
Defining Specific Cell States of MPTP-Induced Parkinson’s Disease by Single-Nucleus RNA Sequencing
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
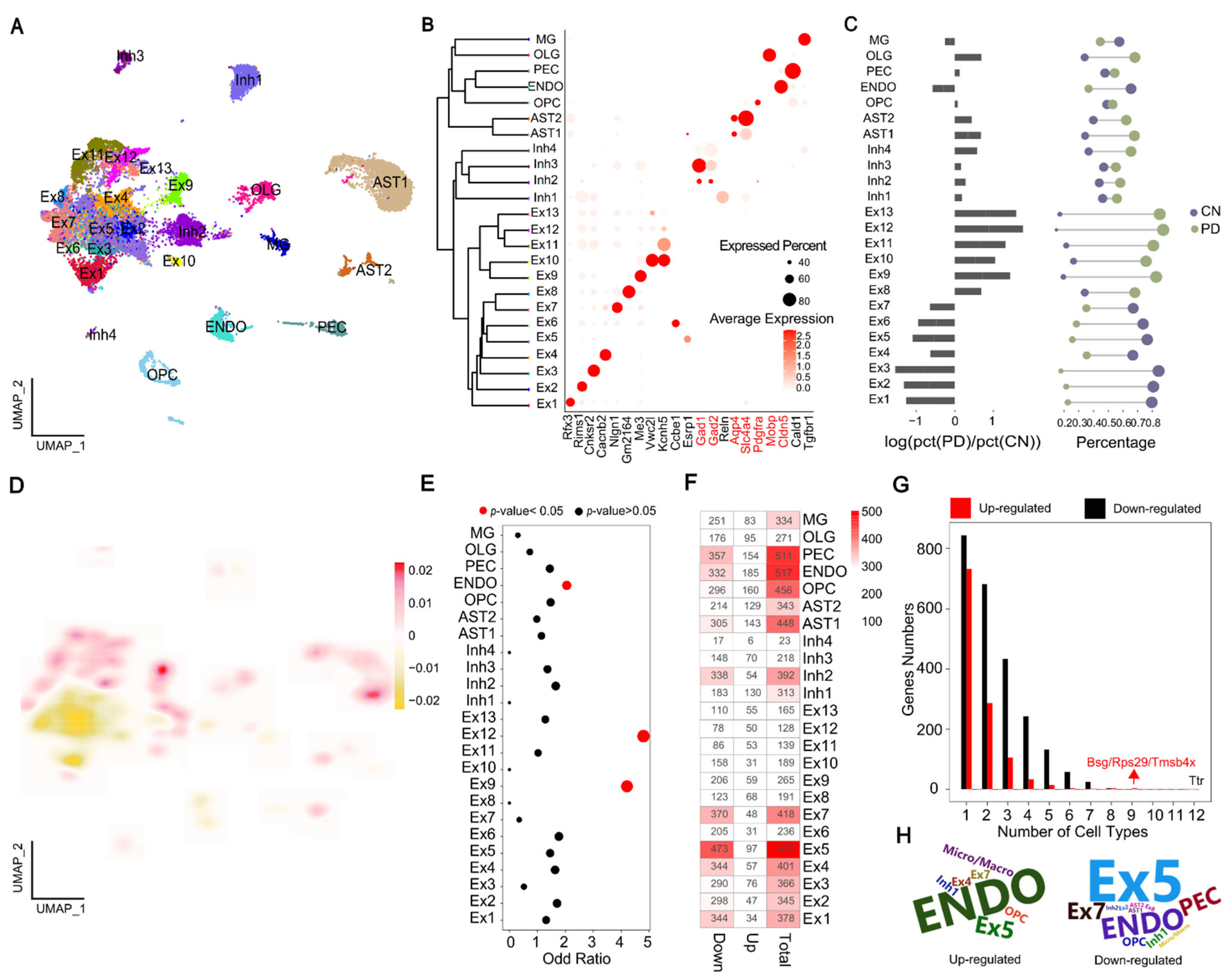
2.1. Single-Nucleus Transcriptome Profiling to Identify Cell Populations
2.2. Multi-Dimensional Validation of MPTP-PD Specific Cells
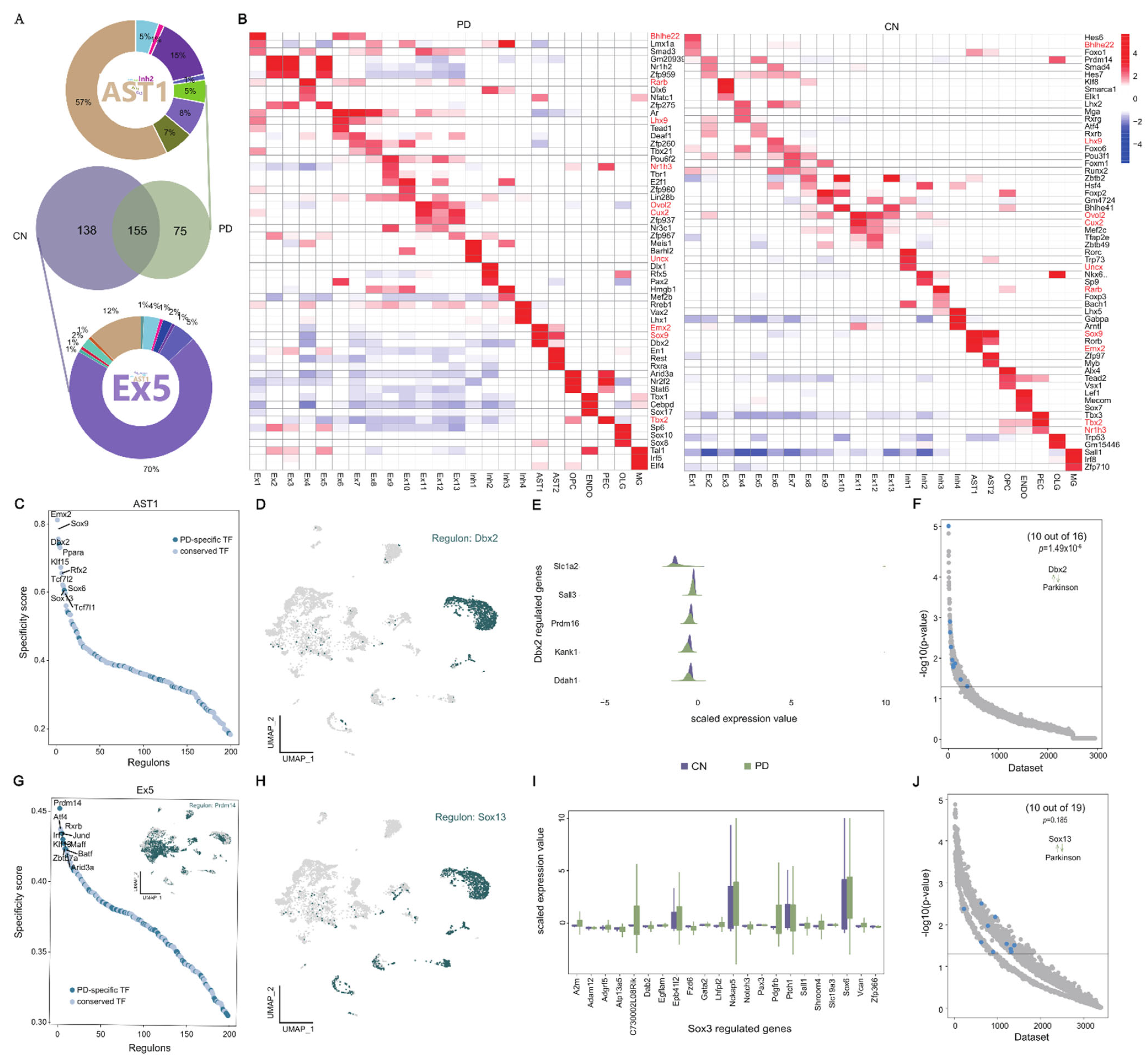
2.3. Transcriptional Regulation of Disease-Specific Astrocytes
2.4. Trajectory Reconstruction of MTPT-PD-Associated Astrocytes and Endothelial Cells
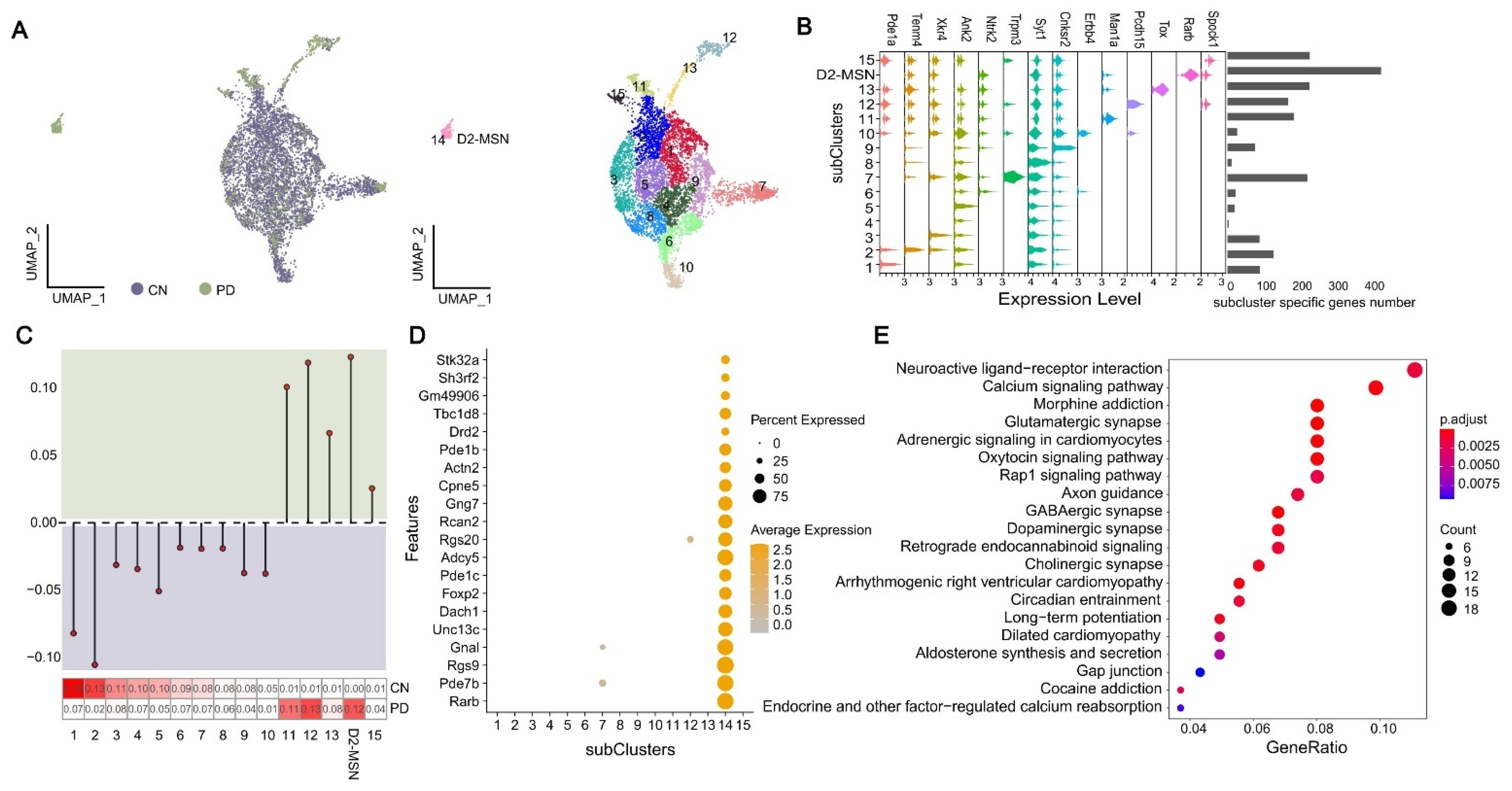
2.5. Characterization of Transcriptomic State of Neuron Cells
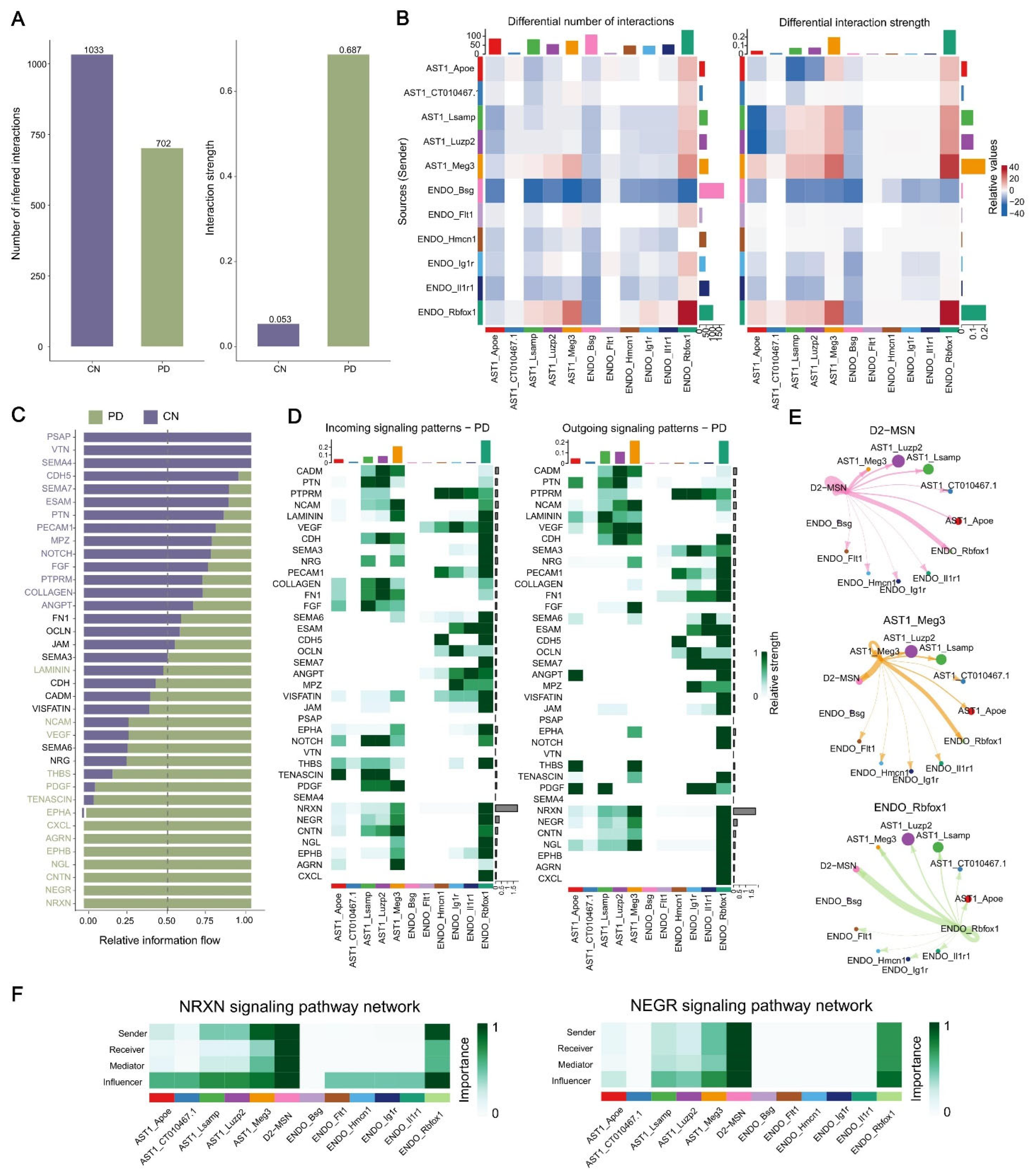
2.6. Analysis of Cell–Cell Communication in MPTP-PD Specific Cells
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Tissue Dissection and Nuclear Extraction
4.3. Library Construction and Sequencing
4.4. Data Demultiplexing and Quality Control
4.5. Clustering and Cell Annotation
4.6. Differential Expressed Genes Analysis
4.7. Inference of Regulon, Quantify Cell-Type Specificity Score and Functional Validation
4.8. Trajectory Analysis Using Monocle3
4.9. Cell–Cell Communication Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tibar, H.; El Bayad, K.; Bouhouche, A.; Ben Haddou, E.; Benomar, A.; Yahyaoui, M.; Benazzouz, A.; Regragui, W. Non-Motor Symptoms of Parkinson’s Disease and Their Impact on Quality of Life in a Cohort of Moroccan Patients. Front. Neurol. 2018, 9, 170. [Google Scholar] [CrossRef] [PubMed]
- Smolders, S.; Broeckhoven, C.V. Genetic perspective on the synergistic connection between vesicular transport, lysosomal and mitochondrial pathways associated with Parkinson’s disease pathogenesis. Acta Neuropathol. Com. 2021, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Mahul-Mellier, A.L.; Burtscher, J.; Maharjan, N.; Weerens, L.; Croisier, M.; Kuttler, F.; Leleu, M.; Knott, G.W.; Lashuel, H.A. The process of Lewy body formation, rather than simply alpha-synuclein fibrillization, is one of the major drivers of neurodegeneration. Proc. Natl. Acad. Sci. USA 2020, 117, 4971–4982. [Google Scholar] [CrossRef] [PubMed]
- Armand, E.J.; Li, J.; Xie, F.; Luo, C.; Mukamel, E.A. Single-Cell Sequencing of Brain Cell Transcriptomes and Epigenomes. Neuron 2021, 109, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Kee, N.; Volakakis, N.; Kirkeby, A.; Dahl, L.; Storvall, H.; Nolbrant, S.; Lahti, L.; Bjorklund, A.K.; Gillberg, L.; Joodmardi, E.; et al. Single-Cell Analysis Reveals a Close Relationship between Differentiating Dopamine and Subthalamic Nucleus Neuronal Lineages. Cell Stem Cell 2017, 20, 29–40. [Google Scholar] [CrossRef]
- La Manno, G.; Gyllborg, D.; Codeluppi, S.; Nishimura, K.; Salto, C.; Zeisel, A.; Borm, L.E.; Stott, S.R.W.; Toledo, E.M.; Villaescusa, J.C.; et al. Molecular Diversity of Midbrain Development in Mouse, Human, and Stem Cells. Cell 2016, 167, 566–580. [Google Scholar] [CrossRef]
- Langston, R.G.; Beilina, A.; Reed, X.; Singleton, A.B.; Blauwendraat, C.; Gibbs, J.R.; Cookson, M.R. Association of a Common Genetic Variant with Parkinson’s Disease is Propagated through Microglia. Sci. Transl. Med. 2022, 12. [Google Scholar]
- Smajić, S.; Prada-Medina, C.A.; Landoulsi, Z.; Dietrich, C.; Jarazo, J.; Henck, J.; Balachandran, S.; Pachchek, S.; Morris, C.M.; Antony, P.; et al. Single-cell sequencing of human midbrain reveals glial activation and a Parkinson-specific neuronal state. Brain 2022, 145, 964–978. [Google Scholar] [CrossRef]
- Zhong, J.; Tang, G.; Zhu, J.; Qiu, X.; Wu, W.; Li, G.; Lin, X.; Liang, L.; Chai, C.; Zeng, Y.; et al. Single-cell brain atlas of Parkinson’s disease mouse model. J. Genet. Genom. 2021, 48, 277–288. [Google Scholar] [CrossRef]
- Picklo, M.J.; Montine, T.J.; Zhu, X. Mitochondrial effects of lipid-derived neurotoxins. J. Alzheimers Dis. 2007, 12, 185–193. [Google Scholar] [CrossRef]
- Freyaldenhoven, T.E.; Ali, S.F. Role of Heat Shock Proteins in MPTP-Induced Neurotoxicity. Ann. N. Y. Acad. Sci. 1997, 825, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, R.; Avila, M.; Gonzalez, J.; El-Bacha, R.S.; Baez, E.; Garcia-Segura, L.M.; Jurado Coronel, J.C.; Capani, F.; Cardona-Gomez, G.P.; Barreto, G.E. Astrocytic modulation of blood brain barrier: Perspectives on Parkinson’s disease. Front. Cell. Neurosci. 2014, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Jia, E.; Pan, M.; Liu, Z.; Zhou, Y.; Zhao, X.; Dong, J.; Bai, Y.; Ge, Q. Transcriptomic profiling of differentially expressed genes and related pathways in different brain regions in Parkinson’s disease. Neurosci. Lett. 2020, 732, 135074. [Google Scholar] [CrossRef] [PubMed]
- Cakar, A.; Durmus-Tekce, H.; Parman, Y. Familial Amyloid Polyneuropathy. Noropsikiyatri Ars. 2019, 56, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Duan, W.; Zhu, X.; Ladenheim, B.; Yu, Q.S.; Guo, Z.; Oyler, J.; Cutler, R.G.; Cadet, J.L.; Greig, N.H.; Mattson, M.P. p53 inhibitors preserve dopamine neurons and motor function in experimental parkinsonism. Ann. Neurol. 2002, 52, 597–606. [Google Scholar] [CrossRef]
- Sun, M.; Nafees, A.; Zhang, R.; Jochen, G. Crybb2 associates with Tmsb4X and is crucial for dendrite morphogenesis. Biochem. Bioph. Res. Commun. 2018, 503, 123–130. [Google Scholar] [CrossRef]
- Morabito, S.; Miyoshi, E.; Michael, N.; Shahin, S.; Martini, A.C.; Head, E.; Silva, J.; Leavy, K.; Perez-Rosendahl, M.; Swarup, V. Single-nucleus chromatin accessibility and transcriptomic characterization of Alzheimer’s disease. Nat. Genet. 2021, 53, 1143–1155. [Google Scholar] [CrossRef]
- Sande, B.; Flerin, C.; Davie, K.; Waegeneer, M.D.; Hulselmans, G.; Aibar, S.; Seurinck, R.; Saelens, W.; Cannoodt, R.; Rouchon, Q. A scalable SCENIC workflow for single-cell gene regulatory network analysis. Nat. Protoc. 2020, 15, 2247–2276. [Google Scholar] [CrossRef]
- Xu, Y.; Cao, B.; Chen, Y.P.; Ou, R.W.; Wei, Q.Q.; Yang, J.; Zhao, B.; Song, W.; Shang, H.F. SLC1A2 rs3794087 are associated with susceptibility to Parkinson’s disease, but not essential tremor, amyotrophic lateral sclerosis or multiple system atrophy in a Chinese population. J. Neurol. Sci. 2016, 365, 96–100. [Google Scholar] [CrossRef]
- Wang, W.; Li, W.; Jiang, W.; Lin, H.; Zhang, D. Genome-wide DNA methylation analysis of cognitive function in middle and old-aged Chinese monozygotic twins. J. Psych. Res. 2021, 136, 571–580. [Google Scholar] [CrossRef]
- Nassir, N.; Bankapur, A.; Samara, B.; Ali, A.; Ahmed, A.; Inuwa, I.; Zarrei, M.; Shabestari, S.; AlBanna, A.; Howe, J.; et al. Single-cell transcriptome identifies molecular subtype of autism spectrum disorder impacted by de novo loss-of-function variants regulating glial cells. Hum. Genom. 2021, 15, 68. [Google Scholar] [CrossRef] [PubMed]
- Whittle, N.; Li, L.; Chen, W.Q.; Yang, J.W.; Sartori, S.B.; Lubec, G.; Singewald, N. Changes in brain protein expression are linked to magnesium restriction-induced depression-like behavior. Amino Acids 2011, 40, 1231–1248. [Google Scholar] [CrossRef]
- Forstner, A.J.; Fischer, S.B.; Schenk, L.M.; Strohmaier, J.; Cichon, S. Whole-exome sequencing of 81 individuals from 27 multiply affected bipolar disorder families. Transl. Psychiat. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Tl, A.; Jia, S.A.; Qz, A.; Sc, A.; Yl, A.; Xin, Z.B.; Zm, A.; Txa, C.; Xg, A. The role of 5-HT 7 R in the memory impairment of mice induced by long-term isoflurane anesthesia. Neurobiol. Learn. Mem. 2022, 188, 107584. [Google Scholar]
- Wang, Y.; Ristevski, S.; Harley, V.R. SOX13 Exhibits a Distinct Spatial and Temporal Expression Pattern During Chondrogenesis, Neurogenesis, and Limb Development. J. Histochem. Cytochem. Offici. J. Histochem. Soc. 2006, 54, 1327. [Google Scholar] [CrossRef] [PubMed]
- Gardini, E.; Giorgi, F.M.; Decherchi, S.; Cavalli, A. Spathial: An R package for the evolutionary analysis of biological data. Bioinformatics 2020, 36, 4664–4667. [Google Scholar] [CrossRef]
- Huang, H.; Zheng, S.; Lu, M. Downregulation of lncRNA MEG3 is involved in Parkinson’s disease. Metab. Brain Dis. 2021, 36, 2323–2328. [Google Scholar] [CrossRef]
- Tan, M.M.; Lawton, M.A.; Jabbari, E.; Reynolds, R.H.; Morris, H.R. Genome-wide association studies of cognitive and motor progression in Parkinson’s disease. Mov. Disord. 2021, 36, 424–433. [Google Scholar] [CrossRef]
- Swaminathan, S.; Kim, S.; Shen, L.; Risacher, S.L.; Foroud, T.; Pankratz, N.; Potkin, S.G.; Huentelman, M.J.; Craig, D.W.; Weiner, M.W. Genomic Copy Number Analysis in Alzheimer’s Disease and Mild Cognitive Impairment: An ADNI Study. Int. J. Alzheimer’s Dis. 2011, 2011, 729478. [Google Scholar] [CrossRef]
- Goes, F.S.; Mcgrath, J.; Avramopoulos, D.; Wolyniec, P.; Pirooznia, M.; Ruczinski, I.; Nestadt, G.; Kenny, E.E.; Vacic, V.; Peters, I. Genome-wide association study of schizophrenia in Ashkenazi Jews. Am. J. Med. Genet. B 2016, 168, 649–659. [Google Scholar] [CrossRef]
- Loo, S.K.; Shtir, C.; Doyle, A.E.; Mick, E.; Mcgough, J.J.; Mccracken, J.; Biederman, J.; Smalley, S.L.; Cantor, R.M.; Faraone, S.V. Genome-Wide Association Study of Intelligence: Additive Effects of Novel Brain Expressed Genes. J. Am. Acad. Child Adolesc. Psychiatry 2012, 51, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Deste Fa No, A.L.; Au, R.; Massaro, J.M.; Beiser, A.S.; Kellyhayes, M.; Kase, C.S.; D ′Agostino, R.B.; Decarli, C.; Atwood, L.D. Genetic correlates of brain aging on MRI and cognitive test measures: A genome-wide association and linkage analysis in the Framingham study. Bmc. Med. Gen. 2007, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, E.; Nogueira, F.C.S.; Velasquez, I.; Schmitt, A.; Falkai, P.; Domont, G.B.; Martins-de-Souza, D. Synaptosomal Proteome of the Orbitofrontal Cortex from Schizophrenia Patients Using Quantitative Label-Free and iTRAQ-Based Shotgun Proteomics. J. Proteome Res. 2017, 16, 4481–4494. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Okerstrom, K.L.; Kingyon, J.R.; Anderson, S.W.; Cavanagh, J.F.; Narayanan, N.S. Startle Habituation and Midfrontal Theta Activity in Parkinson Disease. J. Cogn. Neurosci. 2016, 28, 1923–1932. [Google Scholar] [CrossRef]
- Lee, W.; Koh, S.; Hwang, S.; Kim, S.H. Presynaptic Dysfunction by Familial Factors in Parkinson Disease. Int. Neurourol. J. 2018, 22, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Spencer, B.; Crews, L.; Masliah, E. Climbing the scaffolds of Parkinson’s disease pathogenesis. Neuron 2007, 53, 469–470. [Google Scholar] [CrossRef]
- Cheng, P.; Qiu, Z.; Du, Y. Potassium channels and autism spectrum disorder-an overview. Int. J. Dev. Neurosci. 2021, 81, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Kratschmer, P.; Lowe, S.A.; Buhl, E.; Chen, K.; Kullmann, D.M.; Pittman, A.; Hodge, J.J.L.; Jepson, J.E.C. Impaired Pre-Motor Circuit Activity and Movement in a Drosophila Model of KCNMA1-Linked Dyskinesia. Mov. Disord. 2021, 36, 1158–1169. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Bea, A.; Walker, M.A.; Hyde, T.M.; Kleinman, J.E.; Harrison, P.J.; Lane, T.A. Metabotropic glutamate receptor 3 (mGlu3; mGluR3; GRM3) in schizophrenia: Antibody characterisation and a semi-quantitative western blot study. Schizophr. Res. 2016, 177, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Naruhashi, K.; Kadomatsu, K.; Igakura, T.; Fan, Q.W.; Kuno, N.; Muramatsu, H.; Miyauchi, T.; Hasegawa, T.; Itoh, A.; Muramatsu, T. Abnormalities of sensory and memory functions in mice lacking Bsg gene. Biochem. Bioph. Res. Commun. 1997, 236, 733–737. [Google Scholar] [CrossRef]
- Kasap, M.; Dwyer, D.S. Na+ leak-current channel (NALCN) at the junction of motor and neuropsychiatric symptoms in Parkinson’s disease. J. Neural. Transm. 2021, 128, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Urena-Peralta, J.R.; Alfonso-Loeches, S.; Cuesta-Diaz, C.M.; García-García, F.; Guerri, C. Deep sequencing and miRNA profiles in alcohol-induced neuroinflammation and the TLR4 response in mice cerebral cortex. Sci. Rep. 2018, 8, 15913–15929. [Google Scholar] [CrossRef]
- Cooke, J.P.; Ghebremariam, Y.T. Endothelial Nicotinic Acetylcholine Receptors and Angiogenesis. Trends. Cardiovas. Med. 2008, 18, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Xu, Y.; He, L.; Choi, W.; Simons, M. The Role of Venous Endothelial Cells in Developmental and Pathologic Angiogenesis. Circulation 2021, 144, 1308–1322. [Google Scholar] [CrossRef] [PubMed]
- Laquerriere, A.; Jaber, D.; Abiusi, E.; Maluenda, J.; Melki, J. Phenotypic spectrum and genomics of undiagnosed arthrogryposis multiplex congenita. J. Med. Genet. 2022, 59, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Benton, M.C.; Sutherland, H.G.; Macartney-Coxson, D.; Haupt, L.M.; Griffiths, L.R. Methylome-wide association study of whole blood DNA in the Norfolk Island isolate identifies robust loci associated with age. Aging 2017, 9, 753–766. [Google Scholar] [CrossRef]
- Kellendonk, C.; Simpson, E.H.; Polan, H.J.; Malleret, G.; Vronskaya, S.; Winiger, V.; Moore, H.; Kandel, E.R. Transient and selective overexpression of dopamine D2 receptors in the striatum causes persistent abnormalities in prefrontal cortex functioning. Neuron 2006, 49, 603–615. [Google Scholar] [CrossRef]
- Torri, F.; Akelai, A.; Lupoli, S.; Sironi, M.; Amannzalcenstein, D.; Fumagalli, M.; Fiume, C.D.; Benasher, E.; Kanyas, K.; Cagliani, R. Fine mapping of AHI1 as a schizophrenia susceptibility gene: From association to evolutionary evidence. Faseb. J. 2010, 24, 3066. [Google Scholar] [CrossRef]
- Jose, A.; Morales-Garcia, D.; Aguilar-Morante, E.; Hernandez-Encinas, S.; Alonso-Gil, C. Silencing phosphodiesterase 7B gene by lentiviral-shRNA interference attenuates neurodegeneration and motor deficits in hemiparkinsonian mice. Neurobiol. Aging 2015, 36, 1160–1173. [Google Scholar]
- Psigfogeorgou, K.; Terzi, D.; Papachatzaki, M.M.; Varidaki, A.; Ferguson, D.; Gold, S.J.; Zachariou, V. A Unique Role of RGS9-2 in the Striatum as a Positive or Negative Regulator of Opiate Analgesia. J. Neurosci. 2011, 31, 5617–5624. [Google Scholar] [CrossRef]
- Gerfen, C.; Engber, T.; Mahan, L.; Susel, Z.; Chase, T.; Monsma, F.; Sibley, D. D1 and D2 dopamine receptor-regulated gene expression of striatonigral and striatopallidal neurons. Science 1990, 250, 1429. [Google Scholar] [CrossRef] [PubMed]
- Dobricic, V.; Kresojevic, N.; Westenberger, A.; Svetel, M.; Tomic, A.; Ralic, V.; Petrovic, L.; Lukic, M.J.; Lohmann, K.; Novakovic, L. De novo mutation in the GNAL gene causing seemingly sporadic dystonia in a Serbian patient. Mov. Disord. 2015, 29, 1190–1193. [Google Scholar] [CrossRef] [PubMed]
- Valentina, C.; Andrea, F.; Marco, O.; Dickinson, B.V.; Zhen, L.; Raffaele, I.; Romina, V.; Bulfamante, G.P.; Luca, M.; Gianvito, M. Dynamic and Cell-Specific DACH1 Expression in Human Neocortical and Striatal Development. Cereb. Cortex 2018, 29, 2115–2124. [Google Scholar]
- Niccolini, F.; Mencacci, N.E.; Yousaf, T.; Rabiner, E.A.; Salpietro, V.; Pagano, G.; Balint, B.; Efthymiou, S.; Houlden, H.; Gunn, R.N. PDE10A and ADCY5 mutations linked to molecular and microstructural basal ganglia pathology. Mov. Disord. 2018, 33, 1961–1965. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Ahnn, J. Regulator of Calcineurin (RCAN): Beyond Down Syndrome Critical Region. Mol. Cells 2020, 43, 671–685. [Google Scholar] [PubMed]
- Geertsma, H.M.; Suk, T.R.; Ricke, K.M.; Horsthuis, K.; Parmasad, J.; Fisk, Z.A.; Callaghan, S.M.; Rousseaux, M. Constitutive nuclear accumulation of endogenous alpha-synuclein in mice causes motor impairment and cortical dysfunction, independent of protein aggregation. Hum. Mol. Genet. 2022, 1–16. [Google Scholar] [CrossRef]
- Beker, M.C.; Kilic, E. The role of circadian rhythm in the regulation of cellular protein profiles in the brain. Turk. J. Med. Sci. 2021, 51, 2705–2715. [Google Scholar] [CrossRef]
- Yang, H.; Lin, Q.; Chen, N.; Luo, Z.; Li, H. LncRNA NR_030777 Alleviates Paraquat-Induced Neurotoxicity by Regulating Zfp326 and Cpne5. Toxicol. Sci. 2020, 178, 173–188. [Google Scholar] [CrossRef]
- Kong, Y.; Liang, X.; Liu, L.; Zhang, D.; Wan, C.; Gan, Z.; Yuan, L. High Throughput Sequencing Identifies MicroRNAs Mediating α-Synuclein Toxicity by Targeting Neuroactive-Ligand Receptor Interaction Pathway in Early Stage of Drosophila Parkinson’s Disease Model. PLoS ONE 2015, 10, e0137432. [Google Scholar] [CrossRef]
- Gray, M.T.; Woulfe, J.M. Striatal blood-brain barrier permeability in Parkinson’s disease. J. Cerebr. Blood. F. Met. 2015, 35, 747–750. [Google Scholar] [CrossRef]
- Stelt, M.; Fox, S.H.; Hill, M.; Crossman, A.R.; Brotchie, J.M. A role for endocannabinoids in the generation of parkinsonism and levodopa-induced dyskinesia in MPTP-lesioned non-human primate models of Parkinson’s disease. Faseb J. 2005, 19, 1140. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.C.; Zhao, S.H.; Zeng, W.L.; Yang, Q.T.; Li, C.C. Xenopus Dbx2 is involved in primary neurogenesis and early neural plate patterning. Biochem. Bioph. Res. Commun. 2011, 412, 170–174. [Google Scholar] [CrossRef] [PubMed]
- Lacin, H.; Zhu, Y.; Wilson, B.A.; Skeath, J.B. dbx mediates neuronal specification and differentiation through cross-repressive, lineage-specific interactions with eve and hb9. Development 2009, 136, 3257. [Google Scholar] [CrossRef] [PubMed]
- Lattke, M.; Goldstone, R.; Guillemot, F. Extensive transcriptional and chromatin changes underlie astrocyte maturation in vivo and in culture. Nat. Commun. 2021, 69, 121–122. [Google Scholar] [CrossRef]
- Schlierf, B.; Friedrich, R.P.; Roerig, P.; Felsberg, J.; Reifenberger, G.; Wegner, M. Expression of SoxE and SoxD genes in human gliomas. Neuropath. Appl. Neuro. 2007, 33, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.P.; Tang, D.; Pang, Y.; Li, X.J.; Ma, W. Immunohistochemical basigin expression level in thyroid cancer tissues. J. Surg. Oncol. 2020, 18, 240. [Google Scholar] [CrossRef] [PubMed]
- Wada, K.; Arai, H.; Takanashi, M.; Fukae, J.; Oizumi, H.; Yasuda, T.; Mizuno, Y.; Mochizuki, H. Expression levels of vascular endothelial growth factor and its receptors in Parkinson’s disease. Neuroreport 2006, 17, 705–709. [Google Scholar] [CrossRef]
- Nalls, M.A.; Blauwendraat, C.; Vallerga, C.L.; Heilbron, K.; Castrillo, J. Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: A meta-analysis of genome-wide association studies. Lancet Neurol. 2019, 18, 1091–1102. [Google Scholar] [CrossRef]
- Thobois, S.; Brefel-Courbon, C.; Le Bars, D.; Sgambato-Faure, V. Molecular Imaging of Opioid System in Idiopathic Parkinson’s Disease. Int. Rev. Neurobiol. 2018, 141, 275–303. [Google Scholar]
- Graziano, S.; Marchiò, S.; Bussolino, F.; Arese, M. A peptide from the extracellular region of the synaptic protein α Neurexin stimulates angiogenesis and the vascular specific tyrosine kinase Tie2. Biochem. Biph. Res. Commun. 2013, 432, 574–579. [Google Scholar] [CrossRef]
- Südhof, T.C. Neuroligins and neurexins link synaptic function to cognitive disease. Nature 2008, 455, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Jamain, S.; Radyushkin, K.; Hammerschmidt, K.; Granon, S.; Boretius, S.; Varoqueaux, F.; Ramanantsoa, N.; Gallego, J.; Ronnenberg, A.; Winter, D. Reduced social interaction and ultrasonic communication in a mouse model of monogenic heritable autism. Proc. Nat. Acad. Sci. USA 2008, 105, 1710–1715. [Google Scholar] [CrossRef] [PubMed]
- Cheon, Y.; Yoo, A.; Seo, H.; Yun, S.Y.; Lee, S. Na/K-ATPase beta1-subunit associates with neuronal growth regulator 1 (NEGR1) to participate in intercellular interactions. BMB Rep. 2020, 54, 164–169. [Google Scholar] [CrossRef]
- Stuart, T.; Butler, A.; Hoffman, P.; Hafemeister, C.; Satija, R. Comprehensive Integration of Single-Cell Data. Cell 2019, 177, 1888–1902. [Google Scholar] [CrossRef]
- Zhang, X.; Lan, Y.; Xu, J.; Quan, F.; Zhao, E.; Deng, C.; Luo, T.; Xu, L.; Liao, G.; Yan, M. CellMarker: A manually curated resource of cell markers in human and mouse. Nucleic Acids Res. 2018, 47, 721–728. [Google Scholar] [CrossRef]
- Oscar, F.; Li-Ming, G.; Johan, B. PanglaoDB: A web server for exploration of mouse and human single-cell RNA sequencing data. J. Biolog. Data. 2019, 2019, baz046. [Google Scholar]
- Yu, G.C.; Wang, L.G.; Han, Y.Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. J. Integrat. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Zhu, Q.; Wong, A.K.; Krishnan, A.; Aure, M.R.; Tadych, A.; Zhang, R.; Corney, D.C.; Greene, C.S.; Bongo, L.A.; Kristensen, V.N. Targeted exploration and analysis of large cross-platform human transcriptomic compendia. Nat. Methods 2015, 12, 211–215. [Google Scholar] [CrossRef]
- Jin, S.; Guerrero-Juarez, C.F.; Zhang, L.; Chang, I.; Ramos, R.; Kuan, C.H.; Myung, P.; Plikus, M.V.; Nie, Q. Inference and analysis of cell-cell communication using CellChat. Nat. Commun. 2021, 12, 1088. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Ma, J.; Huang, H.; Xu, J.; Jiang, C.; Ye, K.; Chang, N.; Ge, Q.; Wang, G.; Zhao, X. Defining Specific Cell States of MPTP-Induced Parkinson’s Disease by Single-Nucleus RNA Sequencing. Int. J. Mol. Sci. 2022, 23, 10774. https://doi.org/10.3390/ijms231810774
Guo Y, Ma J, Huang H, Xu J, Jiang C, Ye K, Chang N, Ge Q, Wang G, Zhao X. Defining Specific Cell States of MPTP-Induced Parkinson’s Disease by Single-Nucleus RNA Sequencing. International Journal of Molecular Sciences. 2022; 23(18):10774. https://doi.org/10.3390/ijms231810774
Chicago/Turabian StyleGuo, Yunxia, Junjie Ma, Hao Huang, Jitao Xu, Chao Jiang, Kaiqiang Ye, Ning Chang, Qinyu Ge, Guangzhong Wang, and Xiangwei Zhao. 2022. "Defining Specific Cell States of MPTP-Induced Parkinson’s Disease by Single-Nucleus RNA Sequencing" International Journal of Molecular Sciences 23, no. 18: 10774. https://doi.org/10.3390/ijms231810774