
From Floral Induction to Blooming: The Molecular Mysteries of Flowering in Woody Plants


1
Co-Innovation Center for Sustainable Forestry in Southern China, College of Biology and the Environment, Nanjing Forestry University, Nanjing 210037, China
2
Department of Biology, The Pennsylvania State University, University Park, State College, PA 16802, USA
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(18), 10959; https://doi.org/10.3390/ijms231810959
Submission received: 23 August 2022
/
Revised: 12 September 2022
/
Accepted: 16 September 2022
/
Published: 19 September 2022
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:Flowering is a pivotal developmental process in response to the environment and determines the start of a new life cycle in plants. Woody plants usually possess a long juvenile nonflowering phase followed by an adult phase with repeated flowering cycles. The molecular mechanism underlying flowering regulation in woody plants is believed to be much more complex than that in annual herbs. In this review, we briefly describe the successive but distinct flowering processes in perennial trees, namely the vegetative phase change, the floral transition, floral organogenesis, and final blooming, and summarize in detail the most recent advances in understanding how woody plants regulate flowering through dynamic gene expression. Notably, the florigen gene FLOWERING LOCUS T(FT) and its antagonistic gene TERMINAL FLOWER 1 (TFL1) seem to play a central role in various flowering transition events. Flower development in different taxa requires interactions between floral homeotic genes together with AGL6 conferring floral organ identity. Finally, we illustrate the issues and corresponding measures of flowering regulation investigation. It is of great benefit to the future study of flowering in perennial trees.
1. Introduction
The transition from vegetative growth to flowering in plants is the most critical developmental event during the whole life history [1]. In annual plants such as Arabidopsis, the reproductive transition includes two successive but distinct stages, namely the vegetative phase change (the juvenile-to-adult vegetative transition) and the floral transition (vegetative-to-reproductive transition). Using the excellent model system Arabidopsis and well-developed genetic tools, studies in past decades have extensively unveiled complex genetic networks controlling flowering time. The five major genetically defined pathways refer to the photoperiod, vernalization, the gibberellin pathway, autonomous, and the aging pathway [2,3,4,5].
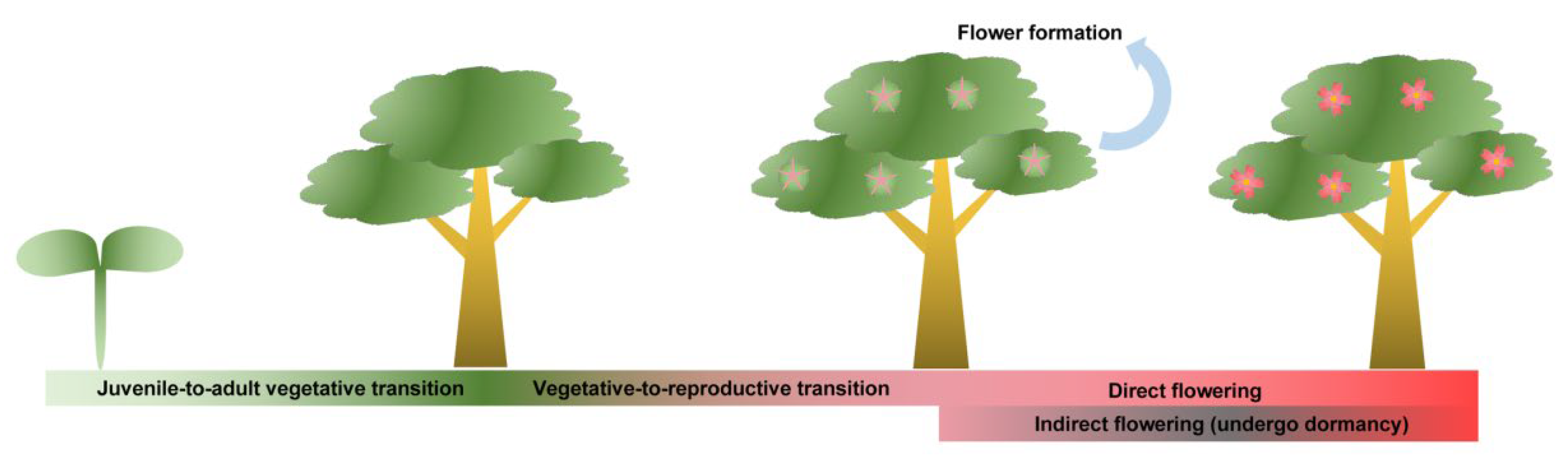
Annual herbaceous plants have quite short life cycles and commonly only spend several months completing the sole reproductive process. Distinctly unlike annual herbs, perennial trees usually have quite long life cycles with relatively complicated reproductive processes (Figure 1) [6]. First, they undergo a much longer juvenile phase before they first acquire flowering ability, usually lasting for several years or even decades (Table 1). Subsequently, adult woody plants show repeated cycling between vegetative and reproductive growth or rather seasonal periodicity of the floral transition. In most perennial woody plants, especially subtropical and temperate deciduous trees (apple, peach, pear, etc.), the flowering cycle spans two successive years. This means that floral induction and blooming are universally separated by a period of rest (usually characterized by the winter dormancy) [7,8,9]. These seasonal-flowering species are also called the “indirect flowering” group. In comparison, “direct flowering” species (mango, jujuba, etc.) will finish their complete reproductive cycle (i.e., the floral transition, flower bud differentiation, and blooming) during a single growing season without a dormancy period [10,11]. Even some rosaceous species (e.g., Rosa chinensis and Rosa hybrida, ‘Little White Pet’) can recurrently flower within one year [12]. What is noteworthy is that some “indirect flowering” trees can directly bloom following flower bud differentiation. For example, a portion of the floral buds of Magnolia × soulangeana ‘Changchun’ is able to skip winter dormancy and bloom in the current summer [13]. The diversity is not only exhibited in the flowering phenologic rhythm but also in flower formation in trees. Normally, the floral organogenesis process mainly involves the regulation of meristem fate and floral organ identity. All facts mentioned above imply that the regulatory mechanism may be different between annual herbaceous and perennial trees, although some flowering pathways are perhaps conserved. The genetic control of flowering time is potentially more complex in perennial plants than that in annuals and is also less understood [14]. Here, on the basis of recent research progress, we review the molecular mechanisms regulating flowering in woody plants, focusing on floral induction, floral organogenesis, and final blooming. It will enhance our understanding of how perennial trees integrate endogenous developmental processes with exogenous environments and decide whether to flower.
2. Molecular Regulation of Flowering Phase Transition
2.1. The Role of PEPB Gene Family in the Flowering Phase Transition
In angiosperms, two different clades of the phosphatidyl ethanolamine-binding protein (PEBP) gene family are well-characterized, namely FLOWERING LOCUS T(FT) and TERMINAL FLOWER 1 (TFL1) [24]. They have been considered the important regulators controlling the time when woody trees become mature and are able to flower, indicating their age-dependent regulatory roles (Table 2). Therefore, FT, the florigen gene, is known as a floral-promoting factor. In early-flowering walnut (Juglans regia), which can flower at the age of 1 or 2 years old, the expression of the FT gene is dramatically higher than that in late-flowering seedlings [25]. Shen et al. isolated two FT orthologs (PsFT1 and PsFT2) from Populus simonii, and over-expressing them in transgenic poplar clone T89 lines led to early flowering that was much earlier than the normal poplar flowering age of 8–20 years [22]. Likewise, the over-expression of Populus deltoides FT2 shortens the juvenile phase of transgenic 717-1B4 poplar (Populus alba × Populus tremula) and controls first-time flowering [26]. On the contrary, FT-RNAi transgenic lines of Jatropha curcas show nonflowering phenotypes [27]. Evidence provided by Tränkner et al. also proves that the MdFT1 gene is responsible for inducing flowering in juvenile plants and plays a conserved function in both annual herbaceous species (Arabidopsis) and perennial woody trees (apple and poplar) [28]. Interestingly, coconut palm (Cocos nucifera) FT is alternatively spliced, and the exclusive presence of the shorter FT variant is highly associated with the early-flowering characteristic in dwarf coconut varieties [29]. In Liriodendron Chinense, an FT alternative splice variant is specific to the super long blooming 1 (slb1) mutant whose inbred offspring have much shorter juvenility (~4 months) than the wild type (usually 8–10 years) [20]. In contrast, TFL1 contributes to the maintenance of the juvenile/vegetative phase and functions as the floral repressor. In transgenic apple (Malus × domestica) seedlings, the expression of endogenous MdTFL1 is suppressed by its antisense RNA, thereby leading to a reduction in vegetative growth and an early-flowering phenotype [30,31]. Similar results were observed in a transgenic European pear ‘Spadona’ (Pyrus communis) genotype by the RNAi silencing of PcTFL1-1 and PcTFL1-2 [32]. Interestingly, Apple latent spherical virus (ALSV)-induced gene silencing of MdTFL1 exhibits a similar strong acceleration of flowering [33]. PopCEN1, a member of the CENTRORADIALIS (CEN)/TFL1 subfamily, controls shoot meristem identity, and PopCEN1-RNAi transgenic poplars with earlier first flowering imply its negative function in the regulation of the juvenile–adult phase transition [34]. Similarly, the CRISPR/Cas9-mediated mutagenesis of the kiwifruit (Actinidia chinensis) CEN-like genes AcCEN4 and AcCEN causes rapid terminal flowering [35]. In addition, overexpressing two TFL1 homologs of dogwood species (Cornus L.) in Arabidopsis wild-type and tfl1 mutant results in delaying flowering time and rescuing the late flowering time phenotype, respectively [36]. All these results have demonstrated that the TFL/CEN1 genes are functionally conserved in eudicots evolution.
In addition to the juvenile–adult phase transition, the seasonal vegetative–reproductive phase transition has also been proven to be regulated by the FT and TFL1 genes (Table 2). Some Rosa species can flower many times within a civil year. A survey of Rosa hybrida screened out floral integrators (FT, AP1, and LFY) as postulated candidates regulating the recurrent flowering character [12]. Compared with seasonal-flowering reference plants, a loss of function or silencing of floral-repressing TFL1-like genes causes perpetual-flowering phenotypes in Rosaceae species, providing additional evidence that TFL1-like genes can also switch the transition from vegetative into reproductive growth [31,32,37]. Jones et al. [14] isolated the FT gene of Eucalyptus globulus subsp. globulus (Myrtaceae) and further examined its expression over a 2-year period using quantitative RT-PCR. The results demonstrate that the expression level of the FT homologue is temporally associated with the annual flower bud initiation (i.e., the annual transition from vegetative to reproductive growth). In adult satsuma mandarin (Citrus unshiu) citrus trees, a seasonal increase in the mRNA level of the citrus FLOWERING LOCUS T homologue CiFT finally stimulates floral induction [38]. Similarly, the drastically increased expression of FT promotes adult ‘Washington’ navel orange (Citrus sinensis) trees to flower [39]. Evidence from Hsu et al. has confirmed that FT2 also plays an additional role in the initiation of seasonal flowering in poplar [26]. In Japanese pear (Pyrus pyrifolia), flower bud formation can be induced by manual photoperiod treatments, and far-red light at 730 nm is the most efficacious wavelength. During this process, PpTFL1 is downregulated, while PpFT1a is positively correlated with flower bud formation [40]. The expression of two copies of the MdTFL1 gene rapidly decreases during floral induction in apple, and ectopic expression delays flowering time in Arabidopsis [41].
As a well-known flowering activator, the FT gene seems not only to play an important role in floral induction but also function in (perpetual) flowering (Table 2). Meng et al. found that the distinct flowering characteristic in Chinese jujube (Ziziphus jujuba) is dominantly regulated by the photoperiod pathway. ZjFT is induced by an unregulated photoperiod and is highly expressed before flowering [11]. To a certain extent, research on the blueberry provided extra experimental evidence, namely that the over-expression of VcFT cloned from Vaccinium corymbosum in ‘Aurora’ results in the continuous occurrence of flower bud formation, flowering, and skipping the normal dormancy stage [42]. What is noteworthy is that some “indirect flowering” trees can directly bloom following flower bud differentiation. It is a very common phenomenon occurring in magnoliaceous species. For example, the slb1 mutant of L. Chinense has a specific FT splice variant that is perhaps causal to perpetual flowering [20]. In tree peony (Paeonia suffruticosa), the upregulation of some flowering-related genes, such as FT, is associated with reblooming [43].
2.2. MicroRNAs in Flowering Phase Transition
Plant microRNAs are endogenous ~21 nt small noncoding RNA molecules that can regulate target gene expression via mRNA destabilization and translational inhibition (Aukerman and Sakai, 2003; Jones-Rhoades et al. 2006). In Arabidopsis, the interactions between the miR156 and miR172 family are considered to play an important role in the juvenile-to-adult vegetative phase transition via mediating the age pathway [44]. They function as inhibitors of the target SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) and APETALA2 (AP2)-like genes, respectively [45]. These two evolutionarily conserved miRNAs are also responsible for the vegetative phase transition in perennial woody plants [46]. miR156 is highly expressed in young seedlings and decreases with aging in some forest species (Acacia confusa, Acacia colei, E. globulus, Hedera helix, Quercus acutissima, and Populus × canadensis), while the expression pattern of miR172 is completely inverse [47]. Similar results have been observed in fruit trees as well as in ornamental flowers, such as apple (M. × domestica) [48], Chinese crabapple (Malus hupehensis) [49], kiwifruit [50], trifoliate orange (Poncirus trifoliata) [51], mango (Mangifera indica) [52], macadamia (Macadamia integrifolia) [52], Prunus mume [53], and Rhododendron arboreum [54]. Autotetraploid Lycium ruthenicum and its diploid progenitor have late- and early-flowering characteristics, respectively, which might be caused by differential expression levels of miR156-SPLs and miR172-AP2 [55]. Furthermore, the over-expression of miR156 drastically prolongs the juvenile phase in transgenic P. × canadensis [47]. Conversely, the ectopic over-expression of JcmiR172a from Jatropha curcas significantly reduces vegetative growth time in Arabidopsis and shortens the juvenile stage of transgenic Jatropha when it grows in a subtropical area [19]. Additionally, miRNAs also function in the floral transition. A multiomics analysis revealed a potential role of miRNAs in stimulating the transition from vegetative growth to reproductive growth in M. × soulangeana ‘Chuangchun’, in which the miR172 family and several other novel miRNAs were differentially expressed and integrated into the GA pathway [56]. In the ‘Golden Delicious’ apple tree, several miRNAs have been found to be vegetative-bud-enriched (miR156, miR159, miR398, and miR408) or floral-bud-enriched (miR164, etc.). Correspondingly, target genes, such as SPLs and ARFs, are down- or upregulated and are speculated to control the floral transition [57]. The upregulation of miRNA167h is considered to be associated with late flowering in Prunus sibirica via participating in the trehalose-6-phosphate (Tre6P) signaling pathway [58]. However, it is regrettable that these results obtained from bioinformatics analyses still need further experimental verification.
3. Regulation Mechanism of Floral Organogenesis
Flower formation is a key regulatory event from floral induction to final blooming in angiosperms, which marks the beginning of the reproductive phase of development [59]. The first developmental transition morphologically reflects that the shoot apical meristem (SAM) transits into the floral meristem (FM), which requires the activity of the LEAFY (LFY) gene [60]. Previous studies of flowering trees have shown that LFY homologues are highly expressed in flower buds in poplar and hickory [61,62]. Further temporal expression analyses of fruit trees (avocado, mango, peach, etc.) have demonstrated that LFY homologues are initiated and highly expressed in the early stages of the floral transition and floral organ differentiation [63,64,65]. The ectopic expression of LFY homologues from Cedrela fissilis, peach, and fig (Ficus carica) rescues the floral defect phenotype in Arabidopsis lfy mutants [64,66,67]. The RNAi-mediated suppression of LFY in P. alba female 6K10 clones results in floral knockdown phenotypes showing the presence of undeveloped carpels that lack stigmatic structures [68,69]. Similarly, in JcLFY-silenced Jatropha curcas, the presence of bracts and shoot buds are observed in abnormal inflorescences, and abnormal flowers are surrounded by 20 sepaloid organs [27]. These results have uncovered the crucial role of the LFY gene in the determination of meristem identity.
Once SAM transits into FM, floral organ primordia arises, and the floral homeotic genes are responsible for floral organ identity during floral organogenesis. According to the interpretation of gene functions learned from floral mutants, several floral homeotic genes and miRNAs have been identified. On the basis of studies in the model plants A. thaliana and Antirrhinum majus, Coen and Meyerowitz [70] summarized and proposed the classic ABC model to explain the identity of successive floral elements. In Arabidopsis, these floral homeotic genes refer to APETALA1 (AP1), APETALA2 (AP2), APETALA3 (AP3), PISTALLATA (PI), and AGAMOUS (AG). A- (AP1 and AP2) and C- (AG) class genes control calyx and carpel formation, respectively. The interaction of A- and B- (AP3 and PI) class genes is responsible for petal development, while stamens develop as a result of the synergetic activity of B- and C-class genes. A revised vision of the ‘ABC’ model, known as the ‘ABCE’ model (Figure 2), has been proposed in combination with a further understanding of SEPALLATA (SEP) gene function [71]. In addition, D-class genes, including SEEDSTICK (STK), SHATTERPROOF1 (SHP1), and SHATTERPROOF2 (SHP2), are sufficient to induce ovule development [72]. The genetic mechanism underlying the floral organogenesis of most angiosperm trees can be interpreted under the framework of the ‘ABC(E)’ model. Considering difficulties in establishing a genetic transformation system of woody species, the ectopic expression of floral homeotic genes in wild-type or corresponding mutants of model plants (i.e., Arabidopsis and tobacco) has been widely applied to investigate gene function. For example, Lemmetyinen et al. isolated two MADS-box genes from silver birch (Betula pendula), namely BpMADS1 and BpMADS6, which are homologues of AtSEP3 and AtAG [73]. Ectopic expression showed detective petals in both 35S::BpMADS6 transgenic Arabidopsis and tobacco. As expected, 35S::BpMADS1 causes global flower defects, which is consistent with its E function. Both PI genes from Catalpa bungei and Argania spinosa rescue petal and stamen identity when they are ectopically expressed in an Arabidopsis pi-1 mutant [74,75]. RNAi-mediated gene silencing is another more effective and direct approach. For example, the RNAi-AG sweetgum trees (Liquidambar styraciflua) have modified inflorescence and floral morphology, with anthers and carpels converting to flat leaf-like structures [76]. The expression suppression of AG genes caused by RNAi finally leads to ‘carpel-inside-carpel’ phenotypes in the poplar clone 6K10 (P. alba) [77]. However, in the basal angiosperms and Magnoliids, floral organs are not clearly divided into different types, all of which exhibit gradual morphological transition (e.g., Amborella trichopoda and Persea americana) [78,79]. These undefined floral architectures might be attributed to a variant of the ABC model, namely ‘the fading borders model’ (Figure 2). This model is characterized by expression domains of floral identity genes that partially overlap with each other, and their expression levels are weaker in the outermost and innermost margins of the expression domains [80,81,82]. The expression patterns of floral homeotic genes determined by quantitative real-time PCR (qRT-PCR) in several Magnolia species provide more valid evidence. For example, AG is expressed in the perianth elements of M. stellata and shows centrifugally reduced activity [83]. In M. grandiflora, a weak expression of AP3 was detected in the carpels [84]. Further gene cloning and ectopic expression experiments revealed that A/B/C-class floral homeotic genes have conserved biological functions in basal angiosperm trees and in core eudicots [85,86,87,88,89]. The AG gene from M. stellata can rescue carpel defects rather than stamen in an Arabidopsis ag-1 mutant. Nevertheless, the ectopic expression of AG alternative splice variants that lack C function causes changes in the perianth elements of wild-type Arabidopsis [87]. Over-expressing M. wufengensis AG in Arabidopsis results in the homeotic conversion of petals into stamenoid organs, indicating its C function [85]. Both AP3 and PI from M. wufengensis can partially rescue the loss of function in the corresponding mutant [86,88].
Interestingly, beyond floral homeotic genes, AGL6-like genes are identified as extra regulators involved in specifying floral organ identity [90]. Members of the ancient AGL6 subfamily of plant MADS-box genes possess diverse functions in different taxa. On one hand, AGL6 genes perform the ‘E’ function, the same as well-characterized SEP genes in herbaceous plants (petunia, rice, and maize). On another hand, they are also considered to be responsible for the development of floral organs in the first whorl, which means they possess ‘A’ function, at least partially [91]. Studies in both gymnosperms and some basal angiosperms have enriched our understanding of how AGL6 genes regulate sexual organ development in woody plants [83,92,93]. When ectopically expressing the CpAGL6 gene from wintersweet (Chimonanthus praecox), transgenic plants show no ectopic floral organs but have abnormal stamen and carpel development, indicating its potential ‘E’ function [93]. Conversely, the role of the A-class gene function in Magnolia species can be assumed by AGL6 (proposed by Soltis et al. [81] and further proved by Ma et al. [94] through genetic experiments). Li et al. found that the AP1 gene from M. wufengensis cannot restore the sepal and petal formation of Arabidopsis ap1 mutants, revealing the absence of A function [89]. As expected, the M. wufengensis AGL6-2 gene can preferentially regulate tepal morphogenesis [94].
Except structural genes, miR172 has been considered to regulate floral organ identity by directly targeting the AP2 gene. In peach (Prunus persica), an AP2 transcription-factor-encoding gene (Prupe.6G242400) was screened out as the candidate regulating single or double traits through high-resolution linkage mapping. Further study showed that a deletion spanning the miR172 binding site confers AP2 gene miR172 resistance, resulting in double-flower formation [95]. Similarly, in kiwifruit alternative splicing leads an AP2 transcript to lose the miR172 targeting site, thereby escaping miR172-mediated cleavage. Finally, abnormal AP2 accumulation results in multiple perianth whorls and extended petaloid features in the mutant ‘Pukekohe dwarf’ [50]. In addition, a transposable element insertion event leads to the creation of an miR172-resistant RcAP2-like variant, which is highly associated with the double flower phenotype in roses (Rosa chinensis) [96].
4. DAM/SVP Genes Associated with Dormancy and “Indirect Flowering”
Dormancy is the representative event during the flowering cycle in “indirect flowering” deciduous trees and a survival strategy to escape the deleterious effects of winter temperatures [97]. The dormancy cycle is divided into three different stages, namely endodormancy, ecodormancy, and paradormancy [98]. Endodormancy is induced by daylight shortening and decreasing temperatures during autumn and winter and requires a certain period of chilling accumulation. Once chilling requirements have been fulfilled, floral or vegetative buds will release from endodormancy and enter ecodormancy, where the growth of buds is inhibited by adverse environmental conditions [99]. The maintenance or release of dormancy is under unique genetic control. SHORT VEGETATIVE PHASE (SVP), an Arabidopsis floral regulator, can represses FT transcription and be integrated into the thermosensory pathway [100]. Recently, the SVP gene and its homologue SVP-LIKE (SVL) have also been implicated in regulating flowering and growth–dormancy cycles in perennials. 35S:SVL transgenic poplars exhibit abnormally late floral bud break to a certain degree [101]. Chromatin immunoprecipitation (ChIP) assays showed that aspen SVL binds to a CArG box on the FT1 promoter, directly suppressing FT1 expression [102]. In sweet cherry (Prunus avium), the expression pattern of PavSVP is closely associated with the suppression of flowering during the dormancy period. Ectopically expressing PavSVP in Arabidopsis delays flowering [103]. Kiwifruit SVP2 has been functionally characterized as a repressor of precocious bud break during dormancy through transgenic experiments. The prolonged dormancy duration in transgenic kiwifruit can be overcome by sufficient winter chilling [104]. In addition to SVPs, DORMANCY-ASSOCIATED MADS-BOX (DAM) genes in rosaceous plants are central regulators of dormancy but are clustered separately in the gene tree [105]. The first report was the evergrowing peach mutant: the deletion of six DAM genes led to a complete lack of dormancy [106,107]. The biological function of DAMs seems distinct due to their diverse season-dependent expression patterns during the dormancy cycle (see Figures 1 and 3 in the previous review [108]). Peach (Prunus persica) is a good example for deeply investigating DAM genes. PpeDAM1, 2, 3, and 4 display the pattern with an approximate peak of expression during bud set. Nevertheless, the expression levels of PpeDAM5 and PpeDAM6 increase over the winter, suggesting they are responsible for the maintenance of bud endodormancy [109]. Additional experiments in a manually controlled environment further confirmed that the expression of PpeDAM5 and PpeDAM6 can be induced by photoperiod, temperature, and exogenous cyanamide [109,110]. Endodormancy release in almond cultivars is accompanied by continuously downregulated DAM-like expression levels [111]. In apple (M. × domestica ‘Royal Gala’), MdDAMb plays a similar role in maintaining bud dormancy [8]. The RNAi-mediated repression of all DAM and SVP genes results in evergrowing apple trees with a precocious-flowering phenotype. Compared with wild-type plants, the expression of the MdFT2 gene is elevated in the terminal buds of RNAi lines [112]. Yeast one-hybrid and dual-luciferase transient expression assays provide limited experimental evidence that DAMs can directly bind to the promoter region of the FT2 gene in pear [113]. Quite interestingly, VvDAM-SVPs gene expression is regulated by vvFT in grapevine (Vitis vinifera) [114]. In addition, Zhao K. et al. found protein interactions among DAMs, and their specific expression patterns contribute to endodormancy in P. mume [115]. Importantly, DAM transcription factors integrate ABA signaling, GA biosynthesis, and catabolism, ultimately mediating dormancy and bud break in perennials [105].
5. Epigenetic Modification in Flowering Regulation
Epigenetic regulation plays an important role in plant growth and development as well as in the response to environmental stresses. The control of flowering progression, including bud dormancy release, is a complex process mediated by different types of epigenetic regulation, i.e., DNA methylation, histone modification, chromatin remodeling, and small interference RNAs (siRNAs) (Table 3) [105,116].
Cytosine DNA methylation, a stable epigenetic mark, is associated with a repressed chromatin state and the inhibition of gene expression [117,118]. During plant developmental processes, a DNA hypermethylation–hypomethylation wave often occurs in the promoter region and specifically in the bodies of active genes. In apple and tree peony (P. suffruticosa ‘Luhehong’), dormancy release is induced by chill conditions and accompanied by a decrease in total DNA methylation [119,120]. Additionally, apple flower bud formation is attributed to high expression levels of flowering-related genes (e.g., SOC1, AP1, and SPLs), which are associated with low methylation levels in the gene-body regions [121]. In the apple cultivar ‘Fuji’, dynamic patterns of DNA methylation are associated with mRNA and siRNA expression, and high CG and CHG methylation were forcefully maintained at the early stage of flower induction [122]. Similarly, in basket willow (Salix viminalis), the application of 5-azacytidine (5-azaC), a DNA methylation inhibitor, leads to hypomethylation in leaves at the floral transition stage and thus promotes the floral initiation and subsequent flower growth [123]. As for tree peony, 5-azaC application significantly reduces the DNA methylation level in the PsFT promoter region and induces higher expression of the PsFT gene, thereby triggering flowering [124]. In sweet cherry, Rothkegel et al. found the silencing of the PavMADS1 and 2 genes during cold accumulation and dormancy release is related to DNA methylation and siRNAs [125]. Their recent study indicated that DNA methylation might act as an early response to low temperatures in the endodormancy period, thus regulating gene expression in a genotype-dependent manner [126]. Similar results were also observed in almond (Prunus dulcis) cultivars with early- and late-flowering phenotypes [127]. The DNA methylation levels in the apical buds of chestnut (Castanea sativa) increase and decrease during bud set and bud burst, respectively. In comparison to DNA methylation, an opposite abundance pattern of H4ac coincides with changes in bud dormancy [128], and the histone-modification-related genes HUB2 and GCN5L are differentially expressed in dormant and germinating buds [129]. During the floral transition in azalea, global DNA methylation and H4ac have opposite and particular dynamics, namely increased DNA methylation levels in contrast to decreased H4ac levels [130,131,132].
Recently, more studies have uncovered the role of histone modification in dormancy and flowering regulation. Genome-wide histone modification gene families have been identified in apple, including 71 histone methyltransferases, 44 histone demethylases, 57 histone acetylases, and 26 histone deacetylases, and most of them are involved in and respond to flower induction [133]. The PavDAM5 gene plays an important role in endodormancy maintenance in sweet cherry, and its expression level is positively related to changes over time in H3K4me3 [134]. In the Japanese pear ‘Kosui’, the reduction in active histone mark H3K4me3 contributes to the decreased expression of the DAM homolog PpMADS13-1 towards endodormancy release [135]. Following the demethylation of H3K4 and the deacetylation of H3 in the region of translation start of the DAM6 gene, H3K27me3 in the DAM6 promoter, the coding region, and the second large intron is consistent with the repression of DAM6, which is responsible for dormancy release in peach [136]. Zhu et al. found multiple epigenetic events, including sRNA expression and H3K27me3 and CHH methylation, that affect the dynamic expression of DAM genes, thereby regulating dormancy maintenance and release in peach [137]. Therefore, the DAM1/2/4/5 genes are significantly enriched in H3K27me3 in dormancy-released buds [138]. A study of leafy spurge (Euphorbia esula) revealed that a decreased enrichment of H3K27me3 and an increase in H3K4me3 were observed in the DAM1 promoter region during endodormancy release [139]. However, the DAM genes are not the only target sites of histone modification. Previously, the SVP gene was characterized experimentally as a flowering repressor. The reduction in AcSVP2 expression towards dormancy release in kiwifruit is attributed to a reduction in H3K4me3 and H3ac but not H3K27me3 and H3K9me3 [140]. Different from the DAM/SVP genes, the early bud-break 1 (EBB1) gene encoding an APETALA2/ethylene-responsive factor (AP2/ERF) transcription factor has been identified as a positive regulator of bud break in poplar [141]. It is worth noting that the regulatory mechanism is conserved among wood species. In peach, higher levels of H3K4me3 in the 50-upstream and start codon regions of the PpEBB gene are associated with the induced PpEBB expression level, which might contribute to bud break and flowering [142]. On the contrary, the activatory mark H3K4me3 enrichment in the promoter region of CcMADS19 is probably the cause of its higher expression, which inhibits floral induction in ‘Moncada’ mandarin (Citrus clementina × (C. unshiu × C. nobilis)) [143]. In addition, Fu et al. [144] proposed that bud endodormancy during chilling accumulation in peach is associated with endoplasmic reticulum stress and the unfolded protein response, which is similar to the report on Arabidopsis [145]. During bud dormancy release in hybrid poplar (P. tremula × P. alba), proteins involved in the primary metabolic pathways are differentially acetylated [146].
6. Final Remarks and Perspectives
The transition to flowering marks a key adaptive developmental switch in plants, which impacts their survival and fitness. In perennial woody plants, this process can be divided into four periods, namely the vegetative phase change, the floral transition, flower organ formation, and finally blooming. The latter three periods compose the repeated flowering cycles during the entire life. All studies summarized in this review have progressively deciphered the complex molecular mechanism of flowering regulation in perennial trees (Figure 2). Notably, the flowering phase transition (including dormancy) is controlled by an antagonistic central gene pair (FT and TFL), miR156/SPL and miR172/AP2 modules, and dormancy-associated genes in response to environmental cues. Therefore, the florigen gene, FT, acts as a central regulator, balancing the exogenous signaling and internal development process in a diverse flowering transition. For “indirect flowering” species, DAM/SVP genes play important roles in dormancy maintenance and release. As for flower development, floral homeotic genes, together with AGL6 and the miR172/AP2 module, are responsible for floral organ identity in trees. In addition, epigenetic regulation involving DNA modification, histone modification, and RNA modification widely participates in flowering events in woody species.
However, there are still some major issues that need to be addressed in the future. First, considering the complexity of woody plants, several genes may execute diverse functions at distinct tissues or developmental stages or are functional redundancies with other paralogs. That means the well-applied ectopic expression in model herbs with simple life histories may not exactly reflect gene function. Natural mutants of perennial woody plants will provide a good chance to investigate the gene function associated with flower development and flowering time regulation. Nonetheless, how to ingeniously select mutant–control pairs is still worth pondering. Except for natural mutant individuals such as Liriodendron slb1 and evergrowing peach, mutational buds/branches from the original mother trees are also welcome. For example, Magnolia × soulangeana ‘Changchun’, whose floral buds can bloom in two distinct seasons, provides a compelling case for investigating dormancy and flowering as well as exploring flower development. Beyond common double-flower mutants of, e.g., peach, kiwifruit, and rose, some tree species with unisexual or polygamous flowers can be sufficient to survey their flower organ formation, such as Woonyoungia septentrionalis, Osmanthus, and persimmon (Diospyros kaki). However, the spontaneous mutation of gene sequences can rarely create ideal mutants with remarkable morphology differences. The reason lies in that the rate of the production of quasineutral, potentially adaptive genetic variance in quantitative characters is an order of magnitude smaller than the total mutational variance because mutations with large phenotypic effects tend to be strongly detrimental. The artificial induction of mutation through radiation (UV, X-ray, radioisotope, etc.), chemical mutagens (alkylating agent, nucleoside analog, NaN3, and colchicine), environment, and virus infection can accelerate mutation rates in order to generate target mutants in a very short period. This will benefit the development of forward genetic approaches in woody plants. Second, the lack of a stable regeneration system in vitro and a genetic transformation system in most woody species inhibits the development of reverse genetics and brings many more challenges in exploring specific genes related to flowering. To a great extent, plant somatic embryogenesis depends on species genotypes. Recently, using WUSCHEL2 (WUS2) and BABY BOOM (BBM) gene, the morphogene-assisted transformation (MAT) has been proven to overcome genotype-dependent disorder in somatic embryogenesis in crops [147]. It might be an effective attempt to introduce this transformation system in order to make its genetic modification available in woody plants. On the other hand, a tobacco rattle virus (TRV)-dependent delivery system can help further gene editing by bypassing tissue culture [148]. Once the difficulties in stable regeneration are overcome in the era of the genome editing technology, the construction of tree early-flowering knockout or knockdown mutants using the CRISPR-Cas9 system will be expected to directly and clearly authenticate their function. Additionally, the publishing of the reference genome makes it easier to understand variation information (e.g., SNP, indel, CNV, and SV) and will also be the genetic basis of genome-wide surveys (WGBS, ChIP-seq, RIP-seq, etc.), thereby shedding light on the molecular mechanism underlying flowering regulation in woody plants.
Author Contributions
Z.Y. and L.S. conceived and wrote the original manuscript. T.N. and Y.C. collected and analyzed studies and illustrated the figures. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the Jiangsu Forestry Science and Technology Innovation and Promotion Project (LYKJ [2019]44), the Postgraduate Research and Practice Innovation Program of Jiangsu Province (Grant Number SJKY19_0917), A Project Funded by the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the Doctorate Fellowship Foundation of Nanjing Forestry University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article, as no datasets were generated or analyzed during the current study.
Conflicts of Interest
All authors declare that they have no conflict of interest.
References
- Rosas, U.; Mei, Y.; Xie, Q.G.; Banta, J.A.; Zhou, R.W.; Seufferheld, G.; Gerard, S.; Chou, L.; Bhambhra, N.; Parks, J.D.; et al. Variation in Arabidopsis flowering time associated with cis-regulatory variation in CONSTANS. Nat. Commun. 2014, 5, 3651. [Google Scholar] [CrossRef] [PubMed]
- Mouradov, A.; Cremer, F.; Coupland, G. Control of fowering time: Interacting pathways as a basis for diversity. Plant Cell 2002, 14, S111–S130. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, A.; Schmid, M. Regulation of fowering time: All roads lead to Rome. Cell Mol. Life Sci. 2011, 68, 2013–2037. [Google Scholar] [CrossRef] [PubMed]
- Andrés, F.; Coupland, G. The genetic basis of flowering responses to seasonal cues. Nat. Rev. Genet. 2012, 13, 627–639. [Google Scholar] [CrossRef]
- Teotia, S.; Tang, G. To bloom or not to bloom: Role of microRNAs in plant flowering. Mol. Plant 2015, 8, 359–377. [Google Scholar] [CrossRef]
- Albani, M.C.; Coupland, G. Comparative analysis of flowering in annual and perennial plants. Curr. Top. Dev. Biol. 2010, 91, 323–348. [Google Scholar]
- Wang, D.L.; Gao, Z.Z.; Du, P.Y.; Xiao, W.; Tan, Q.P.; Chen, X.D.; Li, L.; Gao, D.S. Expression of ABA metabolism-related genes suggests similarities and differences between seed dormancy and bud dormancy of Peach (Prunus persica). Front. Plant Sci. 2016, 6, 1248. [Google Scholar] [CrossRef]
- Wu, R.M.; Tomes, S.; Karunairetnam, S.; Tustin, S.D.; Hellens, R.P.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. SVP-like MADS box genes control dormancy and budbreak in apple. Front. Plant Sci. 2017, 8, 477. [Google Scholar] [CrossRef]
- Ito, A.; Tuan, P.A.; Saito, T.; Bai, S.L.; Kita, M.; Moriguchi, T. Changes in phytohormone content and associated gene expression throughout the stages of pear (Pyrus pyrifolia Nakai) dormancy. Tree Physiol. 2019, 41, 529–543. [Google Scholar] [CrossRef]
- Meir, M.; Ransbotyn, V.; Raveh, E.; Barak, S.; Tel-Zur, N.; Zaccai, M. Dormancy release and flowering time in Ziziphus jujuba Mill., a “direct flowering” fruit tree, has a facultative requirement for chilling. J. Plant Physiol. 2016, 192, 118–127. [Google Scholar] [CrossRef]
- Meng, X.W.; Li, Y.; Yuan, Y.; Zhang, Y.; Li, H.T.; Zhao, J.; Liu, M.J. The regulatory pathways of distinct flowering characteristics in Chinese jujube. Hortic. Res. 2020, 7, 123. [Google Scholar] [CrossRef]
- Remay, A.; Lalanne, D.; Thouroude, T.; Le Couviour, F.; Oyant, L.H.-S.; Foucher, F. A survey of flowering genes reveals the role of gibberellins in floral control in rose. Theor. Appl. Genet. 2009, 119, 767–781. [Google Scholar] [CrossRef]
- Jiang, Z.; Sun, L.Y.; Wei, Q.; Ju, Y.; Zou, X.; Wan, X.X.; Liu, X.; Yin, Z.F. A new insight into flowering regulation: Molecular basis of flowering initiation in Magnolia × soulangeana ‘Changchun’. Genes 2020, 11, 15. [Google Scholar] [CrossRef]
- Jones, R.C.; Valérie, F.G.H.; Potts, B.M.; Vaillancourt, R.E.; Weller, J.L. Expression of a FLOWERING LOCUS T homologue is temporally associated with annual flower bud initiation in Eucalyptus globulus subsp. globulus (Myrtaceae). Aust. J. Bot. 2011, 59, 756–769. [Google Scholar] [CrossRef]
- Su, M.Y.; Wang, N.; Jiang, S.H.; Fang, H.C.; Xu, H.F.; Wang, Y.C.; Zhang, Z.; Zhang, J.; Xu, L.; Zhang, Z.Y.; et al. Molecular characterization and expression analysis of the critical floral gene MdAGL24-like in red-fleshed apple. Plant Sci. 2018, 276, 189–198. [Google Scholar] [CrossRef]
- Wei, X.J.; Ma, J.L.; Wang, K.; Liang, X.J.; Lan, J.X.; Li, Y.J.; Li, K.X.; Liang, H.Y. Early flowering induction in golden Camellia seedlings and effects of paclobutrazol. HortScience 2018, 53, 1849–1854. [Google Scholar] [CrossRef]
- Velázquez, K.; Agüero, J.; Vives, M.C.; Aleza, P.; Pina, J.A.; Moreno, P.; Navarro, L.; Guerri, J. Precocious flowering of juvenile citrus induced by a viral vector based on citrus leaf blotch virus: A new tool for genetics and breeding. Plant Biotechnol. J. 2016, 14, 1976–1985. [Google Scholar] [CrossRef]
- Jordan, G.J.; Potts, B.M.; Chalmers, P.; Wiltshire, R.J.E. Quantitativegenetic evidence that the timing of vegetative phase change in Eucalyptus globulus ssp. globulusis an adaptive trait. Aust. J. Bot. 2000, 48, 561–567. [Google Scholar] [CrossRef]
- Tang, M.Y.; Bai, X.; Niu, L.J.; Chai, X.; Chen, M.S.; Xu, Z.F. miR172 regulates both vegetative and reproductive development in the perennial woody plant Jatropha curcas. Plant Cell Physiol. 2018, 59, 2549–2563. [Google Scholar] [CrossRef]
- Sheng, Y.; Hao, Z.D.; Peng, Y.; Liu, S.Q.; Hu, L.F.; Shen, Y.B.; Shi, J.S.; Chen, J.H. Morphological, phenological, and transcriptional analyses provide insight into the diverse flowering traits of a mutant of the relic woody plant Liriodendron chinense. Hortic. Res. 2021, 8, 174. [Google Scholar] [CrossRef]
- Yuceer, C.; Kubiske, M.E.; Harkess, R.L.; Land, S.B. Effects of induction treatments on flowering in Populus deltoides. Tree Physiol. 2003, 23, 489–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.L.; Chen, Y.; Su, X.H.; Zhang, S.G.; Pan, H.X.; Huang, M.R. Two FT orthologs from Populus simonii Carrière induce early flowering in Arabidopsis and poplar trees. Plant Cell Tiss. Organ Cult. 2012, 108, 371–379. [Google Scholar] [CrossRef]
- Xing, W.; Wang, Z.; Wang, X.Q.; Bao, M.Z.; Ning, G.G. Over-expression of an FT homolog from Prunus mume reduces juvenile phase and induces early flowering in rugosa rose. Sci. Hortic. 2014, 172, 68–72. [Google Scholar] [CrossRef]
- Karlgren, A.; Gyllenstrand, N.; Kallman, T.; Sundstrom, J.F.; Moore, D.; Lascoux, M.; Lagercrantz, U. Evolution of the PEBP gene family in plants: Functional diversification in seed plant evolution. Plant Physiol. 2011, 156, 1967–1977. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Mo, R.L.; Chen, W.X.; Zhang, Q.L.; Sheng, F.; Wu, C.Y.; Zhang, R.; Luo, Z.R. Identification and comparative analysis of genes and microRNAs involved in the floral transition of the Xinjiang early-flowering walnut (Juglans regia L.). Horticulturae 2022, 8, 136. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Liu, Y.X.; Luthe, D.S.; Yuceer, C. Poplar FT2 shortens the juvenile phase and promotes seasonal flowering. Plant Cell 2006, 18, 1846–1861. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Ke, J.P.; Huang, P.; Fatima, I.; Cheng, T.; Tang, M.Y. Promotion of natural flowers by JcFT depends on JcLFY in the perennial woody species Jatropha curcas. Plant Sci. 2022, 318, 111236. [Google Scholar] [CrossRef]
- Tränkner, C.; Lehmann, S.; Hoenicka, H.; Hanke, M.-V.; Fladung, M.; Lenhardt, D.; Dunemann, F.; Gau, A.; Schlangen, K.; Malnoy, M.; et al. Over-expression of an FT-homologous gene of apple induces early flowering in annual and perennial plants. Planta 2010, 232, 1309–1324. [Google Scholar] [CrossRef]
- Xia, W.; Liu, R.; Zhang, J.; Mason, A.S.; Li, Z.Y.; Gong, S.F.; Zhong, Y.Z.; Dou, Y.J.; Sun, X.W.; Fan, H.K.; et al. Alternative splicing of flowering time gene FT is associated with halving of time to flowering in coconut. Sci. Rep. 2020, 10, 11640. [Google Scholar] [CrossRef]
- Kotoda, N.; Iwanami, H.; Takahashi, S.; Abe, K. Antisense expression of MdTFL1, a TFL1-like gene, reduces the juvenile phase in apple. J. Amer. Soc. Hort. Sci. 2006, 131, 74–81. [Google Scholar] [CrossRef]
- Flachowsky, H.; Szankowski, I.; Waidmann, S.; Peil, A.; Tränkner, C.; Hanke, M.V. The MdTFL1 gene of apple (Malus × domestica Borkh.) reduces vegetative growth and generation time. Tree Physiol. 2012, 32, 1288–1301. [Google Scholar] [CrossRef]
- Freiman, A.; Shlizerman, L.; Golobovitch, S.; Yablovitz, Z.; Korchinsky, R.; Cohen, Y.; Samach, A.; Chevreau, E.; Le Roux, P.M.; Patocchi, A.; et al. Development of a transgenic early flowering pear (Pyrus communis L.) genotype by RNAi silencing of PcTFL1-1 and PcTFL1-2. Planta 2012, 235, 1239–1251. [Google Scholar] [CrossRef]
- Sasaki, S.; Yamagishi, N.; Yoshikawa, N. Efficient virus-induced gene silencing in apple, pear and Japanese pear using Apple latent spherical virus vectors. Plant Methods 2011, 7, 15. [Google Scholar] [CrossRef]
- Mohamed, R.; Wang, C.T.; Ma, C.; Shevchenko, O.; Dye, S.J.; Puzey, J.R.; Etherington, E.; Sheng, X.Y.; Meilan, R.; Strauss, S.H.; et al. Populus CEN/TFL1 regulates first onset of flowering, axillary meristem identity and dormancy release in Populus. Plant J. 2010, 62, 674–688. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Wang, T.C.; Voogd, C.; Jeon, S.; Drummond, R.S.M.; Gleave, A.P.; Allan, A.C. Mutagenesis of kiwifruit CENTRORADIALIS-like genes transforms a climbing woody perennial with long juvenility and axillary flowering into a compact plant with rapid terminal flowering. Plant Biotechnol. J. 2019, 17, 869–880. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, J.; Abuahmad, A.; Franks, R.G.; Xie, D.Y.; Xiang, Q.Y. Analysis of two TFL1 homologs of dogwood species (Cornus L.) indicates functional conservation in control of transition to flowering. Planta 2016, 243, 1129–1141. [Google Scholar] [CrossRef]
- Iwata, H.; Gaston, A.; Remay, A.; Thouroude, T.; Jeauffre, J.; Kawamura, K.; Oyant, L.H.-S.; Araki, T.; Denoyes, B.; Foucher, F. The TFL1 homologue KSN is a regulator of continuous flowering in rose and strawberry. Plant J. 2016, 69, 116–125. [Google Scholar] [CrossRef]
- Nishikawa, F.; Endo, T.; Shimada, T.; Fujii, H.; Shimizu, T.; Omura, M.; Ikoma, Y. Increased CiFT abundance in the stem correlates with floral induction by low temperature in Satsuma mandarin (Citrus unshiu Marc.). J. Exp. Bot. 2007, 58, 3915–3927. [Google Scholar] [CrossRef]
- Tang, L.; Lovatt, C.J. Effects of low temperature and gibberellic acid on floral gene expression and floral determinacy in ‘Washington’ navel orange (Citrus sinensis L. Osbeck). Sci. Hortic. 2019, 243, 92–100. [Google Scholar] [CrossRef]
- Ito, A.; Saito, T.; Nishijima, T.; Moriguchi, T. Effect of extending the photoperiod with low-intensity red or far-red light on the timing of shoot elongation and flower-bud formation of 1-year-old Japanese pear (Pyrus pyrifolia). Tree Physiol. 2014, 34, 534–546. [Google Scholar] [CrossRef]
- Zou, X.Y.; Xiang, W.; Zhang, L.Z.; Gao, C.; An, N.; Xing, L.B.; Ma, J.J.; Zhao, C.P.; Zhang, D. Identification of apple TFL1-interacting proteins uncovers an expanded flowering network. Plant Cell Rep. 2021, 40, 2325–2340. [Google Scholar]
- Song, G.Q.; Walworth, A.; Zhao, D.; Ning, J.; Hancock, J.F. The Vaccinium corymbosum FLOWERING LOCUS T-like gene (VcFT): A flowering activator reverses photoperiodic and chilling requirements in blueberry. Plant Cell Rep. 2013, 32, 1759–1769. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.L.; Gao, J.; Xue, J.Q.; Xue, Y.Q.; Li, D.D.; Guan, Y.R.; Zhang, X.X. De novo sequencing of tree peony (Paeonia suffruticosa) transcriptome to identify critical genes involved in flowering and floral organ development. BMC Genom. 2019, 20, 572. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.G.; Jackson, S. Floral induction and flower formation—The role and potential applications of miRNAs. Plant Biotechnol. J. 2015, 13, 282–292. [Google Scholar] [CrossRef]
- Wu, G.; Park, M.Y.; Conway, S.R.; Wang, J.W.; Weigel, D.; Poethig, R.S. The sequential action of miR156 and miR172 regulates developmental timing in Arabidopsis. Cell 2009, 138, 750–759. [Google Scholar] [CrossRef]
- Fang, L.S.; Wang, Y.M. MicroRNAs in woody plants. Front. Plant Sci. 2021, 12, 686831. [Google Scholar] [CrossRef]
- Wang, J.W.; Park, M.Y.; Wang, L.J.; Koo, Y.; Chen, X.Y.; Weigel, D.; Poethig, R.S. MiRNA control of vegetative phase change in trees. PLoS Genet. 2011, 7, e1002012. [Google Scholar] [CrossRef]
- Jia, X.L.; Chen, Y.K.; Xu, X.Z.; Shen, F.; Zheng, Q.B.; Du, Z.; Wang, Y.; Wu, T.; Xu, X.F.; Han, Z.H.; et al. miR156 switches on vegetative phase change under the regulation of redox signals in apple seedlings. Sci. Rep. 2017, 7, 14223. [Google Scholar] [CrossRef]
- Xing, L.B.; Zhang, D.; Li, Y.M.; Zhao, C.P.; Zhang, S.W.; Shen, Y.W.; An, N.; Han, M.Y. Genome-wide identification of vegetative phase transition-associated microRNAs and target predictions using degradome sequencing in Malus hupehensis. BMC Genom. 2014, 15, 1125. [Google Scholar] [CrossRef]
- Varkonyi-Gasic, E.; Lough, R.H.; Moss, S.M.A.; Wu, R.M.; Hellens, R.P. Kiwifruit floral gene APETALA2 is alternatively spliced and accumulates in aberrant indeterminate flowers in the absence of miR172. Plant Mol. Biol. 2012, 78, 417–429. [Google Scholar] [CrossRef]
- Zhang, X.N.; Li, X.; Liu, J.H. Identification of conserved and novel cold-responsive microRNAs in trifoliate orange (Poncirus trifoliata (L.) Raf.) using high-throughput sequencing. Plant Mol. Biol. Rep. 2014, 32, 328–341. [Google Scholar] [CrossRef]
- Ahsan, M.U.; Hayward, A.; Irihimovitch, V.; Fletcher, S.; Tanurdzic, M.; Pocock, A.; Beveridge, C.A.; Mitter, N. Juvenility and vegetative phase transition in tropical/subtropical tree crops. Front. Plant Sci. 2019, 10, 729. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Pan, H.; Wang, J.; Yang, W.R.; Cheng, T.R.; Zhang, Q.X. Identification and profiling of novel and conserved microRNAs during the flower opening process in Prunus mume via deep sequencing. Mol. Genet. Genom. 2014, 289, 169–183. [Google Scholar] [CrossRef]
- Choudhary, S.; Thakur, S.; Najar, R.A.; Majeed, A.; Singh, A.; Bhardwaj, P. Transcriptome characterization and screening of molecular markers in ecologically important Himalayan species (Rhododendron arboreum). Genome 2018, 61, 417–428. [Google Scholar] [CrossRef]
- Rao, S.P.; Li, Y.; Chen, J.H. Combined analysis of microRNAs and target Genes revealed miR156-SPLs and miR172-AP2 are involved in a delayed flowering phenomenon after chromosome doubling in black Goji (Lycium ruthencium). Front. Genet. 2021, 12, 706930. [Google Scholar] [CrossRef]
- Sun, L.Y.; Jiang, Z.; Ju, Y.; Zou, X.; Wan, X.X.; Chen, Y.; Yin, Z.F. A potential endogenous gibberellin-mediated signaling cascade regulated foral transition in Magnolia × soulangeana ‘Changchun’. Mol. Genet. Genom. 2021, 296, 207–222. [Google Scholar] [CrossRef]
- Guo, X.W.; Ma, Z.Y.; Zhang, Z.H.; Cheng, L.L.; Zhang, X.R.; Li, T.H. Small RNA-sequencing links physiological changes and RdDM process to vegetative-to-floral transition in apple. Front. Plant Sci. 2017, 8, 873. [Google Scholar] [CrossRef]
- Xu, W.Y.; Bao, W.Q.; Liu, H.M.; Chen, C.; Bai, H.K.; Huang, M.Z.; Zhu, G.P.; Zhao, H.; Gou, N.N.; Chen, Y.X.; et al. Insights into the molecular mechanisms of late flowering in Prunus sibirica by whole-genome and transcriptome analyses. Front. Plant Sci. 2022, 12, 802827. [Google Scholar] [CrossRef]
- Wellmer, F.; Riechmann, J.L. Gene networks controlling theinitiation of flower development. Trends Genet. 2010, 26, 519–527. [Google Scholar] [CrossRef]
- Weigel, D.; Alvarez, J.; Smyth, D.R.; Yanofsky, M.F.; Meyerowitz, E.M. LEAFY controls floral meristem identity in Arabidopsis. Cell 1992, 69, 843–859. [Google Scholar] [CrossRef]
- Rottmann, W.H.; Meilan, R.; Sheppard, L.A.; Brunner, A.M.; Skinner, J.S.; Ma, C.P.; Cheng, S.P.; Jouanin, L.; Pilate, G.; Strauss, S.H. Diverse effects of overexpression of LEAFY and PTLF, a poplar (Populus) homolog of LEAFY/FLORICAULA, in transgenic poplar and Arabidopsis. Plant J. 2000, 22, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Huang, J.Q.; Huang, Y.J.; Chen, F.F.; Zheng, B.S. Cloning and characterization of a homologue of the FLORICAULA/LEAFY gene in hickory (Carya cathayensis Sarg). Plant Mol. Biol. Rep. 2012, 30, 794–805. [Google Scholar] [CrossRef]
- Acosta-Rangel, A.; Li, R.; Mauk, P.; Santiago, L.; Lovatt, C.J. Effects of temperature, soil moisture and light intensity on the temporal pattern of floral gene expression and flowering of avocado buds (Persea americana cv. Hass). Sci. Hortic. 2021, 280, 109940. [Google Scholar] [CrossRef]
- An, L.J.; Lei, H.J.; Shen, X.J.; Li, T.H. Identification and characterization of PpLFL, a homolog of FLORICAULA/LEAFY in peach (Prunus persica). Plant Mol. Biol. Rep. 2012, 30, 1488–1495. [Google Scholar] [CrossRef]
- Wang, Y.H.; Yu, H.X.; He, X.H.; Lu, T.T.; Huang, X.; Luo, C. Isolation and functional characterization of a LEAFY gene in mango (Mangifera indica L.). Int. J. Mol. Sci. 2022, 23, 3974. [Google Scholar] [CrossRef] [PubMed]
- Dornelas, M.C.; Rodriguez, A.P.M. The tropical cedar tree (Cedrela fissilis Vell., Meliaceae) homolog of the Arabidopsis LEAFY gene is expressed in reproductive tissues and can complement Arabidopsis leafy mutants. Planta 2006, 223, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; An, Y.Y.; Li, J.; Wang, L.J. Cloning and characterization of a homologue of the FLORICAULA/LEAFY gene in Ficus carica L., FcLFY, and its role in flower bud differentiation. Sci. Hortic. 2020, 261, 109014. [Google Scholar] [CrossRef]
- Klocko, A.L.; Brunner, A.M.; Huang, J.; Meilan, R.; Lu, H.W.; Ma, C.; Morel, A.; Zhao, D.Z.; Ault, K.; Dow, M.; et al. Containment of transgenic trees by suppression of LEAFY. Nat. Biotechnol. 2016, 34, 918–922. [Google Scholar] [CrossRef]
- Klocko, A.L.; Goddard, A.L.; Jacobson, J.R.; Magnuson, A.C.; Strauss, S.H. RNAi suppression of LEAFY gives stable floral sterility, and reduced growth rate and leaf size, in field-grown poplars. Plants 2021, 10, 1594. [Google Scholar] [CrossRef]
- Coen, E.S.; Meyerowitz, E.M. The war of the whorls: Genetic interactions controlling flower development. Nature 1991, 353, 31–37. [Google Scholar] [CrossRef]
- Pelaz, S.; Ditta, G.S.; Baumann, E.; Wisman, E.; Yanofsky, M.F. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 2000, 405, 200–203. [Google Scholar] [CrossRef]
- Favaro, R.; Pinyopich, A.; Battaglia, R.; Kooiker, M.; Borghi, L.; Ditta, G.; Yanofsky, M.F.; Kater, M.M.; Colombo, L. MADS-Box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 2003, 15, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Lemmetyinen, J.; Hassinen, M.; Elo, A.; Porali, I.; Sopanen, T. Functional characterization of SEPALLATA3 and AGAMOUS orthologues in silver birch. Physiol. Plant. 2010, 121, 149–162. [Google Scholar] [CrossRef]
- Jing, D.L.; Xia, Y.; Chen, F.J.; Wang, Z.; Zhang, S.G.; Wang, J.H. Ectopic expression of a Catalpa bungei (Bignoniaceae) PISTILLATA homologue rescues the petal and stamen identities in Arabidopsis pi-1 mutant. Plant Sci. 2015, 231, 40–51. [Google Scholar] [CrossRef]
- Louati, M.; Salazar-Sarasua, B.; Roque, E.; Beltrán, J.P.; Hannachi, A.S.; Gómez-Mena, C. Isolation and functional analysis of a PISTILLATA-like MADS-box gene from argan tree (Argania spinosa). Plants 2021, 10, 1665. [Google Scholar] [CrossRef]
- Klocko, A.L.; Brunner, A.M.; Ma, C.; Etherington, E.; Rosenstiel, K.; Magnuson, A.; Taylor, B.J.; JLockwood, T.C.; Covarrubias, N.; Bao, M.Z.; et al. RNAi of AGAMOUS genes in sweetgum alters reproductive organ identity and decreases fruit persistence. Plant Direct 2020, 4, e00225. [Google Scholar] [CrossRef]
- Lu, H.W.; Klocko, A.L.; Brunner, A.M.; Ma, C.; Magnuson, A.C.; Howe, G.T.; An, X.M.; Strauss, S.H. RNAi suppression of AGAMOUS and SEEDSTICK alters floral organ identity and impairs floral organ determinacy, ovule differentiation, and seed-hair development in Populus. New Phytol. 2018, 222, 923–937. [Google Scholar] [CrossRef]
- Buzgo, M.; Soltis, P.S.; Soltis, D.E. Floral developmental morphology of Amborella trichopoda (Amborellaceae). Int. J. Plant Sci. 2004, 165, 925–947. [Google Scholar] [CrossRef]
- Chanderbali, A.S.; Albert, V.A.; Leebens-Mack, J.; Altman, N.S.; Soltis, D.E.; Soltis, P.S. Transcriptional signatures of ancient floral developmental genetics in avocado (Persea americana; Lauraceae). Proc. Natl. Acad. Sci. USA 2009, 106, 8929–8934. [Google Scholar] [CrossRef]
- Luo, H.L.; Chen, S.M.; Jiang, J.F.; Chen, Y.; Chen, F.D.; Teng, N.J.; Yin, D.M.; Huang, C.B. The expression of floral organ identity genes in contrasting water lily cultivars. Plant Cell Rep. 2011, 30, 1909–1918. [Google Scholar] [CrossRef]
- Soltis, D.E.; Chanderbali, A.S.; Kim, S.; Buzgo, M.; Soltis, P.S. The ABC model and its applicability to basal angiosperms. Ann. Bot. 2007, 100, 155–163. [Google Scholar] [CrossRef]
- Soltis, D.E.; Ma, H.; Frohlich, M.W.; Soltis, P.S.; Albert, V.A.; Oppenheimer, D.G.; Altman, N.S.; dePamphilis, C.; Leebens-Mack, J. The floral genome: An evolutionary history of gene duplication and shifting patterns of gene expression. Trends Plant Sci. 2007, 12, 358–367. [Google Scholar] [CrossRef]
- Wróblewska, M.; Dołzbłasz, A.; Zagórska-Marek, B. The role of ABC genes in shaping perianth phenotype in the basal angiosperm Magnolia. Plant Biol. 2016, 18, 230–238. [Google Scholar] [CrossRef]
- Kim, S.; Koh, J.; Yoo, M.-J.; Kong, H.Z.; Hu, Y.; Ma, H.; Soltis, P.S.; Soltis, D.E. Expression of floral MADS-box genes in basal angiosperms: Implications for the evolution of floral regulators. Plant J. 2005, 43, 724–744. [Google Scholar] [CrossRef]
- Wu, W.T.; Chen, F.J.; Jing, D.L.; Liu, Z.X.; Ma, L.Y. Isolation and characterization of an AGAMOUS-Like gene from Magnolia wufengensis (Magnoliaceae). Plant Mol. Biol. Rep. 2012, 30, 690–698. [Google Scholar] [CrossRef]
- Jing, D.L.; Liu, Z.X.; Zhang, B.; Ma, J.; Han, Y.Y.; Chen, F.J. Two ancestral APETALA3 homologs from the basal angiosperm Magnolia wufengensis (Magnoliaceae) can affect flower development of Arabidopsis. Gene 2014, 537, 100–107. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Z.X.; Ma, J.; Song, Y.; Chen, F.J. Alternative splicing of the AGAMOUS orthologous gene in double flower of Magnolia stellata (Magnoliaceae). Plant Sci. 2015, 241, 277–285. [Google Scholar] [CrossRef]
- Liu, W.; Shen, X.L.; Liang, H.W.; Wang, Y.B.; He, Z.Q.; Zhang, D.C.; Chen, F.J. Isolation and functional analysis of PISTILLATA homolog from Magnolia wufengensis. Front. Plant Sci. 2018, 9, 1743. [Google Scholar] [CrossRef] [PubMed]
- Li, C.J.; Chen, L.Y.; Fan, X.N.; Qi, W.J.; Ma, J.; Tian, T.; Zhou, T.; Ma, L.Y.; Chen, F.J. MawuAP1 promotes flowering and fruit development in the basal angiosperm Magnolia wufengensis (Magnoliaceae). Tree Physiol. 2020, 40, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Dreni, L.; Zhang, D.B. Flower development: The evolutionary history and functions of the AGL6 subfamily MADS-box genes. J. Exp. Bot. 2016, 67, 1625–1638. [Google Scholar] [CrossRef] [PubMed]
- Pabón-Mora, N.; Suárez-Baron, H.; Ambrose, B.A.; González, F. Flower development and perianth identity candidate genes in the basal angiosperm Aristolochia fimbriata (Piperales: Aristolochiaceae). Front. Plant Sci. 2015, 6, 1095. [Google Scholar] [CrossRef]
- Mouradov, A.; Glassick, T.V.; Hamdorf, B.A.; Murphy, L.C.; Marla, S.S.; Yang, Y.M.; Teasdale, R.D. Family of MADS-box genes expressed early in male and female reproductive structures of Monterey pine. Plant Physiol. 1998, 117, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.G.; Zhang, Q.; Wang, L.G.; Duan, K.; Pan, A.H.; Tang, X.M.; Sui, S.Z.; Li, M.Y. The AGL6-like gene CpAGL6, a potential regulator of floral time and organ identity in wintersweet (Chimonanthus praecox). J. Plant Growth Regul. 2011, 30, 343–352. [Google Scholar] [CrossRef]
- Ma, J.; Deng, S.X.; Chen, L.Y.; Jia, Z.K.; Sang, Z.Y.; Zhu, Z.L.; Ma, L.Y.; Chen, F.J.; Plomion, C. Gene duplication led to divergence of expression patterns, protein–protein interaction patterns and floral development functions of AGL6-like genes in the basal angiosperm Magnolia wufengensis (Magnoliaceae). Tree Physiol. 2019, 39, 861–876. [Google Scholar] [CrossRef] [PubMed]
- Gattolin, S.; Cirilli, M.; Pacheco, I.; Ciacciulli, A.; da Silva Linge, C.; Mauroux, J.-B.; Lambert, P.; Cammarata, E.; Bassi, D.; Pascal, T.; et al. Deletion of the miR172 target site in a TOE-type gene is a strong candidate variant for dominant double-flower trait in Rosaceae. Plant J. 2018, 96, 358–371. [Google Scholar] [CrossRef] [PubMed]
- François, L.; Verdenaud, M.; Fu, X.P.; Ruleman, D.; Dubois, A.; Vandenbussche, M.; Bendahmane, A.; Raymond, O.; Just, J.; Bendahmane, M. A miR172 target-deficient AP2-like gene correlates with the double flower phenotype in roses. Sci. Rep. 2018, 8, 12912. [Google Scholar] [CrossRef] [PubMed]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2017, 152, 28–36. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, para-, and ecodormancy: Physiological terminology and classification for dormancy research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- Guillamón, J.G.; Dicenta, F.; Sánchez-Pérez, R. Advancing endodormancy release in temperate fruit trees using agrochemical treatments. Front. Plant Sci. 2022, 12, 812621. [Google Scholar] [CrossRef]
- Lee, J.H.; Yoo, S.J.; Park, S.H.; Hwang, I.; Lee, J.S.; Ahn, J.H. Role of SVP in the control of flowering time by ambient temperature in Arabidopsis. Genes Dev. 2007, 21, 397–402. [Google Scholar] [CrossRef]
- Goralogia, G.S.; Howe, G.T.; Brunner, A.M.; Helliwell, E.; Nagle, M.F.; Ma, C.; Lu, H.W.; Goddard, A.L.; Magnuson, A.C.; Klocko, A.L.; et al. Overexpression of SHORT VEGETATIVE PHASE-LIKE (SVL) in Populus delays onset and reduces abundance of flowering in field-grown trees. Hortic. Res. 2021, 8, 167. [Google Scholar] [CrossRef]
- Singh, R.K.; Maurya, J.P.; Azeez, A.; Miskolczi, P.; Tylewicz, S.; Stojkovič, K.; Delhomme, N.; Busov, V.; Bhalerao, R.P. A genetic network mediating the control of bud break in hybrid aspen. Nat. Commun. 2018, 9, 4173. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.Y.; Jiu, S.T.; Xu, Y.; Sabir, I.A.; Wang, L.; Ma, C.; Xu, W.P.; Wang, S.P.; Zhang, C.X. SVP-like gene PavSVP potentially suppressing flowering with PavSEP, PavAP1, and PavJONITLESS in sweet cherries (Prunus avium L.). Plant Physiol. Bioch. 2021, 159, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.M.; Wang, T.C.; Warren, B.A.W.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. Kiwifruit SVP2 gene prevents premature budbreak during dormancy. J. Exp. Bot. 2017, 68, 1071–1082. [Google Scholar] [CrossRef]
- Yang, Q.S.; Gao, Y.H.; Wu, X.Y.; Moriguchi, T.; Bai, S.L.; Teng, Y.W. Bud endodormancy in deciduous fruit trees: Advances and prospects. Hortic. Res. 2021, 8, 139. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Wang, Y.; Fan, S.; Reighard, G.L.; Scorza, R.; Abbott, A.G. A deletion affecting several gene candidates is present in the evergrowing peach mutant. J. Hered. 2004, 95, 436–444. [Google Scholar] [CrossRef]
- Bielenberg, D.G.; Wang, Y.; Li, Z.G.; Zhebentyayeva, T.; Fan, S.H.; Reighard, G.L.; Scorza, R.; Abbott, A.G. Sequencing and annotation of the evergrowing locus in peach [Prunus persica (L.) Batsch] reveals a cluster of six MADS-box transcription factors as candidate genes for regulation of terminal bud formation. Tree Genet. Genomes 2008, 4, 495–507. [Google Scholar] [CrossRef]
- Falavigna, V.S.; Guitton, B.; Costes, E.; Andrés, F. I want to (bud) break free: The potential role of DAM and SVP-like genes in regulating dormancy cycle in temperate fruit trees. Front. Plant Sci. 2019, 9, 1990. [Google Scholar] [CrossRef]
- Li, Z.G.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated MADS genes from the EVG locus of peach [Prunus persica (L.) Batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [CrossRef]
- Yamane, H.; Ooka, T.; Jotatsu, H.; Hosaka, Y.; Sasaki, R.; Tao, R. Expressional regulation of PpDAM5 and PpDAM6, peach (Prunus persica) dormancy-associated MADS-box genes, by low temperature and dormancy-breaking reagent treatment. J. Exp. Bot. 2011, 62, 3481–3488. [Google Scholar] [CrossRef] [PubMed]
- Prudencio, A.S.; Hoeberichts, F.A.; Dicenta, F.; Martínez-Gómez, P.; Sánchez-Pérez, R. Identification of early and late flowering time candidate genes inendodormant and ecodormant almond flower buds. Tree Physiol. 2021, 41, 589–605. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.M.; Cooney, J.; Tomes, S.; Rebstock, R.; Karunairetnam, S.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. RNAi-mediated repression of dormancy-related genes results in evergrowing apple trees. Tree Physiol. 2021, 41, 1510–1523. [Google Scholar] [CrossRef] [PubMed]
- Niu, Q.F.; Li, J.Z.; Cai, D.Y.; Qian, M.J.; Jia, H.M.; Bai, S.L.; Hussain, S.; Liu, G.Q.; Teng, Y.W.; Zheng, X.Y. Dormancy-associated MADS-box genes and microRNAs jointly control dormancy transition in pear (Pyrus pyrifolia white pear group) flower bud. J. Exp. Bot. 2015, 67, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Vergara, R.; Noriega, X.; Pérez, F.J. VvDAM-SVPs genes are regulated by FLOWERING LOCUS T (VvFT) and not by ABA/low temperature-induced VvCBFs transcription factors in grapevine buds. Planta 2021, 253, 31. [Google Scholar] [CrossRef]
- Zhao, K.; Zhou, Y.Z.; Ahmad, S.; Yong, X.; Xie, X.H.; Han, Y.; Li, Y.S.; Sun, L.D.; Zhang, Q.X. PmCBFs synthetically affect PmDAM6 by alternative promoter binding and protein complexes towards the dormancy of bud for Prunus mume. Sci. Rep. 2018, 8, 4527. [Google Scholar] [CrossRef]
- Zhebentyayeva, T.N.; Fan, S.H.; Chandra, A.; Bielenberg, D.G.; Reighard, G.L.; Okie, W.R.; Abbott, A.G. Dissection of chilling requirement and bloom date QTLs in peach using a whole genome sequencing of sibling trees from an F2 mapping population. Tree Genet. Genomes 2014, 10, 35–51. [Google Scholar] [CrossRef]
- Klose, R.J.; Bird, A.P. Genomic DNA methylation: The mark and its mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef]
- Kumar, G.; Rattan, U.K.; Singh, A.K. Chilling-mediated DNA methylation changes during dormancy and its release reveal the importance of epigenetic regulation during winter dormancy in apple (Malus × domestica Borkh.). PLoS ONE 2016, 11, e0149934. [Google Scholar] [CrossRef]
- Xin, H.; Zhang, Y.X.; Wang, X.T.; Liu, C.Y.; Feng, W.R.; Gai, S.P. Morphological, anatomical and DNA methylation changes of tree peony buds during chilling induced dormancy release. Plant Physiol. Biochem. 2019, 144, 64–72. [Google Scholar] [CrossRef]
- Xing, L.B.; Li, Y.M.; Qi, S.Y.; Zhang, C.G.; Ma, W.C.; Zou, X.Y.; Liang, J.Y.; Gao, C.; Jia, P.; Shah, K.; et al. Comparative RNA-sequencing and DNA methylation analyses of apple (Malus domestica Borkh.) buds with diverse flowering capabilities reveal novel insights into the regulatory mechanisms of flower bud formation. Plant Cell Physiol. 2019, 60, 1702–1721. [Google Scholar] [CrossRef]
- Fan, S.; Gao, X.H.; Gao, C.; Yang, Y.; Zhu, X.Z.; Feng, W.; Li, R.M.; Tahir, M.M.; Zhang, D.; Han, M.Y.; et al. Dynamic cytosine DNA methylation patterns associated with mRNA and siRNA expression profiles in alternate bearing apple trees. J. Agric. Food Chem. 2019, 67, 5250–5264. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Peng, X.Y.; Yu, Y.C.; Sun, Z.Y.; Han, L. The effects of dna methylation inhibition on flower development in the dioecious plant Salix Viminalis. Forests 2019, 10, 173. [Google Scholar] [CrossRef]
- Sun, K.R.; Xue, Y.Q.; Prijic, Z.; Wang, S.L.; Markovic, T.; Tian, C.H.; Wang, Y.Y.; Xue, J.Q.; Zhang, X.X. DNA demethylation induces tree peony flowering with a low deformity rate compared to gibberellin by inducing PsFT expression under forcing culture conditions. Int. J. Mol. Sci. 2022, 23, 6632. [Google Scholar] [CrossRef]
- Rothkegel, K.; Sánchez, E.; Montes, C.; Greve, M.; Tapia, S.; Bravo, S.; Prieto, H.; Almeida, A.M. DNA methylation and small interference RNAs participate in the regulation of MADS-box genes involved in dormancy in sweet cherry (Prunus avium L.). Tree Physiol. 2017, 37, 1739–1751. [Google Scholar] [CrossRef]
- Rothkegel, K.; Sandoval, P.; Soto, E.; Ulloa, L.; Riveros, A.; Lillo-Carmona, V.; Cáceres-Molina, J.; Almeida, A.M.; Meneses, C. Dormant but active: Chilling accumulation modulates the epigenome and transcriptome of Prunus avium during bud dormancy. Front. Plant Sci. 2020, 11, 1115. [Google Scholar] [CrossRef]
- Prudencio, Á.S.; Werner, O.; Martínez-García, P.J.; Dicenta, F.; Ros, R.M.; Martínez-Gómez, P. DNA methylation analysis of dormancy release in almond (Prunus dulcis) flower buds using pi-genotyping by sequencing. Int. J. Mol. Sci. 2018, 19, 3542. [Google Scholar] [CrossRef]
- Santamaría, M.E.; Hasbún, R.; Valera, M.J.; Meijón, M.; Valledor, L.; Rodríguez, J.L.; Toorop, P.E.; Cañal, M.J.; Rodríguez, R. Acetylated H4 histone and genomic DNA methylation patterns during bud set and bud burst in Castanea sativa. J. Plant Physiol. 2009, 166, 1360–1369. [Google Scholar] [CrossRef]
- Santamaría, M.E.; Rodríguez, R.; Cañal, M.J.; Toorop, P.E. Transcriptome analysis of chestnut (Castanea sativa) tree buds suggests a putative role for epigenetic control of bud dormancy. Ann. Bot. 2011, 108, 485–498. [Google Scholar] [CrossRef]
- Meijón, M.; Valledor, L.; Santamaría, M.E.; Testillano, P.S.; Risueño, C.; Rodríguez, R.; Feito, I.; Cañal, M.J. Epigenetic characterization of the vegetative and floral stages of azalea buds: Dynamics of DNA methylation and histone H4 acetylation. J. Plant Physiol. 2009, 166, 1624–1636. [Google Scholar] [CrossRef]
- Meijón, M.; Feito, I.; Valledor, L.; Rodríguez, R.; Cañal, M.J. Dynamics of DNA methylation and Histone H4 acetylation during floral bud differentiation in azalea. BMC Plant Biol. 2010, 10, 10. [Google Scholar] [CrossRef]
- Meijón, M.; Cañal, M.J.; Valledor, L.; Rodríguez, R.; Feito, I. Epigenetic and physiological effects of gibberellin inhibitors and chemical pruners on the floral transition of azalea. Physiol. Plant. 2011, 141, 276–288. [Google Scholar] [CrossRef]
- Fan, S.; Wang, J.; Lei, C.; Gao, C.; Yang, Y.; Li, Y.M.; An, N.; Zhang, D.; Han, M.Y. Identification and characterization of histone modification gene family reveal their critical responses to flower induction in apple. BMC Plant Biol. 2018, 18, 173. [Google Scholar] [CrossRef]
- Vimont, N.; Quah, F.X.; Schöepfer, D.G.; Roudier, F.; Dirlewanger, E.; Wigge, P.A.; Wenden, B.; Cortijo, S. ChIP-seq and RNA-seq for complex and low-abundance tree buds reveal chromatin and expression co-dynamics during sweet cherry bud dormancy. Tree Genet. Genomes 2020, 16, 9. [Google Scholar] [CrossRef]
- Saito, T.; Bai, S.L.; Imai, T.; Ito, A.; Nakajima, I.; Moriguchi, T. Histone modification and signalling cascade of the dormancy-associated MADS-box gene, PpMADS13-1, in Japanese pear (Pyrus pyrifolia) during endodormancy. Plant Cell Environ. 2015, 38, 1157–1166. [Google Scholar] [CrossRef] [PubMed]
- Leida, C.; Conesa, A.; Llácer, G.; Badenes, M.L.; Ríos, G. Histone modifications and expression of DAM6 gene in peach are modulated during bud dormancy release in a cultivar-dependent manner. New Phytol. 2012, 193, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Chen, P.Y.; Zhong, S.L.; Dardick, C.; Callahan, A.; An, Y.Q.; van Knocker, S.; Yang, Y.Z.; Zhong, G.Y.; Abbott, A.; et al. Thermal-responsive genetic and epigenetic regulation of DAM cluster controlling dormancy and chilling requirement in peach floral buds. Hortic. Res. 2020, 7, 114. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente, L.; Conesa, A.; Lloret, A.; Badenes, M.L.; Ríos, G. Genome-wide changes in histone H3 lysine 27 trimethylation associated with bud dormancy release in peach. Tree Genet. Genomes 2015, 11, 45. [Google Scholar] [CrossRef]
- Horvath, D.P.; Sung, S.; Kim, D.; Chao, W.; Anderson, J. Characterization, expression and function of DORMANCY ASSOCIATED MADS-BOX genes from leafy spurge. Plant Mol. Biol. 2010, 73, 169–179. [Google Scholar] [CrossRef]
- Wu, R.M.; Wang, T.C.; Richardson, A.C.; Allan, A.C.; Macknight, R.C.; Varkonyi-Gasic, E. Histone modification and activation by SOC1-like and drought stress-related transcription factors may regulate AcSVP2 expression during kiwifruit winter dormancy. Plant Sci. 2019, 281, 242–250. [Google Scholar] [CrossRef]
- Yordanov, Y.S.; Ma, C.; Strauss, S.H.; Busov, V.B. EARLY BUD-BREAK 1 (EBB1) is a regulator of release from seasonal dormancy in poplar trees. Proc. Natl. Acad. Sci. USA 2014, 111, 10001–10006. [Google Scholar] [CrossRef]
- Tuan, P.A.; Bai, S.L.; Saito, T.; Imai, T.; Ito, A.; Moriguchi, T. Involvement of EARLY BUD-BREAK, an AP2/ERF transcription factor gene, in bud break in Japanese pear (Pyrus pyrifolia Nakai) lateral flower buds: Expression, histone modifications and possible target genes. Plant Cell Physiol. 2016, 57, 1038–1047. [Google Scholar] [CrossRef] [Green Version]
- Agustí, A.; Mesejo, C.; Muñoz-Fambuena, N.; Vera-Sirera, F.; de Lucas, M.; Martínez-Fuentes, A.; Reig, C.; Iglesias, D.J.; Primo-Millo, E.; Blázquez, M.A. Fruit-dependent epigenetic regulation of flowering in Citrus. New Phytol. 2020, 225, 376–384. [Google Scholar] [CrossRef]
- Fu, X.L.; Xiao, W.; Wang, D.L.; Chen, M.; Tan, Q.P.; Li, L.; Chen, X.D.; Gao, D.S. Roles of endoplasmic reticulum stress and unfolded protein response associated genes in seed stratification and bud endodormancy during chilling accumulation in Prunus persica. PLoS ONE 2014, 9, e101808. [Google Scholar] [CrossRef]
- Song, Z.T.; Sun, L.; Lu, S.J.; Tian, Y.K.; Ding, Y.; Liu, J.X. Transcription factor interaction with COMPASS-like complex regulates histone H3K4 trimethylation for specific gene expression in plants. Proc. Natl. Acad. Sci. USA 2015, 112, 2900–2905. [Google Scholar] [CrossRef]
- Liao, X.L.; Li, Y.; Hu, Z.Z.; Lin, Y.; Zheng, B.; Ding, J.H. Poplar acetylome profiling reveals lysine acetylation dynamics in seasonal bud dormancy release. Plant Cell Environ. 2021, 44, 1830–1845. [Google Scholar] [CrossRef]
- Aregawi, K.; Shen, J.Q.; Pierroz, G.; Sharma, M.K.; Dahlberg, J.; Owiti, J.; Lemaux, P.G. Morphogene-assisted transformation of Sorghum bicolor allows more efficient genome editing. Plant Biotechnol. J. 2021, 20, 748–760. [Google Scholar] [CrossRef]
- Ellison, E.E.; Nagalakshmi, U.; Gamo, M.E.; Huang, P.J.; Dinesh-Kumar, S.; Voytas, D.F. Multiplexed heritable gene editing using RNA viruses and mobile single guide RNAs. Nat. Plants 2020, 6, 620–624. [Google Scholar] [CrossRef]
Figure 1.
Schematic presentation of flowering phenology in woody trees. The juvenile-to-adult vegetative transition refers to a long period before woody trees first acquire flowering ability; the vegetative-to-reproductive transition commonly shows repeated cycling or rather seasonal periodicity, indicating flower formation follows the floral transition; “Direct flowering” refers to trees that finish their complete reproductive cycle during a single growing season without a dormancy period, while “indirect flowering” means the trees undergo winter dormancy before final blooming.
Figure 1.
Schematic presentation of flowering phenology in woody trees. The juvenile-to-adult vegetative transition refers to a long period before woody trees first acquire flowering ability; the vegetative-to-reproductive transition commonly shows repeated cycling or rather seasonal periodicity, indicating flower formation follows the floral transition; “Direct flowering” refers to trees that finish their complete reproductive cycle during a single growing season without a dormancy period, while “indirect flowering” means the trees undergo winter dormancy before final blooming.

Figure 2.
Diagram illustrating the molecular model underlying flowering regulation in perennial trees, which incorporates important/central genes and microRNAs as well as epigenetic modification. Black solid arrows indicate gene regulation; green dotted arrows indicate epigenetic modification participating in gene regulatory modules; brown dotted arrows represent responses to environmental cues.
Figure 2.
Diagram illustrating the molecular model underlying flowering regulation in perennial trees, which incorporates important/central genes and microRNAs as well as epigenetic modification. Black solid arrows indicate gene regulation; green dotted arrows indicate epigenetic modification participating in gene regulatory modules; brown dotted arrows represent responses to environmental cues.


{kind=link}
{kind=link}
Table 1.
Time of juvenile-to-adult vegetative transition in several representative woody trees.
| Tree Species | Juvenile-to-Adult Vegetative Transition | Reference |
|---|---|---|
| Malus × domestica | 4–8 years | [15] |
| Camellia chrysantha | 6–8 years | [16] |
| Citrus spp. | 6–10 years | [17] |
| Eucalyptus globulus ssp. globulusis | 1–5 years | [18] |
| Jatropha curcas | 5 years in subtropical areas | [19] |
| Liriodendron Chinense | 8–10 years | [20] |
| Populus deltoides var. deltoides | 5–10 years | [21] |
| poplar T89 (Populus tremula L. × P. tremuloides) | 8–20 years | [22] |
| Rosa rugosa ‘Bao White’ | >3 years | [23] |
Table 2.
Summary of PEPB gene family mediated flowering regulation in woody plants.
| Species | Technique | Regulators | Phase Transition | References |
|---|---|---|---|---|
| Actinidia chinensis | CRISPR/Cas9 | CEN *, CEN4 * | Juvenile–adult phase transition | [35] |
| Citrus sinensis | qRT-PCR | FT | Vegetative–reproductive phase transition | [39] |
| Citrus unshiu | qRT-PCR | FT | Vegetative–reproductive phase transition | [38] |
| Cocos nucifera | RNA-seq | FT | Juvenile–adult phase transition | [29] |
| Cornus spp. | Over-expression | TFL1 * | Juvenile–adult phase transition | [36] |
| Eucalyptus globulus | qRT-PCR | FT | Vegetative–reproductive phase transition | [14] |
| Fragaria vesca | Phenotyping | KSN | Vegetative–reproductive phase transition | [37] |
| Jatropha curcas | RNAi | FT * | Flowering transition | [27] |
| Juglans regia | RNA-seq | FT | Juvenile–adult phase transition | [25] |
| Liriodendron Chinense | RNA-seq | FT | Juvenile–adult phase transition; Perpetual flowering | [20] |
| Malus × domestica | Antisense expression | FT1 * | Juvenile–adult phase transition | [30] |
| VIGS | TFL1 * | Juvenile–adult phase transition | [33] | |
| qRT-PCR; Over-expression | TFL1-1 *, TFL1-2 * | Vegetative–reproductive phase transition | [41] | |
| Malus × domestica ‘Pinova’ | Over-expression; | FT1 * | Juvenile–adult phase transition | [28] |
| Antisense expression | FT1 * | Juvenile–adult phase transition; Vegetative–reproductive phase transition | [31] | |
| Paeonia suffruticosa | RNA-seq; qRT-PCR | FT | Reblooming | [43] |
| Populus deltoides | RT-PCR; Over-expression | FT2 * | Juvenile–adult phase transition; Vegetative–reproductive phase transition | [26] |
| Populus simonii | Over-expression | FT1*, FT2 * | Juvenile–adult phase transition | [22] |
| Populus trichocarpa | qRT-PCR; RNAi; Over-expression | CEN1 * | Juvenile–adult phase transition | [34] |
| Pyrus communis | RNAi | TFL1-1 *, TFL1-2 * | Juvenile–adult phase transition; Vegetative–reproductive phase transition | [32] |
| Pyrus pyrifolia | qRT-PCR | FT1; TFL1 | Vegetative–reproductive phase transition | [40] |
| Rosa spp. | (q)RT-PCR; Phenotyping | KSN | Vegetative–reproductive phase transition | [37] |
| Rosa hybrida | (q)RT-PCR | FT | Vegetative–reproductive phase transition | [12] |
| Vaccinium corymbosum | Over-expression | FT * | Perpetual flowering | [42] |
| Ziziphus jujuba | (q)RT-PCR | FT | Direct flowering | [11] |
* Gene function has been verified via genetics experiments.
Table 3.
Epigenetic regulation of flowering in woody angiosperms.
| Species | Epigenetic Modification | Modified Site | Developmental Transition | Reference |
|---|---|---|---|---|
| Actinidia chinensis | H3K4me3; H3ac | AcSVP2 | Dormancy release | [140] |
| Rhododendron spp. | DNA methylation; H4ac | — | Floral transition | [130,131,132] |
| Castanea sativa | DNA methylation; Histone modification; H4ac | — | Bud dormancy | [128,129] |
| Citrus (‘Moncada’ mandarin) | H3K4me3 | CcMADS19 locus | Floral induction | [143] |
| Euphorbia esula | H3K4me3; H3K27me3 | DAM1 | Endodormancy release | [139] |
| Malus ×domestica | DNA methylation | — | Dormancy release | [119] |
| DNA methylation | SOC1, AP1, SPLs, etc. | Floral transition | [121] | |
| Histone methylation/acetylation | — | Flower induction | [133] | |
| Paeonia suffruticosa | DNA methylation | PsFT | Flowering | [124] |
| Paeonia suffruticosa ‘Luhehong’ | DNA methylation | — | Dormancy release | [120] |
| Populus tremula × Populus alba | Lysine acetylation | Metabolic enzymes | Dormancy release | [146] |
| Prunus avium | DNA methylation; siRNAs | PavMADS1, PavMADS2 | Bud dormancy and flowering | [125] |
| DNA methylation | — | Endodormancy | [126] | |
| H3K4me3 | PavDAM5 | Bud dormancy | [134] | |
| Prunus dulcis | DNA methylation | — | Dormancy release | [127] |
| Prunus persica | H3K4me3; H3K27me3; H3ac | DAM6 | Dormancy release | [136] |
| H3K27me3 | DAM1/4/5/6 | Bud dormancy release | [138] | |
| siRNAs; H3K27me3; CHH methylation | DAMs | Dormancy | [137] | |
| Endoplasmic reticulum stress; unfolded protein response | — | Endodormancy | [144] | |
| Pyrus pyrifolia ‘Kosui’ | H3K4me3 | PpMADS13-1 locus | Endodormancy phase transition | [135] |
| H3K4me3 | PpEBB | Bud break and flowering | [142] | |
| Salix viminalis | DNA methylation | — | Floral transition | [123] |
Note: Histone modification includes H3K4me3, H3K27me3, H3ac, and H4ac.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sun, L.; Nie, T.; Chen, Y.; Yin, Z. From Floral Induction to Blooming: The Molecular Mysteries of Flowering in Woody Plants. Int. J. Mol. Sci. 2022, 23, 10959. https://doi.org/10.3390/ijms231810959
AMA Style
Sun L, Nie T, Chen Y, Yin Z. From Floral Induction to Blooming: The Molecular Mysteries of Flowering in Woody Plants. International Journal of Molecular Sciences. 2022; 23(18):10959. https://doi.org/10.3390/ijms231810959
Chicago/Turabian StyleSun, Liyong, Tangjie Nie, Yao Chen, and Zengfang Yin. 2022. "From Floral Induction to Blooming: The Molecular Mysteries of Flowering in Woody Plants" International Journal of Molecular Sciences 23, no. 18: 10959. https://doi.org/10.3390/ijms231810959
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.