
Microglial CD74 Expression Is Regulated by TGFβ Signaling

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
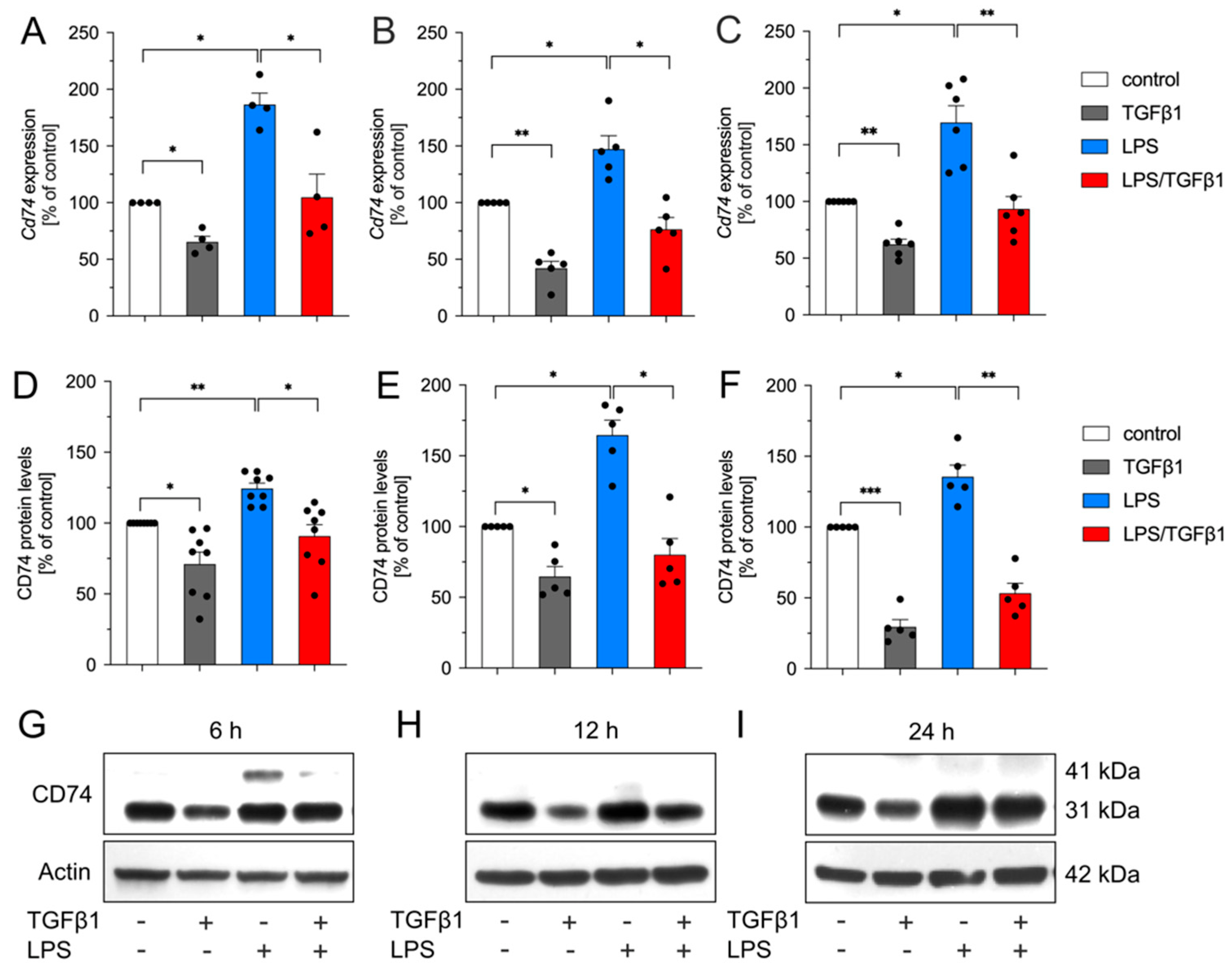
2.1. TGFβ1 Inhibits LPS-Mediated Upregulation of Cd74 in BV2 Cells
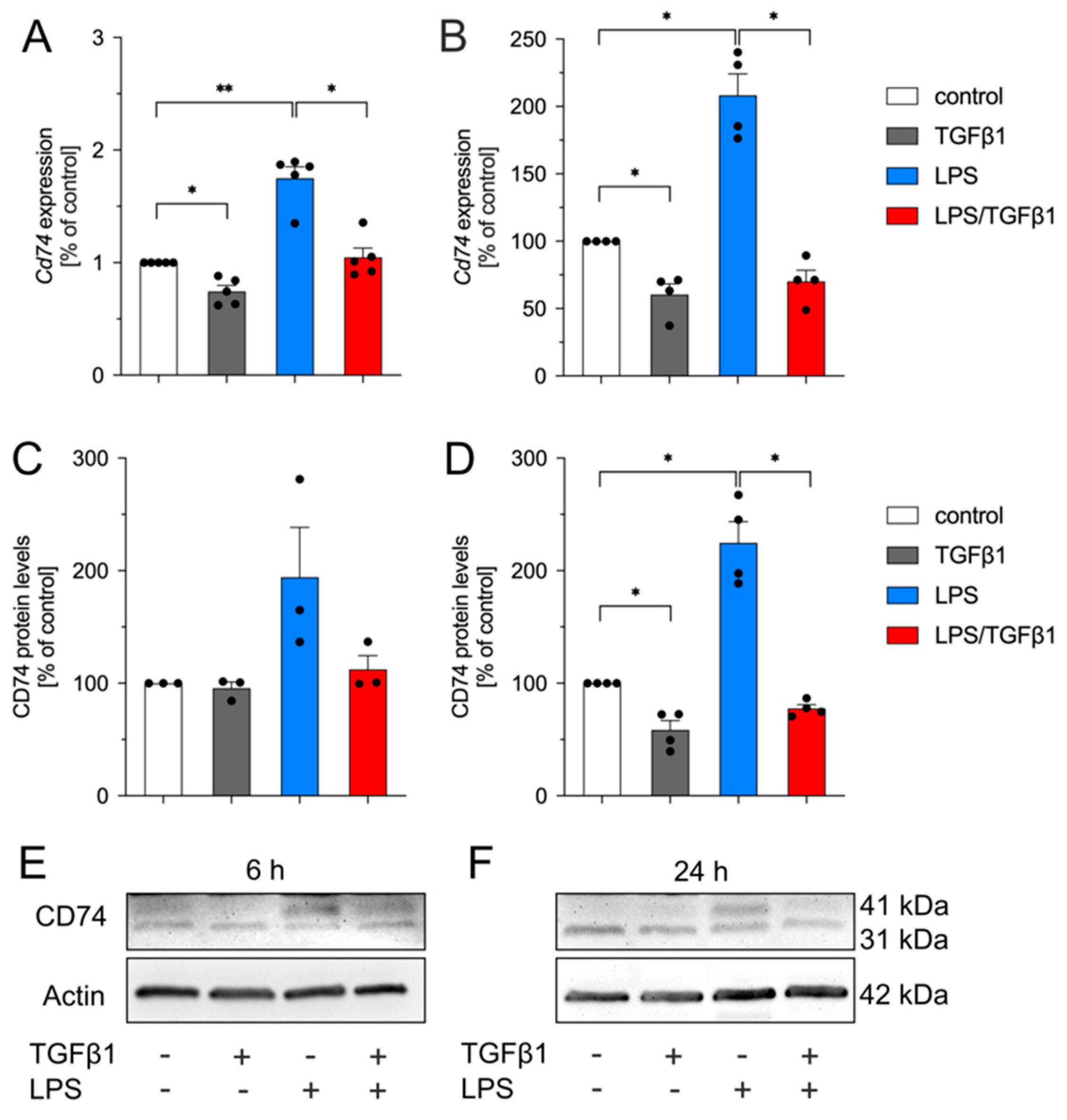
2.2. TGFβ1 Inhibits LPS-Mediated Upregulation of Cd74 in Primary Mouse Microglia
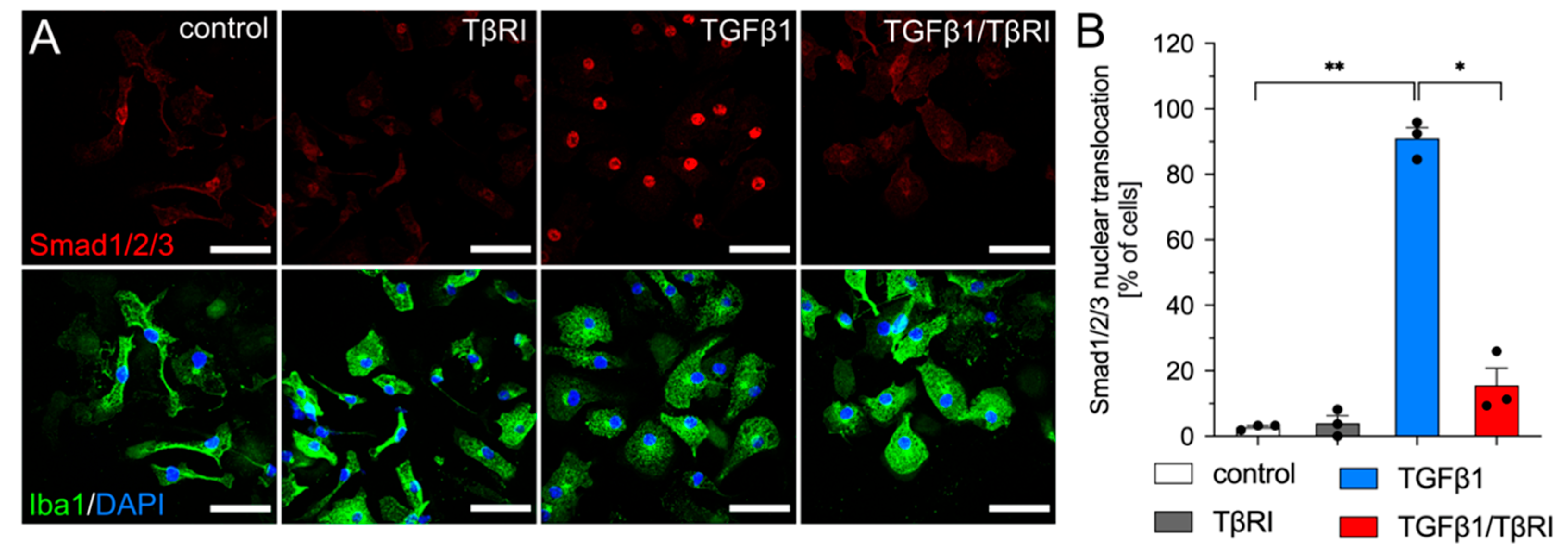
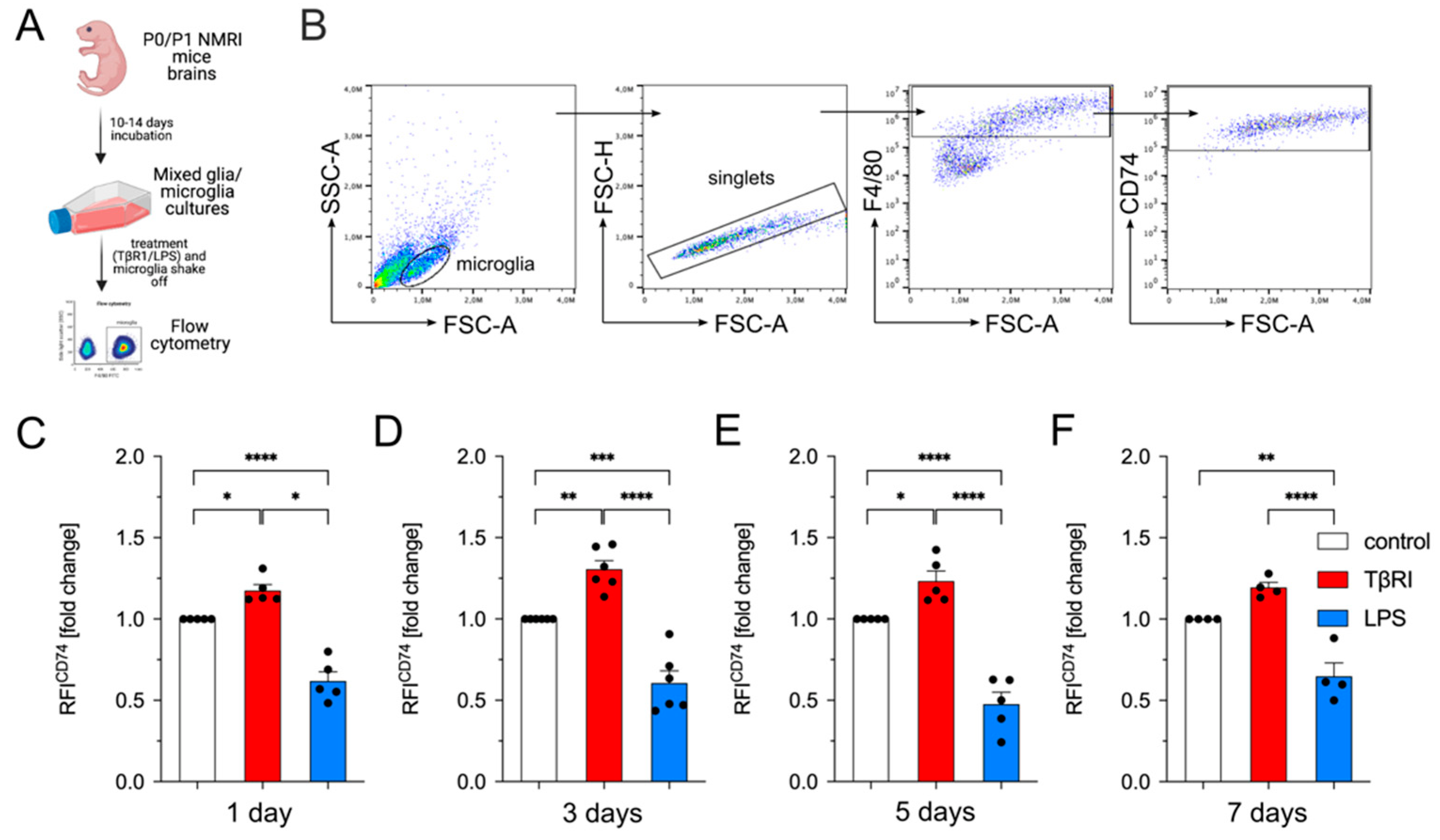
2.3. Inhibition of TGFβ Signaling Increases CD74 Cell Surface Levels in Primary Mouse Microglia
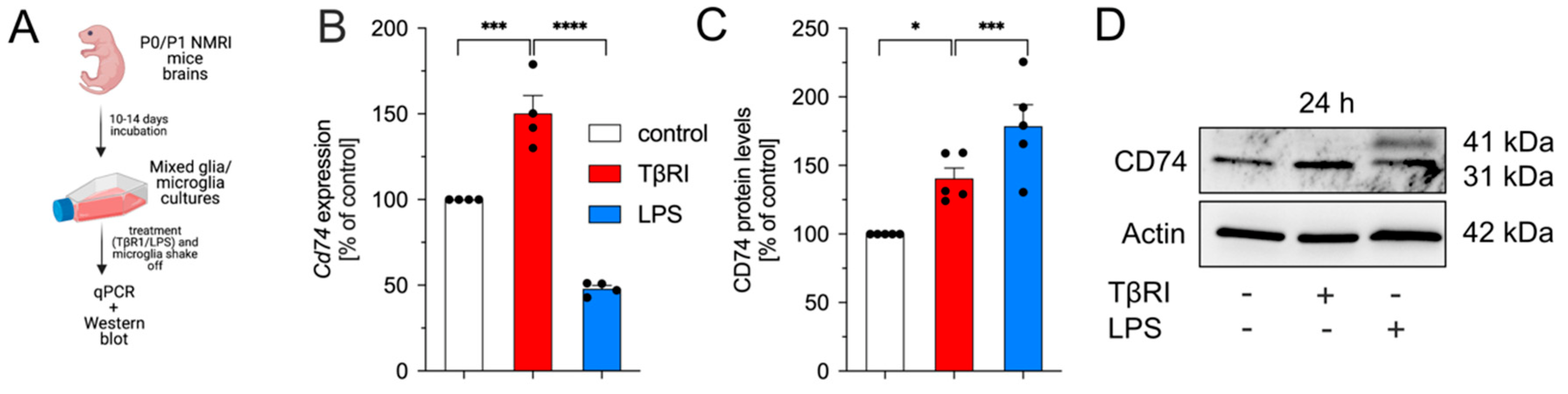
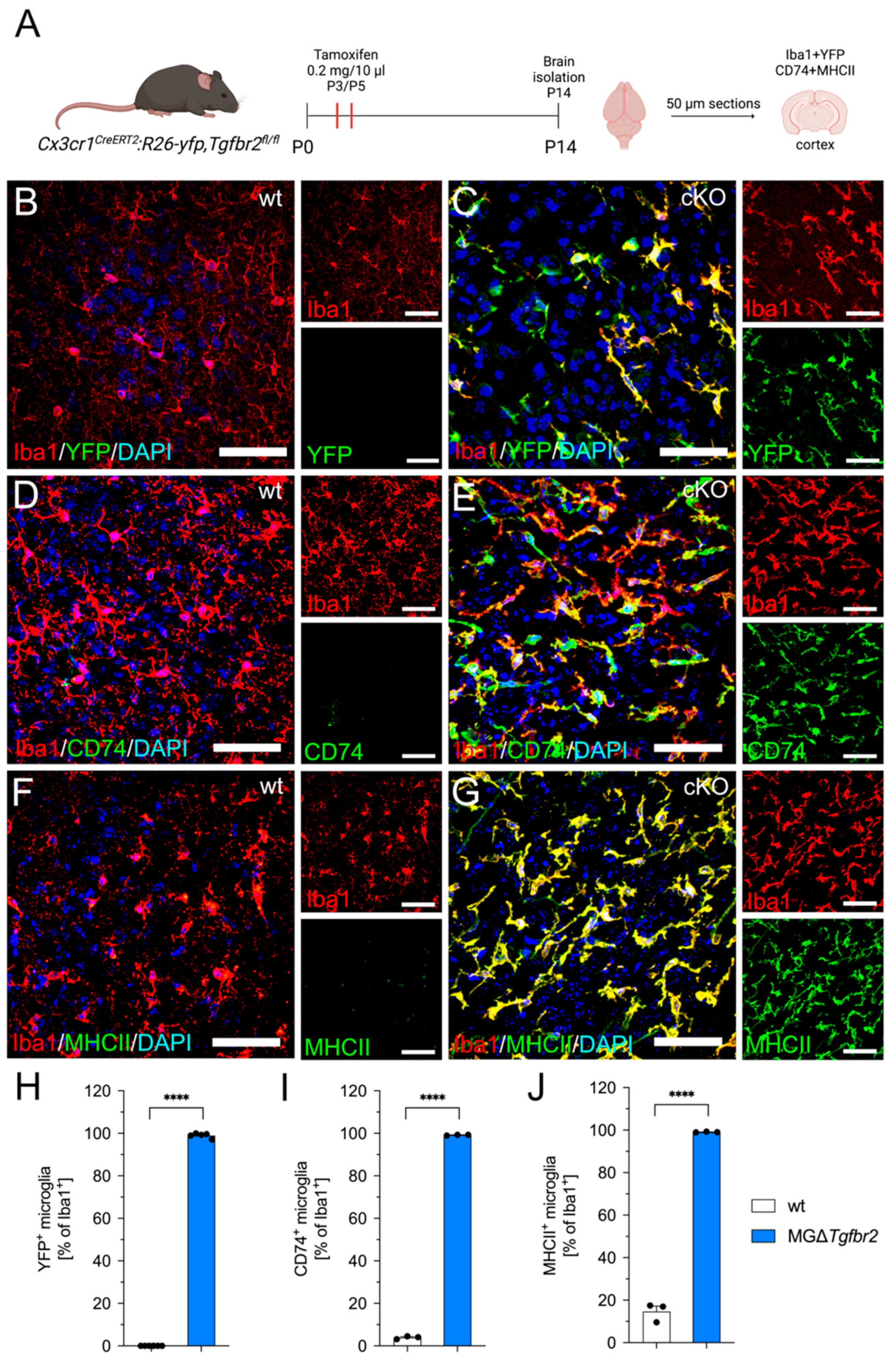
2.4. Microglia-Specific Knockout of Tgfbr2 Increases Microglial CD74 and MHCII Expression In Vivo
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents
4.3. BV2 Cell Culture
4.4. Primary Microglia Cultures
4.5. RNA Isolation, Reverse Transcription, and Quantitative RT-PCR
4.6. Protein Isolation and Western Blotting
4.7. Immunocytochemistry and Immunohistochemistry
4.8. Flow Cytometry
4.9. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prinz, M.; Jung, S.; Priller, J. Microglia Biology: One Century of Evolving Concepts. Cell 2019, 179, 292–311. [Google Scholar] [CrossRef] [PubMed]
- Prinz, M.; Priller, J. Microglia and Brain Macrophages in the Molecular Age: From Origin to Neuropsychiatric Disease. Nat. Rev. Neurosci. 2014, 15, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Nimmerjahn, A.; Kirchhoff, F.; Helmchen, F. Resting Microglial Cells Are Highly Dynamic Surveillants of Brain Parenchyma in Vivo. Science 2005, 308, 1314–1318. [Google Scholar] [CrossRef] [PubMed]
- Kigerl, K.A.; de Rivero Vaccari, J.P.; Dietrich, W.D.; Popovich, P.G.; Keane, R.W. Pattern Recognition Receptors and Central Nervous System Repair. Exp. Neurol. 2014, 258, 5–16. [Google Scholar] [CrossRef]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E.; et al. Identification of a Unique TGF-β-Dependent Molecular and Functional Signature in Microglia. Nat. Neurosci. 2014, 17, 131–143. [Google Scholar] [CrossRef]
- Hickman, S.E.; Kingery, N.D.; Ohsumi, T.K.; Borowsky, M.L.; Wang, L.-C.; Means, T.K.; El Khoury, J. The Microglial Sensome Revealed by Direct RNA Sequencing. Nat. Neurosci. 2013, 16, 1896–1905. [Google Scholar] [CrossRef]
- Keren-Shaul, H.; Spinrad, A.; Weiner, A.; Matcovitch-Natan, O.; Dvir-Szternfeld, R.; Ulland, T.K.; David, E.; Baruch, K.; Lara-Astaiso, D.; Toth, B.; et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer’s Disease. Cell 2017, 169, 1276–1290.e17. [Google Scholar] [CrossRef]
- Butovsky, O.; Weiner, H.L. Microglial Signatures and Their Role in Health and Disease. Nat. Rev. Neurosci. 2018, 19, 622–635. [Google Scholar] [CrossRef]
- Zöller, T.; Attaai, A.; Potru, P.S.; Ruß, T.; Spittau, B. Aged Mouse Cortical Microglia Display an Activation Profile Suggesting Immunotolerogenic Functions. Int. J. Mol. Sci. 2018, 19, 706. [Google Scholar] [CrossRef]
- Jin, C.; Shao, Y.; Zhang, X.; Xiang, J.; Zhang, R.; Sun, Z.; Mei, S.; Zhou, J.; Zhang, J.; Shi, L. A Unique Type of Highly-Activated Microglia Evoking Brain Inflammation via Mif/Cd74 Signaling Axis in Aged Mice. Aging Dis. 2021, 12, 2125–2139. [Google Scholar] [CrossRef]
- Attaai, A.; Neidert, N.; von Ehr, A.; Potru, P.S.; Zöller, T.; Spittau, B. Postnatal Maturation of Microglia Is Associated with Alternative Activation and Activated TGFβ Signaling. Glia 2018, 66, 1695–1708. [Google Scholar] [CrossRef]
- Arnold, T.D.; Lizama, C.O.; Cautivo, K.M.; Santander, N.; Lin, L.; Qiu, H.; Huang, E.J.; Liu, C.; Mukouyama, Y.-S.; Reichardt, L.F.; et al. Impaired AVβ8 and TGFβ Signaling Lead to Microglial Dysmaturation and Neuromotor Dysfunction. J. Exp. Med. 2019, 216, 900–915. [Google Scholar] [CrossRef]
- Kim, W.-K.; Hwang, S.-Y.; Oh, E.-S.; Piao, H.Z.; Kim, K.-W.; Han, I.-O. TGF-Beta1 Represses Activation and Resultant Death of Microglia via Inhibition of Phosphatidylinositol 3-Kinase Activity. J. Immunol. 2004, 172, 7015–7023. [Google Scholar] [CrossRef]
- Zhou, X.; Zöller, T.; Krieglstein, K.; Spittau, B. TGFβ1 Inhibits IFNγ-Mediated Microglia Activation and Protects MDA Neurons from IFNγ-Driven Neurotoxicity. J. Neurochem. 2015, 134, 125–134. [Google Scholar] [CrossRef]
- Zöller, T.; Schneider, A.; Kleimeyer, C.; Masuda, T.; Potru, P.S.; Pfeifer, D.; Blank, T.; Prinz, M.; Spittau, B. Silencing of TGFβ Signalling in Microglia Results in Impaired Homeostasis. Nat. Commun. 2018, 9, 4011. [Google Scholar] [CrossRef]
- Qin, Y.; Garrison, B.S.; Ma, W.; Wang, R.; Jiang, A.; Li, J.; Mistry, M.; Bronson, R.T.; Santoro, D.; Franco, C.; et al. A Milieu Molecule for TGF-β Required for Microglia Function in the Nervous System. Cell 2018, 174, 156–171.e16. [Google Scholar] [CrossRef]
- Schröder, B. The Multifaceted Roles of the Invariant Chain CD74—More than Just a Chaperone. Biochim. Biophys. Acta 2016, 1863, 1269–1281. [Google Scholar] [CrossRef]
- Marsh, L.M.; Cakarova, L.; Kwapiszewska, G.; von Wulffen, W.; Herold, S.; Seeger, W.; Lohmeyer, J. Surface Expression of CD74 by Type II Alveolar Epithelial Cells: A Potential Mechanism for Macrophage Migration Inhibitory Factor-Induced Epithelial Repair. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 296, L442–L452. [Google Scholar] [CrossRef]
- Fan, H.; Hall, P.; Santos, L.L.; Gregory, J.L.; Fingerle-Rowson, G.; Bucala, R.; Morand, E.F.; Hickey, M.J. Macrophage Migration Inhibitory Factor and CD74 Regulate Macrophage Chemotactic Responses via MAPK and Rho GTPase. J. Immunol. 2011, 186, 4915–4924. [Google Scholar] [CrossRef]
- Leng, L.; Metz, C.N.; Fang, Y.; Xu, J.; Donnelly, S.; Baugh, J.; Delohery, T.; Chen, Y.; Mitchell, R.A.; Bucala, R. MIF Signal Transduction Initiated by Binding to CD74. J. Exp. Med. 2003, 197, 1467–1476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryan, K.J.; Zhu, X.; Harris, P.L.; Perry, G.; Castellani, R.J.; Smith, M.A.; Casadesus, G. Expression of CD74 Is Increased in Neurofibrillary Tangles in Alzheimer’s Disease. Mol. Neurodegener. 2008, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Ghoochani, A.; Schwarz, M.A.; Yakubov, E.; Engelhorn, T.; Doerfler, A.; Buchfelder, M.; Bucala, R.; Savaskan, N.E.; Eyüpoglu, I.Y. MIF-CD74 Signaling Impedes Microglial M1 Polarization and Facilitates Brain Tumorigenesis. Oncogene 2016, 35, 6246–6261. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.K.; Park, J.H.; Lee, T.-K.; Kim, D.W.; Yoo, K.-Y.; Ahn, J.H.; Kim, Y.H.; Cho, J.H.; Kim, Y.-M.; Won, M.-H.; et al. CD74-Immunoreactive Activated M1 Microglia Are Shown Late in the Gerbil Hippocampal CA1 Region Following Transient Cerebral Ischemia. Mol. Med. Rep. 2017, 15, 4148–4154. [Google Scholar] [CrossRef] [PubMed]
- Shachar, I.; Elliott, E.A.; Chasnoff, B.; Grewal, I.S.; Flavell, R.A. Reconstitution of Invariant Chain Function in Transgenic Mice In Vivo by Individual P31 and P41 Isoforms. Immunity 1995, 3, 373–383. [Google Scholar] [CrossRef]
- Bikoff, E.K.; Kenty, G.; Van Kaer, L. Distinct Peptide Loading Pathways for MHC Class II Molecules Associated with Alternative Ii Chain Isoforms. J. Immunol. 1998, 160, 3101–3110. [Google Scholar]
- Fortin, J.-S.; Cloutier, M.; Thibodeau, J. Exposing the Specific Roles of the Invariant Chain Isoforms in Shaping the MHC Class II Peptidome. Front. Immunol. 2013, 4, 443. [Google Scholar] [CrossRef]
- Al Abdulmonem, W.; Rasheed, Z.; Aljohani, A.S.M.; Omran, O.M.; Rasheed, N.; Alkhamiss, A.; AM Al Salloom, A.; Alhumaydhi, F.; Alblihed, M.A.; Al Ssadh, H.; et al. Absence of CD74 Isoform at 41kDa Prevents the Heterotypic Associations between CD74 and CD44 in Human Lung Adenocarcinoma-Derived Cells. Immunol. Investig. 2021, 50, 891–905. [Google Scholar] [CrossRef]
- Shishido, T.; Kohyama, M.; Nakai, W.; Matsumoto, M.; Miyata, H.; Suenaga, T.; Arase, H. Invariant Chain P41 Mediates Production of Soluble MHC Class II Molecules. Biochem. Biophys. Res. Commun. 2019, 509, 216–221. [Google Scholar] [CrossRef]
- Mihelic, M.; Dobersek, A.; Guncar, G.; Turk, D. Inhibitory Fragment from the P41 Form of Invariant Chain Can Regulate Activity of Cysteine Cathepsins in Antigen Presentation. J. Biol. Chem. 2008, 283, 14453–14460. [Google Scholar] [CrossRef]
- Xu, S.; Zhang, H.; Yang, X.; Qian, Y.; Xiao, Q. Inhibition of Cathepsin L Alleviates the Microglia-Mediated Neuroinflammatory Responses through Caspase-8 and NF-ΚB Pathways. Neurobiol. Aging 2018, 62, 159–167. [Google Scholar] [CrossRef]
- Spittau, B.; Wullkopf, L.; Zhou, X.; Rilka, J.; Pfeifer, D.; Krieglstein, K. Endogenous Transforming Growth Factor-Beta Promotes Quiescence of Primary Microglia in Vitro. Glia 2013, 61, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Gil-Yarom, N.; Radomir, L.; Sever, L.; Kramer, M.P.; Lewinsky, H.; Bornstein, C.; Blecher-Gonen, R.; Barnett-Itzhaki, Z.; Mirkin, V.; Friedlander, G.; et al. CD74 Is a Novel Transcription Regulator. Proc. Natl. Acad. Sci. USA 2017, 114, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Wang, Y.; Zhou, Y.; Zhu, Z.; Zhang, Q.; Zhang, X.; Wang, W.; Gu, X.; Guo, A.; Wang, Y. Macrophage Migration Inhibitory Factor Activates Inflammatory Responses of Astrocytes through Interaction with CD74 Receptor. Oncotarget 2017, 8, 2719–2730. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.-C.; Wu, T.-N.; Chen, Y.-C.; Lu, C.-H.; Wabitsch, M.; Tian, Y.-F.; Hsieh, P.-S. Targetted Inhibition of CD74 Attenuates Adipose COX-2-MIF-Mediated M1 Macrophage Polarization and Retards Obesity-Related Adipose Tissue Inflammation and Insulin Resistance. Clin. Sci. 2018, 132, 1581–1596. [Google Scholar] [CrossRef]
- Noer, J.B.; Talman, M.-L.M.; Moreira, J.M.A. HLA Class II Histocompatibility Antigen γ Chain (CD74) Expression Is Associated with Immune Cell Infiltration and Favorable Outcome in Breast Cancer. Cancers 2021, 13, 6179. [Google Scholar] [CrossRef]
- Tobin, R.P.; Mukherjee, S.; Kain, J.M.; Rogers, S.K.; Henderson, S.K.; Motal, H.L.; Newell Rogers, M.K.; Shapiro, L.A. Traumatic Brain Injury Causes Selective, CD74-Dependent Peripheral Lymphocyte Activation That Exacerbates Neurodegeneration. Acta Neuropathol. Commun. 2014, 2, 143. [Google Scholar] [CrossRef]
- Goldmann, T.; Wieghofer, P.; Müller, P.F.; Wolf, Y.; Varol, D.; Yona, S.; Brendecke, S.M.; Kierdorf, K.; Staszewski, O.; Datta, M.; et al. A New Type of Microglia Gene Targeting Shows TAK1 to Be Pivotal in CNS Autoimmune Inflammation. Nat. Neurosci. 2013, 16, 1618–1626. [Google Scholar] [CrossRef]
- Srinivas, S.; Watanabe, T.; Lin, C.S.; William, C.M.; Tanabe, Y.; Jessell, T.M.; Costantini, F. Cre Reporter Strains Produced by Targeted Insertion of EYFP and ECFP into the ROSA26 Locus. BMC Dev. Biol. 2001, 1, 4. [Google Scholar] [CrossRef]
- Chytil, A.; Magnuson, M.A.; Wright, C.V.E.; Moses, H.L. Conditional Inactivation of the TGF-Beta Type II Receptor Using Cre:Lox. Genesis 2002, 32, 73–75. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jahn, J.; Bollensdorf, A.; Kalischer, C.; Piecha, R.; Weiß-Müller, J.; Potru, P.S.; Ruß, T.; Spittau, B. Microglial CD74 Expression Is Regulated by TGFβ Signaling. Int. J. Mol. Sci. 2022, 23, 10247. https://doi.org/10.3390/ijms231810247
Jahn J, Bollensdorf A, Kalischer C, Piecha R, Weiß-Müller J, Potru PS, Ruß T, Spittau B. Microglial CD74 Expression Is Regulated by TGFβ Signaling. International Journal of Molecular Sciences. 2022; 23(18):10247. https://doi.org/10.3390/ijms231810247
Chicago/Turabian StyleJahn, Jannik, Antonia Bollensdorf, Christopher Kalischer, Robin Piecha, Jana Weiß-Müller, Phani Sankar Potru, Tamara Ruß, and Björn Spittau. 2022. "Microglial CD74 Expression Is Regulated by TGFβ Signaling" International Journal of Molecular Sciences 23, no. 18: 10247. https://doi.org/10.3390/ijms231810247