
Functional Characterization of the N-Terminal Disordered Region of the piggyBac Transposase

 , ,
, ,  and
and 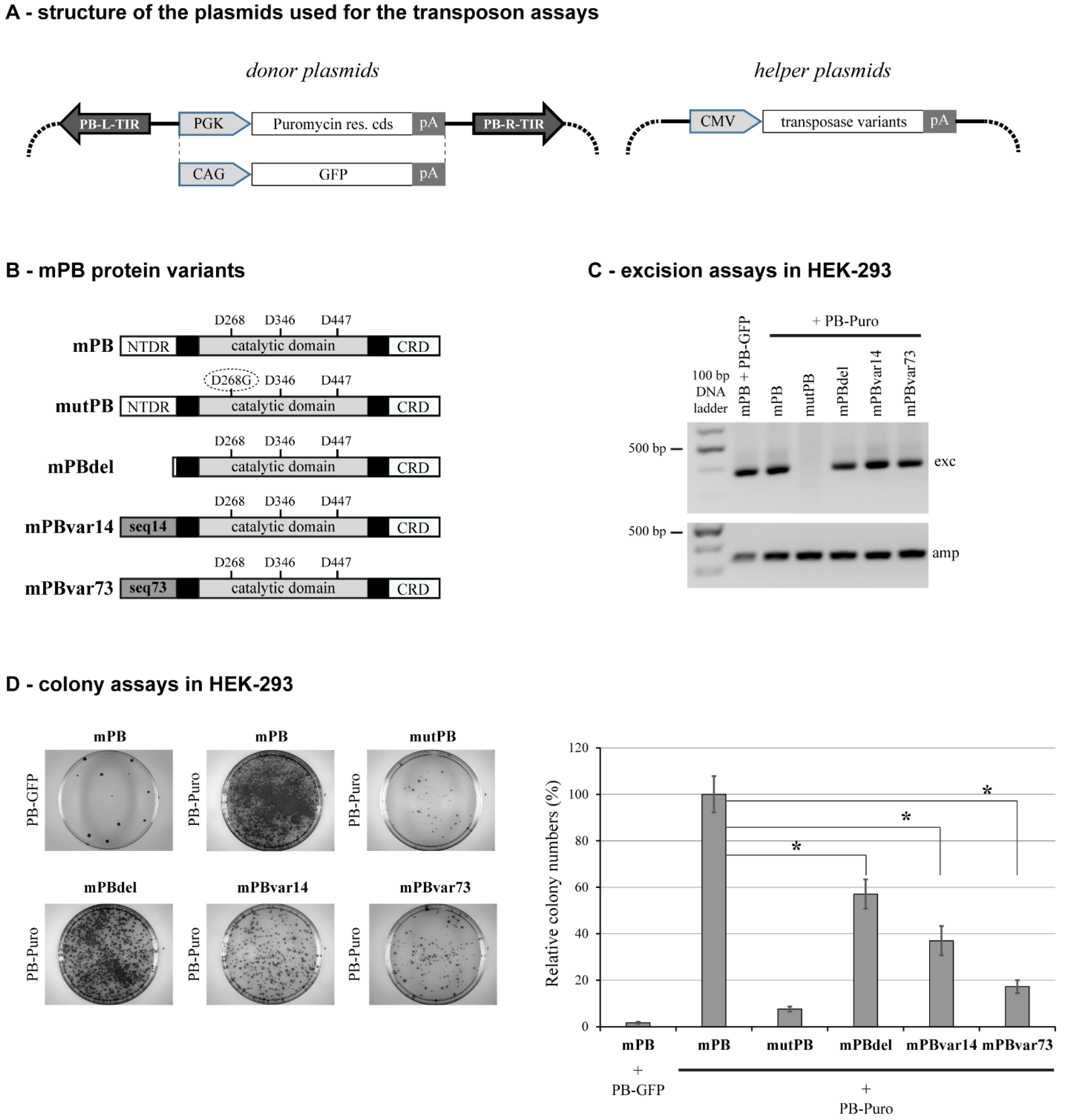{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
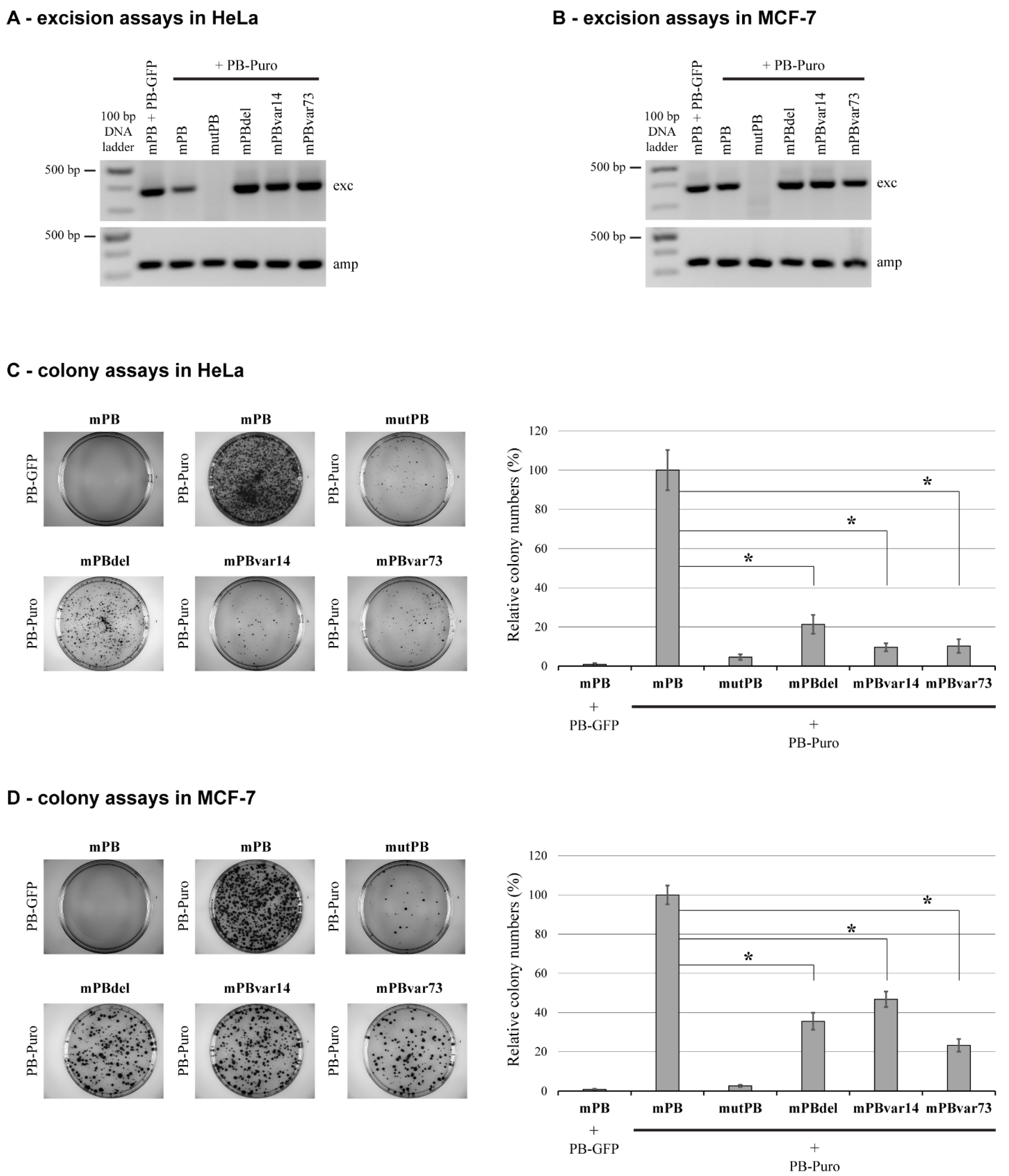
2.1. Deletion of NTDR Significantly Decreases PB Transposition in a Cell Type-Specific Manner
2.2. The Sequence of NTDR Is Important for Its Proper Function
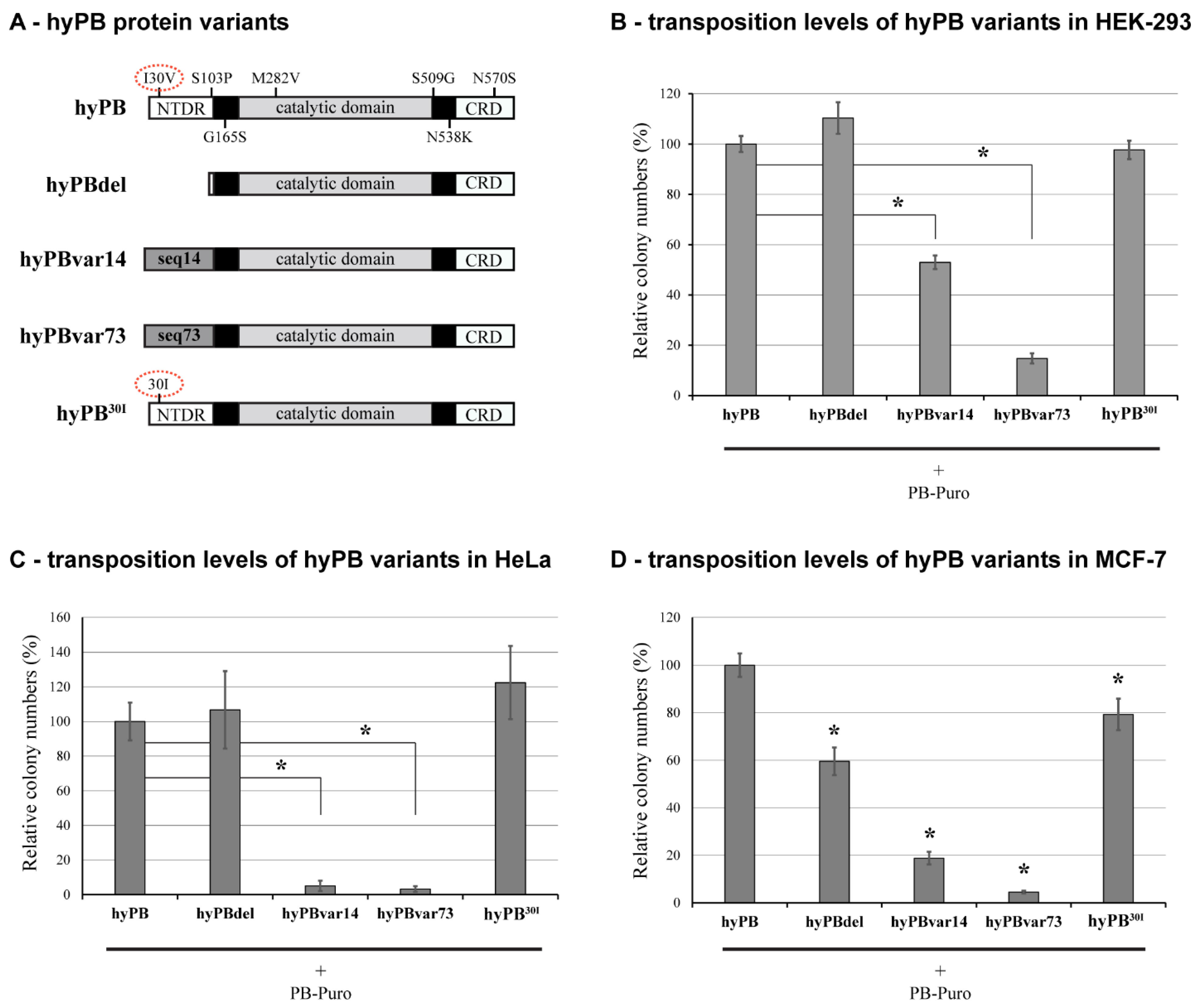
2.3. The Effect of NTDR Deletion on the Hyperactive PB Variant
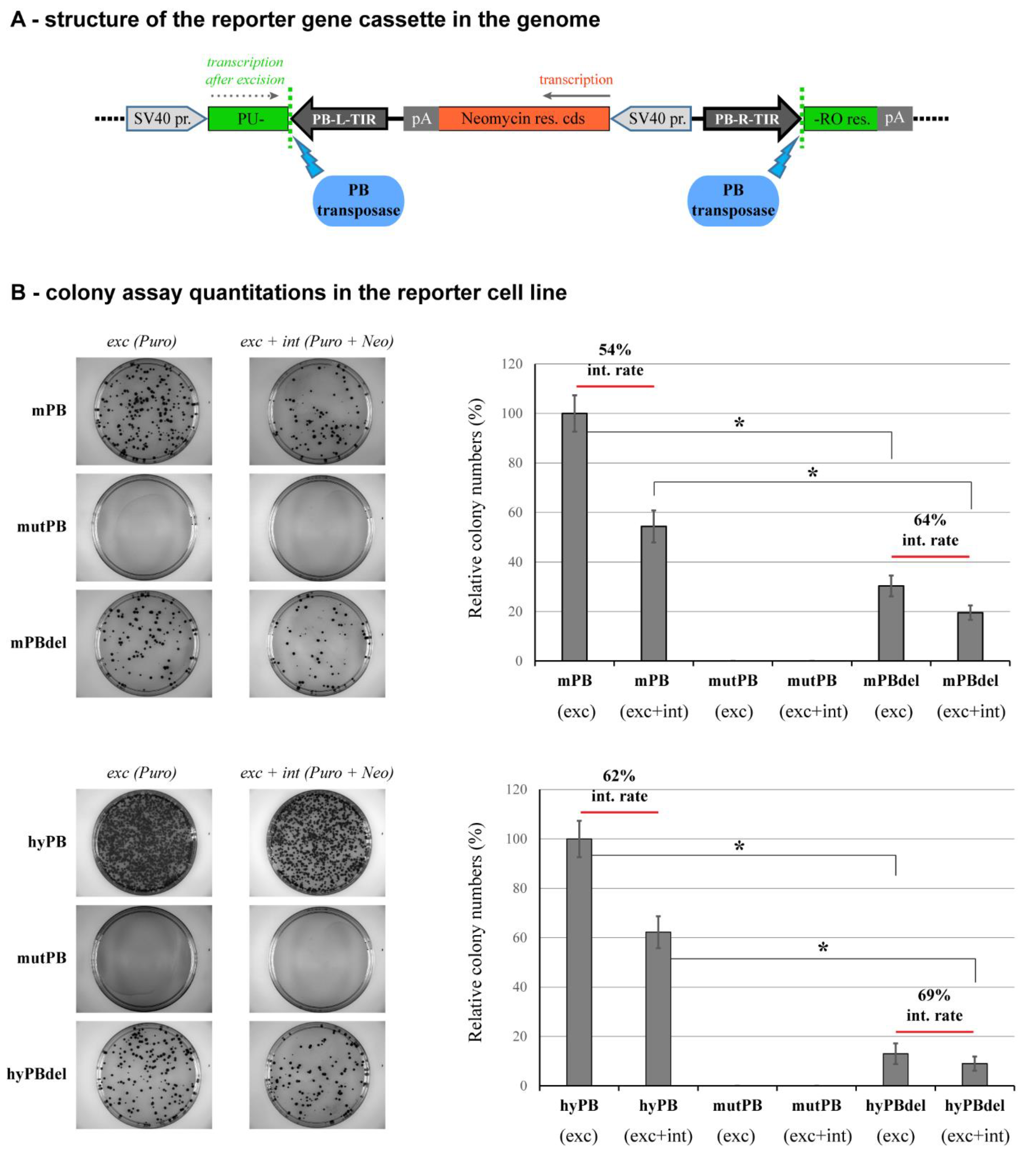
2.4. NTDR Deletion Has a Higher Impact on the Excision Step of Transposition
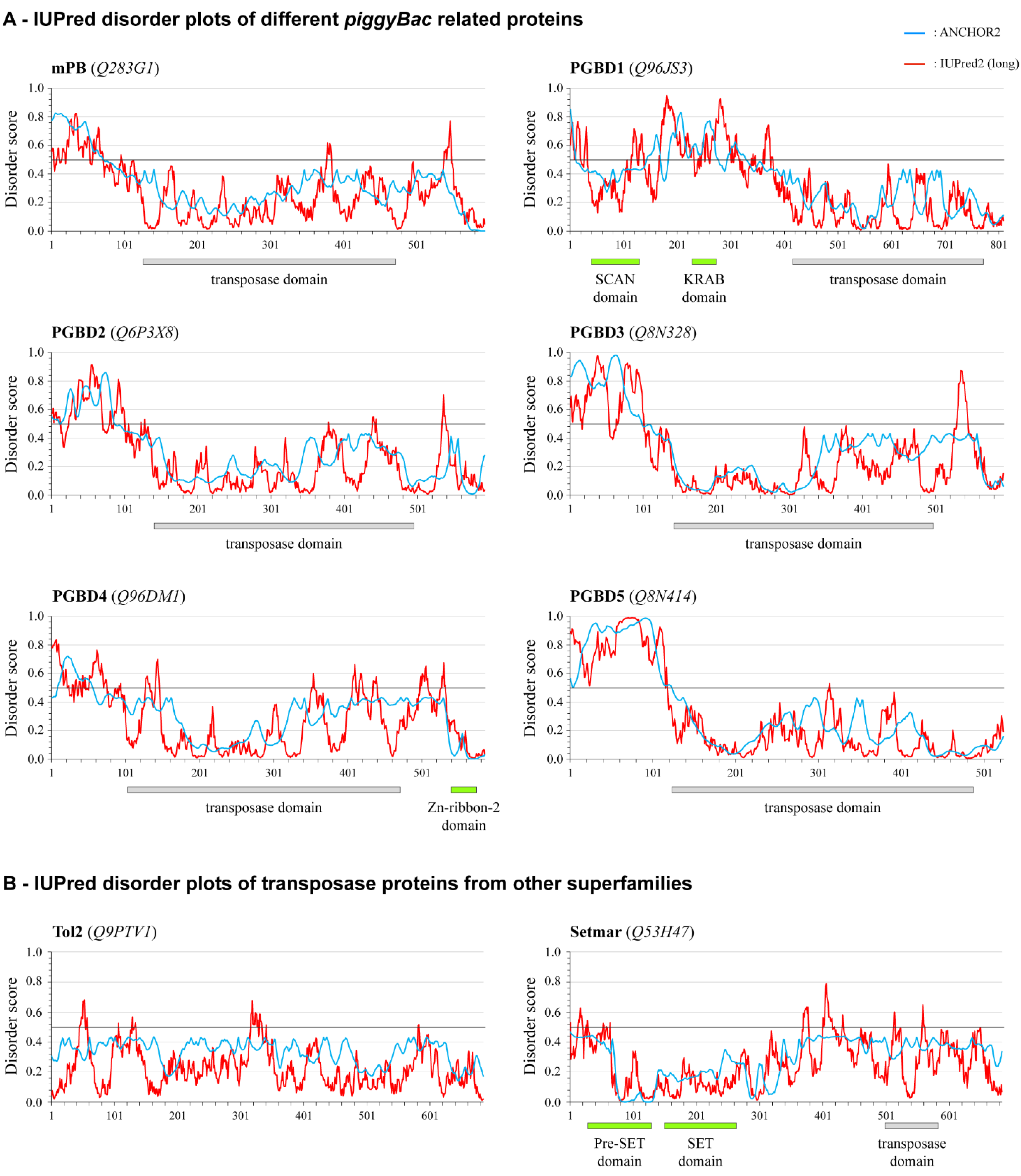
2.5. Conservation of the NTDR in Domesticated PB-Derived Proteins
3. Discussion
4. Materials and Methods
4.1. Plasmid Constructs
4.2. Cell Culturing and Transfection Methods
4.3. Transposition Assays
4.4. Analyzing piggyBac Sequences and Disorder Predictions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Canapa, A.; Barucca, M.; Biscotti, M.A.; Forconi, M.; Olmo, E. Transposons, Genome Size, and Evolutionary Insights in Animals. Cytogenet. Genome Res. 2015, 147, 217–239. [Google Scholar] [CrossRef] [PubMed]
- Wells, J.N.; Feschotte, C. A Field Guide to Eukaryotic Transposable Elements. Annu. Rev. Genet. 2020, 54, 539–561. [Google Scholar] [CrossRef]
- Munoz-Lopez, M.; Garcia-Perez, J.L. DNA transposons: Nature and applications in genomics. Curr. Genom. 2010, 11, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Volff, J.N. Turning junk into gold: Domestication of transposable elements and the creation of new genes in eukaryotes. Bioessays 2006, 28, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Jangam, D.; Feschotte, C.; Betran, E. Transposable Element Domestication as an Adaptation to Evolutionary Conflicts. Trends Genet. 2017, 33, 817–831. [Google Scholar] [CrossRef]
- Burns, K.H. Our Conflict with Transposable Elements and Its Implications for Human Disease. Annu. Rev. Pathol. 2020, 15, 51–70. [Google Scholar] [CrossRef]
- Babarinde, I.A.; Ma, G.; Li, Y.; Deng, B.; Luo, Z.; Liu, H.; Abdul, M.M.; Ward, C.; Chen, M.; Fu, X.; et al. Transposable element sequence fragments incorporated into coding and noncoding transcripts modulate the transcriptome of human pluripotent stem cells. Nucleic Acids Res. 2021, 49, 9132–9153. [Google Scholar] [CrossRef]
- Judd, J.; Sanderson, H.; Feschotte, C. Evolution of mouse circadian enhancers from transposable elements. Genome Biol. 2021, 22, 193. [Google Scholar] [CrossRef]
- Cosby, R.L.; Chang, N.C.; Feschotte, C. Host-transposon interactions: Conflict, cooperation, and cooption. Genes Dev. 2019, 33, 1098–1116. [Google Scholar] [CrossRef]
- Cosby, R.L.; Judd, J.; Zhang, R.; Zhong, A.; Garry, N.; Pritham, E.J.; Feschotte, C. Recurrent evolution of vertebrate transcription factors by transposase capture. Science 2021, 371, eabc6405. [Google Scholar] [CrossRef]
- Rubin, G.M.; Spradling, A.C. Genetic transformation of Drosophila with transposable element vectors. Science 1982, 218, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Wu, X.; Li, G.; Han, M.; Zhuang, Y.; Xu, T. Efficient transposition of the piggyBac (PB) transposon in mammalian cells and mice. Cell 2005, 122, 473–483. [Google Scholar] [CrossRef]
- Cadinanos, J.; Bradley, A. Generation of an inducible and optimized piggyBac transposon system. Nucleic Acids Res. 2007, 35, e87. [Google Scholar] [CrossRef]
- Ivics, Z.; Hackett, P.B.; Plasterk, R.H.; Izsvak, Z. Molecular reconstruction of Sleeping Beauty, a Tc1-like transposon from fish, and its transposition in human cells. Cell 1997, 91, 501–510. [Google Scholar] [CrossRef]
- Kawakami, K.; Shima, A. Identification of the Tol2 transposase of the medaka fish Oryzias latipes that catalyzes excision of a nonautonomous Tol2 element in zebrafish Danio rerio. Gene 1999, 240, 239–244. [Google Scholar] [CrossRef]
- Kawakami, K.; Shima, A.; Kawakami, N. Identification of a functional transposase of the Tol2 element, an Ac-like element from the Japanese medaka fish, and its transposition in the zebrafish germ lineage. Proc. Natl. Acad. Sci. USA 2000, 97, 11403–11408. [Google Scholar] [CrossRef] [PubMed]
- Mates, L.; Chuah, M.K.; Belay, E.; Jerchow, B.; Manoj, N.; Acosta-Sanchez, A.; Grzela, D.P.; Schmitt, A.; Becker, K.; Matrai, J.; et al. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat. Genet. 2009, 41, 753–761. [Google Scholar] [CrossRef]
- Yusa, K.; Zhou, L.; Li, M.A.; Bradley, A.; Craig, N.L. A hyperactive piggyBac transposase for mammalian applications. Proc. Natl. Acad. Sci. USA 2011, 108, 1531–1536. [Google Scholar] [CrossRef]
- Hudecek, M.; Izsvak, Z.; Johnen, S.; Renner, M.; Thumann, G.; Ivics, Z. Going non-viral: The Sleeping Beauty transposon system breaks on through to the clinical side. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 355–380. [Google Scholar] [CrossRef]
- Henssen, A.G.; Henaff, E.; Jiang, E.; Eisenberg, A.R.; Carson, J.R.; Villasante, C.M.; Ray, M.; Still, E.; Burns, M.; Gandara, J.; et al. Genomic DNA transposition induced by human PGBD5. Elife 2015, 4, e10565. [Google Scholar] [CrossRef] [PubMed]
- Beckermann, T.M.; Luo, W.; Wilson, C.M.; Veach, R.A.; Wilson, M.H. Cognate restriction of transposition by piggyBac-like proteins. Nucleic Acids Res. 2021, 49, 8135–8144. [Google Scholar] [CrossRef] [PubMed]
- Kolacsek, O.; Wachtl, G.; Fothi, A.; Schamberger, A.; Sandor, S.; Pergel, E.; Varga, N.; Rasko, T.; Izsvak, Z.; Apati, A.; et al. Functional indications for transposase domestications—Characterization of the human piggyBac transposase derived (PGBD) activities. Gene 2022, 834, 146609. [Google Scholar] [CrossRef] [PubMed]
- Yusa, K. piggyBac Transposon. Microbiol. Spectr. 2015, 3, MDNA3-0028. [Google Scholar] [CrossRef]
- Tipanee, J.; VandenDriessche, T.; Chuah, M.K. Transposons: Moving Forward from Preclinical Studies to Clinical Trials. Hum. Gene Ther. 2017, 28, 1087–1104. [Google Scholar] [CrossRef] [PubMed]
- Woodard, L.E.; Wilson, M.H. piggyBac-ing models and new therapeutic strategies. Trends Biotechnol. 2015, 33, 525–533. [Google Scholar] [CrossRef]
- Kaji, K.; Norrby, K.; Paca, A.; Mileikovsky, M.; Mohseni, P.; Woltjen, K. Virus-free induction of pluripotency and subsequent excision of reprogramming factors. Nature 2009, 458, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Woltjen, K.; Michael, I.P.; Mohseni, P.; Desai, R.; Mileikovsky, M.; Hamalainen, R.; Cowling, R.; Wang, W.; Liu, P.; Gertsenstein, M.; et al. piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. Nature 2009, 458, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Villegas, N.; Nurieva, W.; Amberger, M.; Ivics, Z. Contemporary Transposon Tools: A Review and Guide through Mechanisms and Applications of Sleeping Beauty, piggyBac and Tol2 for Genome Engineering. Int. J. Mol. Sci. 2021, 22, 5084. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Sim, C.; Hong, Y.S.; Hogan, J.R.; Fraser, M.J.; Robertson, H.M.; Collins, F.H. Molecular evolutionary analysis of the widespread piggyBac transposon family and related “domesticated” sequences. Mol. Genet. Genom. 2003, 270, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Pagan, H.J.; Smith, J.D.; Hubley, R.M.; Ray, D.A. PiggyBac-ing on a primate genome: Novel elements, recent activity and horizontal transfer. Genome Biol. Evol. 2010, 2, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Bouallegue, M.; Rouault, J.D.; Hua-Van, A.; Makni, M.; Capy, P. Molecular Evolution of piggyBac Superfamily: From Selfishness to Domestication. Genome Biol. Evol. 2017, 9, 323–339. [Google Scholar] [CrossRef]
- Mitra, R.; Li, X.; Kapusta, A.; Mayhew, D.; Mitra, R.D.; Feschotte, C.; Craig, N.L. Functional characterization of piggyBat from the bat Myotis lucifugus unveils an active mammalian DNA transposon. Proc. Natl. Acad. Sci. USA 2013, 110, 234–239. [Google Scholar] [CrossRef]
- Cary, L.C.; Goebel, M.; Corsaro, B.G.; Wang, H.G.; Rosen, E.; Fraser, M.J. Transposon mutagenesis of baculoviruses: Analysis of Trichoplusia ni transposon IFP2 insertions within the FP-locus of nuclear polyhedrosis viruses. Virology 1989, 172, 156–169. [Google Scholar] [CrossRef]
- Fraser, M.J.; Smith, G.E.; Summers, M.D. Acquisition of Host Cell DNA Sequences by Baculoviruses: Relationship Between Host DNA Insertions and FP Mutants of Autographa californica and Galleria mellonella Nuclear Polyhedrosis Viruses. J. Virol. 1983, 47, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Mitra, R.; Fain-Thornton, J.; Craig, N.L. piggyBac can bypass DNA synthesis during cut and paste transposition. EMBO J. 2008, 27, 1097–1109. [Google Scholar] [CrossRef] [PubMed]
- Keith, J.H.; Schaeper, C.A.; Fraser, T.S.; Fraser, M.J., Jr. Mutational analysis of highly conserved aspartate residues essential to the catalytic core of the piggyBac transposase. BMC Mol. Biol. 2008, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Nesmelova, I.V.; Hackett, P.B. DDE transposases: Structural similarity and diversity. Adv. Drug Deliv. Rev. 2010, 62, 1187–1195. [Google Scholar] [CrossRef]
- Hickman, A.B.; Chandler, M.; Dyda, F. Integrating prokaryotes and eukaryotes: DNA transposases in light of structure. Crit. Rev. Biochem. Mol. Biol. 2010, 45, 50–69. [Google Scholar] [CrossRef]
- Chen, Q.; Luo, W.; Veach, R.A.; Hickman, A.B.; Wilson, M.H.; Dyda, F. Structural basis of seamless excision and specific targeting by piggyBac transposase. Nat. Commun. 2020, 11, 3446. [Google Scholar] [CrossRef]
- Keith, J.H.; Fraser, T.S.; Fraser, M.J., Jr. Analysis of the piggyBac transposase reveals a functional nuclear targeting signal in the 94 c-terminal residues. BMC Mol. Biol. 2008, 9, 72. [Google Scholar] [CrossRef] [Green Version]
- Morellet, N.; Li, X.; Wieninger, S.A.; Taylor, J.L.; Bischerour, J.; Moriau, S.; Lescop, E.; Bardiaux, B.; Mathy, N.; Assrir, N.; et al. Sequence-specific DNA binding activity of the cross-brace zinc finger motif of the piggyBac transposase. Nucleic Acids Res. 2018, 46, 2660–2677. [Google Scholar] [CrossRef] [PubMed]
- Helou, L.; Beauclair, L.; Dardente, H.; Arensburger, P.; Buisine, N.; Jaszczyszyn, Y.; Guillou, F.; Lecomte, T.; Kentsis, A.; Bigot, Y. The C-terminal Domain of piggyBac Transposase Is Not Required for DNA Transposition. J. Mol. Biol. 2021, 433, 166805. [Google Scholar] [CrossRef] [PubMed]
- Guerineau, M.; Bessa, L.; Moriau, S.; Lescop, E.; Bontems, F.; Mathy, N.; Guittet, E.; Bischerour, J.; Betermier, M.; Morellet, N. The unusual structure of the PiggyMac cysteine-rich domain reveals zinc finger diversity in PiggyBac-related transposases. Mob. DNA 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Tompa, P.; Schad, E.; Tantos, A.; Kalmar, L. Intrinsically disordered proteins: Emerging interaction specialists. Curr. Opin. Struct. Biol. 2015, 35, 49–59. [Google Scholar] [CrossRef]
- Meszaros, B.; Erdos, G.; Dosztanyi, Z. IUPred2A: Context-dependent prediction of protein disorder as a function of redox state and protein binding. Nucleic Acids Res. 2018, 46, W329–W337. [Google Scholar] [CrossRef]
- Erdos, G.; Dosztanyi, Z. Analyzing Protein Disorder with IUPred2A. Curr. Protoc. Bioinf. 2020, 70, e99. [Google Scholar] [CrossRef] [PubMed]
- Raskó, T.; Pande, A.; Radscheit, K.; Zink, A.; Singh, M.; Sommer, C.; Wachtl, G.; Kolacsek, O.; Inak, G.; Szvetnik, A.; et al. A novel gene controls a new structure: PiggyBac Transposable Element-derived 1, unique to mammals, controls mammal-specific neuronal paraspeckles. Mol Biol Evol. 2022, msac175. [Google Scholar] [CrossRef]
- Meszaros, B.; Simon, I.; Dosztanyi, Z. Prediction of protein binding regions in disordered proteins. PLoS Comput. Biol. 2009, 5, e1000376. [Google Scholar] [CrossRef] [PubMed]
- Kolacsek, O.; Erdei, Z.; Apati, A.; Sandor, S.; Izsvak, Z.; Ivics, Z.; Sarkadi, B.; Orban, T.I. Excision efficiency is not strongly coupled to transgenic rate: Cell type-dependent transposition efficiency of sleeping beauty and piggyBac DNA transposons. Hum. Gene Ther. Methods 2014, 25, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Burnight, E.R.; Cooney, A.L.; Malani, N.; Brady, T.; Sander, J.D.; Staber, J.; Wheelan, S.J.; Joung, J.K.; McCray, P.B., Jr.; et al. piggyBac transposase tools for genome engineering. Proc. Natl. Acad. Sci. USA 2013, 110, E2279–E2287. [Google Scholar] [CrossRef] [Green Version]
- Li, M.A.; Pettitt, S.J.; Eckert, S.; Ning, Z.; Rice, S.; Cadinanos, J.; Yusa, K.; Conte, N.; Bradley, A. The piggyBac transposon displays local and distant reintegration preferences and can cause mutations at noncanonical integration sites. Mol. Cell Biol. 2013, 33, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Walisko, O.; Izsvak, Z.; Szabo, K.; Kaufman, C.D.; Herold, S.; Ivics, Z. Sleeping Beauty transposase modulates cell-cycle progression through interaction with Miz-1. Proc. Natl. Acad. Sci. USA 2006, 103, 4062–4067. [Google Scholar] [CrossRef] [PubMed]
- Sigalov, A.B.; Zhuravleva, A.V.; Orekhov, V.Y. Binding of intrinsically disordered proteins is not necessarily accompanied by a structural transition to a folded form. Biochimie 2007, 89, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Goyal, S.; Gupta, G.; Qin, H.; Upadya, M.H.; Tan, Y.J.; Chow, V.T.; Song, J. VAPC, an human endogenous inhibitor for hepatitis C virus (HCV) infection, is intrinsically unstructured but forms a “fuzzy complex” with HCV NS5B. PLoS ONE 2012, 7, e40341. [Google Scholar] [CrossRef] [PubMed]
- Tantos, A.; Szabo, B.; Lang, A.; Varga, Z.; Tsylonok, M.; Bokor, M.; Verebelyi, T.; Kamasa, P.; Tompa, K.; Perczel, A.; et al. Multiple fuzzy interactions in the moonlighting function of thymosin-beta4. Intrinsically Disord. Proteins 2013, 1, e26204. [Google Scholar] [CrossRef] [PubMed]
- Haarmann, C.S.; Green, D.; Casarotto, M.G.; Laver, D.R.; Dulhunty, A.F. The random-coil ‘C’ fragment of the dihydropyridine receptor II-III loop can activate or inhibit native skeletal ryanodine receptors. Biochem. J. 2003, 372, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Schad, E.; Kalmar, L.; Tompa, P. Exon-phase symmetry and intrinsic structural disorder promote modular evolution in the human genome. Nucleic Acids Res. 2013, 41, 4409–4422. [Google Scholar] [CrossRef] [PubMed]
- Miskey, C.; Izsvak, Z.; Plasterk, R.H.; Ivics, Z. The Frog Prince: A reconstructed transposon from Rana pipiens with high transpositional activity in vertebrate cells. Nucleic Acids Res. 2003, 31, 6873–6881. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wachtl, G.; Schád, É.; Huszár, K.; Palazzo, A.; Ivics, Z.; Tantos, Á.; Orbán, T.I. Functional Characterization of the N-Terminal Disordered Region of the piggyBac Transposase. Int. J. Mol. Sci. 2022, 23, 10317. https://doi.org/10.3390/ijms231810317
Wachtl G, Schád É, Huszár K, Palazzo A, Ivics Z, Tantos Á, Orbán TI. Functional Characterization of the N-Terminal Disordered Region of the piggyBac Transposase. International Journal of Molecular Sciences. 2022; 23(18):10317. https://doi.org/10.3390/ijms231810317
Chicago/Turabian StyleWachtl, Gerda, Éva Schád, Krisztina Huszár, Antonio Palazzo, Zoltán Ivics, Ágnes Tantos, and Tamás I. Orbán. 2022. "Functional Characterization of the N-Terminal Disordered Region of the piggyBac Transposase" International Journal of Molecular Sciences 23, no. 18: 10317. https://doi.org/10.3390/ijms231810317