
Repair of Retinal Degeneration by Human Amniotic Epithelial Stem Cell–Derived Photoreceptor–like Cells

 , , , and
, , , and 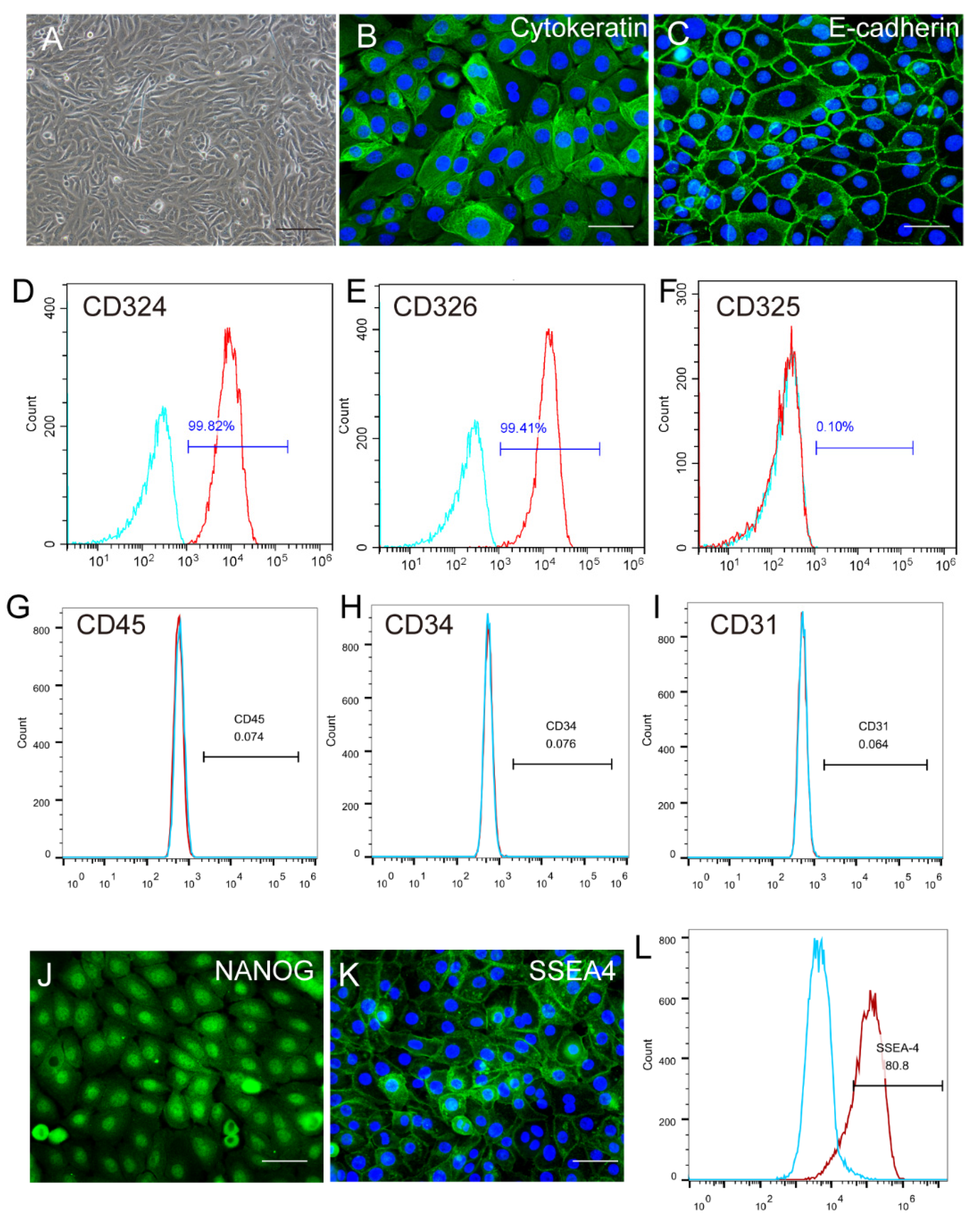{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of Human Amniotic Epithelial Stem Cells
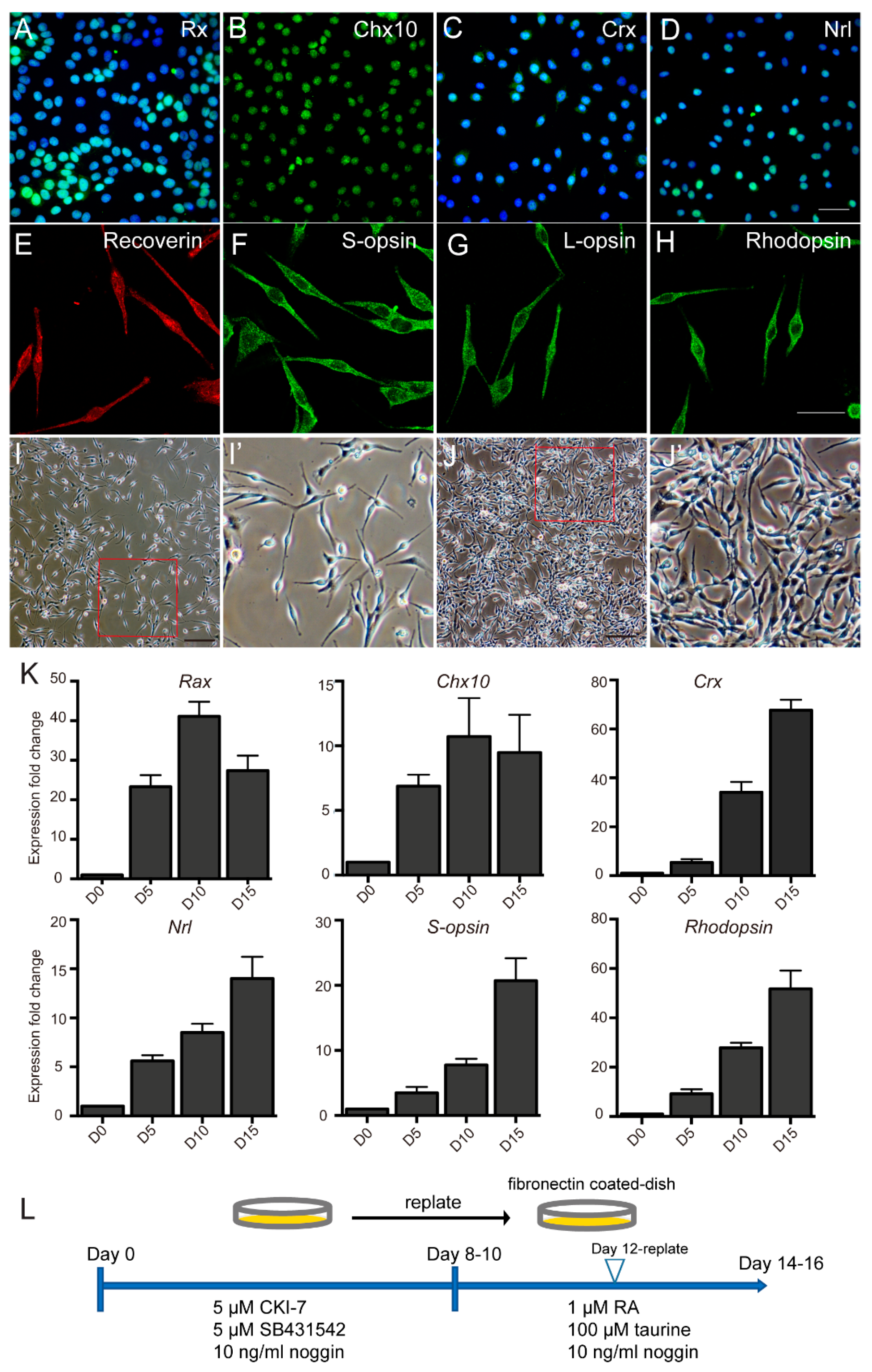
2.2. PR–like Cells Were Obtained from hAESCs by a Two–Step Combined Induction
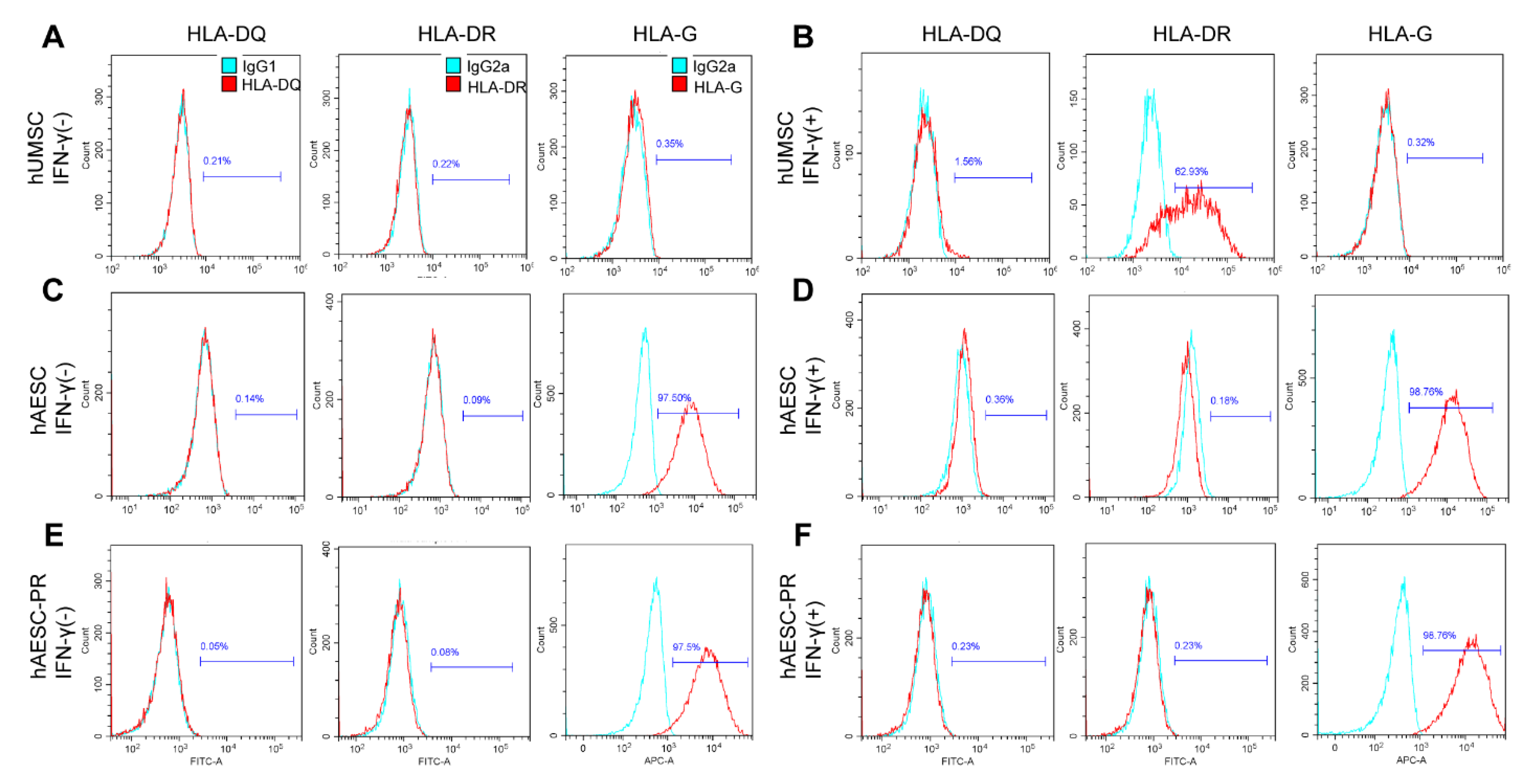
2.3. The Immune Privilege of hAESCs–Derived PR–Like Cells
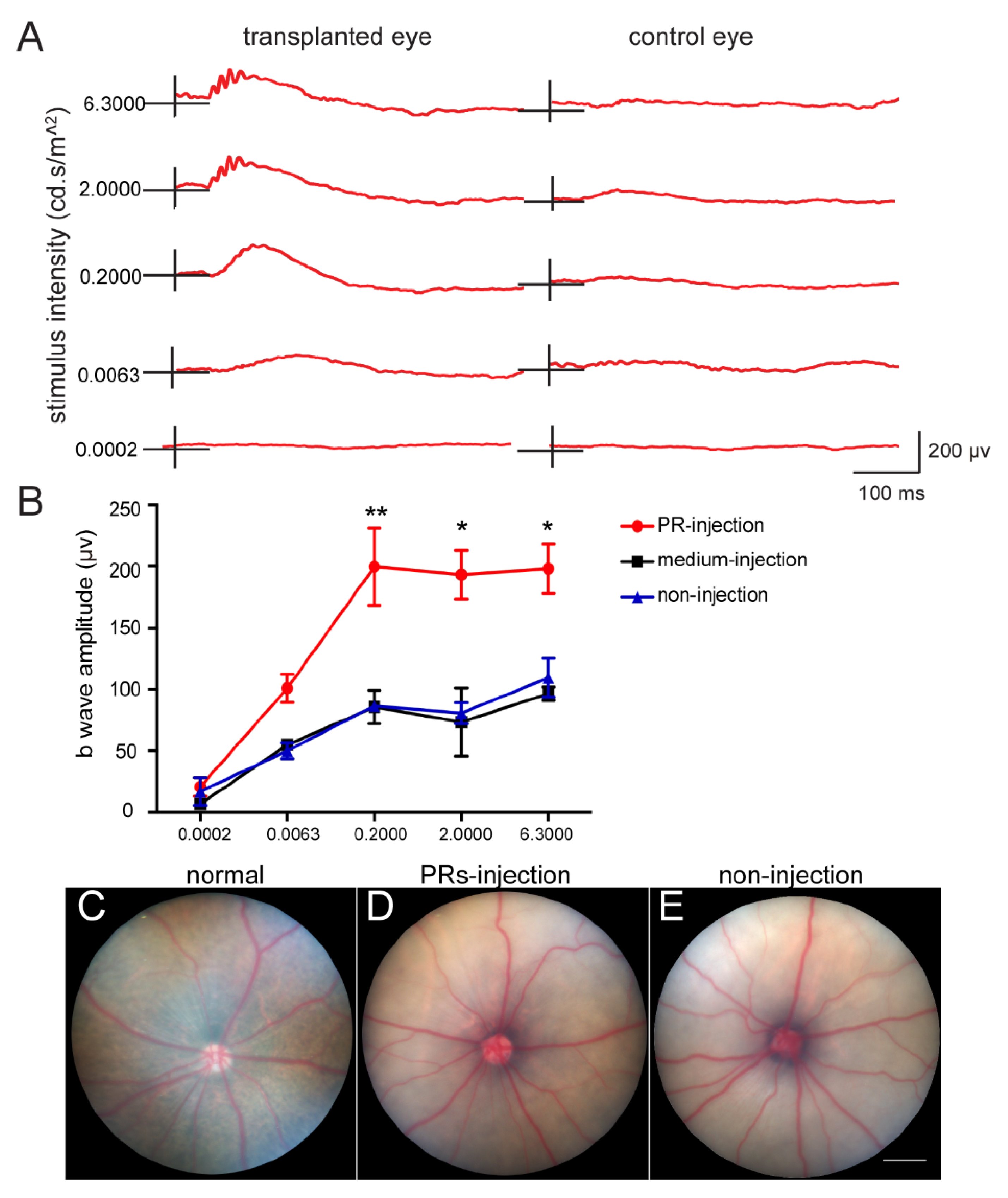
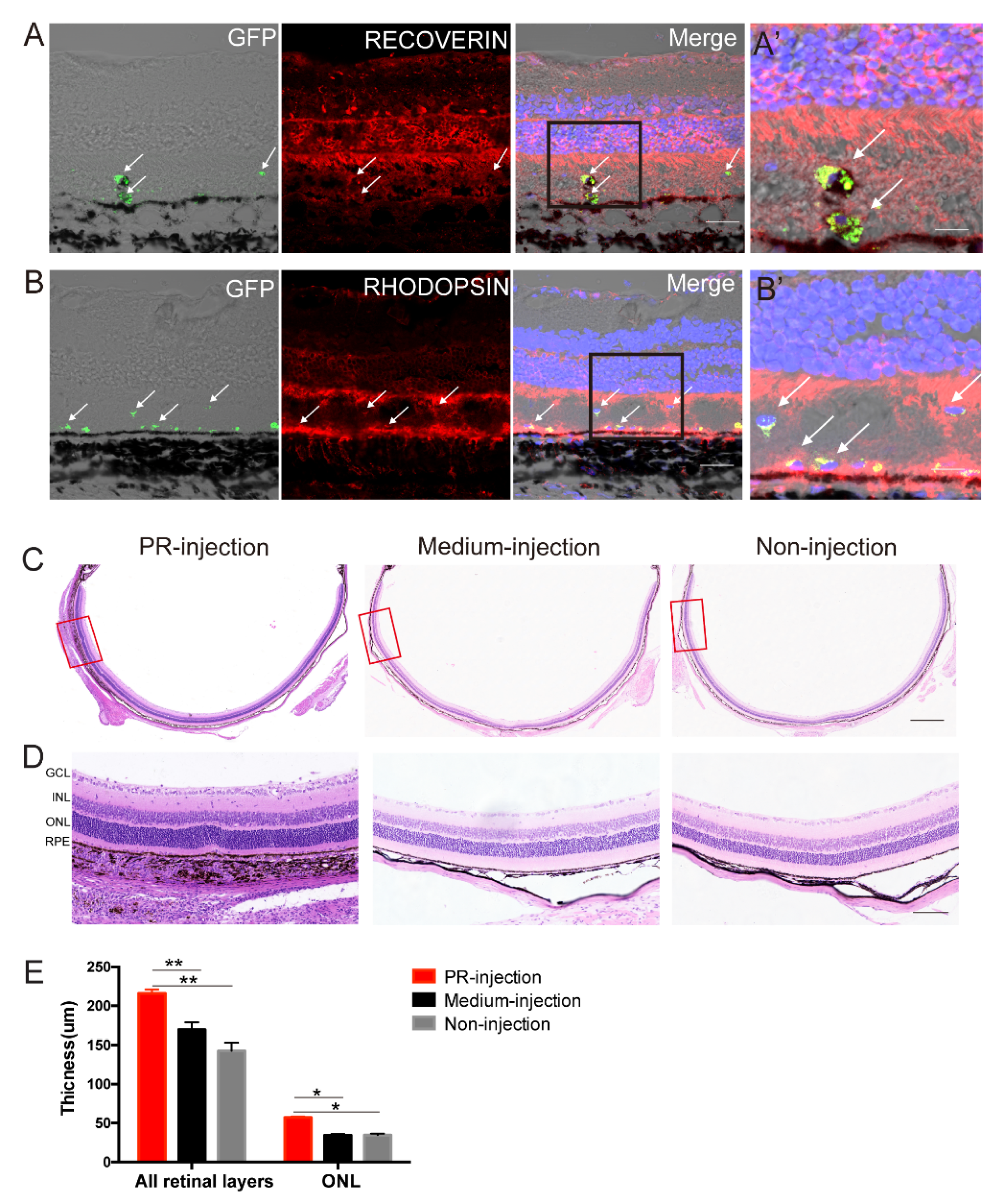
2.4. Subretinal Transplantation of hAESCs–PR–like Cells Resulted in Functional Retinal Rescue in RCS Rats
3. Discussion
4. Materials and Methods
4.1. Separation of hAESCs
4.2. Differentiation of PR–like Cells from hAESCs
4.3. Quantitative Reverse–Transcription PCR
4.4. Immunostaining
4.5. Flow Cytometry
4.6. Animals
4.7. Transplantation of hAESCs–PR–Like Cells in RCS Rats
4.8. Electroretinograms (ERG)
4.9. Histology and Immunohistochemical (IHC) Staining
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zarbin, M. Cell–Based Therapy for Degenerative Retinal Disease. Trends Mol. Med. 2016, 22, 115–134. [Google Scholar] [CrossRef] [PubMed]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsden, C.M.; Powner, M.B.; Carr, A.J.; Smart, M.J.; da Cruz, L.; Coffey, P.J. Stem cells in retinal regeneration: Past, present and future. Development 2013, 140, 2576–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLaren, R.E.; Pearson, R.A.; MacNeil, A.; Douglas, R.H.; Salt, T.E.; Akimoto, M.; Swaroop, A.; Sowden, J.C.; Ali, R.R. Retinal repair by transplantation of photoreceptor precursors. Nature 2006, 444, 203–207. [Google Scholar] [CrossRef]
- Barber, A.C.; Hippert, C.; Duran, Y.; West, E.L.; Bainbridge, J.W.B.; Warre–Cornish, K.; Luhmann, U.F.O.; Lakowski, J.; Sowden, J.C.; Ali, R.R.; et al. Repair of the degenerate retina by photoreceptor transplantation. Proc. Natl. Acad. Sci. USA 2013, 110, 354–359. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.S.; Issa, P.C.; Butler, R.; Martin, C.; Lipinski, D.M.; Sekaran, S.; Barnard, A.R.; MacLaren, R.E. Reversal of end–stage retinal degeneration and restoration of visual function by photoreceptor transplantation. Proc. Natl. Acad. Sci. USA 2013, 110, 1101–1106. [Google Scholar] [CrossRef] [Green Version]
- Mandai, M.; Watanabe, A.; Kurimoto, Y.; Hirami, Y.; Morinaga, C.; Daimon, T.; Fujihara, M.; Akimaru, H.; Sakai, N.; Shibata, Y.; et al. Autologous Induced Stem–Cell–Derived Retinal Cells for Macular Degeneration. New Engl. J. Med. 2017, 376, 1038–1046. [Google Scholar] [CrossRef]
- Da Cruz, L.; Fynes, K.; Georgiadis, O.; Kerby, J.; Luo, Y.H.; Ahmado, A.; Vernon, A.; Daniels, J.T.; Nommiste, B.; Hasan, S.M.; et al. Phase 1 clinical study of an embryonic stem cell–derived retinal pigment epithelium patch in age–related macular degeneration. Nat. Biotechnol. 2018, 36, 328–337. [Google Scholar] [CrossRef] [Green Version]
- Mehat, M.S.; Sundaram, V.; Ripamonti, C.; Robson, A.G.; Smith, A.J.; Borooah, S.; Robinson, M.; Rosenthal, A.N.; Innes, W.; Weleber, R.G.; et al. Transplantation of Human Embryonic Stem Cell–Derived Retinal Pigment Epithelial Cells in Macular Degeneration. Ophthalmology 2018, 125, 1765–1775. [Google Scholar] [CrossRef] [Green Version]
- Miki, T. Stem cell characteristics and the therapeutic potential of amniotic epithelial cells. Am. J. Reprod. Immunol. 2018, 80, e13003. [Google Scholar] [CrossRef]
- Tan, B.; Yuan, W.X.; Li, J.Y.; Yang, P.J.; Ge, Z.; Liu, J.; Qiu, C.; Zhu, X.L.; Qiu, C.; Lai, D.M.; et al. Therapeutic effect of human amniotic epithelial cells in murine models of Hashimoto's thyroiditis and Systemic lupus erythematosus. Cytotherapy 2018, 20, 1247–1258. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Qiu, C.; Zhang, Z.; Yuan, W.; Ge, Z.; Tan, B.; Yang, P.; Liu, J.; Zhu, X.; Qiu, C.; et al. Subretinal Transplantation of Human Amniotic Epithelial Cells in the Treatment of Autoimmune Uveitis in Rats. Cell Transpl. 2018, 27, 1504–1514. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.-J.; Yuan, W.-X.; Liu, J.; Li, J.-Y.; Tan, B.; Qiu, C.; Zhu, X.-L.; Qiu, C.; Lai, D.-M.; Guo, L.-H.; et al. Biological characterization of human amniotic epithelial cells in a serum–free system and their safety evaluation. Acta Pharmacol. Sin. 2018, 39, 1305–1316. [Google Scholar] [CrossRef]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Buhring, H.J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise review: Isolation and characterization of cells from human term placenta: Outcome of the first international Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem cell characteristics of amniotic epithelial cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [Green Version]
- Miki, T.; Strom, S.C. Amnion–derived pluripotent/multipotent stem cells. Stem Cell Rev. 2006, 2, 133–142. [Google Scholar] [CrossRef]
- Ishii, T.; Ohsugi, K.; Nakamura, S.; Sato, K.; Hashimoto, M.; Mikoshiba, K.; Sakuragawa, N. Gene expression of oligodendrocyte markers in human amniotic epithelial cells using neural cell–type–specific expression system. Neurosci. Lett. 1999, 268, 131–134. [Google Scholar] [CrossRef]
- Sakuragawa, N.; Misawa, H.; Ohsugi, K.; Kakishita, K.; Ishii, T.; Thangavel, R.; Tohyama, J.; Elwan, M.; Yokoyama, Y.; Okuda, O.; et al. Evidence for active acetylcholine metabolism in human amniotic epithelial cells: Applicable to intracerebral allografting for neurologic disease. Neurosci. Lett. 1997, 232, 53–56. [Google Scholar] [CrossRef]
- Niknejad, H.; Peirovi, H.; Ahmadiani, A.; Ghanavi, J.; Jorjani, M. Differentiation Factors That Influence Neuronal Markers Expression in Vitro from Human Amniotic Epithelial Cells. Eur. Cells Mater. 2010, 19, 22–29. [Google Scholar] [CrossRef]
- Strom, S.C.; Gramignoli, R. Human amnion epithelial cells expressing HLA–G as novel cell–based treatment for liver disease. Hum. Immunol. 2016, 77, 734–739. [Google Scholar] [CrossRef]
- Akle, C.A.; Welsh, K.I.; Adinolfi, M.; Leibowitz, S. Immunogenicity of Human Amniotic Epithelial–Cells after Transplantation into Volunteers. Lancet 1981, 2, 1003–1005. [Google Scholar] [CrossRef]
- Osakada, F.; Ikeda, H.; Sasai, Y.; Takahashi, M. Stepwise differentiation of pluripotent stem cells into retinal cells. Nat. Protoc. 2009, 4, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Osakada, F.; Jin, Z.B.; Hirami, Y.; Ikeda, H.; Danjyo, T.; Watanabe, K.; Sasai, Y.; Takahashi, M. In vitro differentiation of retinal cells from human pluripotent stem cells by small–molecule induction. J. Cell Sci. 2009, 122, 3169–3179. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Gutierrez, C.; Xue, T.; Hampton, C.; Vergara, M.N.; Cao, L.H.; Peters, A.; Park, T.S.; Zambidis, E.T.; Meyer, J.S.; et al. Generation of three–dimensional retinal tissue with functional photoreceptors from human iPSCs. Nat. Commun. 2014, 5, 4047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elwan, M.A.; Sakuragawa, N. Evidence for synthesis and release of catecholamines by human amniotic epithelial cells. Neuroreport 1997, 8, 3435–3438. [Google Scholar] [CrossRef]
- Elwan, M.A. Synthesis of dopamine from L–3,4–dihydroxyphenylalanine by human amniotic epithelial cells. Eur. J. Pharm. 1998, 354, R1–R2. [Google Scholar] [CrossRef]
- Wu, Z.; Hui, G.; Lu, Y.; Liu, T.; Huang, Q.; Guo, L. Human amniotic epithelial cells express specific markers of nerve cells and migrate along the nerve fibers in the corpus callosum. Neural Regen. Res. 2012, 7, 41–45. [Google Scholar]
- Gonzalez–Cordero, A.; West, E.L.; Pearson, R.A.; Duran, Y.; Carvalho, L.S.; Chu, C.J.; Naeem, A.; Blackford, S.J.I.; Georgiadis, A.; Lakowski, J.; et al. Photoreceptor precursors derived from three–dimensional embryonic stem cell cultures integrate and mature within adult degenerate retina. Nat. Biotechnol. 2013, 31, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Chen, H.; Yang, X.; Guo, L.; Hui, G. Treatment of intracerebral haemorrhage in rats with intraventricular transplantation of human amniotic epithelial cells. Cell Biol. Int. 2010, 34, 573–577. [Google Scholar] [CrossRef]
- Zhang, R.; Cai, Y.; Xiao, R.; Zhong, H.; Li, X.; Guo, L.; Xu, H.; Fan, X. Human amniotic epithelial cell transplantation promotes neurogenesis and ameliorates social deficits in BTBR mice. Stem Cell Res. 2019, 10, 153. [Google Scholar] [CrossRef]
- Liu, T.; Wu, J.; Huang, Q.; Hou, Y.; Jiang, Z.; Zang, S.; Guo, L. Human amniotic epithelial cells ameliorate behavioral dysfunction and reduce infarct size in the rat middle cerebral artery occlusion model. Shock 2008, 29, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Gu, E.; Chen, W.Y.; Gu, J.; Burridge, P.; Wu, J.C. Molecular Imaging of Stem Cells: Tracking Survival, Biodistribution, Tumorigenicity, and Immunogenicity. Theranostics 2012, 2, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nauta, A.J.; Westerhuis, G.; Kruisselbrink, A.B.; Lurvink, E.G.A.; Willemze, R.; Fibbe, W.E. Donor–derived mesenchymal stem cells are immunogenic in an allogeneic host and stimulate donor graft rejection in a nonmyeloablative setting. Blood 2006, 108, 2114–2120. [Google Scholar] [CrossRef]
- Pearson, R.A.; Gonzalez–Cordero, A.; West, E.L.; Ribeiro, J.R.; Aghaizu, N.; Goh, D.; Sampson, R.D.; Georgiadis, A.; Waldron, P.V.; Duran, Y.; et al. Donor and host photoreceptors engage in material transfer following transplantation of post–mitotic photoreceptor precursors. Nat. Commun. 2016, 7, 13029. [Google Scholar] [CrossRef] [Green Version]
- Santos–Ferreira, T.; Llonch, S.; Borsch, O.; Postel, K.; Haas, J.; Ader, M. Retinal transplantation of photoreceptors results in donor–host cytoplasmic exchange. Nat. Commun. 2016, 7, 13028. [Google Scholar] [CrossRef] [Green Version]
- Singh, M.S.; Balmer, J.; Barnard, A.R.; Aslam, S.A.; Moralli, D.; Green, C.M.; Barnea–Cramer, A.; Duncan, I.; MacLaren, R.E. Transplanted photoreceptor precursors transfer proteins to host photoreceptors by a mechanism of cytoplasmic fusion. Nat. Commun. 2016, 7, 13537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamba, D.A.; Gust, J.; Reh, T.A. Transplantation of human embryonic stem cell–derived photoreceptors restores some visual function in Crx–deficient mice. Cell Stem Cell 2009, 4, 73–79. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Qiu, C.; Zhou, J.; Wei, Y.; Yuan, W.; Liu, J.; Cui, W.; Huang, J.; Qiu, C.; Guo, L.; et al. Repair of Retinal Degeneration by Human Amniotic Epithelial Stem Cell–Derived Photoreceptor–like Cells. Int. J. Mol. Sci. 2022, 23, 8722. https://doi.org/10.3390/ijms23158722
Li J, Qiu C, Zhou J, Wei Y, Yuan W, Liu J, Cui W, Huang J, Qiu C, Guo L, et al. Repair of Retinal Degeneration by Human Amniotic Epithelial Stem Cell–Derived Photoreceptor–like Cells. International Journal of Molecular Sciences. 2022; 23(15):8722. https://doi.org/10.3390/ijms23158722
Chicago/Turabian StyleLi, Jinying, Chen Qiu, Jiayi Zhou, Yang Wei, Weixin Yuan, Jia Liu, Wenyu Cui, Jianan Huang, Cong Qiu, Lihe Guo, and et al. 2022. "Repair of Retinal Degeneration by Human Amniotic Epithelial Stem Cell–Derived Photoreceptor–like Cells" International Journal of Molecular Sciences 23, no. 15: 8722. https://doi.org/10.3390/ijms23158722