
Temporal Expression of Myogenic Regulatory Genes in Different Chicken Breeds during Embryonic Development
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Temporal Expression of Myogenic Regulatory Genes during Embryonic Development
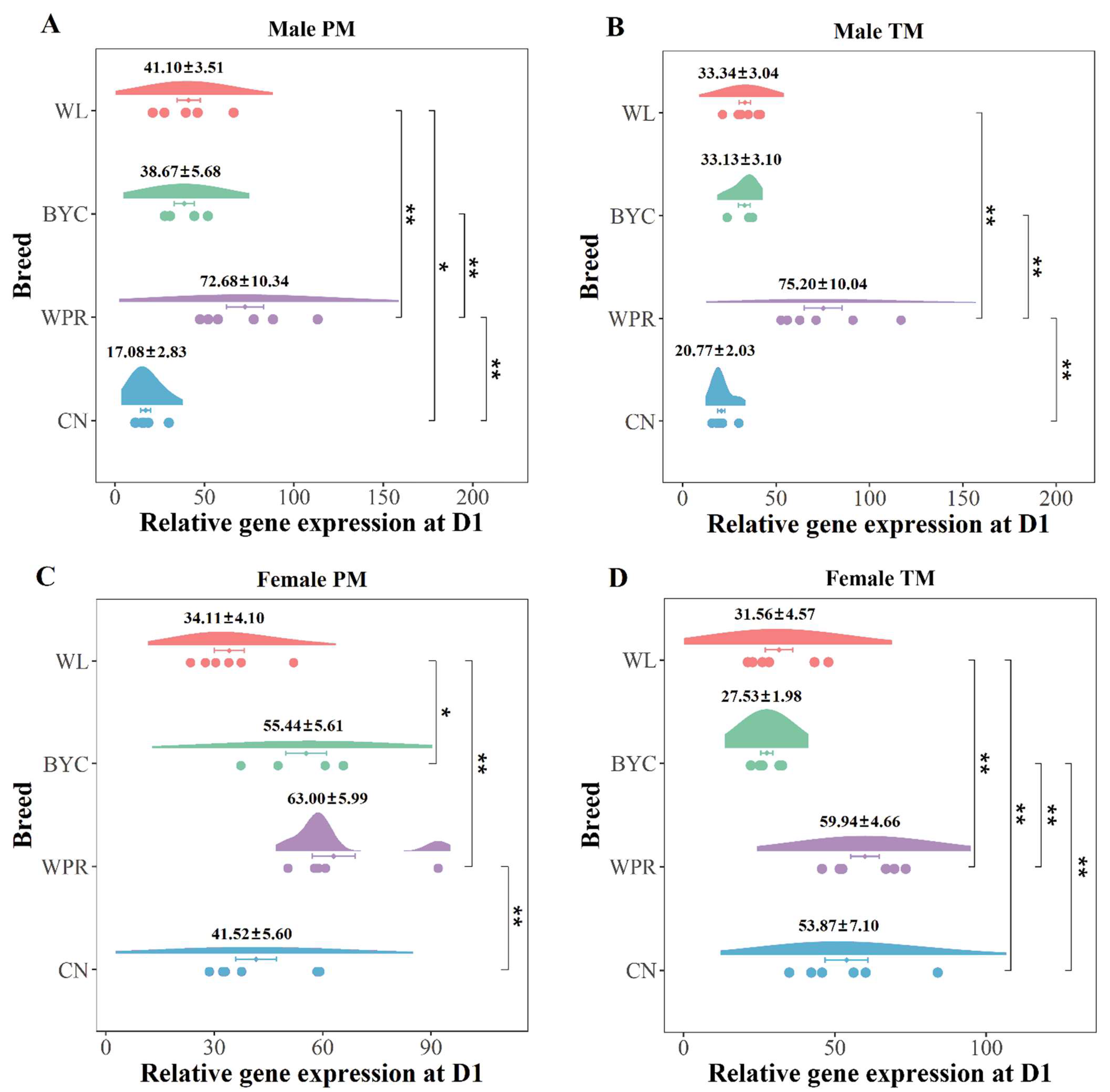
2.2. Meat-Type Chicken Breed Had the Highest Gene Expression of Myomaker at the Crucial Period
2.3. Various Gene Expression of MYOD among the Four Chicken Breeds at D1
2.4. Inhibition of MSTN for the Myoblast Fusion
3. Discussion
4. Materials and Methods
4.1. Bird and Sample Collection
4.2. Sex Determination
4.3. Total RNA Extraction and Cdna Synthesis
4.4. Quantitative Real-Time PCR
4.5. Hematoxylin-Eosin (HE) Staining
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| WL | White Leghorn |
| BYC | Beijing-You Chicken |
| WPR | White Plymouth Rock |
| CN | Cornish |
| PM | Pectoral muscle |
| TM | Thigh muscle |
| PMM | Pectoral muscle of males |
| PMF | Pectoral muscle of females |
| TMM | Thigh muscle of males |
| TMF | Thigh muscle of females |
References
- Janssen, I.; Heymsfield, S.B.; Wang, Z.; Ross, R. Skeletal muscle mass and distribution in 468 men and women aged 18–88 yr. J. Appl. Physiol. 2000, 89, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Velleman, S.G. Muscle Development in the Embryo and Hatchling. Poult. Sci. 2007, 86, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Bi, P.; Ramirez-Martinez, A.; Li, H.; Cannavino, J.; McAnally, J.R.; Shelton, J.M.; Sánchez-Ortiz, E.; Bassel-Duby, R.; Olson, E.N. Control of muscle formation by the fusogenic micropeptide myomixer. Science 2017, 356, 323–327. [Google Scholar] [CrossRef] [PubMed]
- Millay, D.P.; O’Rourke, J.R.; Sutherland, L.B.; Bezprozvannaya, S.; Shelton, J.M.; Bassel-Duby, R.; Olson, E.N. Myomaker is a membrane activator of myoblast fusion and muscle formation. Nature 2013, 499, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Cai, M.; Si, Y.; Zhang, J.; Du, S. Knockout of myomaker results in defective myoblast fusion, reduced muscle growth and increased adipocyte infiltration in zebrafish skeletal muscle. Hum. Mol. Genet. 2018, 27, 3542–3554. [Google Scholar] [CrossRef]
- Si, Y.; Wen, H.; Du, S. Genetic Mutations in jamb, jamc, and myomaker Revealed Different Roles on Myoblast Fusion and Muscle Growth. Mar. Biotechnol. 2019, 21, 111–123. [Google Scholar] [CrossRef]
- Landemaine, A.; Rescan, P.-Y.; Gabillard, J.-C. Myomaker mediates fusion of fast myocytes in zebrafish embryos. Biochem. Biophys. Res. Commun. 2014, 451, 480–484. [Google Scholar] [CrossRef]
- Luo, W.; Li, E.; Nie, Q.; Zhang, X. Myomaker, Regulated by MYOD, MYOG and miR-140-3p, Promotes Chicken Myoblast Fusion. Int. J. Mol. Sci. 2015, 16, 26186–26201. [Google Scholar] [CrossRef]
- Hernández-Hernández, J.M.; García-González, E.G.; Brun, C.E.; Rudnicki, M.A. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Semin. Cell Dev. Biol. 2017, 72, 10–18. [Google Scholar] [CrossRef]
- Braun, T.; Gautel, M. Transcriptional mechanisms regulating skeletal muscle differentiation, growth and homeostasis. Nat. Rev. Mol. Cell Biol. 2011, 12, 349–361. [Google Scholar] [CrossRef]
- Legerlotz, K.; Smith, H.K. Role of MyoD in denervated, disused, and exercised muscle. Muscle Nerve 2008, 38, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.C.; Rould, M.A.; Weintraub, H.; Pabo, C.O. Crystal structure of MyoD bHLH domain-DNA complex: Perspectives on DNA recognition and implications for transcriptional activation. Cell 1994, 77, 451–459. [Google Scholar] [CrossRef]
- Kambadur, R.; Sharma, M.; Smith, T.P.; Bass, J.J. Mutations in myostatin (GDF8) in Double-Muscled Belgian Blue and Piedmontese Cattle. Genome Res. 1997, 7, 910–915. [Google Scholar] [CrossRef] [PubMed]
- Trendelenburg, A.U.; Meyer, A.; Rohner, D.; Boyle, J.; Hatakeyama, S.; Glass, D.J. Myostatin reduces Akt/TORC1/p70S6K signaling, inhibiting myoblast differentiation and myotube size. Am. J. Physiol. Physiol. 2009, 296, C1258–C1270. [Google Scholar] [CrossRef]
- Kim, G.-D.; Lee, J.H.; Song, S.; Kim, S.W.; Han, J.S.; Shin, S.P.; Park, B.-C.; Park, T.S. Generation of myostatin-knockout chickens mediated by D10A-Cas9 nickase. FASEB J. 2020, 34, 5688–5696. [Google Scholar] [CrossRef] [PubMed]
- Posey, A.D.; Demonbreun, A.; McNally, E.M. Ferlin Proteins in Myoblast Fusion and Muscle Growth. Curr. Top. Dev. Biol. 2011, 96, 203–230. [Google Scholar] [CrossRef]
- Melendez, J.; Sieiro, D.; Salgado, D.; Morin, V.; Dejardin, M.-J.; Zhou, C.; Mullen, A.C.; Marcelle, C. TGFβ signalling acts as a molecular brake of myoblast fusion. Nat. Commun. 2021, 12, 749. [Google Scholar] [CrossRef]
- Chen, W.; Lv, Y.T.; Zhang, H.X.; Ruan, D.; Wang, S.; Lin, Y.C. Developmental specificity in skeletal muscle of late-term avian embryos and its potential manipulation. Poult. Sci. 2013, 92, 2754–2764. [Google Scholar] [CrossRef]
- Givisiez, P.E.N.; Moreira Filho, A.L.B.; Santos, M.R.B.; Oliveira, H.B.; Ferket, P.R.; Oliveira, C.J.B.; Malheiros, R.D. Chicken embryo development: Metabolic and morphological basis for in ovo feeding technology. Poult. Sci. 2020, 99, 6774–6782. [Google Scholar] [CrossRef]
- Dai, D.; Zhang, H.-J.; Qiu, K.; Qi, G.-H.; Wang, J.; Wu, S.-G. Supplemental L-Arginine Improves the Embryonic Intestine Development and Microbial Succession in a Chick Embryo Model. Front. Nutr. 2021, 8, 692305. [Google Scholar] [CrossRef]
- Smith, J.H. Relation of Body Size to Muscle Cell Size and Number in the Chicken. Poult. Sci. 1963, 42, 283–290. [Google Scholar] [CrossRef]
- Wardle, F.C. Master control: Transcriptional regulation of mammalian Myod. J. Muscle Res. Cell Motil. 2019, 40, 211–226. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, O.; Fujisawa-Sehara, A.; Nabeshima, Y.-I.; Periasamy, M. Expression of myogenic factors in denervated chicken breast muscle: Isolation of the chicken Myf5 gene. Nucleic Acids Res. 1993, 21, 2503–2509. [Google Scholar] [CrossRef] [PubMed]
- Langley, B.; Thomas, M.; Bishop, A.; Sharma, M.; Gilmour, S.; Kambadur, R. Myostatin Inhibits Myoblast Differentiation by Down-regulating MyoD Expression. J. Biol. Chem. 2002, 277, 49831–49840. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Aparnathi, M.K.; Patel, A.K.; Joshi, C.G. In vitro silencing of myostatin gene by shRNAs in chicken embryonic myoblast cells. Biotechnol. Prog. 2013, 29, 425–431. [Google Scholar] [CrossRef]
- Sundaresan, N.; Saxena, V.; Singh, R.; Jain, P.; Singh, K.; Anish, D.; Singh, N.; Saxena, M.; Ahmed, K. Expression profile of myostatin mRNA during the embryonic organogenesis of domestic chicken (Gallus gallus domesticus). Res. Veter- Sci. 2008, 85, 86–91. [Google Scholar] [CrossRef]
- Raja, J.S.; Hoffman, M.L.; Govoni, K.E.; Zinn, S.A.; Reed, S.A. Restricted maternal nutrition alters myogenic regulatory factor expression in satellite cells of ovine offspring. Animal 2016, 10, 1200–1203. [Google Scholar] [CrossRef]
- Rodrigues, G.D.A.; Júnior, D.T.V.; Soares, M.H.; da Silva, C.B.; Fialho, F.A.; Barbosa, L.M.D.R.; Neves, M.M.; Rocha, G.C.; Duarte, M.D.S.; Saraiva, A. L-Arginine Supplementation for Nulliparous Sows during the Last Third of Gestation. Animals 2021, 11, 3476. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, S.; Shibasaki, A.; Naka, A.; Saito, H.; Iida, K. Lactate Promotes Myoblast Differentiation and Myotube Hypertrophy via a Pathway Involving MyoD In Vitro and Enhances Muscle Regeneration In Vivo. Int. J. Mol. Sci. 2018, 19, 3649. [Google Scholar] [CrossRef]
- Liu, X.; Wang, J.; Li, R.; Yang, X.; Sun, Q.; Albrecht, E.; Zhao, R. Maternal dietary protein affects transcriptional regulation of myostatin gene distinctively at weaning and finishing stages in skeletal muscle of Meishan pigs. Epigenetics 2011, 6, 899–907. [Google Scholar] [CrossRef] [Green Version]
- Larsen, A.; Tunstall, R.; Carey, K.; Nicholas, G.; Kambadur, R.; Crowe, T.; Cameron-Smith, D. Actions of Short-Term Fasting on Human Skeletal Muscle Myogenic and Atrogenic Gene Expression. Ann. Nutr. Metab. 2006, 50, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, I.; González, T.; López, M.; Señarís, R.; Devesa, J.; Arce, V.M. Myostatin expression is regulated by underfeeding and neonatal programming in rats. J. Physiol. Biochem. 2012, 69, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Buzała, M.; Janicki, B.; Czarnecki, R. Consequences of different growth rates in broiler breeder and layer hens on embryogenesis, metabolism and metabolic rate: A review. Poult. Sci. 2015, 94, 728–733. [Google Scholar] [CrossRef]
- Emmerson, D. Commercial approaches to genetic selection for growth and feed conversion in domestic poultry. Poult. Sci. 1997, 76, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Zhang, Y.; Chen, Y.; Yang, N.; Wang, X.-J.; Zhu, D. Systematic identification of genes involved in divergent skeletal muscle growth rates of broiler and layer chickens. BMC Genom. 2009, 10, 87. [Google Scholar] [CrossRef] [PubMed]
- Al-Musawi, S.L.; Lock, F.; Simbi, B.H.; Bayol, S.A.; Stickland, N.C. Muscle specific differences in the regulation of myogenic differentiation in chickens genetically selected for divergent growth rates. Differentiation 2011, 82, 127–135. [Google Scholar] [CrossRef]
- Fridolfsson, A.-K.; Cheng, H.; Copeland, N.G.; Jenkins, N.A.; Liu, H.-C.; Raudsepp, T.; Woodage, T.; Chowdhary, B.; Halverson, J.; Ellegren, H. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proc. Natl. Acad. Sci. USA 1998, 95, 8147–8152. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Gu, S.; Wen, C.L.; Li, J.Y.; Wu, G.Q.; Li, G.Q.; Han, W.P.; Yan, Y.Y.; Sun, C.J.; Yang, N. Comparison of muscle fiber characteristics during late embryonic development among four breeds of chickens. J. Northeast. Agric. Univ. 2021, 52, 9–16. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) | Annealing Temperature (°C) |
|---|---|---|
| CHD1 | F-GTTACTGATTCGTCTACGAGA | 52 |
| R-ATTGAAATGATCCAGTGCTTG | ||
| β-actin | F-ATCTTTCTTGGGTATGGAGTC | 60 |
| R-GCCAGGGTACATTGTGG | ||
| Myomaker | F-TGGGTGTCCCTGATGGC | 60 |
| R-CCCGATGGGTCCTGAGTAG | ||
| MYOD | F-GCTACTACACGGAATCACCAAAT | 60 |
| R-CTGGGCTCCACTGTCACTCA | ||
| MSTN | F-ACAGTAGCGATGGCTCTTT | 60 |
| R-CCGTTGTAGGTTTTTGGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.; Wen, C.; Li, J.; Liu, H.; Huang, Q.; Zheng, J.; Sun, C.; Yang, N. Temporal Expression of Myogenic Regulatory Genes in Different Chicken Breeds during Embryonic Development. Int. J. Mol. Sci. 2022, 23, 10115. https://doi.org/10.3390/ijms231710115
Gu S, Wen C, Li J, Liu H, Huang Q, Zheng J, Sun C, Yang N. Temporal Expression of Myogenic Regulatory Genes in Different Chicken Breeds during Embryonic Development. International Journal of Molecular Sciences. 2022; 23(17):10115. https://doi.org/10.3390/ijms231710115
Chicago/Turabian StyleGu, Shuang, Chaoliang Wen, Junying Li, Honghong Liu, Qiang Huang, Jiangxia Zheng, Congjiao Sun, and Ning Yang. 2022. "Temporal Expression of Myogenic Regulatory Genes in Different Chicken Breeds during Embryonic Development" International Journal of Molecular Sciences 23, no. 17: 10115. https://doi.org/10.3390/ijms231710115