
Medicinal Herbs and Their Derived Ingredients Protect against Cognitive Decline in In Vivo Models of Alzheimer’s Disease

Abstract
:1. Introduction
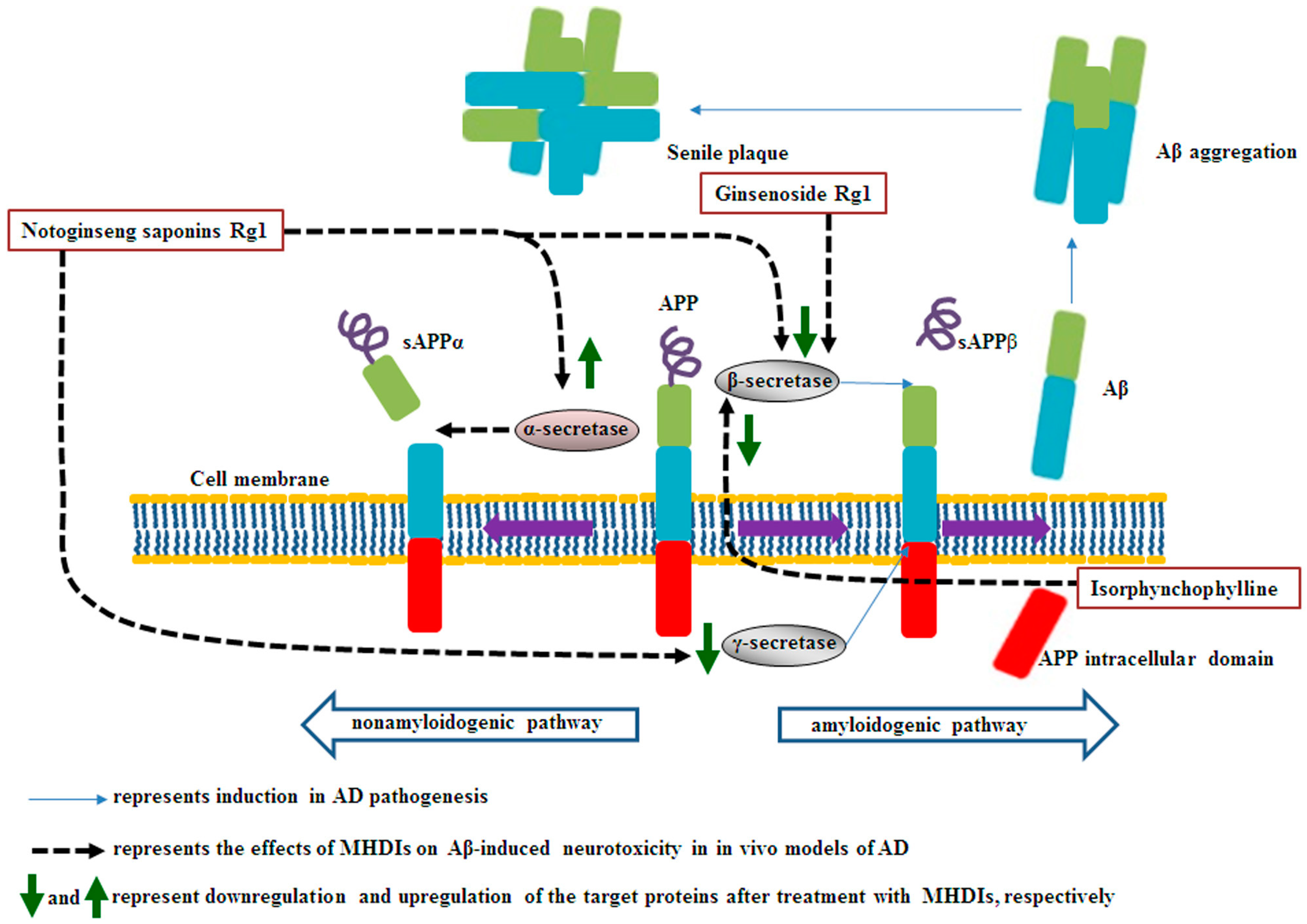
2. MHDI-Mediated Suppression of Aβ Accumulation
2.1. Effects of MHDIs on Aβ Accumulation through α-, β-, and γ-Secretase Activity Regulation
2.2. Summary
3. MHDI-Mediated Inhibition of Aβ-Induced Oxidative Stress
3.1. Involvement of Decreased Antioxidant Status and Increased Lipid Peroxidation in Aβ-Induced Oxidative Stress
3.2. Effects of MHDIs on Aβ-Induced Oxidative Stress through Antioxidant Activity and Lipid Oxidation Regulation
3.3. Summary
4. MHDI-Mediated Downregulation of Tau Hyperphosphorylation
4.1. Effects of MHDIs on Aβ-Induced Tau Hyperphosphorylation through PP2A, CDK5, and GSK-3β Expression Regulation
4.2. Summary
5. MHDI-Mediated Reduction of Aβ-Induced Inflammation
5.1. Effects of MHDIs on Aβ-Induced Inflammation through Inflammatory Mediator Regulation
5.2. Effects of MHDIs on Aβ-Induced Inflammation through Receptor for Advanced Glycation End Product- and MAPK-Mediated Signaling Regulation
5.3. Summary
6. MHDI-Mediated Amelioration of Aβ-Induced Synaptic Dysfunction
6.1. Involvement of Synaptic Protein Expression in Aβ-Induced Synaptic Dysfunction
6.2. Effects of MHDIs on Aβ-Induced Synaptic Dysfunction through Synaptic Protein Expression Regulation
6.3. Involvement of Acetylcholine Release in Aβ-Induced Synaptic Dysfunction
6.4. Effects of MHDIs on Aβ-Induced Synaptic Dysfunction through ChAT, ACh, and AChE Level Regulation
6.5. Involvement of Postsynaptic Receptor and Protein Expression in Aβ-Induced Synaptic Dysfunction
6.6. Effects of MHDIs on Aβ-Induced Synaptic Dysfunction through Postsynaptic Receptor and Protein Expression Regulation
6.7. Summary
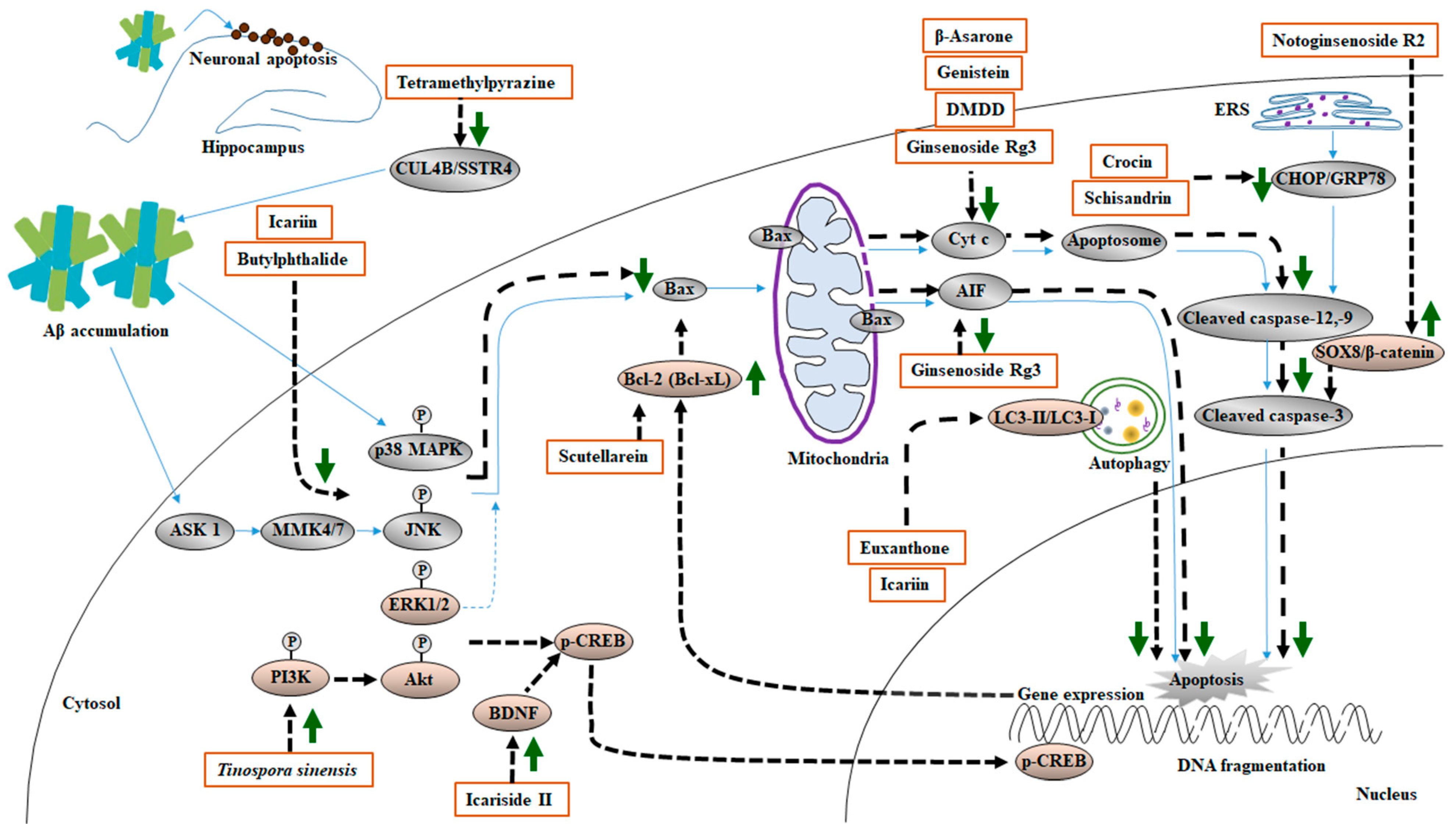
7. MHDI-Mediated Attenuation of Aβ-Induced Apoptosis
7.1. Involvement of MAPK and PI3K/Akt Signaling in Aβ-Induced Apoptosis
7.2. Effects of MHDIs on Aβ-Induced Apoptosis through MAPK-, PI3K/Akt-, and BDNF/CREB-Mediated Signaling Regulation
7.3. Involvement of Mitochondria-Mediated Apoptotic Cascades in Aβ-Induced Apoptosis
7.4. Effects of MHDIs on Aβ-Induced Apoptosis through Bax-, Cullin 4B-, and β-Catenin-Mediated Signaling Regulation
7.5. Effects of MHDIs on Aβ-Induced Apoptosis through Endoplasmic Reticulum Stress and Autophagy Signaling Regulation
7.6. Summary
8. Conclusions
9. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sadigh-Eteghad, S.; Sabermarouf, B.; Majdi, A.; Talebi, M.; Farhoudi, M.; Mahmoudi, J. Amyloid-beta: A crucial factor in Alzheimer’s disease. Med. Princ. Pract. 2015, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Ma, S.; Lv, J.; Wang, X.; Afewerky, H.K.; Li, H.; Lu, Y. The emerging role of exosomes in Alzheimer’s disease. Ageing Res. Rev. 2021, 68, 101321. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.J.T.; Frendo-Cumbo, S.; MacPherson, R.E.K. Resveratrol and Metformin Recover Prefrontal Cortex AMPK Activation in Diet-Induced Obese Mice but Reduce BDNF and Synaptophysin Protein Content. J. Alzheimers Dis. 2019, 71, 945–956. [Google Scholar] [CrossRef]
- Kincheski, G.C.; Valentim, I.S.; Clarke, J.R.; Cozachenco, D.; Castelo-Branco, M.T.L.; Ramos-Lobo, A.M.; Rumjanek, V.; Donato, J., Jr.; De Felice, F.G.; Ferreira, S.T. Chronic sleep restriction promotes brain inflammation and synapse loss, and potentiates memory impairment induced by amyloid-beta oligomers in mice. Brain Behav. Immun. 2017, 64, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, J.; Yang, X.; Jin, Y.; Yang, Z.; Wang, R.; Zhang, F.; Linhardt, R.J. Akebia saponin D reverses corticosterone hypersecretion in an Alzheimer’s disease rat model. Biomed. Pharmacother. 2018, 107, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Nobakht, M.; Hoseini, S.M.; Mortazavi, P.; Sohrabi, I.; Esmailzade, B.; Rahbar Rooshandel, N.; Omidzahir, S. Neuropathological changes in brain cortex and hippocampus in a rat model of Alzheimer’s disease. Iran. Biomed. J. 2011, 15, 51–58. [Google Scholar]
- Sheng, C.; Peng, W.; Xia, Z.A.; Wang, Y.; Chen, Z.; Su, N.; Wang, Z. The impact of ginsenosides on cognitive deficits in experimental animal studies of Alzheimer’s disease: A systematic review. BMC Complement. Altern. Med. 2015, 15, 386. [Google Scholar] [CrossRef]
- Weng, G.; Zhou, B.; Liu, T.; Huang, Z.; Huang, S. Tetramethylpyrazine Improves Cognitive Function of Alzheimer’s Disease Mice by Regulating SSTR4 Ubiquitination. Drug Des. Devel. Ther. 2021, 15, 2385–2399. [Google Scholar] [CrossRef]
- Wong, W. Economic burden of Alzheimer disease and managed care considerations. Am. J. Manag. Care. 2020, 26, S177–S183. [Google Scholar]
- Song, F.X.; Wang, L.; Liu, H.; Wang, Y.L.; Zou, Y. Brain cell apoptosis inhibition by butylphthalide in Alzheimer’s disease model in rats. Exp. Ther. Med. 2017, 13, 2771–2774. [Google Scholar] [CrossRef]
- Wei, M.; Liu, Z.; Liu, Y.; Li, S.; Hu, M.; Yue, K.; Liu, T.; He, Y.; Pi, Z.; Liu, Z.; et al. Urinary and plasmatic metabolomics strategy to explore the holistic mechanism of lignans in S. chinensis in treating Alzheimer’s disease using UPLC-Q-TOF-MS. Food Funct. 2019, 10, 5656–5668. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.F.; Dong, Y.; Chen, J.Y.; Lu, J.H. The effect and underlying mechanisms of garlic extract against cognitive impairment and Alzheimer’s disease: A systematic review and meta-analysis of experimental animal studies. J. Ethnopharmacol. 2021, 280, 114423. [Google Scholar] [CrossRef] [PubMed]
- Chiroma, S.M.; Baharuldin, M.T.H.; Mat Taib, C.N.; Amom, Z.; Jagadeesan, S.; Ilham Adenan, M.; Mahdi, O.; Moklas, M.A.M. Centella asiatica Protects d-Galactose/AlCl3 Mediated Alzheimer’s Disease-Like Rats via PP2A/GSK-3beta Signaling Pathway in Their Hippocampus. Int. J. Mol. Sci. 2019, 20, 1871. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hossain, S.; Abdullah, N.; Aminudin, N. Brain proteomics links oxidative stress with metabolic and cellular stress response proteins in behavioural alteration of Alzheimer’s disease model rats. AIMS Neurosci. 2019, 6, 299–315. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, B.; Shao, J.; Wang, T.T.; Cai, R.Z.; Ma, C.J.; Han, T.; Du, J. Genistein suppresses the mitochondrial apoptotic pathway in hippocampal neurons in rats with Alzheimer’s disease. Neural. Regen. Res. 2016, 11, 1153–1158. [Google Scholar] [PubMed]
- Park, Y.M.; Lee, B.G.; Park, S.H.; Oh, H.G.; Kang, Y.G.; Kim, O.J.; Kwon, L.S.; Kim, Y.P.; Choi, M.H.; Jeong, Y.S.; et al. Prolonged oral administration of Gastrodia elata extract improves spatial learning and memory of scopolamine-treated rats. Lab. Anim. Res. 2015, 31, 69–77. [Google Scholar] [CrossRef]
- Ali, T.; Yoon, G.H.; Shah, S.A.; Lee, H.Y.; Kim, M.O. Osmotin attenuates amyloid beta-induced memory impairment, tau phosphorylation and neurodegeneration in the mouse hippocampus. Sci. Rep. 2015, 5, 11708. [Google Scholar] [CrossRef]
- Kim, J.H.; Lim, D.K.; Suh, Y.H.; Chang, K.A. Long-Term Treatment of Cuban Policosanol Attenuates Abnormal Oxidative Stress and Inflammatory Response via Amyloid Plaques Reduction in 5xFAD Mice. Antioxidants 2021, 10, 1321. [Google Scholar] [CrossRef]
- Pang, K.; Jiang, R.; Zhang, W.; Yang, Z.; Li, L.L.; Shimozawa, M.; Tambaro, S.; Mayer, J.; Zhang, B.; Li, M.; et al. An App knock-in rat model for Alzheimer’s disease exhibiting Abeta and tau pathologies, neuronal death and cognitive impairments. Cell Res. 2022, 32, 157–175. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, Y.; Zhang, L.; Huang, L.; Yu, P.; Zhu, H.; Deng, W.; Qin, C. Down-Regulated Drebrin Aggravates Cognitive Impairments in a Mouse Model of Alzheimer’s Disease. Int. J. Mol. Sci. 2017, 18, 800. [Google Scholar] [CrossRef]
- Ghasemi, R.; Zarifkar, A.; Rastegar, K.; Maghsoudi, N.; Moosavi, M. Insulin protects against Abeta-induced spatial memory impairment, hippocampal apoptosis and MAPKs signaling disruption. Neuropharmacology 2014, 85, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.; Wang, X.; Jin, S.; Hu, J.; Wu, Y.; Li, Y.; Wu, X. Activation of GPR55 attenuates cognitive impairment and neurotoxicity in a mouse model of Alzheimer’s disease induced by Abeta1-42 through inhibiting RhoA/ROCK2 pathway. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2022, 112, 110423. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.L.; Zhang, P.; Lin, L.; Zhang, H.M.; Deng, S.D.; Wu, Z.Q.; Ou, S.; Liu, S.H.; Wang, J.Y. Protective effects of bajijiasu in a rat model of Abeta(2)(5)(-)(3)(5)-induced neurotoxicity. J. Ethnopharmacol. 2014, 154, 206–217. [Google Scholar] [CrossRef] [PubMed]
- Zussy, C.; Brureau, A.; Keller, E.; Marchal, S.; Blayo, C.; Delair, B.; Ixart, G.; Maurice, T.; Givalois, L. Alzheimer’s disease related markers, cellular toxicity and behavioral deficits induced six weeks after oligomeric amyloid-beta peptide injection in rats. PLoS ONE 2013, 8, e53117. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.Q.; Lai, X.X.; Xu, L.T.; Shen, Y.; Lin, J.W.; Gao, S.Y.; Hu, H.Y. Qingxin Kaiqiao Fang Inhibits Abeta 25-35-Induced Apoptosis in Primary Cultured Rat Hippocampal Neuronal Cells via the p38 MAPK Pathway: An Experimental Validation and Network Pharmacology Study. Evid. Based Complement. Alternat. Med. 2020, 2020, 9058135. [Google Scholar]
- Zhu, Q.; Zhang, N.; Hu, N.; Jiang, R.; Lu, H.; Xuan, A.; Long, D.; Chen, Y. Neural stem cell transplantation improves learning and memory by protecting cholinergic neurons and restoring synaptic impairment in an amyloid precursor protein/presenilin 1 transgenic mouse model of Alzheimer’s disease. Mol. Med. Rep. 2020, 21, 1172–1180. [Google Scholar] [CrossRef]
- Belaya, I.; Ivanova, M.; Sorvari, A.; Ilicic, M.; Loppi, S.; Koivisto, H.; Varricchio, A.; Tikkanen, H.; Walker, F.R.; Atalay, M.; et al. Astrocyte remodeling in the beneficial effects of long-term voluntary exercise in Alzheimer’s disease. J. Neuroinflammation 2020, 17, 271. [Google Scholar] [CrossRef]
- Luo, R.; Su, L.Y.; Li, G.; Yang, J.; Liu, Q.; Yang, L.X.; Zhang, D.F.; Zhou, H.; Xu, M.; Fan, Y.; et al. Activation of PPARA-mediated autophagy reduces Alzheimer disease-like pathology and cognitive decline in a murine model. Autophagy 2020, 16, 52–69. [Google Scholar] [CrossRef]
- Liu, S.; Li, X.; Gao, J.; Liu, Y.; Shi, J.; Gong, Q. Icariside II, a Phosphodiesterase-5 Inhibitor, Attenuates Beta-Amyloid-Induced Cognitive Deficits via BDNF/TrkB/CREB Signaling. Cell Physiol. Biochem. 2018, 49, 985. [Google Scholar] [CrossRef]
- Wu, T.Y.; Chen, C.P.; Jinn, T.R. Traditional Chinese medicines and Alzheimer’s disease. Taiwan J. Obstet. Gynecol. 2011, 50, 131–135. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Su, M.; Chu, X.; Li, S.; Yue, Y.; Zhang, X.; Wang, J.; Han, F. Brain metabolomics study for the protective effects of Rhodiola crenulata extract on Alzheimer’s disease by HPLC coupled with Fourier transform-ion cyclotron resonance mass spectrometry. J. Sep. Sci. 2020, 43, 3216–3223. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.M.; Schiapparelli, L.; Salazar-Colocho, P.; Cuadrado-Tejedor, M.; Escribano, L.; Lopez de Maturana, R.; Del Rio, J.; Perez-Mediavilla, A.; Frechilla, D. Overexpression of wild-type human APP in mice causes cognitive deficits and pathological features unrelated to Abeta levels. Neurobiol. Dis. 2009, 33, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Deng, Y.; Gao, J.M.; Lv, C.; Lang, L.H.; Shi, J.S.; Yu, C.Y.; Gong, Q.H. Icariside II inhibits lipopolysaccharide-induced inflammation and amyloid production in rat astrocytes by regulating IKK/IkappaB/NF-kappaB/BACE1 signaling pathway. Acta Pharmacol. Sin. 2020, 41, 154–162. [Google Scholar] [CrossRef]
- Tambini, M.D.; Norris, K.A.; D’Adamio, L. Opposite changes in APP processing and human Abeta levels in rats carrying either a protective or a pathogenic APP mutation. eLife 2020, 9, e52612. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Z.; Cheng, W.; Shao, J.W.; Gu, Y.F.; Zhu, Y.Y.; Dong, Q.J.; Bai, S.Y.; Wang, P.; Lin, L. Notoginseng Saponin Rg1 Prevents Cognitive Impairment through Modulating APP Processing in Abeta1-42-injected Rats. Curr. Med. Sci. 2019, 39, 196–203. [Google Scholar] [CrossRef]
- Manczak, M.; Kandimalla, R.; Yin, X.; Reddy, P.H. Hippocampal mutant APP and amyloid beta-induced cognitive decline, dendritic spine loss, defective autophagy, mitophagy and mitochondrial abnormalities in a mouse model of Alzheimer’s disease. Hum. Mol. Genet. 2018, 27, 1332–1342. [Google Scholar] [CrossRef]
- Furcila, D.; Dominguez-Alvaro, M.; DeFelipe, J.; Alonso-Nanclares, L. Subregional Density of Neurons, Neurofibrillary Tangles and Amyloid Plaques in the Hippocampus of Patients With Alzheimer’s Disease. Front. Neuroanat. 2019, 13, 99. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, L.; Lu, J.; Jiao, J.; Yang, Y.; Zhao, H.; Liang, Z.; Zheng, H. Ginsenoside Rg1 improves cognitive capability and affects the microbiota of large intestine of tree shrew model for Alzheimer’s disease. Mol. Med. Rep. 2021, 23, 291. [Google Scholar] [CrossRef]
- Li, J.; Du, Q.; Li, N.; Du, S.; Sun, Z. Alpiniae oxyphyllae Fructus and Alzheimer’s disease: An update and current perspective on this traditional Chinese medicine. Biomed. Pharmacother. 2021, 135, 111167. [Google Scholar] [CrossRef]
- Vassar, R.; Kovacs, D.M.; Yan, R.; Wong, P.C. The beta-secretase enzyme BACE in health and Alzheimer’s disease: Regulation, cell biology, function, and therapeutic potential. J. Neurosci. 2009, 29, 12787–12794. [Google Scholar] [CrossRef]
- Neumann, U.; Ufer, M.; Jacobson, L.H.; Rouzade-Dominguez, M.L.; Huledal, G.; Kolly, C.; Luond, R.M.; Machauer, R.; Veenstra, S.J.; Hurth, K.; et al. The BACE-1 inhibitor CNP520 for prevention trials in Alzheimer’s disease. EMBO Mol. Med. 2018, 10, e9316. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Q.; Ip, S.P.; Yuan, Q.J.; Zheng, G.Q.; Tsim, K.K.W.; Dong, T.T.X.; Lin, G.; Han, Y.; Liu, Y.; Xian, Y.F.; et al. Isorhynchophylline ameliorates cognitive impairment via modulating amyloid pathology, tau hyperphosphorylation and neuroinflammation: Studies in a transgenic mouse model of Alzheimer’s disease. Brain Behav. Immun. 2019, 82, 264–278. [Google Scholar] [CrossRef]
- McDade, E.; Voytyuk, I.; Aisen, P.; Bateman, R.J.; Carrillo, M.C.; De Strooper, B.; Haass, C.; Reiman, E.M.; Sperling, R.; Tariot, P.N.; et al. The case for low-level BACE1 inhibition for the prevention of Alzheimer disease. Nat. Rev. Neurol. 2021, 17, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Jo, M.H.; Ikram, M.; Khan, A.; Kim, M.O. Deciphering the Potential Neuroprotective Effects of Luteolin against Abeta1-42-Induced Alzheimer’s Disease. Int. J. Mol. Sci. 2021, 22, 9583. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.T.; Wang, Z.Z.; Yuan, Y.H.; Sun, H.M.; Chen, N.H.; Zhang, Y. Mangiferin: A multipotent natural product preventing neurodegeneration in Alzheimer’s and Parkinson’s disease models. Pharmacol. Res. 2019, 146, 104336. [Google Scholar] [CrossRef] [PubMed]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxid. Med. Cell Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef]
- Huang, X.B.; Chen, Y.J.; Chen, W.Q.; Wang, N.Q.; Wu, X.L.; Liu, Y. Neuroprotective effects of tenuigenin on neurobehavior, oxidative stress, and tau hyperphosphorylation induced by intracerebroventricular streptozotocin in rats. Brain Circ. 2018, 4, 24–32. [Google Scholar]
- Shunan, D.; Yu, M.; Guan, H.; Zhou, Y. Neuroprotective effect of Betalain against AlCl3-induced Alzheimer’s disease in Sprague Dawley Rats via putative modulation of oxidative stress and nuclear factor kappa B (NF-kappaB) signaling pathway. Biomed. Pharmacother. 2021, 137, 111369. [Google Scholar] [CrossRef]
- Xiong, Z.; Liu, C.; Wang, F.; Li, C.; Wang, W.; Wang, J.; Chen, J. Protective effects of breviscapine on ischemic vascular dementia in rats. Biol. Pharm. Bull. 2006, 29, 1880–1885. [Google Scholar] [CrossRef]
- Liu, J.; Yan, X.; Li, L.; Zhu, Y.; Qin, K.; Zhou, L.; Sun, D.; Zhang, X.; Ye, R.; Zhao, G. Ginsennoside rd attenuates cognitive dysfunction in a rat model of Alzheimer’s disease. Neurochem. Res. 2012, 37, 2738–2747. [Google Scholar] [CrossRef]
- Ashabi, G.; Alamdary, S.Z.; Ramin, M.; Khodagholi, F. Reduction of hippocampal apoptosis by intracerebroventricular administration of extracellular signal-regulated protein kinase and/or p38 inhibitors in amyloid beta rat model of Alzheimer’s disease: Involvement of nuclear-related factor-2 and nuclear factor-kappaB. Basic Clin. Pharmacol. Toxicol. 2013, 112, 145–155. [Google Scholar] [PubMed]
- Ferreira, F.S.; Biasibetti-Brendler, H.; Pierozan, P.; Schmitz, F.; Berto, C.G.; Prezzi, C.A.; Manfredini, V.; Wyse, A.T.S. Kynurenic Acid Restores Nrf2 Levels and Prevents Quinolinic Acid-Induced Toxicity in Rat Striatal Slices. Mol. Neurobiol. 2018, 55, 8538–8549. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wei, M.; Fan, M.; Liu, Z.; Wang, A.; Liu, Y.; Men, L.; Pi, Z.; Liu, Z.; Song, F. Pharmacokinetic and metabolomics approach based on UHPLC-MS to evaluate therapeutic effect of lignans from S. Chinensis in alzheimer’s disease. J. Chromatogr. B 2021, 1178, 122859. [Google Scholar] [CrossRef]
- Duffy, S.L.; Lagopoulos, J.; Hickie, I.B.; Diamond, K.; Graeber, M.B.; Lewis, S.J.; Naismith, S.L. Glutathione relates to neuropsychological functioning in mild cognitive impairment. Alzheimers Dement. 2014, 10, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Ruan, Y.Y.; Xu, H.; Shi, X.M.; Wang, Z.X.; Hu, Y.L. Safflower yellow reduces lipid peroxidation, neuropathology, tau phosphorylation and ameliorates amyloid beta-induced impairment of learning and memory in rats. Biomed. Pharmacother. 2015, 76, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.X.; Bao, X.Q.; Li, L.; Yang, H.Y.; Wang, L.; Yu, Y.; Wang, X.L.; Yao, X.S.; Zhang, D. The Protective Effects of Gardenia jasminoides (Fructus Gardenia) on Amyloid-beta-Induced Mouse Cognitive Impairment and Neurotoxicity. Am. J. Chin. Med. 2018, 46, 389–405. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, X.; Wang, S.; Song, S. Ginsenoside Rg3 Prevents Cognitive Impairment by Improving Mitochondrial Dysfunction in the Rat Model of Alzheimer’s Disease. J. Agric. Food Chem. 2019, 67, 10048–10058. [Google Scholar] [CrossRef]
- Yin, S.; Ran, Q.; Yang, J.; Zhao, Y.; Li, C. Nootropic effect of neferine on aluminium chloride-induced Alzheimer’s disease in experimental models. J. Biochem. Mol. Toxicol. 2020, 34, e22429. [Google Scholar] [CrossRef]
- Tampellini, D. Synaptic activity and Alzheimer’s disease: A critical update. Front. Neurosci. 2015, 9, 423. [Google Scholar] [CrossRef]
- Chen, Y.; Han, S.; Huang, X.; Ni, J.; He, X. The Protective Effect of Icariin on Mitochondrial Transport and Distribution in Primary Hippocampal Neurons from 3x Tg-AD Mice. Int. J. Mol. Sci. 2016, 17, 163. [Google Scholar] [CrossRef]
- Zeng, P.; Shi, Y.; Wang, X.M.; Lin, L.; Du, Y.J.; Tang, N.; Wang, Q.; Fang, Y.Y.; Wang, J.Z.; Zhou, X.W.; et al. Emodin Rescued Hyperhomocysteinemia-Induced Dementia and Alzheimer’s Disease-Like Features in Rats. Int. J. Neuropsychopharmacol. 2019, 22, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gong, Q.; Wu, Q.; Li, F.; Lu, Y.; Shi, J. Alkaloids enriched extract from Dendrobium nobile Lindl. attenuates tau protein hyperphosphorylation and apoptosis induced by lipopolysaccharide in rat brain. Phytomedicine 2014, 21, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, X.; Hu, X.; Chu, X.; Li, X.; Han, F. Neuroprotective effects of a Rhodiola crenulata extract on amyloid-beta peptides (Abeta1-42) -induced cognitive deficits in rat models of Alzheimer’s disease. Phytomedicine 2019, 57, 331–338. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Liu, Y.; Xu, Q.Q.; Xian, Y.F.; Lin, Z.X. Sulforaphene Ameliorates Neuroinflammation and Hyperphosphorylated Tau Protein via Regulating the PI3K/Akt/GSK-3beta Pathway in Experimental Models of Alzheimer’s Disease. Oxid. Med. Cell Longev. 2020, 2020, 4754195. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Kayama, T.; Noguchi-Shinohara, M.; Hamaguchi, T.; Yamada, M.; Abe, K.; Kobayashi, S. Rosmarinic acid suppresses tau phosphorylation and cognitive decline by downregulating the JNK signaling pathway. NPJ Sci. Food 2021, 5, 1. [Google Scholar] [CrossRef]
- Xiong, R.; Wang, X.L.; Wu, J.M.; Tang, Y.; Qiu, W.Q.; Shen, X.; Teng, J.F.; Pan, R.; Zhao, Y.; Yu, L.; et al. Polyphenols isolated from lychee seed inhibit Alzheimer’s disease-associated Tau through improving insulin resistance via the IRS-1/PI3K/Akt/GSK-3beta pathway. J. Ethnopharmacol. 2020, 251, 112548. [Google Scholar] [CrossRef]
- Sun, Y.; Wu, A.; Li, X.; Qin, D.; Jin, B.; Liu, J.; Tang, Y.; Wu, J.; Yu, C. The seed of Litchi chinensis fraction ameliorates hippocampal neuronal injury in an Abeta25-35-induced Alzheimer’s disease rat model via the AKT/GSK-3beta pathway. Pharm. Biol. 2020, 58, 35–43. [Google Scholar] [CrossRef]
- Ding, B.; Lin, C.; Liu, Q.; He, Y.; Ruganzu, J.B.; Jin, H.; Peng, X.; Ji, S.; Ma, Y.; Yang, W. Tanshinone IIA attenuates neuroinflammation via inhibiting RAGE/NF-kappaB signaling pathway in vivo and in vitro. J. Neuroinflammation 2020, 17, 302. [Google Scholar] [CrossRef]
- Eikelenboom, P.; Hoozemans, J.J.; Veerhuis, R.; van Exel, E.; Rozemuller, A.J.; van Gool, W.A. Whether, when and how chronic inflammation increases the risk of developing late-onset Alzheimer’s disease. Alzheimers Res. Ther. 2012, 4, 15. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Z.; Zang, C.; Wang, L.; Yang, H.; Sheng, C.; Shang, J.; Zhao, Z.; Yuan, F.; Yu, Y.; et al. GJ-4 ameliorates memory impairment in focal cerebral ischemia/reperfusion of rats via inhibiting JAK2/STAT1-mediated neuroinflammation. J. Ethnopharmacol. 2021, 267, 113491. [Google Scholar] [CrossRef]
- Hickman, S.E.; Allison, E.K.; El Khoury, J. Microglial dysfunction and defective beta-amyloid clearance pathways in aging Alzheimer’s disease mice. J. Neurosci. 2008, 28, 8354–8360. [Google Scholar] [CrossRef]
- Song, L.; Piao, Z.; Yao, L.; Zhang, L.; Lu, Y. Schisandrin ameliorates cognitive deficits, endoplasmic reticulum stress and neuroinflammation in streptozotocin (STZ)-induced Alzheimer’s disease rats. Exp. Anim. 2020, 69, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.; Giannopoulos, P.F.; Ceballos-Diaz, C.; Golde, T.E.; Pratico, D. 5-Lipoxygenase gene transfer worsens memory, amyloid, and tau brain pathologies in a mouse model of Alzheimer disease. Ann. Neurol. 2012, 72, 442–454. [Google Scholar] [CrossRef] [PubMed]
- Guo, E.; Hu, Y.; Du, T.; Zhu, H.; Chen, L.; Qu, W.; Zhang, J.; Xie, N.; Liu, W.; Feng, F.; et al. Effects of Picrasma quassioides and its active constituents on Alzheimer’s disease in vitro and in vivo. Bioorg. Chem. 2019, 92, 103258. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, R.; Zhu, S.; Ma, J.; Pang, L.; Ma, B.; Du, L.; Jin, Y. Nasal timosaponin BII dually sensitive in situ hydrogels for the prevention of Alzheimer’s disease induced by lipopolysaccharides. Int. J. Pharm. 2020, 578, 119115. [Google Scholar] [CrossRef]
- Xie, L.; Zhang, N.; Zhang, Q.; Li, C.; Sandhu, A.F.; Iii, G.W.; Lin, S.; Lv, P.; Liu, Y.; Wu, Q.; et al. Inflammatory factors and amyloid beta-induced microglial polarization promote inflammatory crosstalk with astrocytes. Aging 2020, 12, 22538–22549. [Google Scholar] [PubMed]
- Zhang, L.; Zhou, Z.; Zhai, W.; Pang, J.; Mo, Y.; Yang, G.; Qu, Z.; Hu, Y. Safflower yellow attenuates learning and memory deficits in amyloid beta-induced Alzheimer’s disease rats by inhibiting neuroglia cell activation and inflammatory signaling pathways. Metab. Brain Dis. 2019, 34, 927–939. [Google Scholar] [CrossRef]
- Pang, J.; Hou, J.; Zhou, Z.; Ren, M.; Mo, Y.; Yang, G.; Qu, Z.; Hu, Y. Safflower Yellow Improves Synaptic Plasticity in APP/PS1 Mice by Regulating Microglia Activation Phenotypes and BDNF/TrkB/ERK Signaling Pathway. Neuromolecular Med. 2020, 22, 341–358. [Google Scholar] [CrossRef]
- Tang, J.J.; Huang, L.F.; Deng, J.L.; Wang, Y.M.; Guo, C.; Peng, X.N.; Liu, Z.; Gao, J.M. Cognitive enhancement and neuroprotective effects of OABL, a sesquiterpene lactone in 5xFAD Alzheimer’s disease mice model. Redox. Biol. 2022, 50, 102229. [Google Scholar] [CrossRef]
- Sakata, K.; Mastin, J.R.; Duke, S.M.; Vail, M.G.; Overacre, A.E.; Dong, B.E.; Jha, S. Effects of antidepressant treatment on mice lacking brain-derived neurotrophic factor expression through promoter IV. Eur. J. Neurosci. 2013, 37, 1863–1874. [Google Scholar] [CrossRef]
- Chu, S.; Gu, J.; Feng, L.; Liu, J.; Zhang, M.; Jia, X.; Liu, M.; Yao, D. Ginsenoside Rg5 improves cognitive dysfunction and beta-amyloid deposition in STZ-induced memory impaired rats via attenuating neuroinflammatory responses. Int. Immunopharmacol. 2014, 19, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.F.; Wang, B.J.; Cheng, H.T.; Kuo, L.H.; Wolfe, M.S. Tumor necrosis factor-alpha, interleukin-1beta, and interferon-gamma stimulate gamma-secretase-mediated cleavage of amyloid precursor protein through a JNK-dependent MAPK pathway. J. Biol. Chem. 2004, 279, 49523–49532. [Google Scholar] [CrossRef]
- Alam, J.J. Selective Brain-Targeted Antagonism of p38 MAPKalpha Reduces Hippocampal IL-1beta Levels and Improves Morris Water Maze Performance in Aged Rats. J. Alzheimers Dis. 2015, 48, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.W.; Tsai, F.H.; Lan, W.C.; Cheng, Y.D.; Lee, S.C.; Wu, C.R. Steroid-Enriched Fraction of Achyranthes bidentata Protects Amyloid beta Peptide 1-40-Induced Cognitive Dysfunction and Neuroinflammation in Rats. Mol. Neurobiol. 2019, 56, 5671–5688. [Google Scholar] [CrossRef]
- Wang, Y.; Li, J.; Hua, L.; Han, B.; Zhang, Y.; Yang, X.; Zeng, Z.; Bai, H.; Yin, H.; Lou, J. Effects of caffeic acid on learning deficits in a model of Alzheimer’s disease. Int. J. Mol. Med. 2016, 38, 869–875. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Zhu, Y.; Yan, L.; Zhao, L. Neuroprotective Effect of S-trans, Trans-farnesylthiosalicylic Acid via Inhibition of RAS/ERK Pathway for the Treatment of Alzheimer’s Disease. Drug Des. Devel. Ther. 2019, 13, 4053–4063. [Google Scholar] [CrossRef]
- Tampellini, D.; Capetillo-Zarate, E.; Dumont, M.; Huang, Z.; Yu, F.; Lin, M.T.; Gouras, G.K. Effects of synaptic modulation on beta-amyloid, synaptophysin, and memory performance in Alzheimer’s disease transgenic mice. J. Neurosci. 2010, 30, 14299–14304. [Google Scholar] [CrossRef]
- Zhan, P.Y.; Peng, C.X.; Zhang, L.H. Berberine rescues D-galactose-induced synaptic/memory impairment by regulating the levels of Arc. Pharmacol. Biochem. Behav. 2014, 117, 47–51. [Google Scholar] [CrossRef]
- Vyas, Y.; Montgomery, J.M.; Cheyne, J.E. Hippocampal Deficits in Amyloid-beta-Related Rodent Models of Alzheimer’s Disease. Front. Neurosci. 2020, 14, 266. [Google Scholar] [CrossRef]
- Xia, Z.; Wang, F.; Zhou, S.; Zhang, R.; Huang, J.H.; Wu, E.; Zhang, Y.; Hu, Y. Catalpol protects synaptic proteins from beta-amyloid induced neuron injury and improves cognitive functions in aged rats. Oncotarget 2017, 8, 69303–69315. [Google Scholar] [CrossRef]
- Li, B.; Mao, Q.; Zhao, N.; Xia, J.; Zhao, Y.; Xu, B. Treadmill exercise overcomes memory deficits related to synaptic plasticity through modulating ionic glutamate receptors. Behav. Brain Res. 2021, 414, 113502. [Google Scholar] [CrossRef]
- Ji, Z.H.; Xu, Z.Q.; Zhao, H.; Yu, X.Y. Neuroprotective effect and mechanism of daucosterol palmitate in ameliorating learning and memory impairment in a rat model of Alzheimer’s disease. Steroids 2017, 119, 31–35. [Google Scholar] [CrossRef]
- Cascella, R.; Conti, S.; Tatini, F.; Evangelisti, E.; Scartabelli, T.; Casamenti, F.; Wilson, M.R.; Chiti, F.; Cecchi, C. Extracellular chaperones prevent Abeta42-induced toxicity in rat brains. Biochim. Biophys. Acta. 2013, 1832, 1217–1226. [Google Scholar] [CrossRef]
- Li, Y.; Xu, J.; Xu, P.; Song, S.; Liu, P.; Chi, T.; Ji, X.; Jin, G.; Qiu, S.; Hou, Y.; et al. Xanthoceras sorbifolia extracts ameliorate dendritic spine deficiency and cognitive decline via upregulation of BDNF expression in a rat model of Alzheimer’s disease. Neurosci. Lett. 2016, 629, 208–214. [Google Scholar] [CrossRef]
- Koo, Y.S.; Kim, H.; Park, J.H.; Kim, M.J.; Shin, Y.I.; Choi, B.T.; Lee, S.Y.; Shin, H.K. Indoleamine 2,3-Dioxygenase-Dependent Neurotoxic Kynurenine Metabolism Contributes to Poststroke Depression Induced in Mice by Ischemic Stroke along with Spatial Restraint Stress. Oxid. Med. Cell Longev. 2018, 2018, 2413841. [Google Scholar] [CrossRef]
- Audrain, M.; Fol, R.; Dutar, P.; Potier, B.; Billard, J.M.; Flament, J.; Alves, S.; Burlot, M.A.; Dufayet-Chaffaud, G.; Bemelmans, A.P.; et al. Alzheimer’s disease-like APP processing in wild-type mice identifies synaptic defects as initial steps of disease progression. Mol. Neurodegener. 2016, 11, 5. [Google Scholar] [CrossRef]
- Yi, X.X.; Li, J.Y.; Tang, Z.Z.; Jiang, S.; Liu, Y.H.; Deng, J.G.; Gao, C.H. Marinoid J, a phenylglycoside from Avicennia marina fruit, ameliorates cognitive impairment in rat vascular dementia: A quantitative iTRAQ proteomic study. Pharm. Biol. 2020, 58, 1211–1220. [Google Scholar] [CrossRef]
- Sheng, C.; Xu, P.; Zhou, K.; Deng, D.; Zhang, C.; Wang, Z. Icariin Attenuates Synaptic and Cognitive Deficits in an Abeta1-42-Induced Rat Model of Alzheimer’s Disease. Biomed. Res. Int. 2017, 2017, 7464872. [Google Scholar] [CrossRef]
- Fan, C.H.; Lin, C.W.; Huang, H.J.; Lee-Chen, G.J.; Sun, Y.C.; Lin, W.; Chen, C.M.; Chang, K.H.; Su, M.T.; Hsieh-Li, H.M. LMDS-1, a potential TrkB receptor agonist provides a safe and neurotrophic effect for early-phase Alzheimer’s disease. Psychopharmacology 2020, 237, 3173–3190. [Google Scholar] [CrossRef]
- Yuan, Y.; Shan, X.; Men, W.; Zhai, H.; Qiao, X.; Geng, L.; Li, C. The effect of crocin on memory, hippocampal acetylcholine level, and apoptosis in a rat model of cerebral ischemia. Biomed. Pharmacother. 2020, 130, 110543. [Google Scholar] [CrossRef]
- Huang, G.B.; Zhao, T.; Muna, S.S.; Jin, H.M.; Park, J.I.; Jo, K.S.; Lee, B.H.; Chae, S.W.; Kim, S.Y.; Park, S.H.; et al. Therapeutic potential of Gastrodia elata Blume for the treatment of Alzheimer’s disease. Neural. Regen. Res. 2013, 8, 1061–1070. [Google Scholar]
- Suvorina, M.Y.; Selivanova, O.M.; Grigorashvili, E.I.; Nikulin, A.D.; Marchenkov, V.V.; Surin, A.K.; Galzitskaya, O.V. Studies of Polymorphism of Amyloid-beta42 Peptide from Different Suppliers. J. Alzheimers Dis. 2015, 47, 583–593. [Google Scholar] [CrossRef]
- Yuede, C.M.; Dong, H.; Csernansky, J.G. Anti-dementia drugs and hippocampal-dependent memory in rodents. Behav. Pharmacol. 2007, 18, 347–363. [Google Scholar] [CrossRef]
- Hassan, N.A.; Alshamari, A.K.; Hassan, A.A.; Elharrif, M.G.; Alhajri, A.M.; Sattam, M.; Khattab, R.R. Advances on Therapeutic Strategies for Alzheimer’s Disease: From Medicinal Plant to Nanotechnology. Molecules 2022, 27, 4839. [Google Scholar] [CrossRef]
- Tuzimski, T.; Petruczynik, A. Determination of Anti-Alzheimer’s Disease Activity of Selected Plant Ingredients. Molecules 2022, 27, 3222. [Google Scholar] [CrossRef]
- Meunier, J.; Ieni, J.; Maurice, T. The anti-amnesic and neuroprotective effects of donepezil against amyloid beta 25–35 peptide-induced toxicity in mice involve an interaction with the sigma1 receptor. Br. J. Pharmacol. 2006, 149, 998–1012. [Google Scholar] [CrossRef]
- Takata, K.; Kitamura, Y.; Saeki, M.; Terada, M.; Kagitani, S.; Kitamura, R.; Fujikawa, Y.; Maelicke, A.; Tomimoto, H.; Taniguchi, T.; et al. Galantamine-induced amyloid-β clearance mediated via stimulation of microglial nicotinic acetylcholine receptors. J. Biol. Chem. 2010, 285, 40180–40191. [Google Scholar] [CrossRef]
- Nikiforuk, A.; Potasiewicz, A.; Kos, T.; Popik, P. The combination of memantine and galantamine improves cognition in rats: The synergistic role of the alpha7 nicotinic acetylcholine and NMDA receptors. Behav. Brain Res. 2016, 313, 214–218. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Naz, F.; Rahul; Varshney, H. Comparative study of rivastigmine and galantamine on the transgenic Drosophila model of Alzheimer’s disease. Curr. Res. Pharmacol. Drug Discov. 2022, 3, 100120. [Google Scholar] [CrossRef]
- Seo, E.J.; Fischer, N.; Efferth, T. Phytochemicals as inhibitors of NF-kappaB for treatment of Alzheimer’s disease. Pharmacol. Res. 2018, 129, 262–273. [Google Scholar] [CrossRef]
- Sahoo, A.K.; Dandapat, J.; Dash, U.C.; Kanhar, S. Features and outcomes of drugs for combination therapy as multi-targets strategy to combat Alzheimer’s disease. J. Ethnopharmacol. 2018, 215, 42–73. [Google Scholar] [CrossRef]
- Tang, Y.; Yu, C.; Wu, J.; Chen, H.; Zeng, Y.; Wang, X.; Yang, L.; Mei, Q.; Cao, S.; Qin, D. Lychee seed extract protects against neuronal injury and improves cognitive function in rats with type II diabetes mellitus with cognitive impairment. Int. J. Mol. Med. 2018, 41, 251–263. [Google Scholar] [CrossRef]
- Liu, Z.; Lv, C.; Zhao, W.; Song, Y.; Pei, D.; Xu, T. NR2B-containing NMDA receptors expression and their relationship to apoptosis in hippocampus of Alzheimer’s disease-like rats. Neurochem. Res. 2012, 37, 1420–1427. [Google Scholar] [CrossRef]
- Qiu, Y.; Xue, X.J.; Liu, G.; Shen, M.M.; Chao, C.Y.; Zhang, J.; Guo, Y.Q.; Niu, Q.Q.; Yu, Y.N.; Song, Y.T.; et al. Perillaldehyde improves cognitive function in vivo and in vitro by inhibiting neuronal damage via blocking TRPM2/NMDAR pathway. Chin. Med. 2021, 16, 136. [Google Scholar] [CrossRef]
- Jiang, H.; Ashraf, G.M.; Liu, M.; Zhao, K.; Wang, Y.; Wang, L.; Xing, J.; Alghamdi, B.S.; Li, Z.; Liu, R. Tilianin Ameliorates Cognitive Dysfunction and Neuronal Damage in Rats with Vascular Dementia via p-CaMKII/ERK/CREB and ox-CaMKII-Dependent MAPK/NF-kappaB Pathways. Oxid. Med. Cell Longev. 2021, 2021, 6673967. [Google Scholar] [CrossRef]
- Wang, K.; Sun, W.; Zhang, L.; Guo, W.; Xu, J.; Liu, S.; Zhou, Z.; Zhang, Y. Oleanolic Acid Ameliorates A beta 25–35 Injection-induced Memory Deficit in Alzheimer’s Disease Model Rats by Maintaining Synaptic Plasticity. CNS Neurol. Disord. Drug Targets 2018, 17, 389–399. [Google Scholar] [CrossRef]
- Wei, G.; Chen, Y.B.; Chen, D.F.; Lai, X.P.; Liu, D.H.; Deng, R.D.; Zhou, J.H.; Zhang, S.X.; Li, Y.W.; Lii, H.; et al. beta-Asarone inhibits neuronal apoptosis via the CaMKII/CREB/Bcl-2 signaling pathway in an in vitro model and AbetaPP/PS1 mice. J. Alzheimers Dis. 2013, 33, 863–880. [Google Scholar] [CrossRef]
- Xu, G.; Shi, Y. Apoptosis signaling pathways and lymphocyte homeostasis. Cell Res. 2007, 17, 759–771. [Google Scholar] [CrossRef]
- Liu, J.; Li, C.; Xing, G.; Zhou, L.; Dong, M.; Geng, Y.; Li, X.; Li, J.; Wang, G.; Zou, D.; et al. Beta-asarone attenuates neuronal apoptosis induced by Beta amyloid in rat hippocampus. Yakugaku Zasshi 2010, 130, 737–746. [Google Scholar] [CrossRef]
- Xue, M.Q.; Liu, X.X.; Zhang, Y.L.; Gao, F.G. Nicotine exerts neuroprotective effects against beta-amyloid-induced neurotoxicity in SH-SY5Y cells through the Erk1/2-p38-JNK-dependent signaling pathway. Int. J. Mol. Med. 2014, 33, 925–933. [Google Scholar] [CrossRef]
- Ma, B.; Meng, X.; Wang, J.; Sun, J.; Ren, X.; Qin, M.; Sun, G.; Sun, X. Notoginsenoside R1 attenuates amyloid-beta-induced damage in neurons by inhibiting reactive oxygen species and modulating MAPK activation. Int. Immunopharmacol. 2014, 22, 151–159. [Google Scholar] [CrossRef]
- Zong, N.; Li, F.; Deng, Y.; Shi, J.; Jin, F.; Gong, Q. Icariin, a major constituent from Epimedium brevicornum, attenuates ibotenic acid-induced excitotoxicity in rat hippocampus. Behav. Brain Res. 2016, 313, 111–119. [Google Scholar] [CrossRef]
- Borsello, T.; Forloni, G. JNK signalling: A possible target to prevent neurodegeneration. Curr. Pharm. Des. 2007, 13, 1875–1886. [Google Scholar] [CrossRef]
- Frasca, G.; Carbonaro, V.; Merlo, S.; Copani, A.; Sortino, M.A. Integrins mediate beta-amyloid-induced cell-cycle activation and neuronal death. J. Neurosci. Res. 2008, 86, 350–355. [Google Scholar] [CrossRef]
- Zhou, F.; He, K.; Guan, Y.; Yang, X.; Chen, Y.; Sun, M.; Qiu, X.; Yan, F.; Huang, H.; Yao, L.; et al. Network pharmacology-based strategy to investigate pharmacological mechanisms of Tinospora sinensis for treatment of Alzheimer’s disease. J. Ethnopharmacol. 2020, 259, 112940. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Kao, S.T.; Lee, Y.C. Angelica sinensis extract protects against ischemia-reperfusion injury in the hippocampus by activating p38 MAPK-mediated p90RSK/p-Bad and p90RSK/CREB/BDNF signaling after transient global cerebral ischemia in rats. J. Ethnopharmacol. 2020, 252, 112612. [Google Scholar] [CrossRef]
- Adams, J.M.; Cory, S. The Bcl-2 protein family: Arbiters of cell survival. Science 1998, 281, 1322–1326. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Kaipia, A.; McGee, E.; Lomeli, M.; Hsueh, A.J. Bok is a pro-apoptotic Bcl-2 protein with restricted expression in reproductive tissues and heterodimerizes with selective anti-apoptotic Bcl-2 family members. Proc. Natl. Acad. Sci. USA 1997, 94, 12401–12406. [Google Scholar] [CrossRef]
- Zhao, T.; Fu, Y.; Sun, H.; Liu, X. Ligustrazine suppresses neuron apoptosis via the Bax/Bcl-2 and caspase-3 pathway in PC12 cells and in rats with vascular dementia. IUBMB Life 2018, 70, 60–70. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Li, Z.G. Cathepsin B and phospo-JNK in relation to ongoing apoptosis after transient focal cerebral ischemia in the rat. Neurochem. Res. 2012, 37, 948–957. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, C.M.; Wan, H.Y.; Wu, J.L.; Quan, W.Q.; Wu, K.Y.; Li, D. Neuroprotection against permanent focal cerebral ischemia by ginkgolides A and B is associated with obstruction of the mitochondrial apoptotic pathway via inhibition of c-Jun N-terminal kinase in rats. J. Neurosci. Res. 2014, 92, 232–242. [Google Scholar] [CrossRef]
- Zhou, L.; Ao, L.Y.; Yan, Y.Y.; Li, W.T.; Ye, A.Q.; Li, C.Y.; Shen, W.Y.; Liang, B.W.; Xiong, Z.; Li, Y.M. JLX001 Ameliorates Ischemia/Reperfusion Injury by Reducing Neuronal Apoptosis via Down-Regulating JNK Signaling Pathway. Neuroscience 2019, 418, 189–204. [Google Scholar] [CrossRef]
- Wei, X.; Xu, X.; Chen, Z.; Liang, T.; Wen, Q.; Qin, N.; Huang, W.; Huang, X.; Li, Y.; Li, J.; et al. Protective Effects of 2-Dodecyl-6-Methoxycyclohexa-2,5 -Diene-1,4-Dione Isolated from Averrhoa Carambola L. (Oxalidaceae) Roots on Neuron Apoptosis and Memory Deficits in Alzheimer’s Disease. Cell Physiol. Biochem. 2018, 49, 1064–1073. [Google Scholar] [CrossRef]
- Huang, X.W.; Xu, Y.; Sui, X.; Lin, H.; Xu, J.M.; Han, D.; Ye, D.D.; Lv, G.F.; Liu, Y.X.; Qu, X.B.; et al. Scutellarein suppresses Abeta-induced memory impairment via inhibition of the NF-kappaB pathway in vivo and in vitro. Oncol. Lett. 2019, 17, 5581–5589. [Google Scholar]
- Han, Y.; Wang, N.; Kang, J.; Fang, Y. beta-Asarone improves learning and memory in Abeta1-42-induced Alzheimer’s disease rats by regulating PINK1-Parkin-mediated mitophagy. Metab. Brain Dis. 2020, 35, 1109–1117. [Google Scholar] [CrossRef]
- Wang, H.L.; Chang, N.C.; Weng, Y.H.; Yeh, T.H. XLID CUL4B mutants are defective in promoting TSC2 degradation and positively regulating mTOR signaling in neocortical neurons. Biochim. Biophys. Acta. 2013, 1832, 585–593. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Pina-Crespo, J.; Li, Y. Restoring Wnt/beta-catenin signaling is a promising therapeutic strategy for Alzheimer’s disease. Mol. Brain 2019, 12, 104. [Google Scholar] [CrossRef]
- Xie, S.L.; Fan, S.; Zhang, S.Y.; Chen, W.X.; Li, Q.X.; Pan, G.K.; Zhang, H.Q.; Wang, W.W.; Weng, B.; Zhang, Z.; et al. SOX8 regulates cancer stem-like properties and cisplatin-induced EMT in tongue squamous cell carcinoma by acting on the Wnt/beta-catenin pathway. Int. J. Cancer 2018, 142, 1252–1265. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, L.; Jiang, L.; Liang, N.; Zhu, X.; He, Q.; Qin, H.; Chen, W. Notoginsenoside R2 reduces Abeta25-35-induced neuronal apoptosis and inflammation via miR-27a/SOX8/beta-catenin axis. Hum. Exp. Toxicol. 2021, 40, S347–S358. [Google Scholar] [CrossRef]
- Lin, L.; Liu, G.; Yang, L. Crocin Improves Cognitive Behavior in Rats with Alzheimer’s Disease by Regulating Endoplasmic Reticulum Stress and Apoptosis. Biomed. Res. Int. 2019, 2019, 9454913. [Google Scholar] [CrossRef]
- Yorimitsu, T.; Nair, U.; Yang, Z.; Klionsky, D.J. Endoplasmic reticulum stress triggers autophagy. J. Biol. Chem. 2006, 281, 30299–30304. [Google Scholar] [CrossRef]
- Sheng, R.; Zhang, L.S.; Han, R.; Liu, X.Q.; Gao, B.; Qin, Z.H. Autophagy activation is associated with neuroprotection in a rat model of focal cerebral ischemic preconditioning. Autophagy 2010, 6, 482–494. [Google Scholar] [CrossRef]
- Carloni, S.; Buonocore, G.; Balduini, W. Protective role of autophagy in neonatal hypoxia-ischemia induced brain injury. Neurobiol. Dis. 2008, 32, 329–339. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, L.L.; Lan, Z.; Xiong, F.; Xu, X.; Yin, Y.Y.; Li, P.; Wang, P. Icariin Ameliorates Amyloid Pathologies by Maintaining Homeostasis of Autophagic Systems in Abeta1-42-Injected Rats. Neurochem. Res. 2019, 44, 2708–2722. [Google Scholar] [CrossRef]
- Yuan, H.; Jiang, C.; Zhao, J.; Zhao, Y.; Zhang, Y.; Xu, Y.; Gao, X.; Guo, L.; Liu, Y.; Liu, K.; et al. Euxanthone Attenuates Abeta1-42-Induced Oxidative Stress and Apoptosis by Triggering Autophagy. J. Mol. Neurosci. 2018, 66, 512–523. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Ingredients | Isolated from Medicinal Herbs | Anti-Aβ Accumulation Activities | Models | Reference |
|---|---|---|---|---|
| Notoginseng saponin Rg1 | Panax notoginseng | α-secretase↑, β- secretase↓, γ-secretase↓ | 28 days after Aβ1–42-induced AD | [35] |
| Ginsenoside Rg1 | Bcl-2↑, MAP-2↑, NeuN↑, Bax↓, β-secretase↓ | 6 weeks after Aβ25–35-induced AD | [38] | |
| Isorphynchophylline | Uncaria tomentosa | BACE-1↓, presenilin 1↓, p-APP (Thr668) ↓ | 129 days in TgCRND8 transgenic mice | [42] |
| Major Ingredients | Isolated from Medicinal Herbs | Antioxidative Stress Activities | Models | References |
|---|---|---|---|---|
| Ginsennoside Rd | Panax ginseng | 4-HNE↓ | 5 days after Aβ1–40-induced AD | [50] |
| Lignans | Schisandra chinensis Baill | kynurenic acid↑, Nrf2↑ | 28 days after Aβ25–35-induced AD | [53] |
| Bajijiasu | Morinda officinalis | SOD↑, CAT↑, GSH-Px↑, MDA↓ | 25 days after Aβ25–35-induced AD | [23] |
| Safflower yellow | Carthamus tinctorius | SOD↑, GSH-Px↑, MDA↓ | 28 days after Aβ1–42-induced AD | [55] |
| GJ-4 | Gardenia jasminoides J. Ellis | SOD↑, MDA↓, iNOS↓, COX-2↓, PGE2↓, TNF-α↓ | 10 days after Aβ25–35-induced AD | [56] |
| Tenuigenin | Polygala tenuifolia Willd | SOD↑, GSH-Px↑, MDA↓, 4-HNE↓ p-tau (Ser396) ↓, p-tau (Thr181) ↓ | 28 days after STZ-induced AD | [47] |
| Ginsenoside Rg3 | P. ginseng C. A. Meyer | SOD↑, CAT↑, GSH-Px↑, MDA↓, | 60 days after D-galactose-induced AD | [57] |
| Neferine | Nelumbo nucifera | SOD↑, CAT↑, GSH-Px↑ | 4 days after AlCl3-induced AD | [58] |
| Rhodiola crenulata | GSH-Px↓, arachidonic acid↓ | 28 days after Aβ1–42-induced AD | [31] | |
| Betalin | Beta vulgaris L. | SOD↑, CAT↑, GSH-Px↑, MDA↓, | 28 days after AlCl3-induced AD | [48] |
| Major Ingredients | Isolated from Medicinal Herbs | Anti-p-Tau Activities | Models | References |
|---|---|---|---|---|
| Dendrobium nobile Lindl. | GSK-3β↓ p-tau (Ser199-202) ↓, p-tau (Ser396) ↓, p-tau (Ser404) ↓, p-tau (Thr231), p-tau (Thr205) | 7 days after LPS-induced AD | [62] | |
| Safflower yellow | C. tinctorius | PP2A↑, CDK5↓, GSK-3↓ | 28 days after Aβ1–42-induced AD | [55] |
| Emodin | Rheum officinale | PP2A↑, p-CREB↑, SYP↑, SYN-1↑, BACE-1↓, | 14 days after Hcy-induced AD | [61] |
| Centella asiatica | PP2A↑, Bcl-2 mRNA↑, GSK-3β↓ | 10 weeks after d-galactose/AlCl3-induced AD | [13] | |
| R. crenulata | GSK-3β (Ser9)/GSK-3β↑ | 28 days after Aβ1–42-induced AD | [63] | |
| Sulforaphene | Raphani semen | p-Akt (Ser473) ↑, p-GSK-3β (Ser9) ↑, IL-10↑, TNF-α↓, IL-6↓ | 6 weeks after s STZ-induced AD | [64] |
| Seed of Litchi chinensis | Akt↑, GSK-3β↓ | 28 days after Aβ25–35-induced AD | [67] |
| Major Ingredients | Isolated from Medicinal Herbs | Anti-Inflammation Activities | Models | References |
|---|---|---|---|---|
| Emodin | R. officinale | microglia activation↓, TNF-α↓, IL-6↓, 5-LO↓, NF-κB↓ | 14 days after Hcy-induced AD | [61] |
| Ethyl acetate | Picrasma quassioides Benn | TNF-α↓, IL-1β↓, IL-6↓ | 23 days after Aβ25–35-induced AD | [74] |
| Betalin | B. vulgaris L. | TNF-α mRNA↓, IL-1β mRNA↓, IL-6 mRNA↓, iNOS mRNA↓, COX-2 mRNA↓, NF-κB↓ | 28 days after AlCl3-induced AD | [48] |
| Neferine | N. nucifera | TNF-α↓, IL-1β↓, IL-6↓, iNOS↓, COX-2↓, NF-κB↓ | 4 days after AlCl3-induced AD | [58] |
| Timosaponin BII | Anemarrhena asphodeloides Bunge | TNF-α↓, IL-1β↓, iNOS↓ | 38 days after LPS-induced inflammation and AD | [75] |
| Schisandrin | S. chinensis Baill | Sirtuin 1↑, TNF-α↓, IL-1β↓, IL-6↓, NF-κB↓ | 14 days after STZ-induced AD | [72] |
| Cuban policosanol | Saccharum officinarum | 4-HNE↓, TNF-α↓, IL-1β↓, IL-6↓ | 4 months in 5xFAD transgenic mice | [18] |
| Ginsennoside Rd | Panax ginseng | IL-10↑, HSP70↑, Iba1↓, GFAP↓, TNF-α↓, IL-1β↓, IL-6↓, caspase-3↓ | 5 days after Aβ1–40-induced AD | [50] |
| Ginsenoside Rg5 | P. ginseng | BDNF↑, IGF↑, ChAT↑, TNF-α↓, IL-1β↓, iNOS↓, COX-2↓, AChE↓ | 28 days after STZ-induced AD | [81] |
| Safflower yellow | C. tinctorius L. | TNF-α↓, IL-1β↓, IL-6↓, iNOS mRNA↓, Arg1↑(marker of M2 microglia), YM-1 mRNA↑ (M2-related cytokine), CD206 mRNA↑ (M2-related cytokine) | 28 days after Aβ1–42-induced AD | [77] |
| Tanshinone IIA | salvia miltiorrhiza Bunge | TNF-α↓, IL-1β↓, IL-6↓, RAGE↓, NF-κB↓ | 30 days in APP/PS1 transgenic mice | [68] |
| Caffeic acid | Ocimum gratissimum | p-p38 MAPK↓, NF-κB-p65↓, TNF-α↓, IL-6↓, p53↓, AChE↓, CAT↑, GSH-Px↑ | 14 days after Aβ1–40-induced AD | [85] |
| Achyranthes bidentata | p-p38 MAPK↓, p-JNK↓ TNF-α↓, IL-1β↓, IL-6↓ | 16 days after Aβ1–40-induced AD | [84] | |
| Rosmarinic acid | p-JNK↓, p-c-Jun↓ | 8 months in the triple-transgenic mouse model of AD | [65] | |
| Safflower yellow | C. tinctorius L. | Arg1↑, BDNF ↑, TrkB ↑, p-ERK1/2↑ iNOS↓ | 3 months in APP/PS1 transgenic mice | [78] |
| Major Ingredients | Isolated from Medicinal Herbs | Restoring Synaptic Dysfunction Activities | Models | References |
|---|---|---|---|---|
| Berberine | IEG mRNA & protein↑, Arc mRNA & protein↑ | 7 weeks after D-galactose-induced AD | [88] | |
| Xanthoceras sorbifolia | PSD-95↑, BDNF↑, p-TrkB/TrkB↑, RhoA↓, ROCK2↓ | 18 days after Aβ25–35-induced AD | [94] | |
| Daucosterol palmitate | Alpinia oxyphylla Miq. | SYP↑ | 14 days after Aβ1–42-induced AD | [92] |
| Catalpol | Rehmanniae Radix | dynamin 1↑, SYP↑, PSD-95↑, MAP-2↑ | 2 months in aged rats (23–24 months old) | [90] |
| Icariin | Epimedium brevicornum Maxim | PSD-95↑, BDNF↑, TrkB↑, Akt↑, CREB↑ | 28 days after Aβ1–42-induced AD | [98] |
| Galantamine | Galanthus woronowii | AChE↓ | 7 days after Aβ25–35-induced AD | [106] |
| Galantamine | microglial α7 nAChR↑ | 2 weeks after Aβ42-induced AD | [107] | |
| Galantamine | AChE↓, GSH-Px↑, caspase-9 activity↓, caspase-3 activity↓ | 57 days in the transgenic Drosophila model of AD | [109] | |
| Gastrodia elata Blume | ChAT↑, AChE↓ | 52 days after Aβ25–35-induced AD | [101] | |
| Bajijiasu | Morinda officinalis | ACh↑, AChE↓ | 25 days after Aβ25–35-induced AD | [23] |
| Lychee seed extract | Litchi chinensis | AChE↓ | 8 weeks in a rat model of T2DM and AD | [112] |
| GJ-4 | G. jasminoides J. Ellis | ACh↑, AChE↓ | 10 days after Aβ25–35-induced AD | [56] |
| Lignans | S. chinensis Baill | ACh↑ | 1 week in AD rats | [11] |
| β-Asarone | Acori graminei Rhizoma | CaMKIIα↑, p-CREB↑, Bcl-2↑ | 4 months in APP/PS1 mice | [117] |
| Oleanolic acid | Ligustrum lucidum | NMDAR2B↑, CaMKII↑, PKC↑, BDNF↑, TrkB↑, CREB↑ | 28 days after Aβ25–35-induced AD | [116] |
| Major Ingredients | Isolated from Medicinal Herbs | Anti-Apoptotic Activities | Models | References |
|---|---|---|---|---|
| Icariin | E. brevicornum Maxim | Bcl-2/Bax↑, NF-κB↓,p-ERK1/2/ERK1/2↓, p-p38 MAPK/p38 MAPK↓, p-JNK/JNK↓ | 20 days after IBO-induced AD | [122] |
| Butylphthalide | p38 MAPK mRNA & protein↓ | 30 days after Aβ1–42-induced AD | [10] | |
| Tinospora sinensis | p-PI3K/PI3K↑, p-Akt/Akt↑ | 21 days after Aβ1–40-induced AD | [125] | |
| Icariside II | E. brevicornum Maxim | BDNF↑, TrkB↑, p-CREB/CREB↑ | 5 days after Aβ25–35-induced AD | [29] |
| β-asarone | Acorus tatarinowii Schott | ASK 1↓, p-MKK7↓, p-c-Jun↓, Bad mRNA & protein↓, Bax mRNA & protein↓, cleaved caspase-9 mRNA & protein↓ | 28 days of Aβ1–42-induced AD | [119] |
| Genistein | Bax↓, cyt c↓, caspase-3↓ | 49 days after Aβ25–35-induced AD | [15]. | |
| DMDD | Averrhoa carambola L. | Bcl-2/Bax↑, cleaved caspase-9↓, cleaved caspase-3↓ | 21 days in APP/PS1 transgenic AD mice | [133] |
| Scutellarein | Scutellaria baicalensis | Bcl-2↑, Bax↓, caspase-3↓, nucleus NF-κB↓ | 28 days after Aβ-induced AD | [134] |
| Ginsenoside Rg3 | P. ginseng C. A. Meyer | Bcl-2↑, Bax↓, caspase-9↓, caspase-3↓, AIF↓ | 60 days after D-galactose-induced AD | [57] |
| Tetramethylpyrazine | Ligusticum wallichii | SSTR4↑, CUL4B↓ | 30 days in APP/PS1 transgenic mice | [8] |
| Notoginsenoside R2 | P. notoginseng | SOX8↑, β-catenin↑, cleaved caspase-3↓, COX-2↓ | 20 weeks after Aβ25–35-induced AD | [139] |
| Crocin | Crocus sativus L. | GRP78↓, CHOP↓, Bax↓, caspase-3↓ | 14 days after Aβ25–35-induced AD | [140] |
| Schisandrin | S. chinensis Baill | GRP78↓, CHOP↓, cleaved caspase-12↓ | 14 days after STZ-induced AD | [72] |
| Euxanthone | Polygala caudate | Bcl-2/Bax↑, LC3B-II↑ | 16 days after Aβ1–42-induced AD | [145] |
| Icariin | E. brevicornum Maxim | p-Akt↑, LC3-II/LC3-I↓, Beclin-1↓, Cathepsin D (neurofibrillary degeneration marker) ↓ | 5 days after Aβ1–42-induced AD | [144] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, Y.-T.; Kao, S.-T.; Cheng, C.-Y. Medicinal Herbs and Their Derived Ingredients Protect against Cognitive Decline in In Vivo Models of Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 11311. https://doi.org/10.3390/ijms231911311
Tsai Y-T, Kao S-T, Cheng C-Y. Medicinal Herbs and Their Derived Ingredients Protect against Cognitive Decline in In Vivo Models of Alzheimer’s Disease. International Journal of Molecular Sciences. 2022; 23(19):11311. https://doi.org/10.3390/ijms231911311
Chicago/Turabian StyleTsai, Yueh-Ting, Shung-Te Kao, and Chin-Yi Cheng. 2022. "Medicinal Herbs and Their Derived Ingredients Protect against Cognitive Decline in In Vivo Models of Alzheimer’s Disease" International Journal of Molecular Sciences 23, no. 19: 11311. https://doi.org/10.3390/ijms231911311