
Development and Assessment of Herpes Simplex Virus Type 1 (HSV-1) Amplicon Vectors with Sensory Neuron-Selective Promoters


 , ,
, ,
Abstract
:1. Introduction
2. Results
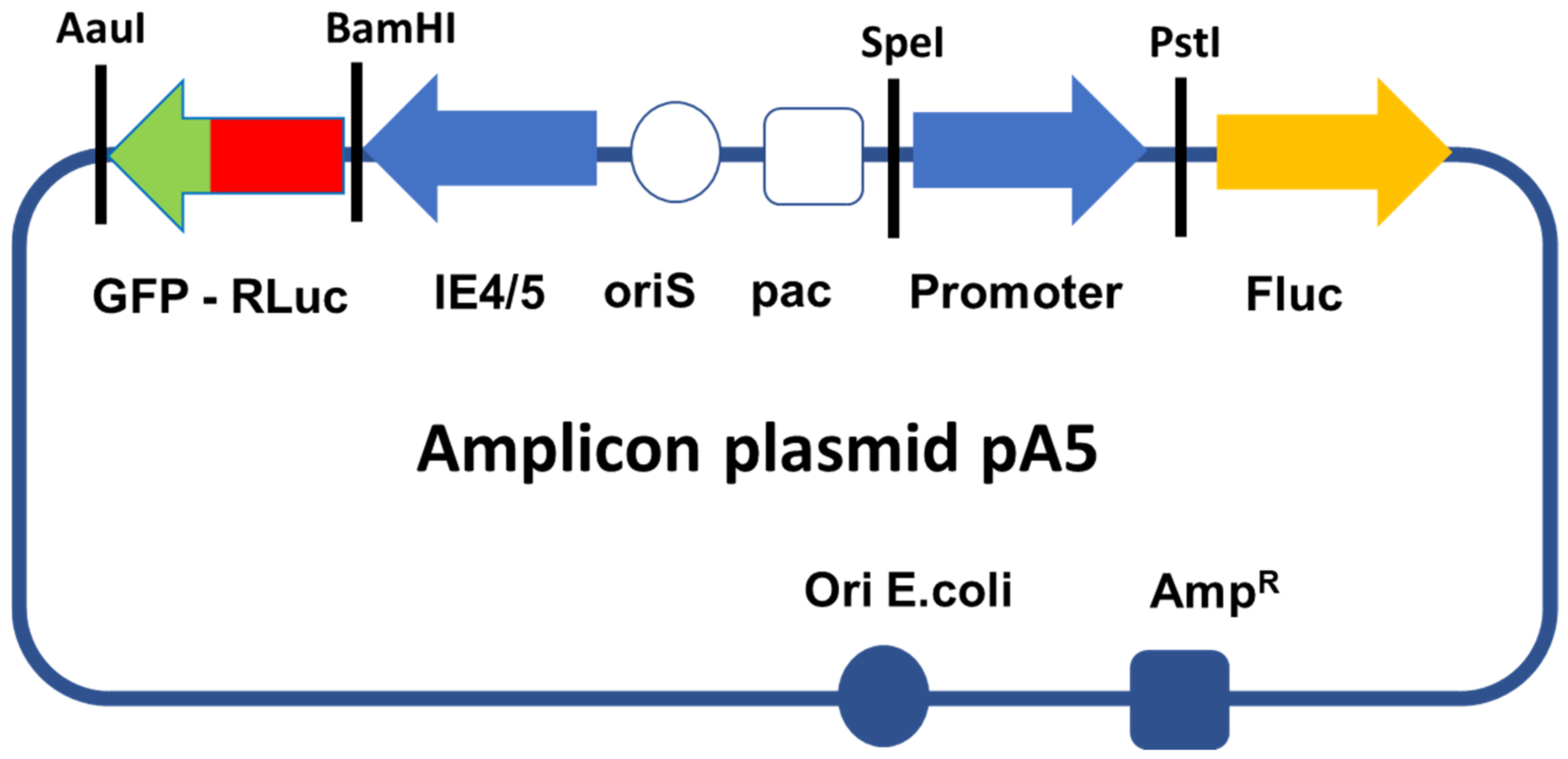
2.1. Construction of Amplicon Vectors
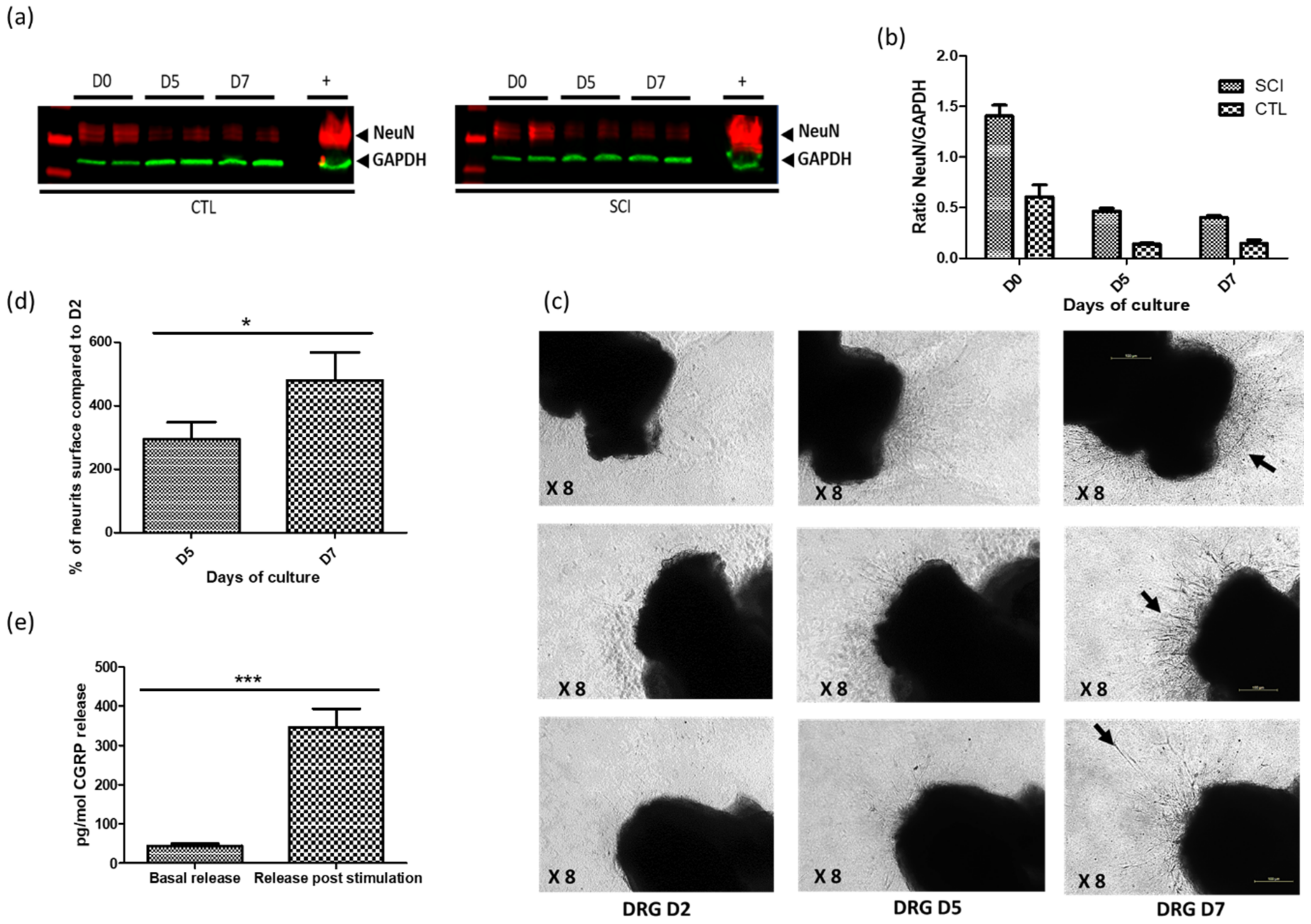
2.2. Establishment of Optimized DRG Organotypic Cultures
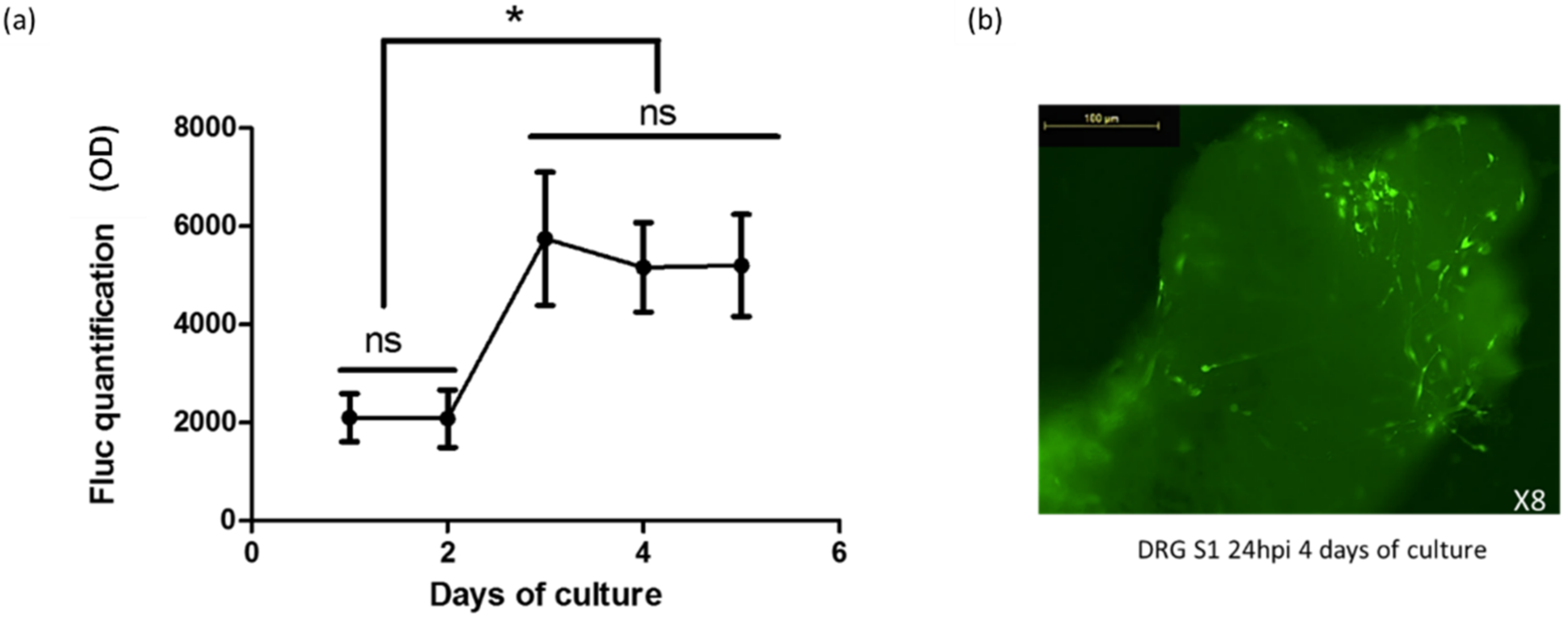
2.3. Gene Transfer in Organotypic DRG Cultures by HSV-1-Based Vectors
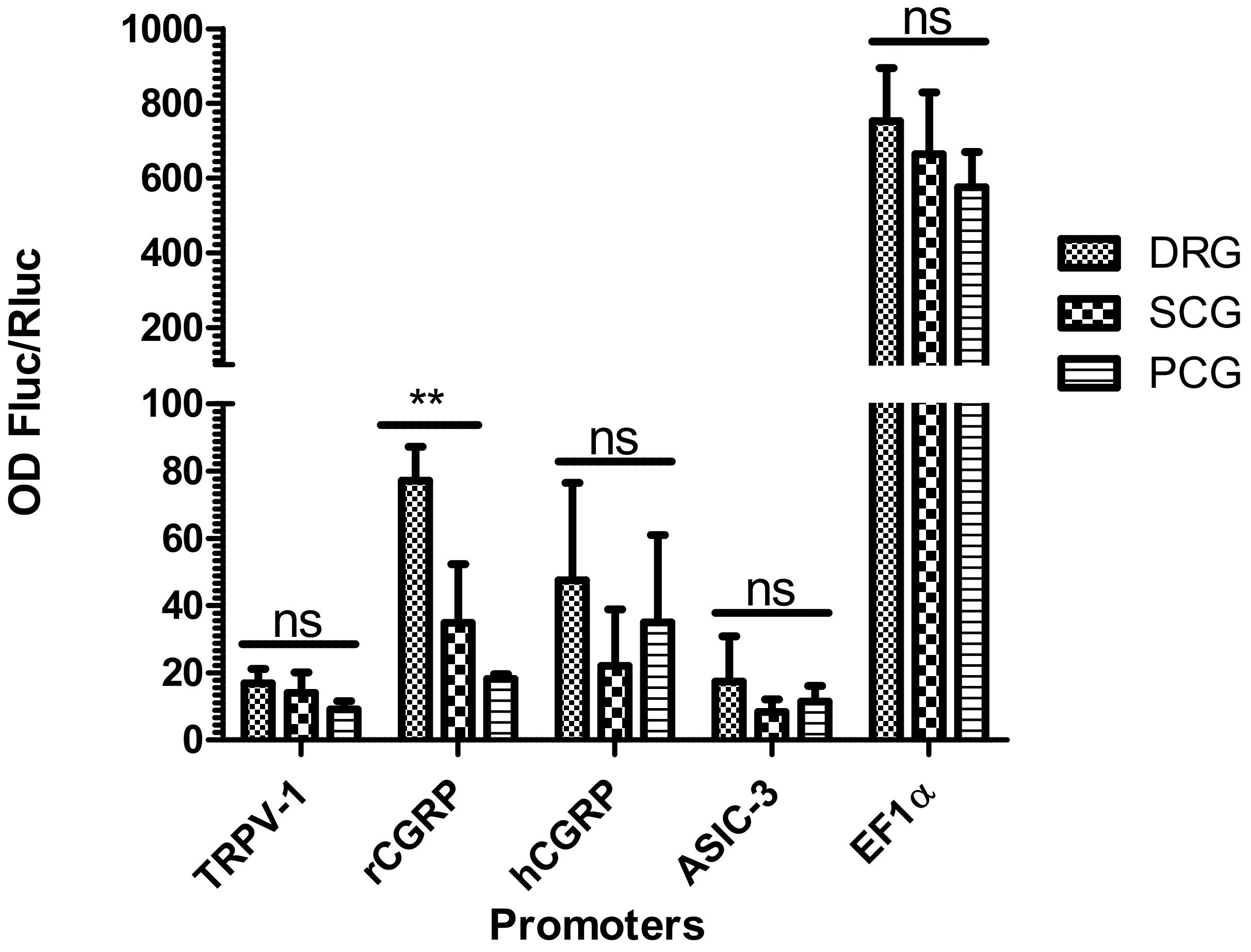
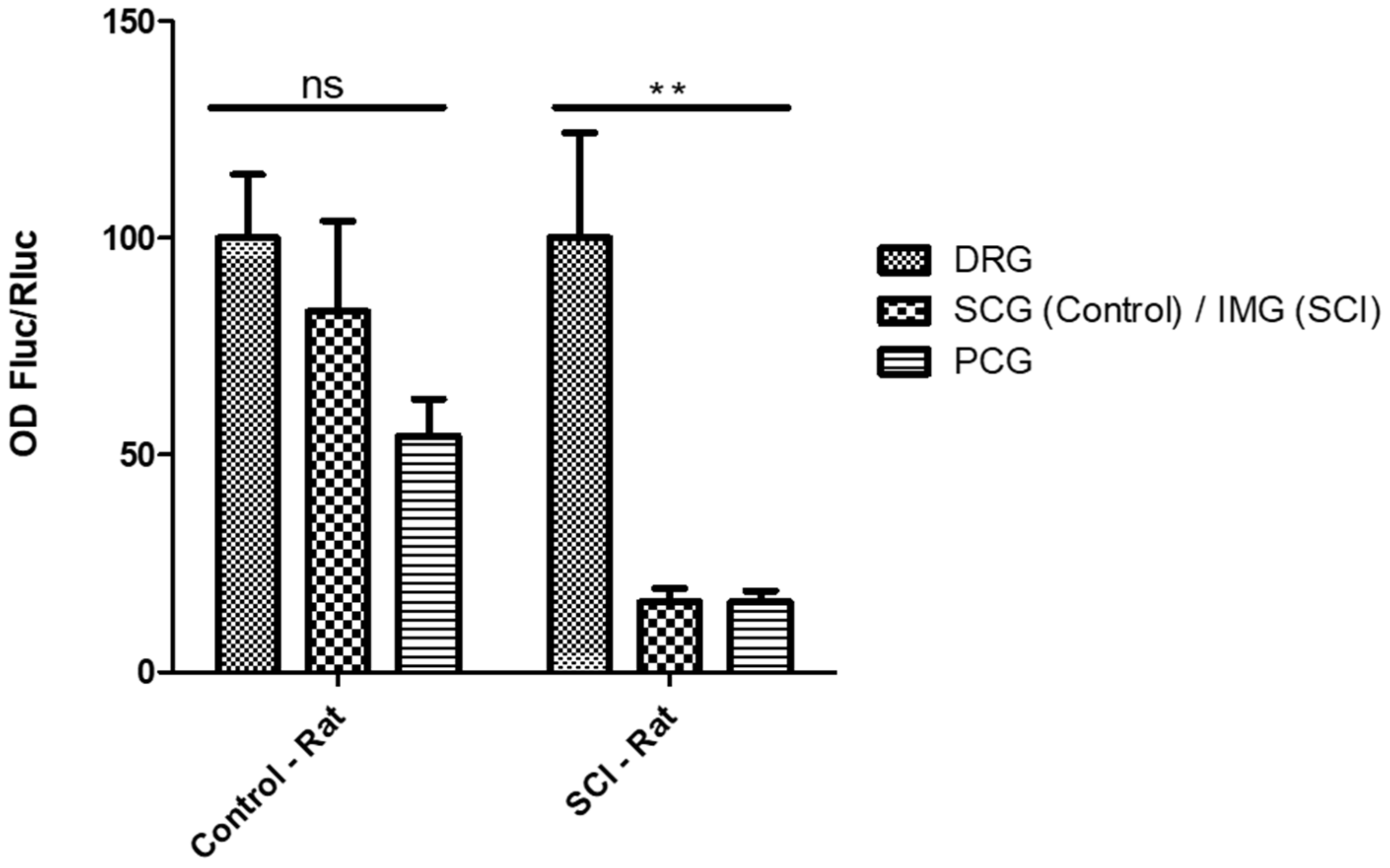
2.4. Selective Expression of RCGRP Promoter in Adult Rat DRG Ganglia
2.5. Selective Expression of the RTRPV-1 Promoter in DRG Ganglia from SCI Rats
3. Discussion
4. Materials and Methods
4.1. Organotypic Cultures of Adult Rat Ganglia
4.2. Construction and Titration of Non-Replicative HSV-1 Amplicon Vectors
- (a)
- Plasmids containing sensory neuron-specific promoters. A family of pUC57-proms plasmids carrying the sequences of 4 promoters that were expected to be selectively expressed in sensory neurons, rat TRPV-1, rat ASIC3, rat CGRP, human CGRP, and EF1α as a control non-selective promoter, was ordered from GenScript Biotech Corps, Piscataway, NJ, USA (Gene ID numbers: rTRPV-1: 83810, rASIC3: 286920, rCGRP: 24241, hCGRP: 796, EF1α: 1915, see Supplementary Materials). All the sequences were engineered to contain a SpeI site at the 5′ end, and PstI (hCGRP, EF1α) or NsiI (TRPV-1, rCGRP, ASIC-3) sites at the 3′ end to facilitate sub-cloning into amplicon plasmids.
- (b)
- Generation of amplicon plasmids. A pUC57 plasmid carrying the sequence of renilla luciferase (RLuc) and fused to a green fluorescent protein (GFP) through an alpha helix (RLuc-GFP, 5′Rluc-αH-GFP3′) was also ordered from GenScript. This sequence, which is flanked by a BamHI site in 5′ and an AauI site in 3′, was sub-cloned into a pGemT-easy plasmid, thus producing plasmid pG-RLuc-GFP. To generate the amplicon vectors used in this study, we modified a previously described amplicon plasmid pA-EUA2-Luc [28]. This plasmid carries 2 independent transcription cassettes: one that expresses GFP driven by an HSV-1 immediate-early promoter (IE4/5), and one that expresses firefly luciferase (Fluc) driven by the immediate early human cytomegalovirus (HCMV) promoter. We first digested pA-EAU2-Luc and pG-RLuc-GFP with BamHI and AauI to delete the GFP open reading frame from pA-EUA2-Luc and replaced this sequence by the RLuc-GFP sequence from pG-RLuc-GFP, thus creating the amplicon plasmid pA-RLuc-GFP. Then, plasmid pA-RLuc-GFP was digested by SpeI and PstI to delete the HCMV promoter, which was replaced by each of the promoters from the pUC57-proms plasmids, thus creating the 5 members of the pA5 family of amplicon plasmids, pA-rTRPV-1-Luc, pA-rASIC3-1-Luc, pA-rCGRP-Luc, pA-hCGRP-Luc, and pA-EF1α -Luc, which express RLuc-GFP driven by the IE4/5 promoter and Fluc driven by each of the different sensory neuron selective promoters (Figure 1).
- (c)
- Production and titration of amplicon vector particles. Amplicon vector stocks were prepared and titrated as previously described [28,29,30]. Cells expressing fluorescent GFP were counted at 24 h post-infection using an inverted fluorescence microscope (Olympus, Tokyo, Japan) with transducing units (TU). Titers of helper virus particles were provided as plaque-forming units (PFU) per milliliter [29,30]. Serial passages of the vector populations were then carried out on 7b cells, thus increasing the TU/PFU ratio to 99/1 or higher.
4.3. Infection and Harvesting
4.4. Western Blots (WB)
4.5. Luciferase Quantification
4.6. Statistics
4.7. Ethics
5. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fehlings, M.G.; Singh, A.; Tetreault, L.; Kalsi-Ryan, S.; Nouri, A. Global prevalence and incidence of traumatic spinal cord injury. Clin. Epidemiol. 2014, 6, 309–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.D. Targeting Recovery: Priorities of the Spinal Cord-Injured Population. J. Neurotrauma 2004, 21, 1371–1383. [Google Scholar] [CrossRef] [PubMed]
- Ruffion, A.; Castro-Diaz, D.; Patel, H.; Khalaf, K.; Onyenwenyi, A.; Globe, D.; LeReun, C.; Teneishvili, M.; Edwards, M. Systematic Review of the Epidemiology of Urinary Incontinence and Detrusor Overactivity among Patients with Neurogenic Overactive Bladder. Neuroepidemiology 2013, 41, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, A.; Lucas, M. Chapter 4: Guidelines for the diagnosis and treatment of overactive bladder (OAB) and neurogenic detrusor overactivity (NDO). Neurourol. Urodyn. 2014, 33 (Suppl. S3), S21–S25. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, D.; Gousse, A.; Keppenne, V.; Sievert, K.-D.; Thompson, C.; Lam, W.; Brin, M.F.; Jenkins, B.; Haag-Molkenteller, C. Phase 3 Efficacy and Tolerability Study of OnabotulinumtoxinA for Urinary Incontinence from Neurogenic Detrusor Overactivity. J. Urol. 2012, 187, 2131–2139. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.; Peyronnet, B.; Aublé, A.; Hascoet, J.; Castel-Lacanal, E.; Miget, G.; Le Doze, S.; Prudhomme, T.; Manunta, A.; Cornu, J.-N.; et al. Long-Term Discontinuation of Botulinum Toxin a Intradetrusor Injections for Neurogenic Detrusor Overactivity: A Multicenter Study. J. Urol. 2018, 201, 769–776. [Google Scholar] [CrossRef] [PubMed]
- de Groat, W.C.; Griffiths, D.; Yoshimura, N. Neural Control of the Lower Urinary Tract. Compr. Physiol. 2014, 5, 327–396. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Chew, D.J.; Biers, S.; Thiruchelvam, N. Electrical nerve stimulation to promote micturition in spinal cord injury patients: A review of current attempts. Neurourol. Urodyn. 2015, 35, 365–370. [Google Scholar] [CrossRef]
- Fink, D.J.; Wechuck, J.; Mata, M.; Glorioso, J.C.; Goss, J.; Krisky, D.; Wolfe, D. Gene therapy for pain: Results of a phase I clinical trial. Ann. Neurol. 2011, 70, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, N.; Doyal, M.F.; Goins, W.F.; Kadekawa, K.; Wada, N.; Kanai, A.J.; de Groat, W.C.; Hirayama, A.; Uemura, H.; Glorioso, J.C.; et al. Morphological changes in different populations of bladder afferent neurons detected by herpes simplex virus (HSV) vectors with cell-type-specific promoters in mice with spinal cord injury. Neuroscience 2017, 364, 190–201. [Google Scholar] [CrossRef] [PubMed]
- de Groat, W.C.; Yoshimura, N. Afferent Nerve Regulation of Bladder Function in Health and Disease. Sens. Nerves 2009, 194, 91–138. [Google Scholar] [CrossRef] [Green Version]
- Corrow, K.; Girard, B.M.; Vizzard, M.A. Expression and response of acid-sensing ion channels in urinary bladder to cyclophosphamide-induced cystitis. Am. J. Physiol. Physiol. 2010, 298, F1130–F1139. [Google Scholar] [CrossRef] [Green Version]
- Kokubun, S.; Sato, T.; Ogawa, C.; Kudo, K.; Goto, K.; Fujii, Y.; Shimizu, Y.; Ichikawa, H. Distribution of TRPV1 and TRPV2 in the human stellate ganglion and spinal cord. Neurosci. Lett. 2015, 590, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Ragionieri, L.; Botti, M.; Gazza, F.; Sorteni, C.; Chiocchetti, R.; Clavenzani, P.; Minelli, L.B.; Panu, R. Localization of peripheral autonomic neurons innervating the boar urinary bladder trigone and neurochemical features of the sympathetic component. Eur. J. Histochem. 2013, 57, e16. [Google Scholar] [CrossRef]
- Chen, C.-C.; England, S.; Akopian, A.N.; Wood, J.N. A sensory neuron-specific, proton-gated ion channel. Proc. Natl. Acad. Sci. USA 1998, 95, 10240–10245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with aδ/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef]
- Xing, J.; Lu, J.; Li, J. Acid-sensing ion channel subtype 3 function and immunolabelling increases in skeletal muscle sensory neurons following femoral artery occlusion. J. Physiol. 2012, 590, 1261–1272. [Google Scholar] [CrossRef] [Green Version]
- Mamet, J.; Lazdunski, M.; Voilley, N. How Nerve Growth Factor Drives Physiological and Inflammatory Expressions of Acid-sensing Ion Channel 3 in Sensory Neurons. J. Biol. Chem. 2003, 278, 48907–48913. [Google Scholar] [CrossRef] [Green Version]
- Smet, P.J.; Edyvane, K.A.; Jonavicius, J.; Marshall, V.R. Neuropeptides and neurotransmitter-synthesizing enzymes in intrinsic neurons of the human urinary bladder. J. Neurocytol. 1996, 25, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Furuta, Y.; Takasu, T.; Fukuda, S.; Inuyama, Y.; Sato, K.C.; Nagazshima, K. Latent Herpes Simplex Virus Type 1 in Human Vestibular Ganglia. Acta Oto-Laryngol. 1993, 113, 85–89. [Google Scholar] [CrossRef]
- Lee, S.; Ives, A.M.; Bertke, A.S. Herpes Simplex Virus 1 Reactivates from Autonomic Ciliary Ganglia Independently from Sensory Trigeminal Ganglia to Cause Recurrent Ocular Disease. J. Virol. 2015, 89, 8383–8391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKie, E.; Graham, D.; Brown, S. Selective astrocytic transgene expression in vitro and in vivo from the GFAP promoter in a HSV RL1 null mutant vector—Potential glioblastoma targeting. Gene Ther. 1998, 5, 440–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foka, P.; Pourchet, A.; Hernandez-Alcoceba, R.; Doumba, P.P.; Pissas, G.; Kouvatsis, V.; Dalagiorgou, G.; Kazazi, D.; Marconi, P.; Foschini, M.; et al. Novel tumour-specific promoters for transcriptional targeting of hepatocellular carcinoma by herpes simplex virus vectors. J. Gene Med. 2010, 12, 956–967. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, J.J.L.; Folkers, E.; Vreeswijk, J. Detection of varicella-zoster virus and herpes simplex virus by the polymerase chain reaction with degenerate primers. J. Virol. Methods 1999, 83, 155–167. [Google Scholar] [CrossRef]
- Smith, A.J.; Clutton, R.E.; Lilley, E.; Hansen, K.E.A.; Brattelid, T. PREPARE: Guidelines for planning animal research and testing. Lab. Anim. 2017, 52, 135–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joussain, C.; Le Coz, O.; Pichugin, A.; Marconi, P.; Lim, F.; Sicurella, M.; Salonia, A.; Montorsi, F.; Wandosell, F.; Foster, K.; et al. Botulinum Neurotoxin Light Chains Expressed by Defective Herpes Simplex Virus Type-1 Vectors Cleave SNARE Proteins and Inhibit CGRP Release in Rat Sensory Neurons. Toxins 2019, 11, E123. [Google Scholar] [CrossRef] [Green Version]
- Behr-Roussel, D.; Oger, S.; Pignol, B.; Pham, E.; Le Maux, A.; Chabrier, P.-E.; Caisey, S.; Compagnie, S.; Picaut, P.; Bernabé, J.; et al. Minimal Effective dose of Dysport and Botox in a Rat Model of Neurogenic Detrusor Overactivity. Eur. Urol. 2012, 61, 1054–1061. [Google Scholar] [CrossRef]
- Lin, G.; Shindel, A.W.; Fandel, T.M.; Bella, A.J.; Lin, C.-S.; Lue, T.F. Neurotrophic effects of brain-derived neurotrophic factor and vascular endothelial growth factor in major pelvic ganglia of young and aged rats. Br. J. Urol. 2009, 105, 114–120. [Google Scholar] [CrossRef] [Green Version]
- Zaupa, C.; Revol-Guyot, V.; Epstein, A.L. Improved Packaging System for Generation of High-Level Noncytotoxic HSV-1 Amplicon Vectors Using Cre-loxP Site-Specific Recombination to Delete the Packaging Signals of Defective Helper Genomes. Hum. Gene Ther. 2003, 14, 1049–1063. [Google Scholar] [CrossRef]
- Krisky, D.; Wolfe, D.; Goins, W.; Marconi, P.; Ramakrishnan, R.; Mata, M.; Rouse, R.; Fink, D.; Glorioso, J. Deletion of multiple immediate–Early genes from herpes simplex virus reduces cytotoxicity and permits long-term gene expression in neurons. Gene Ther. 1998, 5, 1593–1603. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibodies | Brand | Reference | Species | Dilution |
|---|---|---|---|---|
| GAPDH | Millipore | CB1001 | Mouse | 3/4000 |
| NeuN | Abcam | ab177487 | Rabbit | 1/400 |
| Secondary Antibodies | Brand | Reference | Species | |
| Mouse (800) | Li-Cor | 926-32210 | Goat | 3/4000 |
| Rabbit (680) | Li-Cor | 926-68071 | Goat | 3/4000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joussain, C.; Le Coz, O.; Pichugin, A.; Marconi, P.; Lim, F.; Sicurella, M.; Foster, K.; Giuliano, F.; Epstein, A.L.; Aranda Muñoz, A. Development and Assessment of Herpes Simplex Virus Type 1 (HSV-1) Amplicon Vectors with Sensory Neuron-Selective Promoters. Int. J. Mol. Sci. 2022, 23, 8474. https://doi.org/10.3390/ijms23158474
Joussain C, Le Coz O, Pichugin A, Marconi P, Lim F, Sicurella M, Foster K, Giuliano F, Epstein AL, Aranda Muñoz A. Development and Assessment of Herpes Simplex Virus Type 1 (HSV-1) Amplicon Vectors with Sensory Neuron-Selective Promoters. International Journal of Molecular Sciences. 2022; 23(15):8474. https://doi.org/10.3390/ijms23158474
Chicago/Turabian StyleJoussain, Charles, Olivier Le Coz, Andrey Pichugin, Peggy Marconi, Filip Lim, Mariaconcetta Sicurella, Keith Foster, François Giuliano, Alberto L. Epstein, and Alejandro Aranda Muñoz. 2022. "Development and Assessment of Herpes Simplex Virus Type 1 (HSV-1) Amplicon Vectors with Sensory Neuron-Selective Promoters" International Journal of Molecular Sciences 23, no. 15: 8474. https://doi.org/10.3390/ijms23158474