.; Smagghe, G.; Guo, Y.; Liang, G. Molecular Characterization and function analysis of the vitellogenin receptor from the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera, Noctuidae). PLoS ONE 2016, 11, e155785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Sun, Y.; Xiao, L.; Tan, Y.; Jiang, Y.; Bai, L. Vitellogenin and vitellogenin receptor gene expression profiles in Spodoptera exigua are related to host plant suitability. Pest Manag. Sci. 2018, 74, 950–958. [Google Scholar] [CrossRef]
- Havukainen, H.; Underhaug, J.; Wolschin, F.; Amdam, G.; Halskau, O. A vitellogenin polyserine cleavage site: Highly disordered conformation protected from proteolysis by phosphorylation. J. Exp. Biol. 2012, 215, 1837–1846. [Google Scholar] [CrossRef] [Green Version]
- Jing, Y.; Wen, X.; Li, L.; Zhang, S.; Zhang, C.; Zhou, S. The vitellogenin receptor functionality of the migratory locust depends on its phosphorylation by juvenile hormone. Proc. Natl. Acad. Sci. USA 2021, 118, e2106908118. [Google Scholar] [CrossRef] [PubMed]
- Roy-Zokan, E.M.; Cunningham, C.B.; Hebb, L.E.; Mckinney, E.C.; Moor, A.J. Vitellogenin and vitellogenin receptor gene expression is associated with male and female parenting in a subsocial insect. Proc. Biol. Sci. 2015, 282, 20150787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piulachs, M.D.; Guidugli, K.R.; Barchuk, A.R.; Cruz, J.; Simões, Z.L.P.; Bellés, X. The vitellogenin of the honey bee, Apis mellifera: Structural analysis of the cDNA and expression studies. Insect Biochem. Molec. Biol. 2003, 33, 459–465. [Google Scholar] [CrossRef]
- Shang, F.; Niu, J.; Ding, B.; Zhang, Q.; Ye, C.; Zhang, W.; Smagghe, G.; Wang, J. Vitellogenin and its receptor play essential roles in the development and reproduction of the brown citrus aphid, Aphis (Toxoptera) citricidus. Insect Mol. Biol. 2017, 27, 221–233. [Google Scholar] [CrossRef]
- Shu, Y.; Zhou, J.; Tang, W.; Lu, K.; Qiang, Z.; Zhang, G. Molecular characterization and expression pattern of Spodoptera litura (Lepidoptera: Noctuidae) vitellogenin, and its response to lead stress. J. Insect Physiol. 2009, 55, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Han, S.; Qin, Q.; Chen, J.; Wang, D.; He, Y. Molecular identification and functional characterization of vitellogenin receptor from Harmonia axyridis (Coleoptera: Coccinellidae). J. Econ. Entomol. 2022, 115, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.A.; Bogaert, T.; Clinton, W.; Heck, G.R.; Feldmann, P.; Ilagan, O.; Johnson, S.; Plaetinck, G.; Munyikwa, T.; Pleau, M.; et al. Control of coleopteran insect pests through RNA interference. Nat. Biotechnol. 2007, 25, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Sharleeza, M.; Yasmin, O.R.; Ann, H.J. Global trends in research and commercialization of exogenous and endogenous RNAi technologies for crops. Crit. Rev. Biotechnol. 2018, 39, 67–78. [Google Scholar]
- Xu, J.; Wang, X.; Chen, P.; Liu, F.; Zheng, S.; Ye, H.; Mo, M. RNA interference in moths: Mechanisms, applications, and progress. Genes 2016, 7, 88. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Lin, Y.; Shen, G.; Chen, E.; Wang, Y.; Luo, J.; Zhang, H.; Xing, R.; Xia, Q. Female qualities in males: Vitellogenin synthesis induced by ovary transplants into the male silkworm, Bombyx mori. Biochem. Biophys. Res. Commun. 2014, 453, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.; Shu, Y.; Zhou, J.; Zhang, X.; Zhang, X.; Chen, M.; Yao, Q.; Zhou, Q.; Zhang, W. Molecular characterization and RNA interference analysis of vitellogenin receptor from Nilaparvata lugens (Stål). J. Insect Physiol. 2015, 73, 20–29. [Google Scholar] [CrossRef]
- Liu, W.; Guo, S.; Sun, D.; Zhu, L.; Zhu, F.; Lei, C.; Sheng, L.; Phelps, B.; Wang, X. Molecular characterization and juvenile hormone-regulated transcription of the vitellogenin receptor in the cabbage beetle Colaphellus bowringi. Comp. Biochem. Phys. A 2019, 229, 69–75. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Qin, Q.; Wang, D.; Zhou, Y.; He, Y. Selection and evaluation of reference genes for qRT-PCR in Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2021, 12, 902. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purpose | Name | Primer Sequences (5′–3′) |
|---|---|---|
| gene cloning | VgF | ATGAAGTTGTTGGTATTGGC |
| VgR | TTGCTATGTAGTGAGGCTCTTAC | |
| VgRF | ATAATGAAGTATCAAAGCTTGGTATT | |
| VgRR | TTAATTATATTTTAAAATTCTCTGACTCTC | |
| qRT-PCR | qVgF | CAATGAAACTGCTCACAACTACTA |
| qVgR | AGAATCTCACGGTGTCCTAAG | |
| qVgRF | GTGGTTCGGATGAGATACTTT | |
| qVgRR | CCTCGTCTTCACTCTTAGGAC | |
| RNAi | RNAiVgF | TCCAGGGTGCTTTCTTCTAC |
| RNAiVgR | TTCCTTGGTCGCTCTACAA | |
| T7RNAiVgF | taatacgactcactataggTCCAGGGTGCTTTCTTCTAC | |
| T7RNAiVgR | taatacgactcactataggTTCCTTGGTCGCTCTACAA | |
| RNAiVgRF | GCAGTGGATTACTTAGGCG | |
| RNAiVgRR | AGATGCGAGCAGTTGTTGT | |
| T7RNAiVgRF | taatacgactcactataggGCAGTGGATTACTTAGGCG | |
| T7RNAiVgRR | taatacgactcactataggAGATGCGAGCAGTTGTTGT | |
| GFPF | CCACAAGTTCAGCGTGTCCG | |
| GFPR | taatacgactcactataggAAGTTCACCTTGATGCCGTTCT | |
| T7GFPF | CCACAAGTTCAGCGTGTCCG | |
| T7GFPR | taatacgactcactataggAAGTTCACCTTGATGCCGTTCT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, S.; Wang, D.; Song, P.; Zhang, S.; He, Y. Molecular Characterization of Vitellogenin and Its Receptor in Spodoptera frugiperda (J. E. Smith, 1797), and Their Function in Reproduction of Female. Int. J. Mol. Sci. 2022, 23, 11972. https://doi.org/10.3390/ijms231911972
Han S, Wang D, Song P, Zhang S, He Y. Molecular Characterization of Vitellogenin and Its Receptor in Spodoptera frugiperda (J. E. Smith, 1797), and Their Function in Reproduction of Female. International Journal of Molecular Sciences. 2022; 23(19):11972. https://doi.org/10.3390/ijms231911972
Chicago/Turabian StyleHan, Shipeng, Da Wang, Peng Song, Shuo Zhang, and Yunzhuan He. 2022. "Molecular Characterization of Vitellogenin and Its Receptor in Spodoptera frugiperda (J. E. Smith, 1797), and Their Function in Reproduction of Female" International Journal of Molecular Sciences 23, no. 19: 11972. https://doi.org/10.3390/ijms231911972