1. Introduction
Nucleotides act as extracellular messengers that regulate vast functions in almost any cell type present in animal organisms [
1], and their effects are particularly important in the nervous system, both in physiological and pathological conditions [
2,
3]. On cells, nucleotide actions are mediated by P2 receptors, a family of membrane receptors that can be further classified into two different groups: P2Y and P2X receptors [
4,
5]. From these, P2YRs are metabotropic receptors that belong to the G-protein-coupled receptor (GPCR) superfamily. They are formed by seven-membrane-spanning domain proteins coupled to G proteins that are activated selectively by adenosine and uridine tri- and diphosphates. In total, eight different subtypes have been discovered in mammalians: P2Y
1, 2, 4, 6, 11, 12, 13, and
14 [
4]. P2XRs, in turn, are ATP-gated ionic channels consisting of seven distinct subunits (P2X1-7) that are assembled in homotrimers or heterotrimers. The trimeric assembly of the subunits constitutes the functional receptor, whose structure consists of a chalice-like extracellular domain where the three ATP-binding sites are found: the ionic pore, composed by six transmembrane (TM) domains, two TMs for each subunit, and a shorter intracellular domain. Upon nucleotide activation, P2XRs have been shown to elicit membrane depolarization, extracellular calcium influx, and activation of several intracellular signaling pathways, both in a calcium-dependent and -independent manner [
5].
Among the different P2XRs described, the P2X7 receptor is of particular interest for our studies because of its unique structural and pharmacological characteristics. P2X7R is a homotrimer whose subunits show the longest C-termini of all P2X subunits (200 additional amino acids), and its affinity for ATP is 100 times lower than that shown by the other P2X receptors [
5,
6]. Besides its well-known role in the regulation of the immune response [
7,
8] (it was previously known as the P2Z receptor from macrophages and lymphocytes [
9]), P2X7R is implicated in important physiological and pathological processes in the central nervous system, including the modulation of neurotransmitter release, neuritogenesis and synapse formation, the neuroprotective role in cerebellar granule neurons and astrocytes, and several neurodegenerative diseases [
6]. Likewise, multiple intracellular signaling pathways are regulated by P2X7Rs. For instance, P2X7R is known to activate several phospholipases, protein kinases (protein kinases C and D, mitogen-activated protein kinases, and the cellular sarcoma tyrosine kinase), and the phosphoinositide 3-kinase/Akt pathway [
10]. P2X7 receptor is also known to interact with several cytoskeleton-related proteins, such as the extracellular matrix protein laminin-α3, integrin β2, α-actinin, and β-actin [
11], and it has also been reported that P2X7R stimulation in macrophages causes actin-cortex reorganization that is required for the membrane blebbing [
12]. Hence, such tight interaction between P2X7Rs and the cytoskeleton, upon external stimuli, ultimately implies their involvement onto changes in the mechanical properties of the cell. Indeed, previous research in this regard indicated that the signaling of these ionotropic receptors in rat cerebellar astrocytes also includes metabotropic pathways independent from the ionic channel activity, such as MAP kinases ERK1/2 and protein kinase D activation [
13,
14,
15]. Interestingly, while activation of ERK1/2 kinases by P2Y
2/P2Y
4 receptors induce migration of the astrocytes [
16], activation of these kinases by P2X7 receptors cause complex changes in cell morphology [
17]. Therefore, P2X7-originated changes in the cytoskeleton and, by extension, in the mechanical properties of the cell are to be expected. However, a detailed quantification of the triggered mechanical variation has not been reported so far.
In this regard, atomic force microscopy (AFM) is one of the most extended techniques employed to study the mechanical properties of cells and biomaterials. This technique, mostly known for its application on sample topography imaging, can also be used to analyze changes in the stiffness, rheology, and adhesive properties of cells, and it is based on the perpendicular indentation of the cell membrane with a tip located at the end of a cantilever [
18,
19,
20,
21]. The forces resulting from the controlled interaction between the tip and the sample are displayed as force-vs.-distance or force-vs.-time curves (see
Figure 1). Initially, the tip is approached to the sample, and the indentation of the membrane provides information about the deformability and stiffness of the cell [
19]. Upon reaching a previously stablished maximum loading force value (or setpoint), the tip can be maintained at a certain height (stress–relaxation experiments) or can be pushed further in order to keep constant the force applied (creep experiments). Both experimental situations correlate with the rheological properties of the cell and provide quantitative information over factors such as relaxation times and viscosity [
18]. In a last step, the tip is retracted from the sample, causing the cantilever to deflect in the opposite direction due to the adhesion between cell membrane and tip, thus delivering information about the adhesive properties of the cell, such as the maximum adhesive force, the adhesive work required to split the tip-cell contact, and to identify the potential formation of membrane tethers during retracting motion [
20,
22,
23,
24,
25].
On the basis of these principles, previous collaborative work between our groups showed how nucleotide receptor regulation can influence the mechanical properties of cells at the nanoscale, as concluded from the studies performed on P2YR-expressing endothelial HUVEC cells and rat cerebellar astrocytes [
26]. Stimulation of P2Y
2Rs with UTP in both cell types caused an increase in cell stiffness and a decrease in adhesive properties. Further experiments with astrocytes showed that the effects of the purinergic agonist 2MeSADP on the mechanical properties of the cell were mediated by P2Y
13 receptors and negatively modulated by P2Y
1 receptors. As a logical continuation to those initial studies, and attending to what is described above, unravelling the role of P2X7 nucleotide receptors on the mechanical properties of rat cerebellar astrocytes becomes extremely interesting.
Following this, in the present work, we show how stimulation of the P2X7 receptor of rat cerebellar astrocytes with the selective agonist BzATP specifically induced an increase in the cell size, motility, and number of membrane protrusions of the astrocytes in culture. Such effects were reverted when cells were treated with the competitive antagonist of P2X7R, A 438079 (henceforth referred to as A43), prior to treatment with the agonist BzATP. Interestingly, when AFM was employed to further disentangle the role of P2X7 receptor in the modulation of the mechanical properties of this population on the nanoscale, an increase in the stiffness of the cells was observed. However, treatment with A43 did not effectively block such increase in stiffness caused by the agonist, pointing to the fact that the effects observed were not exclusively dependent on the ionotropic activity of the receptor but also on additional effects caused by independent metabotropic pathways that remained unblocked. Similar results were obtained when analyzing the viscous moduli of astrocytes, which indicated potential changes occurring in the cytoskeleton. In order to obtain a more general view on the induced effects, complementary force spectroscopy maps performed over the whole body of treated/untreated cells allowed for the observation of higher stiffness values and stronger adhesive forces throughout all the cell body after treatment with the agonist, further supporting the evidence of changes on the cytoskeleton level.
3. Discussion
Both the presence and activity, upon nucleotide-induced stimulation, of functional P2X7 receptors in rat cerebellar astrocytes in culture were studied and have been described throughout the results reported in this manuscript. Indeed, the identification, quantification, and interpretation of cell responses upon external stimuli remain a topic of great interest for both biologists and biophysicists.
As a main difference from previous studies in the literature, the current experimental conditions and approaches are certainly innovative. For instance, in the present work, astrocytes were plated at very low density (4000 cells/cm
2) in order to follow changes of individual, isolated, cells. This differs from biochemical studies focused on calcium imaging, on which much denser starting populations (400,000 cells/cm
2) were needed, and samples were even incubated for over 24 h until an optimal confluence was achieved [
13,
14,
16]. Another difference would come from the exposure time-range selected (max 4 h) to observe the P2X7R stimulation by nucleotides. For instance, previous studies covered stimulation of P2X7R in rat cerebellar astrocytes with high concentrations of BzATP over a longer period in order to show how such treatment did not induce cell death (in contrast to what was found in mouse cerebellar astrocytes) [
31]. The different, and certainly critical, behavior observed between rat and mouse cells could be hypothesized to stem from differences in the density of the P2X7Rs along the plasma membrane, and/or from their coexistence with other receptors favoring cell survival (P2Y
2R and EGFR) [
16]. In general, such stimuli-induced responses have been observed to a supra-micron scale by monitoring variations on the triggered cell motility, oriented polarization, or changes in size, etc. These factors are extensively described by light-interacting microscopy techniques, from which a true quantification of the forces behind cannot be extracted, unfortunately, unless complementary approaches are considered in parallel (as is the case here).
We first analyzed the cellular behavior upon selective exposure to agonist and antagonist compounds by time-lapse microscopy. As observed in
Figure 2, P2X7R stimulation with BzATP (300 µM) did not affect cell viability and increased the cell size (>20%), cell migration (>200%), and number of protrusions (>50%). In turn, the competitive P2X7R antagonist, A 438079 (or A43, as it has been referred to along the manuscript), significantly reversed the effects of BzATP treatment. These findings agree with previously reported results and highlight the connection between those changes and the mechanical properties of the cell. In order to delve into the intracellular events (i.e., cytoskeletal rearrangement) accounting for these P2X7R-induced responses, we used atomic force microscopy (AFM), which offers a well-established biophysical approach to analyze the potential changes on the mechanical properties of living rat cerebellar astrocytes on the nanometric level. These mechanical parameters were determined after stimulation or blocking of their P2X7 purinergic receptors with BzATP or A43, respectively. A sequential blocking/stimulating combination (A43 followed by exposure to BzATP), which already brought about interesting results for P2YRs to help discriminating between P2Y
1 and P2Y
13 receptor contributions [
26], was also studied. In the current case, AFM-based studies allowed for the identification of effects beyond the mere ionotropic activity of the receptor, which are caused by alternative pathways that remain unblocked. This is here evidenced by more extreme variations in all the analyzed mechanical factors (apparent elastic modulus, adhesion force, and adhesion work) after sequential stimulation of P2X7 receptors with both A43 and BzATP, instead of the initially expected blocking. As a surprising outcome, it must be noted that trends observed for BzATP and A43 + BzATP treatments were overall the same, compared to untreated cells, except for their respective maximum adhesion force values, which went in opposite directions. This factor (F
adh) provides an idea of the strength of the interaction between the indenting tip and the cell, which varied accordingly with the affinity (or selectivity) between both, as well as the availability shown by the cell membrane to be pulled. The latter correlates to a tighter or looser membrane–cytoskeleton connection. Cell stiffness, for instance, is among the (physical) parameters affecting the adhesive behavior of cells. As a general rule, softer samples (cells) usually induce measurement of larger adhesion events, in comparison to stiffer ones. However, in the current case, such an observation was certainly distorted. In terms of stiffness, astrocytes exposed to BzATP appeared in an intermediate level between untreated (control) cells and cells undergoing receptor pre-blocking followed by agonist stimulation (A43 + BzATP). This would lead to predicting a gradual decrease in F
adh from one case to the other. Surprisingly, our measurements suggest a generation of stickier cells upon direct exposure of P2X7 to BzATP in comparison to those generated after A43 blocking. Divagation over this point may be rather diverse: the membrane–cytoskeleton connection was not as strengthened as actin fibers (and other constitutive elements) within the cytoskeleton were (see explanation below); the surface charge was locally affected by means of the activation performed; or, perhaps, a different type of process was triggered. Indeed, a very similar trend was observed in our previous work—when testing exposure to UTP to identify the presence of P2Y
2 receptors in astrocytes. P2Y2R is one of the receptors inducing cell migration, and its stimulation also resulted in stiffer cells, albeit more adhesive [
26].
AFM was also used to determine the viscoelastic behavior of the astrocytes. A good representative of this behavior is the cell viscosity value derived from cytoskeleton contribution, which increased up to a 150% after direct receptor simulation with BzATP when compared with the pre-blocked case (1171.2 vs. 1775.0 Pa s, BzATP and A43 + BzATP, respectively). A larger viscosity clearly pointed towards re-arrangement of cytoskeleton constituents to form a much-reinforced structural supportive mesh. This also correlated with the detection of a stiffer cell when P2X7Rs were initially passivated prior to stimulation, attending to the instantaneous elastic modulus (2.00 vs. 3.06 kPa) behavior. Such reinforcement seemed not to have a dramatic impact at the so-considered cortex level, since
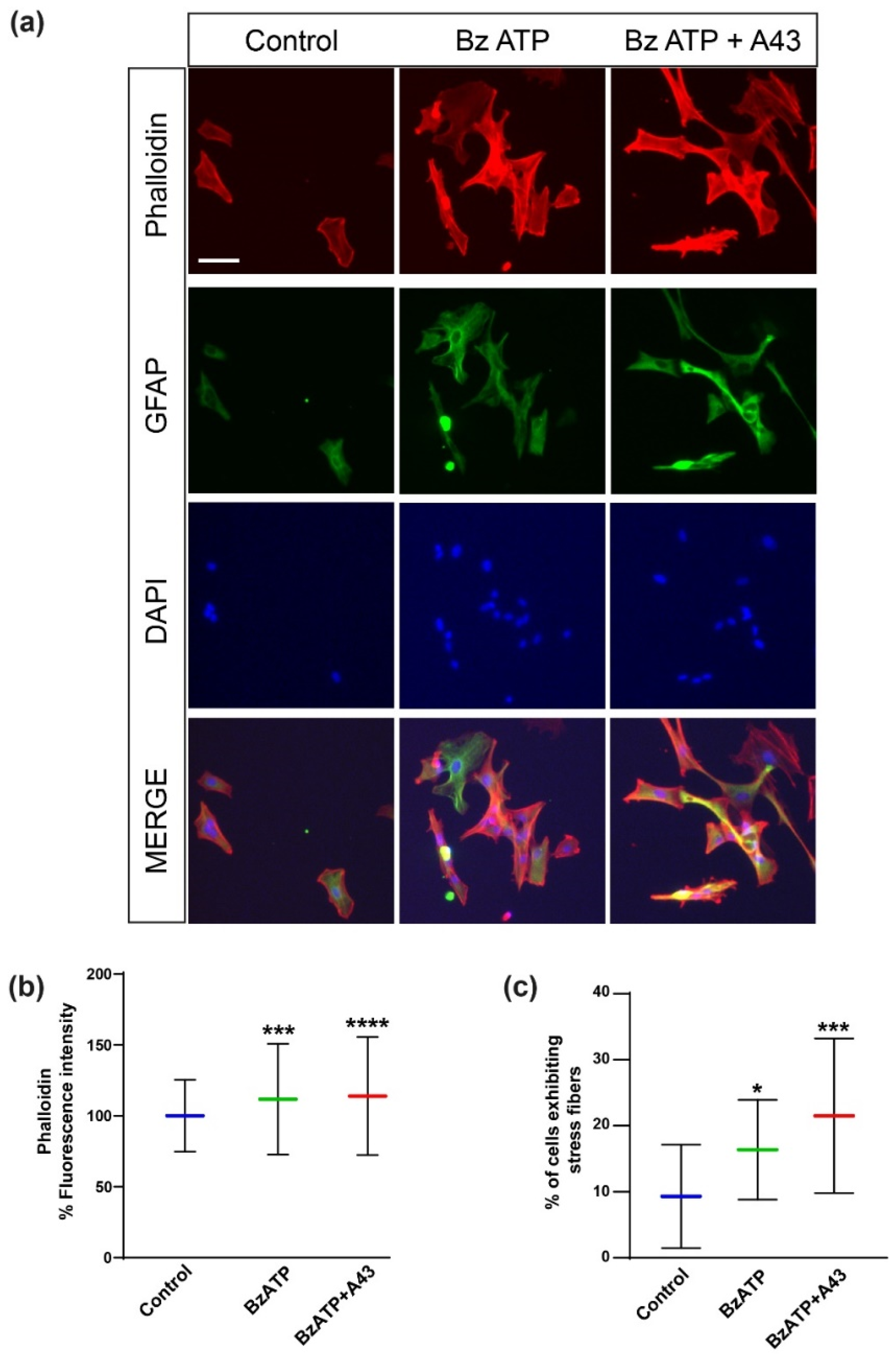
did not vary much, despite an increment of the 58% between A43 + BzATP-treated cells and the control system that should be noted. Additionally, astrocytes treated with BzATP showed more actin stress fibers compared to control conditions (
Figure 8), therefore confirming the implication of actin cytoskeleton in the increase in viscosity observed after P2X7 stimulation.
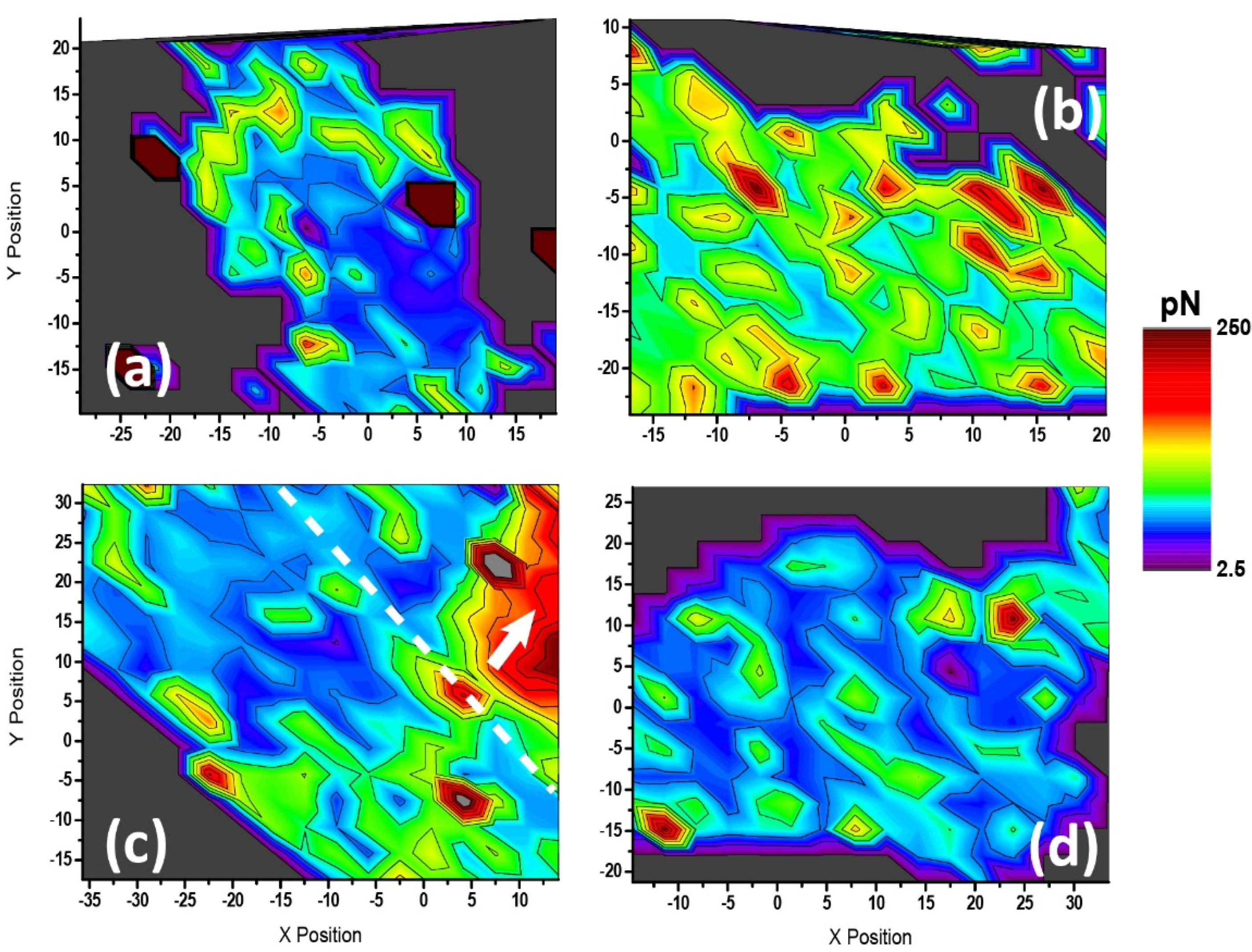
Finally, force spectroscopy maps of the astrocytes (see
Figure 6 and
Figure 7) were generated for the first time. These maps clearly indicated that the cell response upon receptor stimulation occurred all over the cell body, with a special mention to what was observed for adhesion under direct treatment with BzATP: a huge general increase in attachment independently of the cell region, which was not observed under receptor pre-blocking. This result brings us to the discussion above, again showing certain similarities between the effects of P2X7R and P2Y
2R stimulation. In rat cerebellar astrocytes, both P2X7 and P2Y
2 receptors increase intracellular calcium levels and share several intracellular pathways, including protein kinase D (PKD), and mitogen activated protein kinases (MAPKs), such as the extracellular regulated kinases 1 and 2 (ERK1/2) [
14,
15,
16] and p38 [
32]. Furthermore, both receptors also induce transactivation of epidermal growth factor receptor (EGFR), which is essential for the inactivation of MAPKs, preventing MAPK overactivation that would lead to cell death (unpublished results). Despite sharing several intracellular pathways, stimulation of each receptor causes different effects on astrocytes. As mentioned above, P2Y
2 stimulation is linked to astrocyte migration [
16], while P2X7 receptor activation causes morphological changes [
17]. Surprisingly, in the current work, we also determined a slight increment in the migration of astrocytes after P2X7 receptor stimulation. Thus, such an effect could be, at least in part, responsible for the similarity in the changes observed in the mechanical properties of the astrocytes after stimulation of both receptors (stiffer and more adhesive cells). In addition, the activation of MAPKs has been previously linked to actin cortex reorganization in other cell models [
12]. Since MAPKs are activated by both receptors, their implication in the reorganization of the actin cytoskeleton in astrocytes cannot be discarded. Further experiments, employing inhibitors of different MAPKs, are required to determine the effects of this pathways in our cell model.
A final surprising result to mention from the present work is the different effects of the P2X7R antagonist, A43, on both the mechanical properties of the astrocytes and on their activity. A43 completely abolished the increase in migratory capabilities, cell spreading, and membrane protrusions of astrocytes treated with BzATP. However, when analyzing the mechanical properties of the astrocytes with AFM, A43 did not hinder the effects of BzATP on the stiffness and viscosity of the cells. In fact, it appeared to enhance such effects instead. The same applies for the effects observed on the actin cytoskeleton. Such inconsistency in the effects of A43 may lie on the distinctive features of P2X7 receptors. As indicated previously, P2X7 receptors possess the largest C-termini present in P2X receptors. Such a particular feature allows P2X7 receptors to interact with different proteins, such as integrins and β-actin [
11], and also to activate different signaling pathways, such as phospholipase C (PLC), protein kinase C (PKC), and phosphatidil-inositol-3 kinase (PI3K), which are independent from ion channel activation (for a review, see [
10]). While A43 may block the activity of the ionic channel and calcium influx, this type of signaling activated by the C-termini is independent and therefore unaffected by A43 activity. On one hand, the increase on the migratory capabilities of astrocytes after P2X7 stimulation could be dependent on channel ionic activity, and therefore blocked by A43; on the other hand, further effects on the actin cytoskeleton and, by extension, on the mechanical properties of the cell, could be dependent of the activity of the C-termini, and as such, unaffected by A43. Previous work of the group, related to the activation of ERK1/2 in astrocytes after P2X7R stimulation, supports this hypothesis [
32]. Treatment with A43 prior to stimulation of the receptors with BzATP alters the temporal pattern of phosphorylation and activation of ERK1/2, but it does not completely block it [
32]. Although implication of the C-termini in this process is not confirmed, it could explain the activation of ERK1/2 despite the treatment with the antagonist, possibly by means of EGF receptor transactivation [
16,
32]. As stated above, MAPKs, including ERK1/2, are linked to actin cytoskeleton remodeling, and since A43 does not block the activation of these MAPKs after P2X7 stimulation, this could also explain the inability of the antagonist to block the changes in actin cytoskeleton and the mechanical properties of the cell. The enhancement of the effects observed after treatment with both A43 and BzATP, however, suggests complex interactions between different signaling pathways. Unraveling such complex effects elicited by P2X7R stimulation, however, requires further research and is out of the scope of the current work.
4. Materials and Methods
4.1. Chemicals, Materials, and Antibodies
Papain was purchased from Worthington (Lake Wood, NJ, USA). Fetal bovine serum (FBS), Dulbecco’s modified Eagle’s medium (DMEM), penicillin/streptomycin solution, Leibovitz’s-L15 and TrypLE™ Express, rhodamine-conjugated phalloidin, and nuclear counterstain DAPI (4′,6-Diamidino-2-phenylindole) were obtained from Gibco, Thermo-Fisher (Waltham, MA, USA). Plastic Petri dishes and culture flasks were supplied by Falcon Becton Dickinson Labware (Franklin Lakes, NJ, USA). Borosilicate circular cover glasses (diameter: 24 mm, 15 mm, thickness: 0.08–0.12 mm) were purchased from Menzel Gläser, VWR, (Bruchsal, Germany). Bovine serum albumin (BSA), BzATP (2′(3′)-O-(4-benzoylbenzoyl)adenosine 5′-triphosphate), A438079 (3-(5-(2,3-dichlorophenyl)-1H-tetrazol-1-yl)methyl pyridine), and anti-GFAP antibody were provided by Sigma-Aldrich, Merck (Kenilworth, NJ, USA). Alexa Fluor 488 secondary antibody was from Dako/Agilent (Santa Clara, CA, USA). All other non-specified reagents were routinely supplied by Sigma-Aldrich, Merck, or Roche Diagnostics SL (Basel, Switzerland).
4.2. Cell Cultures
Primary cultures of cerebellar astrocytes were prepared at the Complutense University of Madrid, following the International Council for Laboratory Animal Science guidelines, as described previously [
15]. Cerebella from Wistar rat pups (P7) were digested with papain; the resulting cells were resuspended in complete DMEM medium (10% (
v/v) FBS, 2 mM glutamine, 25 mM glucose, 100 U/mL penicillin, and 100 mg/mL streptomycin) and then seeded in culture flasks at a density of 70,000 cells/cm
2. The cells were maintained in culture until they reached confluence, and then they were purified by orbital shaking. Purified astrocytes were then detached from the culture flasks by trypsin digestion and cryopreserved in FBS containing 10% (
v/v) DMSO at −80 °C. Cryopreserved cells were thawed when necessary, resuspended in complete culture medium, and grown in flasks until reaching confluence again.
4.3. Time-Lapse Video Microscopy, Single-Cell Tracking, and Immunofluorescent Staining
24 h before time-lapse video microscopy experiments, astrocytes were seeded at very low density (4000 cells/cm2) onto 15 mm coverslips in 24-well plates in complete culture medium. Before the experiment, cells were serum-starved for 30 min in complete DMEM medium containing only 1% FBS and then stimulated with 300 µM BzATP for 1 h. Where indicated, cells were pretreated with P2X7 antagonist (A 438079, 10 µM) for 5 min, prior to the nucleotide addition.
After treatment, cells were tracked using a NIKON TE-2000 microscope and equipped with a long-distance 20x phase contrast objective (Nikon) and a ZYLA camera from ANDOR [
33]. Cells were kept at a constant temperature of 37 °C and in 5% CO
2, and images were acquired every 5 min over 3 h using the 4.7/NIS-elements software from NIKON.
After finishing the time-lapse experiments, cells were fixed with 4% (w/v) paraformaldehyde for 10 min at room temperature, followed by cell permeabilization with 0.1% (v/v) Triton X-100 in phosphate-buffered saline (PBS) for 15 min at room temperature. Unspecific antibody interactions were blocked by adding 2% (w/v) bovine serum albumin (BSA) in PBS for 1 h at room temperature, and then the samples were incubated with a primary anti-GFAP rabbit antibody and diluted 1:1000 in 1% (w/v) BSA–PBS solution overnight at 4 °C. This was followed by incubation for 1 h at room temperature with a 1% (w/v) solution of BSA–PBS containing a secondary goat anti-rabbit antibody labelled with the fluorophore Alexa Fluor™ 488, diluted 1:500; a rhodamine-labelled phalloidin actin marker, diluted 1:40; and a nuclear counterstain DAPI, with a final concentration of 2 µg/mL. After a final washing step with PBS, samples were air-dried, covered in mounting medium, and stored at 4 °C until their visualization in the fluorescence microscope.
Cell mobility was determined by single-cell tracking, using specific custom software (The Tracking Tool, TTT) [
34]. Migrated distance was determined as the total distance covered by each cell during the 3 h of time-lapse, on the basis of the displacement of the cell nucleus.
4.4. Atomic Force Microscopy
Before AFM experiments (48 h in advance), astrocytes were plated at very low density (4000 cells/cm2) onto 24 mm diameter glass coverslips (previously cleaned with oxygen plasma) in 6-well plates in complete culture medium. Right before the experiments, cells were subjected to trophic factor withdrawal by washing them for 30 min with Leibovitz’s medium, and then the cells were stimulated with 300 µM BzATP in the same medium for 1 h at 37 °C. Where indicated, cells were pretreated with P2X7 antagonist (A 438079, 10 µM) for 5 min, prior to the nucleotide addition.
AFM measurements were carried out in a JPK Nanowizard III (JPK Instruments, Berlin, Germany), mounted on an inverted optical microscope (Axio Observer Z1, Zeiss, Jena, Germany), and equipped with a CellHesion module and a BioCell chamber (JPK Instruments, Berlin, Germany). To ensure the duration and quality of the experiments, cells were maintained in Leibovitz’s medium at 37 °C. Before each experiment, triangular silicon nitride cantilevers equipped with a quadratic pyramidal tip (DNP-10, B, Bruker; r = 22 nm, = half-to-face-angle = 22°) were cleaned with ozone/UV and calibrated on a clean glass surface covered in Leibovitz’s medium at 37 °C, and their spring constant was determined by means of the thermal tune method (nominal spring constant of 0.12 N/m).
Force spectroscopy curves were obtained as previously described [
21,
26,
35]. Cells were indented on top of the nucleus to reduce variability and influence of the substrate on the measurements. The tip was approached at a constant velocity of 5 µm/s, and after reaching a setpoint of 1.5 nN, it was kept in contact at constant height (stress–relaxation measurements) for a period of 5 s. The tip was then retracted at the same constant velocity (5 µm/s). To ensure tip cleanliness, the glass substrate was probed in between the indentation of cells. For statistical purposes, each cell was indented 5 times, and at least 20 cells per condition were analyzed.
Complementary mapping analysis was applied on individual cells, after the selection of an optimal zone and performance of an optical calibration. Then, measurements were run over a 32 × 32 pixel map covering between 30 and 40 µm2 areas, attending to the respective cell shapes. Each of these pixels originated individual force vs. distance plots recorded at the same conditions described above. As a main difference to previous measurements, the pause segment was omitted in the recording of the maps (retraction started at maximum load achievement) due to experimental feasibility, and the tip was immediately retracted at maximum loading force.
4.5. AFM Data Analysis
Force curves were analyzed using JPKSPM Data Processing software (JPK Instruments, Berlin, Germany) and the R afmToolkit [
36]. The batch data processing from JPK was particularly useful in the case of force-volume maps and also offered a good starting point for comparison with those analyzed with R in the case of individual measurements. Regarding R afmToolkit, force spectroscopy curves were imported into the toolkit, and contact point detection, baseline calibration, and zero-point determination were realized using the corresponding algorithms of the toolkit [
37]. Deformation of the sample (
) was determined as indicated in Equation (1):
where
is the extension of the piezo,
is the deformation of the cantilever,
is the force, and
is the spring constant of the cantilever. Apparent Young’s modulus (
) values were obtained for an indentation range of 500 nm, following a Hertz–Sneddon model optimized for the tip geometry employed (quadratic pyramid) according to Equation (2):
where
is the force,
is the Poisson ratio (set to 0.5),
is the half-to-face angle of the tip used (22°), and
is the indentation of the sample.
Stress–relaxation pause segments were used to determine the viscoelastic properties of the cell. A double exponential decay function was fitted to the stress–relaxation segments, and calculation of the different viscoelastic values were determined by applying a Zener model of the cell (a spring in parallel with two Maxwell elements), following Equation (3):
where
is the Poisson ratio,
is the equilibrium modulus (at infinite time),
is the compressive moduli of the respective springs of the Maxwell elements,
is the relaxation times of the viscous dashpots, and
C is a constant that depends on the geometry of the indenter. For pyramidal tips,
C is defined as indicated in Equation (4):
where
is the half-to-face angle of the tip used and
is the indentation value that is kept constant during the stress relaxation phase. Lastly, viscosities of the dashpots (
) were calculated following Equation (5):
where
is the compressive moduli of the respective springs of the Maxwell elements and
is the relaxation times of the viscous dashpots.
Retract curves were adapted similarly, and maximum adhesion force was measured as the lowest point of the curve in the Y-axis. The total number of rupture events, their relative position, and the rupture force associated were also saved for further analysis.
In the case of colored maps, these were obtained after batch processing of the related plots, data exporting of the thus extracted factors, and final plotting in Origin software.
4.6. Statistical Analysis and Data Presentation
Statistical analysis of AFM data was carried out using the OriginPro 2018 (OriginLab, Northhampton, MA, USA) program. At least 20 cells per experimental condition were analyzed, and 5 force curves were obtained from each cell. Values for the apparent Young’s modulus (), cell indentation, maximum adhesive force, compressive moduli, relaxation times, and viscous moduli were obtained as indicated above, and the outliers were removed using Grubb’s test. Statistically significant differences between the different experimental conditions were determined with an analysis of the variance test (one-way ANOVA) followed by Dunnett’s post-test analysis, and they are reported in the figures as * for p < 0.05, ** for p < 0.01, *** for p < 0.001, and **** for p < 0.0001. Data graphs were generated in OriginPro 2018 and further processed using Adobe Illustrator Adobe Inc, San José, CA, USA.
Statistical analysis of time-lapse video microscopy experiments was performed using GraphPadPrism8 (GraphPadsoftware), and one-way ANOVA followed by Dunnett’s multiple comparisons tests was used to analyze the results. The data are presented as the mean ± SD, and each independent experiment shown was reproduced at least three times. A p-value < 0.05 was considered statistically significant.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}