
Maternal High-Fat Diet and Offspring Hypertension



1
Department of Pediatrics, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
2
College of Medicine, Chang Gung University, Taoyuan 333, Taiwan
3
Department of Pharmacy, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan
4
School of Pharmacy, Kaohsiung Medical University, Kaohsiung 807, Taiwan
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(15), 8179; https://doi.org/10.3390/ijms23158179
Submission received: 27 June 2022
/
Revised: 15 July 2022
/
Accepted: 22 July 2022
/
Published: 25 July 2022
(This article belongs to the Special Issue Effect of Maternal High-Fat Diet on Offspring’s Health)
Abstract
:The incidence of hypertension has increased to epidemic levels in the past decades. Increasing evidence reveals that maternal dietary habits play a crucial role in the development of hypertension in adult offspring. In humans, increased fat consumption has been considered responsible for obesity and associated diseases. Maternal diets rich in saturated fats have been widely employed in animal models to study various adverse offspring outcomes. In this review, we discussed current evidence linking maternal high-fat diet to offspring hypertension. We also provided an in-depth overview of the potential mechanisms underlying hypertension of developmental origins that are programmed by maternal high-fat intake from animal studies. Furthermore, this review also presented an overview of how reprogramming interventions can prevent maternal high-fat-diet-induced hypertension in adult offspring. Overall, recent advances in understanding mechanisms behind programming and reprogramming of maternal high-fat diet on hypertension of developmental origins might provide the answers to curtail this epidemic. Still, more research is needed to translate research findings into practice.
1. Introduction
The growing occurrence of cardiometabolic disease is a worldwide health problem that influences all age groups, including women of childbearing age [1,2,3]. Today, a broad spectrum of factors contributes to cardiometabolic disease [3], including genetic susceptibility, intra-uterine growth retardation, imbalanced diet, socioeconomic status, physical activity, etc. Among them, imbalanced diet is a key modifiable factor that can be targeted to control cardiometabolic disease [4]. Recently, the increased consumption of fats, especially saturated fats, has raised interest in finding how high-fat diet increases susceptibility to cardiometabolic disease. A high-fat diet (HFD) is more reflective of the dietary habits in western society. Dietary patterns with high-fat consumption established in early childhood tend to continue during pregnancy.
A mounting body of epidemiological and experimental evidence supports a role for early-life environmental exposures in determining the long-term health of an individual, which is referred as the Developmental Origins of Health and Disease (DOHaD) theory [5]. Fetal development is dependent on maternal nutrition. Under- or over-nutrition has adverse effect on offspring [6]. A maternal HFD can alter the morphology and function of various tissues/organs and thus the offspring become more susceptible to many diseases later in life [7,8].
It is challenging to study maternal dietary patterns in human participants due to difficulty in exactly calculating nutrient content of food intake, and ethical issues related to manipulating the dietary patterns of pregnant women. Animal models provide an invaluable tool to control over diet composition, eliminate confounding factors, and explore the underlying mechanisms of developmental programming [9]. Animal models of maternal HFD demonstrate that adverse outcomes may occur in the offspring, such as obesity, insulin resistance, behavior disorders, endothelial dysfunction, hypertension, hepatic steatosis, dyslipidemia, increased visceral fat mass, glucose intolerance, and adipocyte hypertrophy, etc. [7,8,9,10,11].
Hypertension has a central role in cardiometabolic disease [3,4]. Although blood pressure (BP) shows multifactorial inheritance patterns, known genetic variants only explain ~3.5% of BP trait variability [12]. Increasing evidence indicates that developmental programming can take place in early life, resulting in hypertension later in life [13,14,15]. A wide array of early-life environmental stimuli can induce developmental programming of hypertension [13,14,15,16]. Among them, maternal nutrition has a role in the pathogenesis of hypertension of developmental origins [15]. Maternal HFD is considered as a key early-life trait involving adverse offspring outcomes. Nevertheless, little is known on whether maternal HFD can induce offspring hypertension and the underlying mechanisms.
To examine and identify the evidence around the impact of maternal HFD on offspring hypertension, our search strategy was developed for literature retrieval in the PubMed/MEDLINE databases on maternal HFD, DOHaD, and hypertension. We used the following search terms: “developmental programming”, “DOHaD”, “reprogramming”, “blood pressure”, “mother”, “pregnancy”, “gestation”, “lactation”, “offspring”, “progeny”, “high-fat diet”, and “hypertension”. We further checked fitting reference lists to find additional studies in eligible papers. The last search was conducted on 20 June 2022.
2. Maternal High-Fat Diet Programs Adult Diseases
Relatively little information currently exists concerning the impact of maternal HFD on offspring health in humans. Although some evidence links maternal obesity during pregnancy with an increased risk of obesity in offspring in later life [17,18], maternal obesity does not necessarily correspond to maternal high-fat diet [10]. Most epidemiological studies that pool various participants as well as fat components from different dietary sources bring great risk for diluting any real findings. In contrast, dietary composition can be easily determined for the purpose of comparisons in animal models across species. Hence, much of what is known about the implication of maternal HFD on offspring health is mainly based on animal models. Animal models have been instrumental in indicating the biological plausibility of the relations noticed in epidemiological research and provide proof of causality.
High-fat diets have long been known for generating obesity in animal models and associated diseases [18,19]. Nevertheless, this dietary intervention is not standardized, and the HFD-induced phenotypes vary distinctly among different studies [18,19]. Prior reviews demonstrated that different HF diets with fat fractions ranged from 20% to 60% energy as fat, and the basic constituents of fats vary between plant oils (e.g., corn or coconut oil) and animal-derived fats (e.g., lard or butter) [7,8,19]. Notably, diets rich in saturated fats can lead to health risks, while unsaturated fats are beneficial to heart health [20]. These obviously lead to a considerable variability in the results being reported.
The effect of a maternal HFD on the offspring has been examined in rats [7,8], mice [7,8], rabbits [21], pigs [22], and non-human primates [23]. A systematic review recruiting 17 animal studies demonstrated that maternal HFD may compromise parameters in feeding behavior and body composition of offspring [24]. Another systemic review including 11 studies identified the risk of type 2 diabetes and obesity in male offspring exposed to a maternal HFD [25].
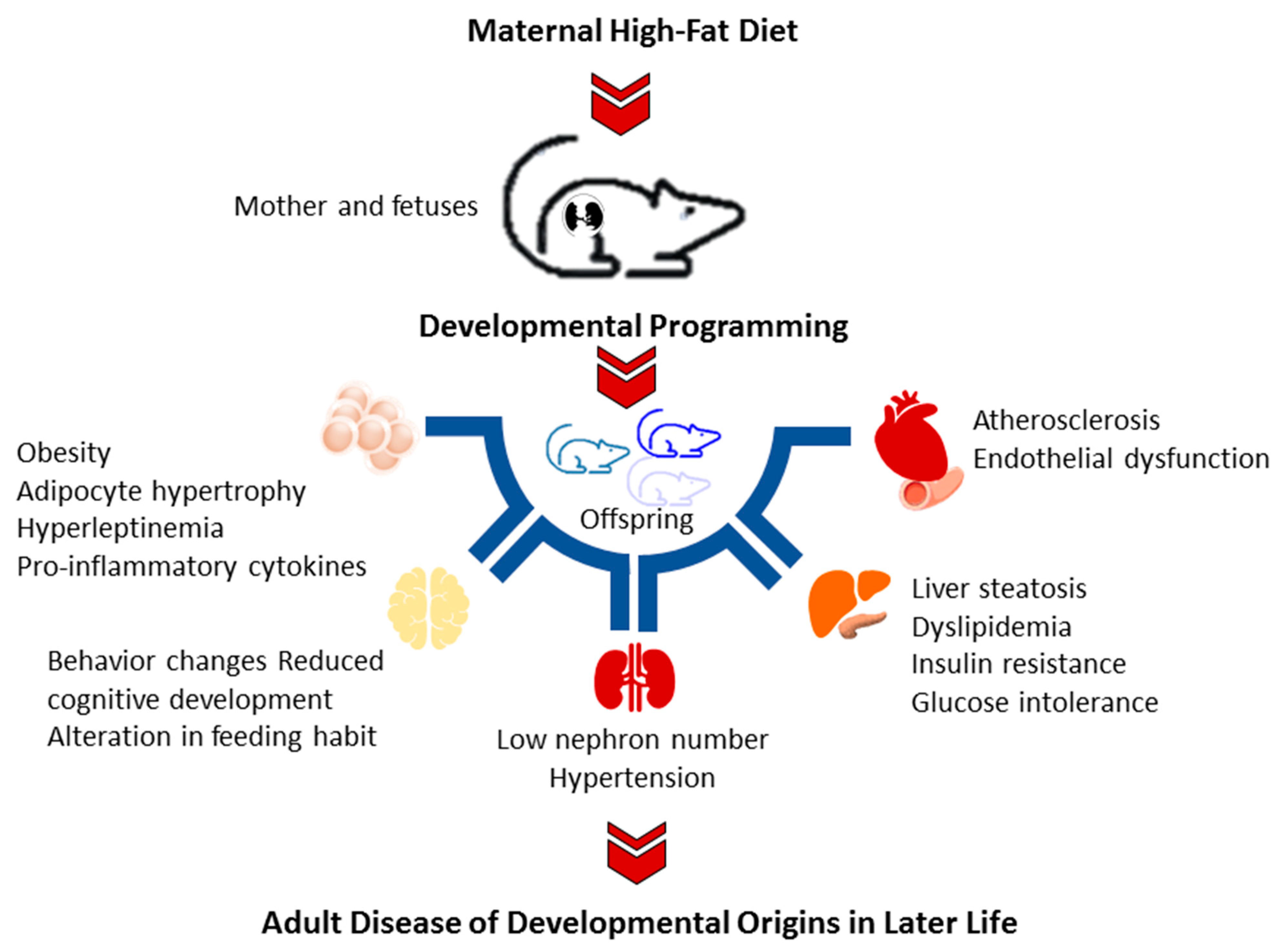
Current evidence has emerged from animal models that offspring exposed to a maternal HFD manifest various components of metabolic syndrome [26,27], including obesity [26], insulin resistance [28], liver steatosis [29], dyslipidemia [29], and hypertension [30]. Additionally, maternal HFD may modify the development of the brain, resulting in reduced cognitive development, increased depressive-like and aggressive behaviors, and alteration in feeding habits in the offspring [8]. A brief summary of adverse offspring outcomes consequent of maternal HFD is depicted in Figure 1.
Although a variety of adverse offspring outcomes related to maternal HFD have been reported, at present, our understanding of how maternal HFD induces offspring hypertension, the mechanisms behind developmental programming, and efficient reprogramming strategies remain largely unknown.
3. High-Fat Diet and Hypertension
Regulation of blood pressure (BP) is a complex integrated response involving a variety of organ systems, including the heart, blood vessels, brain, and kidneys [28]. Besides, the maintenance of normal BP needs the interconnection and coordination of several regulatory mechanisms involving the nitric oxide (NO), renin–angiotensin system (RAS), the sympathetic nervous system, and sodium excretion [31].
3.1. Cardiovascular System
Observational studies in humans linked high saturated fat consumption with atherosclerosis and coronary artery disease, while monounsaturated fat consumption is association with reduction of cardiovascular mortality [32]. Endothelial cells are important constituents of blood vessels that determine cardiovascular homeostasis [33]. Dysfunction of endothelium has been characterized by a shift from executing physiologic functions of endothelium to a pro-inflammatory setting, a pro-thrombotic state, and vasoconstriction. All these together implicate in the pathogenesis of hypertension. Notably, these events can be induced by oxidative stress in the presence of high-fat intake [34]. The vasculature is a major source of NADPH oxidase-derived ROS [35]. High-fat diet causes endothelial dysfunction and vascular oxidative stress is related to increases of NADPH oxidase-derived ROS [36]. Additionally, several endothelium-derived vasoconstrictors, such as angiotensin II (Ang II) [37], urotensin II [38], and vasoconstrictor prostaglandins [39], can be released by endothelium in response to a high-fat diet.
On the other hand, reduced bioavailability of NO, a well-known vasodilator, is considered a hallmark of endothelial dysfunction [40]. The endothelial NO synthase (NOS)-derived NO is responsible for vasodilation in the cardiovascular system. Prior work indicated that high-fat-diet-induced endothelial dysfunction accompanying by reduced eNOS-derived NO [36]. High-fat diet can also increase asymmetric dimethylarginine (ADMA), an endogenous NOS inhibitor [41]. ADMA can uncouple NOS isoenzymes to form superoxide, contributing to endothelial dysfunction [42]. Together, high-fat diet disturbs vascular tone via regulating vasodilators and vasoconstrictors and causes vascular dysfunction, arterial stiffness, atherosclerosis, and vascular remodeling as well [43], all of which contribute to the development of hypertension.
3.2. Central Nervous System
The central nervous system (CNS) organizes regional sympathetic outflow to target organs (e.g., the kidneys and heart) through the integration of autonomic brainstem networks, reflex influences, and input from circulating factors [44]. Overexcitation of the sympathetic nervous system has a crucial role in the pathogenesis of hypertension [44]. Similar to the cardiovascular system, oxidative stress in the CNS is involved in the development of hypertension [45]. ROS increase sympathoexcitatory inputs to rostral ventrolateral medulla (RVLM) neurons, while iNOS-mediated NO production stimulates sympathoinhibition [46]. As a result, an imbalance of ROS and NO in the RVLM increases sympathetic tone, resulting in hypertension [45,46]. Conversely, interventions that reduce brain oxidative stress have been reported to prevent neurogenic hypertension [47].
3.3. Renal System
Several lines of evidence clearly indicate that the kidneys contribute to HFD-induced hypertension. The first are data from spontaneously hypertensive rat (SHR), a commonly used hypertension rat model. High-fat diet causes hypertension and coincides with increased intrarenal lipid concentrations, oxidative stress, renal inflammation, and activation of renal RAS [51]. Secondly, dysregulated sodium transport in the kidneys leads to hypertension [52]. High-fat diets have been found to induce hypertension accompanying by impairing several sodium transporters in the kidneys, like Na+/Cl− cotransporter (NCC), sodium hydrogen exchanger type 3 (NHE3), and Na-K-2Cl cotransporter (NKCC2) [30,53,54]. Another line of evidence comes from the activation of the renal RAS in high-fat-diet-induced hypertension and kidney injury [30,55,56]. It is well known that the kidney is a principal target for the various components of the RAS that are implicated in hypertension and kidney disease [57]. Accordingly, it is suggested that the intrarenal RAS activation plays an important role in hypertension induced by high-fat diet.
Although these organ systems have shown their potential roles in high-fat-diet-induced hypertension, little is known regarding programming effects of maternal HFD in the brain, heart, kidneys, and vessels in hypertension of developmental origins.
4. Hypertension of Developmental Origins: The Impact of Maternal High-Fat Diet
4.1. Animal Models of Maternal HFD-Induced Offspring Hypertension
A growing number of animal models have been generated to study hypertension of developmental origins, as reviewed elsewhere [14,58]. Imbalanced maternal nutrition can program the fetus resulting in hypertension in later life. Table 1 summarizes animal studies documenting offspring hypertension in animal models of high-fat diet, restricting the exposure to critical periods during early development [21,59,60,61,62,63,64,65,66,67,68,69,70,71]. In this review, we only considered studies reporting offspring outcomes starting from childhood.
Rats are the most frequently used animals. Other species such as rabbits [21] and mice [66] have also been used to study hypertension of developmental origins programmed by maternal HFD. In view of the fact that each month of the adult life of a rat corresponds to 3 human years [72], Table 1 shows the timing of developing hypertension in rats ranging from 12 weeks to 1 year of age, which corresponds to humans from childhood to adulthood. In rodents, different maternal HFDs with fat fractions ranged from 18.7% to 58% energy as fat, which were close to previous studies [7,8]. Notably, maternal HFD-induced responses of offspring BP could be increased [21,59,60,61,62,63,64,65,66,67,68,69,70,71] but also unaltered [30,61], mainly depending on sex, age, species, and varied fatty acid compositions. In Table 1, most BP data were obtained from the tail cuff method, except some studies using direct arterial catheter [21] or telemetry method [61,62]. Though BP data detected from the tail cuff method have been reported to correlate well with findings of direct arterial catheter and telemetry methods [73], part of the increased BP in offspring may be related to an increase in sympathetic nerve activity [74].
Additionally, maternal HFD-induced offspring hypertension is associated with developmental programming in several organs, including the kidneys [63,64,67,68,69,70,71], vessels [60,61,62,65,66], and brain [21]. Notably, offspring hypertension does not essentially correspond to obesity in response to maternal HFD. Offspring obesity may or may not appear in these maternal HFD-induced hypertension models. What is absent in the literature is animal models used for studying maternal HFD-induced hypertension target organ damage. Although maternal HFD induced programming effects in various organs, the contributions of organ-specific programming on damage in target organs have not been yet extensively studied in the above-mentioned animal models.
Of note is that the development of hypertension may follow the two-hit model. Recent evidence suggests a “two-hit” hypothesis that illuminates the developmental programming of adult diseases [75]. Hypertension can develop with two sequential hits, the first hit being the response to a prenatal insult, followed by the second hit in response to ongoing programming induced by the first hit. Since the first hit alone may not be sufficient to alter the adult phenotype, another insult may act as a second hit to amplify the underlying defects culminating in a disease state. A number of two-hit models, hence, have been used to evaluate whether two distinct hits affect offspring outcomes synergistically or differently when combined as compared with either hit alone.
In some studies, maternal HFD was applied as the first hit. Maternal HFD causes morphological functional changes of fetal organs, which alone might not be sufficient to alter the adult phenotype. The second hit could amplify ongoing programming processes triggered by maternal HFD, culminating in a disease state. So far, some two-hit models have been used to evaluate whether two distinct hits affect offspring hypertension synergistically or differently when combined as compared with maternal HFD alone. For example, models of a maternal plus post-weaning HFD [68,71] and combined maternal HFD and bisphenol A exposure [69] have been established to study hypertension of developmental origins. To sum up, animal models with various maternal HFDs during different fetal developmental stages generate the same outcome―hypertension in adult life. These findings suggest there might be major basic mechanisms behind hypertension of developmental origins.
4.2. Proposed Mechanisms Underlying Maternal HFD-Induced Offspring Hypertension
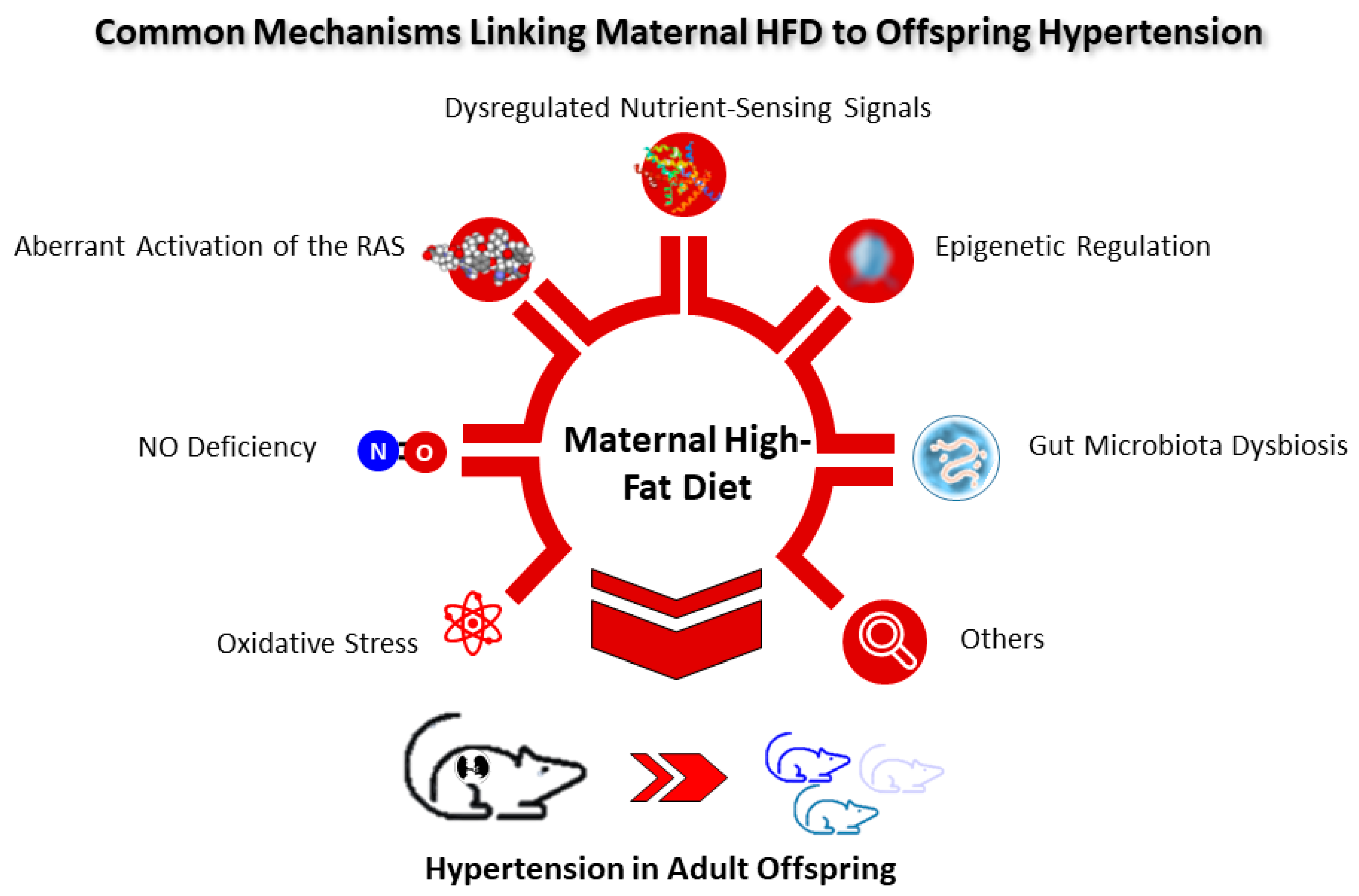
To date, several main mechanisms underlying hypertension of developmental origins have been proposed, such as oxidative stress, NO deficiency, aberrant activation of the RAS, dysregulated nutrient-sensing signals, epigenetic regulation, gut microbiota dysbiosis, etc. [14,57,58,76,77,78,79]. Among them, some are interconnected with maternal HFD and will be discussed in turn (Figure 2).
4.2.1. Oxidative Stress
Oxidative stress, a disturbance in pro-oxidant/antioxidant balance, has been implicated in the pathophysiology of compromised pregnancy and adverse fetal outcomes [80]. During fetal development, the existence of excessive ROS under adverse intrauterine conditions prevails over the antioxidant defense system and causes fetal programming, leading to oxidative stress-related hypertension in adult offspring [81]. Prior reviews revealed that there are a number of maternal insult stimuli connected to oxidative stress in mediating offspring [14,81].
Several mechanisms accompanying by oxidative stress behind hypertension of developmental origins have been reported, including upregulation of ROS-producing enzymes [82], excessive ROS generation [69,83], increases in lipid peroxidation [84], elevated oxidative DNA damage [85], increased peroxynitrite formation [86], and decreased antioxidant capacity [63].
Table 1 shows that increased oxidative stress in the vessels [60,66] and kidneys [63,64,68,69] is associated with maternal HFD-induced hypertension in adult offspring. A commonly used marker of lipid peroxidation, malondialdehyde (MDA), has been used to detect oxidative damage and shown increased in the offspring kidneys in maternal HFD models [63,64]. Additionally, 8-hydroxydeoxyguanosine (8-OHdG), an oxidized nucleoside of DNA, is the most frequently detected DNA lesion [87], whose staining was significantly increased in the kidneys of adult rat offspring born to dams received HFD [68,69]. These findings provide the connections between maternal HFD and oxidative stress that underlie the hypertension of developmental origins.
4.2.2. NO Deficiency
NO depletion in pregnancy can cause fetal programming, leading to programmed hypertension in adult offspring [76,88,89]. Prior work revealed that maternal NO deficiency alters a wide range of signaling pathways using the transcriptomic analysis [77]. Among them, several redox-sensitive signaling pathways contribute to the development of hypertension [90]. Additionally, NO deficiency in embryonic kidneys induced by ADMA can impair nephrogenesis and alter several genes related to hypertension of developmental origins [91]. Conversely, early interventions targeting the NO pathway could be used for reprogramming hypertension in different models of developmental programming [76,89].
As shown in Table 1, reduced NO in the vessels [60,65,66] and kidneys [63,68,69] is related to maternal HFD-induced offspring hypertension. Former work reveals that ADMA-related ROS–NO imbalance in early life causes hypertension in adult life [76]. In a combined maternal HFD and bisphenol A exposure [69], adult offspring develop hypertension coinciding with increased ADMA. However, maternal HFD alone increased offspring BP but has a negligible effect on ADMA level. A previous study reported that atorvastatin can reduce ADMA by increasing its metabolism to protect adult rats against high-fat diet-induced endothelial dysfunction [41]. As many currently used drugs have ADMA-lowering properties that can restore ADMA-NO balance [92], a deeper understanding of the reprogramming effects of NO-targeted intervention in HFD-induced programmed hypertension is warranted.
4.2.3. Aberrant Activation of the RAS
The RAS, a major regulatory network of BP, is tightly connected with hypertension of developmental origins [57]. The RAS consists of several angiotensin (Ang) peptides that regulate BP through distinct receptors [93]. The classic RAS can be defined as the angiotensin-converting enzyme (ACE)/angiotensin (ANG) II/angiotensin II type 1 receptor (AT1R) axis. Activation of the classic RAS elicits vasoconstriction, oxidative stress, and inflammation, resulting in hypertension [94]. Maternal HFD-induced offspring hypertension is related to the aberrant activation of the classic RAS, represented by increases in increased renal mRNA expression of Agt and Ace and protein level of AT1R [67].
Conversely, the non-classic RAS, composed by the ACE2/Ang-(1–7)/Mas receptor, can counterbalance the adverse effects of ANG II [93]. Another study reported that renal Ang-(1–7) level was decreased in 16-week-old male offspring born to dams that received HDF [68]. Prior research indicated there is a transient biphasic response with downregulation of classic RAS axis in neonatal stage that becomes normalized with age [94]. Maternal HFD may disturb this normalization; hence, the classic RAS axis is aberrant activation, resulting in the rising BP in adult offspring.
Given the fact that only few studies addressed the impact of RAS in maternal HFD-induced hypertension, there is need for further investigation of this research gap. Considering maternal HFD increased offspring’s BP coinciding with aberrant activation of the classic RAS, more work is required to explore whether blockade of the RAS can be used as reprogramming interventions for maternal HFD-induced programmed hypertension.
4.2.4. Dysregulated Nutrient-Sensing Signals
During gestation, nutrient-sensing signals regulate fetal metabolism in response to maternal nutritional status [95]. Different signaling pathways that detect intracellular and extracellular levels of specific nutrients, such as fats, are coordinated at the organismal level via hormonal signals [96]. Accordingly, maternal nutritional imbalance resulting in dysregulation of nutrient-sensing signals cause a crucial impact on hypertension of developmental origins [14,93].
Maternal HFD-induced offspring hypertension is correlated to inhibitory AMP-activated protein kinase (AMPK)/peroxisome proliferator-activated receptor-γ (PPARγ) coactivator-1α (PGC-1α) pathway in offspring kidneys [68]. AMPK can phosphorylate PGC-1α. PGC-1α binds to PPARγ and coactivates PPARγ to facilitate the expression of specific sets of PPAR target genes participating in hypertension [97]. A growing body of evidence indicates that downregulation of nutrient-sensing signals, such as AMPK and PGC-1α, is related to hypertension of developmental origins, while AMPK activation can serve as a reprogramming strategy to protect offspring against adverse programmed processes [98]. Although a potential link between nutrient-sensing signals and maternal HFD underlying hypertension of developmental origins exists, whether these signals impact maternal HFD-induced adverse offspring outcomes in an organ-specific manner remains unclear.
4.2.5. Epigenetic Regulation
During pregnancy, epigenetic mechanisms are involved in programming gene expression for fetal development [99]. Recent studies suggest that DNA methylation, histone modification, and noncoding RNAs may be one of the epigenetic mechanisms that program the effects of early-life habits on later-life risk of adult diseases, including hypertension [78,100].
Using next-generation RNA sequencing (NGS) analysis, our prior work reported maternal HFD significantly altered renal transcriptome in 1-week-old female offspring [30]. In total, 251 differential expressed genes (DEGs) (154 up- and 97 downregulated genes) were identified. Among them, several genes were related to regulation of BP, such as Agtr1b and Ace belonging to the RAS, Ddah1 in the NO signaling pathway, and sodium transporter Slc12a3. Notably, a maternal HFD also induces differential alterations of gene expression in the placenta [100], brain [101], and heart [102] in offspring. These data demonstrate that epigenetic regulation may participate the developmental programming of adult diseases in an organ-specific manner.
As well, maternal HFD leads to offspring hypertension and was relevant to increased leptin promoter hypomethylation and leptin expression in adipose tissues of HFD-exposed rat offspring [59]. Similarly, another study demonstrated that maternal HFD may program sympatho-excitatory capacity to induce hypertension in adult rabbit offspring attributed to increased leptin receptor [58]. Hypothalamic leptin signaling can activate specific melanocortin receptors located on sympathetic neurons and consequently result in sympathetic activation of the heart and kidneys and, finally, elevated BP [103]. Thus, maternal HFD can regulated certain genes involved in the regulation of BP. However, the underlying epigenetic mechanisms await further clarification.
4.2.6. Gut Microbiota Dysbiosis
Within the gut reside various microbes coexisting with the host in a mutually beneficial relationship, and thus gut microbiota has potential to affect human health and disease [104]. During pregnancy and lactation, the mothers share gut microbiota and derived metabolites with their offspring, which illuminate the impact of maternal influences in the development of offspring’s gut microbiota [105].
A meta-analysis including 15 studies indicated that intake of high saturated fat may negatively affect microbiota richness and diversity [106]. Maternal HFD was reported to reduce α-diversity in offspring’ microbiota [107]. Loss of α-diversity appears as the most constant finding of gut microbiota dysbiosis, leading to many human diseases [108]. Additionally, a maternal HFD programs offspring’s hypertension coincides with an increased Firmicutes to Bacteroidetes (F/B) ratio. This was tied well with hypertension models showing the F/B ratio was increased and served as a microbial marker of hypertension [109]. Additionally, reduction of beneficial microbes was also a hallmark of gut microbiota dysbiosis [104]. The abundance of both beneficial bacterial strains Lactobacillus and Akkermansia [110,111], were reduced in the maternal HFD model [67,71].
Moreover, maternal HFD increased offspring’s BP accompanying by alterations of gut microbiota-derived metabolites [67,71]. Microbial metabolites such as short-chain fatty acids (SCFAs), trimethylamine (TMA), and trimethylamine N-oxide (TMAO) are involved in BP regulation [112,113,114]. Maternal HFD was reported to reduce fecal propionate level in 3-week-old offspring [71]. As propionate has vasodilatory action via mediating SCFA receptors [112], this finding suggests reduced SCFA might be a possible reason contributing to maternal HFD-induced hypertension. Maternal HFD also caused the increases of TMA levels and decreases of TMAO-to-TMA ratio in adult rat offspring [71]. As microbiota-derived metabolites TMA and TMAO are risk factors for cardiovascular disease [113,114], whether HFD-induced programmed hypertension can be prevented by reducing accumulation of TMA and TMAO warrants further investigation.
4.2.7. Others
With regard to the multiple negative aspects of maternal HFD on offspring outcomes, other possible mechanisms might be involved, for example, dysregulation of H2S or sex differences. Hydrogen sulfide (H2S), the third gasotransmitter, has emerged as an important regulator of BP [115]. Increasing evidence supports the use of H2S-based interventions as a reprogramming approach to protect offspring against hypertension of developmental origins [116]. One previous study revealed maternal HFD caused decreases of plasma H2S levels and renal H2S-releasing activity in male rat offspring [70]. These findings suggest that a link between HFD and H2S might be behind hypertension of developmental origins, although this remains speculative. Sex-dependent differences exist in hypertension of developmental origins [117,118]. It has been noted that male offspring are more susceptible to be hypertension than female offspring [117,118]. This difference has led many researchers to work on predominately male animal research, as listed in Table 1. However, one study reported that maternal HFD programs hypertension in female but not male offspring [61]. Another study revealed there is no difference of maternal HFD on the development of hypertension in each sex [30]. More research on sex differences is required as they may become a potential mechanism in hypertension programmed by maternal HFD.
5. Reprogramming Strategies
With a deeper understanding of the mechanisms underlying maternal HFD-programmed hypertension, the development of mechanism-targeted strategies holds potential for reprogramming. So far, early-life interventions to offset mechanisms governing hypertension of developmental origins that have been evaluated range from avoidance of risk factors, nutritional interventions, pharmacological therapies, to lifestyle modification [14,58,81].
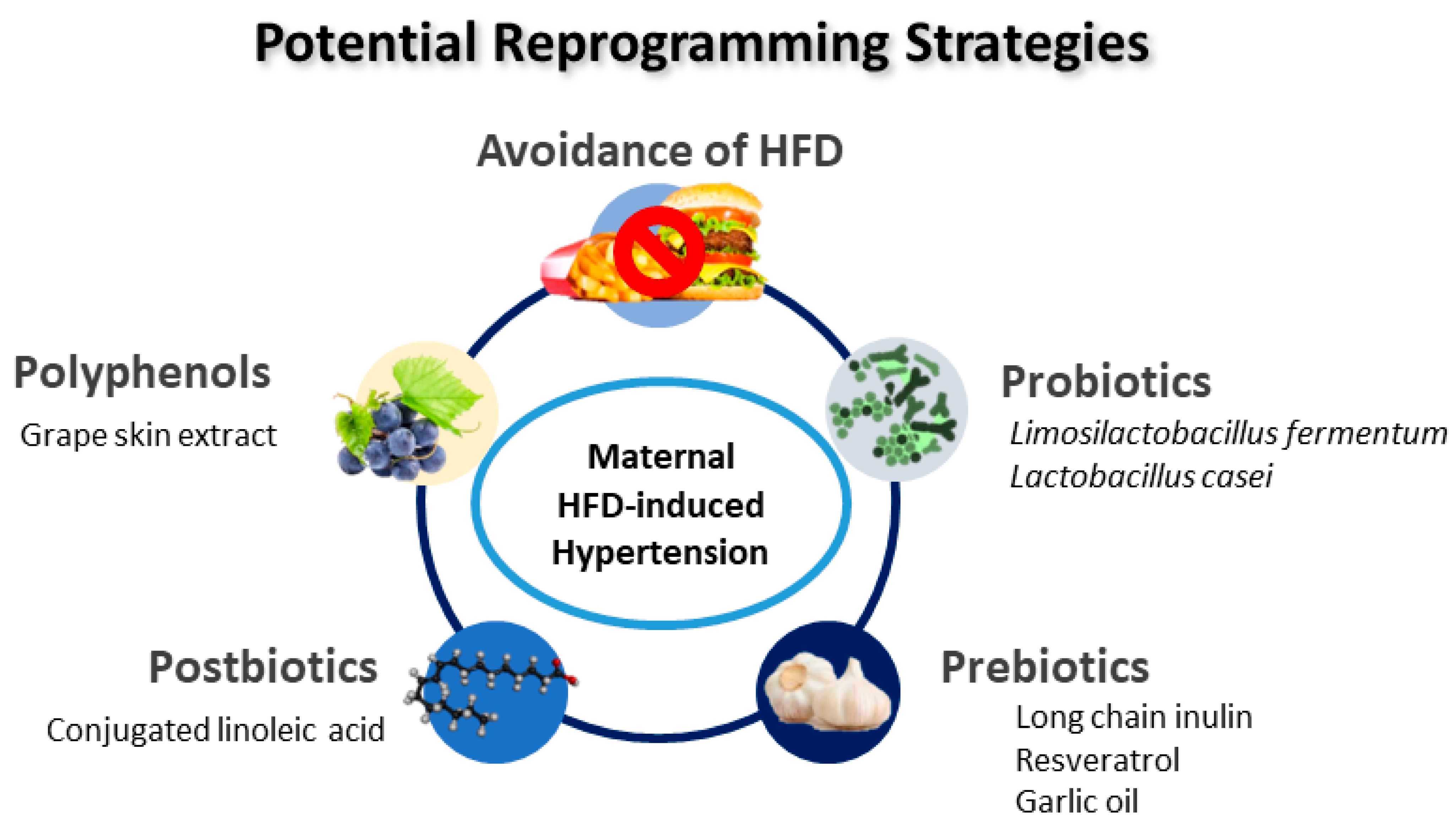
Animal models have been essential in providing potential reprogramming strategies. As described in Table 1, several interventions have been used as reprogramming strategies to prevent hypertension in offspring exposed to maternal HFD, including grape skin extract [60], Limosilactobacillus fermentum [64], conjugated linoleic acid [65], long chain inulin [67], Lactobacillus casei [67], resveratrol [68,69], and garlic oil [70]. A schematic summarizing the potential reprogramming interventions for maternal HFD-induced hypertension of developmental origins is presented in Figure 3.
Interestingly, most reprogramming interventions used in maternal HFD models are targeted on gut microbiota. Probiotics and prebiotics have long been known for their benefits in human health [119,120], including treating hypertension [121], while less attention has been paid to their preventive aspect on hypertension of developmental origins [58]. Probiotic treatment with Limosilactobacillus fermentum [64] or Lactobacillus casei [67] in pregnancy and lactation prevents the development of hypertension in adult offspring exposed to maternal HFD. Additionally, long-chain inulin, a prebiotic, causes a beneficial protective effect against maternal HFD-induced offspring hypertension [67]. Similarly, resveratrol could be used to reprogram maternal HFD-induced hypertension due to its probiotic properties [68,69]. Another study showed that garlic oil, one of the prebiotic foods, protected maternal HFD-induced hypertension in adult rat progeny [70]. Its beneficial effects include increased α-diversity, increased plasma SCFA levels, and increased proportions of beneficial bacteria Bifidobacterium and Lactobacillus.
Apart from probiotics and prebiotics, postbiotics are another gut microbiota-based modality [122]. Postbiotics are bioactive compounds made from metabolic by-products of live probiotic bacteria [122]. Conjugated linoleic acid is a gut microbiota-derived metabolite from dietary polyunsaturated fatty acids. As a postbiotic, maternal conjugated linoleic acid supplementation reversed maternal HFD-induced offspring hypertension [65].
Furthermore, former work reveals that specific nutrient intake can be advantageous for protecting offspring from hypertension of developmental origins in various animal models [123,124]. One of them is polyphenol [124]. One previous study showed that supplementation with grape skin polyphenols during gestation and lactation protects against hypertension induced by a maternal HFD [60].
To summarize, current evidence from animal models supports that early-life interventions such as gut microbiota-targeted therapies may be able to prevent the development of hypertension programmed by maternal HFD in a desired favorable direction.
6. Conclusions and Perspectives
Previous research has shown that maternal HFD causes a variety of adverse offspring outcomes later in life. In this review, we outlined recent advances supporting the influence of maternal HFD on offspring hypertension. Reflecting current knowledge of animal models, our review also shed light on prevention of maternal HFD-induced hypertension via innovative reprogramming strategies. Unfortunately, animal models of maternal HFD are not without limitations. There is no one-size-fits-all model to induce the same adverse offspring outcomes (e.g., hypertension or obesity) in response to maternal HFD. Part of the issue is limited not simply to practical issues regarding study design (compositions of fats, duration, and age at measure) but also to the confounding effects of genetic variation (sex, animal species, and background). Future work could point to improve comparability across studies using “identical” maternal HFD models.
To move the field forward, some unsolved aspects toward clinical translation need to be considered. Although several core mechanisms were addressed in the current review, we may not completely understand the full picture of mechanisms behind maternal HFD-induced offspring hypertension. Considering maternal HFD can program various organs resulting different phenotypes in adult offspring, it is imperative that additional studies simultaneously evaluate organ-specific programming effects in an experiment. What is absent from the literature is a deeper understanding of whether maternal HFD can program offspring hypertension leading to damage of target organs, such as the brain, heart, and kidneys.
Studies to date have found that gut microbiota-targeted therapies can reprogram maternal HFD-induced hypertension. Nevertheless, none of them have been translated in human studies. Despite results from human studies revealing probiotics and prebiotics in the treatment of maternal conditions during pregnancy being beneficial, currently their effectiveness in protecting offspring against hypertension remain largely unclear. Future work in large trials is required to better identify appropriate probiotic species and prebiotics for pregnant women consuming HFD for preventing hypertension in their children.
In summary, the growing body of knowledge suggests maternal dietary habits with excessive fat intake during gestation and lactation should be avoided to minimize offspring risk for developing hypertension in later life. After a greater understanding of maternal HFD-induced offspring hypertension and a remarkable growth achieved in reprogramming strategies, we believe that translating animal results into clinical application is a valuable approach that could curtail the global pandemic of hypertension.
Author Contributions
Conceptualization, Y.-L.T. and C.-N.H.; funding acquisition, Y.-L.T.; project administration, Y.-L.T. and C.-N.H.; data curation, Y.-L.T. and C.-N.H.; writing—original draft, Y.-L.T. and C.-N.H.; writing—review and editing, Y.-L.T. and C.-N.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by grants MOST 110-2314-B-182-020-MY3 (Y.-L.T.) and MOST 110-2314-B-182A-029 (C.-N.H.) from the Ministry of Science and Technology, Taiwan.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Micha, R.; Peñalvo, J.L.; Cudhea, F.; Imamura, F.; Rehm, C.D.; Mozaffarian, D. Association between dietary factors and mortality from heart disease, stroke, and type 2 diabetes in the United States. JAMA 2017, 317, 912. [Google Scholar] [CrossRef] [Green Version]
- Ramlakhan, K.P.; Johnson, M.R.; Roos-Hesselink, J.W. Pregnancy and cardiovascular disease. Nat. Rev. Cardiol. 2020, 17, 718–731. [Google Scholar] [CrossRef] [PubMed]
- Sattar, N.; Gill, J.M.R.; Alazawi, W. Improving prevention strategies for cardiometabolic disease. Nat. Med. 2020, 26, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.; Malik, V.S.; Hu, F.B. Cardiovascular disease prevention by diet modification. J. Am. Coll. Cardiol. 2018, 72, 914–926. [Google Scholar] [CrossRef] [PubMed]
- Haugen, A.C.; Schug, T.T.; Collman, G.; Heindel, J.J. Evolution of DOHaD: The impact of environmental health sciences. J. Dev. Orig. Health Dis. 2014, 6, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Tain, Y.L. The Good, the Bad, and the Ugly of Pregnancy Nutrients and Developmental Programming of Adult Disease. Nutrients 2019, 11, 894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, L.; Seki, Y.; Vuguin, P.M.; Charron, M.J. Animal models of in utero exposure to a high fat diet: A review. Biochim. Biophys. Acta 2014, 1842, 507–519. [Google Scholar] [CrossRef] [Green Version]
- Gawlińska, K.; Gawliński, D.; Filip, M.; Przegaliński, E. Relationship of maternal high-fat diet during pregnancy and lactation to offspring health. Nutr. Rev. 2021, 79, 709–725. [Google Scholar] [CrossRef]
- Litzenburger, T.; Huber, E.K.; Dinger, K.; Wilke, R.; Vohlen, C.; Selle, J.; Kadah, M.; Persigehl, T.; Heneweer, C.; Dötsch, J.; et al. Maternal high-fat diet induces long-term obesity with sex-dependent metabolic programming of adipocyte differentiation, hypertrophy and dysfunction in the offspring. Clin. Sci. 2020, 134, 921–939. [Google Scholar] [CrossRef]
- Ribaroff, G.A.; Wastnedge, E.; Drake, A.J.; Sharpe, R.M.; Chambers, T.J.G. Animal models of maternal high fat diet exposure and effects on metabolism in offspring: A meta-regression analysis. Obes. Rev. 2017, 18, 673–686. [Google Scholar] [CrossRef]
- Seet, E.L.; Yee, J.K.; Jellyman, J.K.; Han, G.; Ross, M.G.; Desai, M. Maternal high-fat-diet programs rat offspring liver fatty acid metabolism. Lipids 2015, 50, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Seidel, E.; Scholl, U.I. Genetic mechanisms of human hypertension and their implications for blood pressure physiology. Physiol. Genom. 2017, 49, 630–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luyckx, V.A.; Bertram, J.F.; Brenner, B.M.; Fall, C.; Hoy, W.E.; Ozanne, S.E.; Vikse, B.E. Effect of fetal and child health on kidney development and long-term risk of hypertension and kidney disease. Lancet 2013, 382, 273–283. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Animal Models for DOHaD Research: Focus on Hypertension of Developmental Origins. Biomedicines 2021, 9, 623. [Google Scholar]
- Bagby, S.P. Maternal nutrition, low nephron number, and hypertension in later life: Pathways of nutritional programming. J. Nutr. 2007, 137, 1066–1072. [Google Scholar] [PubMed] [Green Version]
- Laitinen, J.; Power, C.; Järvelin, M.R. Family social class, maternal body mass index, childhood body mass index, and age at menarche as predictors of adult obesity. Am. J. Clin. Nutr. 2001, 74, 287–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawlor, D.A. The Society for Social Medicine John Pemberton Lecture 2011. Developmental overnutrition—An old hypothesis with new importance? Int. J. Epidemiol. 2013, 42, 7–29. [Google Scholar] [CrossRef]
- Buettner, R.; Schölmerich, J.; Bollheimer, L.C. High-fat diets: Modeling the metabolic disorders of human obesity in rodents. Obesity 2007, 15, 798–808. [Google Scholar] [CrossRef]
- Buettner, R.; Parhofer, K.G.; Woenckhaus, M.; Wrede, C.E.; Kunz-Schughart, L.A.; Schölmerich, J.; Bollheimer, L.C. Defining high-fat-diet rat models: Metabolic and molecular effects of different fat types. J. Mol. Endocrinol. 2006, 36, 485–501. [Google Scholar] [CrossRef] [Green Version]
- Clifton, P.M.; Keogh, J.B. A systematic review of the effect of dietary saturated and polyunsaturated fat on heart disease. Nutr. Metab. Cardiovasc. Dis. 2017, 27, 1060–1080. [Google Scholar] [CrossRef]
- Lim, K.; Burke, S.L.; Marques, F.Z.; Jackson, K.L.; Gueguen, C.; Sata, Y.; Armitage, J.A.; Head, G.A. Leptin and Melanocortin Signaling Mediates Hypertension in Offspring from Female Rabbits Fed a High-Fat Diet During Gestation and Lactation. Front. Physiol. 2021, 12, 693157. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, E.; Liistro, T.; Mainardi, M.; Pardini, S.; Salvadori, P.A.; Vannucci, A.; Burchielli, S.; Iozzo, P. Maternal high-fat feeding leads to alterations of brain glucose metabolism in the offspring: Positron emission tomography study in a porcine model. Diabetologia 2016, 59, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Salati, J.A.; Roberts, V.H.J.; Schabel, M.C.; Lo, J.O.; Kroenke, C.D.; Lewandowski, K.S.; Lindner, J.R.; Grove, K.L.; Frias, A.E. Maternal high-fat diet reversal improves placental hemodynamics in a nonhuman primate model of diet-induced obesity. Int. J. Obes. 2019, 43, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Chaves, W.F.; Pinheiro, I.L.; da Silva, J.M.; Manhães-de-Castro, R.; da Silva Aragão, R. Repercussions of maternal exposure to high-fat diet on offspring feeding behavior and body composition: A systematic review. J. Dev. Orig. Health Dis. 2021, 12, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Ainge, H.; Thompson, C.; Ozanne, S.E.; Rooney, K.B. A systematic review on animal models of maternal high fat feeding and offspring glycaemic control. Int. J. Obes. 2011, 35, 325–335. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, C.M.; Figueiredo, V.P.; Barbosa, M.A.; Cardoso, L.M.; Alzamora, A.C. Maternal high-fat diet triggers metabolic syndrome disorders that are transferred to first and second offspring generations. Br. J. Nutr. 2020, 123, 59–71. [Google Scholar] [CrossRef]
- Sheen, J.M.; Yu, H.R.; Tain, Y.L.; Tsai, W.L.; Tiao, M.M.; Lin, I.C.; Tsai, C.C.; Lin, Y.J.; Huang, L.T. Combined maternal and postnatal high-fat diet leads to metabolic syndrome and is effectively reversed by resveratrol: A multiple-organ study. Sci. Rep. 2018, 8, 5607. [Google Scholar] [CrossRef]
- Saengnipanthkul, S.; Noh, H.L.; Friedline, R.H.; Suk, S.; Choi, S.; Acosta, N.K.; Tran, D.A.; Hu, X.; Inashima, K.; Kim, A.M.; et al. Maternal exposure to high-fat diet during pregnancy and lactation predisposes normal weight offspring mice to develop hepatic inflammation and insulin resistance. Physiol. Rep. 2021, 9, e14811. [Google Scholar] [CrossRef]
- Wankhade, U.D.; Zhong, Y.; Kang, P.; Alfaro, M.; Chintapalli, S.V.; Piccolo, B.D.; Mercer, K.E.; Andres, A.; Thakali, K.M.; Shankar, K. Maternal High-Fat Diet Programs Offspring Liver Steatosis in a Sexually Dimorphic Manner in Association with Changes in Gut Microbial Ecology in Mice. Sci. Rep. 2018, 8, 16502. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lin, Y.J.; Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Chen, C.C.; Tsai, C.C.; Huang, L.T.; Hsu, C.N. High Fat Diets Sex-Specifically Affect the Renal Transcriptome and Program Obesity, Kidney Injury, and Hypertension in the Offspring. Nutrients 2017, 9, 357. [Google Scholar] [CrossRef] [Green Version]
- Oparil, S.; Acelajado, M.C.; Bakris, G.L.; Berlowitz, D.R.; Cífková, R.; Dominiczak, A.F.; Grassi, G.; Jordan, J.; Poulter, N.R.; Rodgers, A.; et al. Hypertension. Nat. Rev. Dis. Primers 2018, 4, 18014. [Google Scholar] [CrossRef] [Green Version]
- Wali, J.A.; Jarzebska, N.; Raubenheimer, D.; Simpson, S.J.; Rodionov, R.N.; O’Sullivan, J.F. Cardio-Metabolic Effects of High-Fat Diets and Their Underlying Mechanisms-A Narrative Review. Nutrients 2020, 12, 1505. [Google Scholar] [CrossRef]
- Konukoglu, D.; Uzun, H. Endothelial Dysfunction and Hypertension. Adv. Exp. Med. Biol. 2017, 956, 511–540. [Google Scholar]
- Moraes-Souza, R.Q.; Vesentini, G.; Paula, V.G.; Sinzato, Y.K.; Soares, T.S.; Gelaleti, R.B.; Volpato, G.T.; Damasceno, D.C. Oxidative Stress Profile of Mothers and Their Offspring after Maternal Consumption of High-Fat Diet in Rodents: A Systematic Review and Meta-Analysis. Oxid. Med. Cell Longev. 2021, 2021, 9073859. [Google Scholar]
- Bengtsson, S.H.; Gulluyan, L.M.; Dusting, G.J.; Drummond, G.R. Novel isoforms of NADPH oxidase in vascular physiology and pathophysiology. Clin. Exp. Pharmacol. Physiol. 2003, 30, 849–854. [Google Scholar]
- Ketonen, J.; Pilvi, T.; Mervaala, E. Caloric restriction reverses high-fat diet-induced endothelial dysfunction and vascular superoxide production in C57Bl/6 mice. Heart Vessel. 2010, 25, 254–262. [Google Scholar]
- Maneesai, P.; Bunbupha, S.; Kukongviriyapan, U.; Prachaney, P.; Tangsucharit, P.; Kukongviriyapan, V.; Pakdeechote, P. Asiatic acid attenuates renin-angiotensin system activation and improves vascular function in high-carbohydrate, high-fat diet fed rats. BMC Complement Altern. Med. 2016, 16, 123. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Xie, L.; Bi, H.; Li, Y.; Li, Y.; Zhao, J. Urantide alleviates the symptoms of atherosclerotic rats in vivo and in vitro models through the JAK2/STAT3 signaling pathway. Eur. J. Pharmacol. 2021, 902, 174037. [Google Scholar]
- Peredo, H.A.; Lee, H.; Donoso, A.S.; Andrade, V.; Sánchez Eluchans, N.; Puyó, A.M. A high-fat plus fructose diet produces a vascular prostanoid alterations in the rat. Auton. Autacoid Pharmacol. 2015, 34, 35–40. [Google Scholar]
- Li, Q.; Youn, J.Y.; Cai, H. Mechanisms and consequences of endothelial nitric oxide synthase dysfunction in hypertension. J. Hypertens. 2015, 33, 1128–1136. [Google Scholar]
- Chen, P.; Xia, K.; Zhao, Z.; Deng, X.; Yang, T. Atorvastatin modulates the DDAH1/ADMA system in high-fat diet-induced insulin-resistant rats with endothelial dysfunction. Vasc. Med. 2012, 17, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Cardounel, A.J.; Cui, H.; Samouilov, A.; Johnson, W.; Kearns, P.; Tsai, A.L.; Berka, V.; Zweier, J.L. Evidence for the pathophysiological role of endogenous methylarginines in regulation of endothelial NO production and vascular function. J. Biol. Chem. 2007, 282, 879–887. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.M.; Leandro, A.; Azul, L.; Seiça, R.; Perry, G. Vascular Oxidative Stress: Impact and Therapeutic Approaches. Front. Physiol. 2018, 9, 1668. [Google Scholar] [CrossRef] [Green Version]
- Hurr, C.; Young, C.N. Neural Control of Non-vasomotor Organs in Hypertension. Curr. Hypertens. Rep. 2016, 18, 30. [Google Scholar] [CrossRef] [Green Version]
- Chan, S.H.; Chan, J.Y. Brain stem NOS and ROS in neural mechanisms of hypertension. Antioxid. Redox Signal. 2014, 20, 146163. [Google Scholar] [CrossRef]
- Chan, J.Y.H.; Chan, S.H.H. Differential impacts of brain stem oxidative stress and nitrosative stress on sympathetic vasomotor tone. Pharmacol. Ther. 2019, 201, 120–136. [Google Scholar] [CrossRef]
- Chan, S.H.H.; Tai, M.H.; Li, C.Y.; Chan, J.Y.H. Reduction in molecular synthesis or enzyme activity of superoxide dismutases and catalase contributes to oxidative stress and neurogenic hypertension in spontaneously hypertensive rats. Free Radic. Biol. Med. 2006, 40, 2028–2039. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E. Effect of High-Fat Diets on Oxidative Stress, Cellular Inflammatory Response and Cognitive Function. Nutrients 2019, 11, 2579. [Google Scholar] [CrossRef] [Green Version]
- Nagae, A.; Fujita, M.; Kawarazaki, H.; Matsui, H.; Ando, K.; Fujita, T. Sympathoexcitation by oxidative stress in the brain mediates arterial pressure elevation in obesity-induced hypertension. Circulation 2009, 119, 978–986. [Google Scholar] [CrossRef]
- Kishi, T.; Hirooka, Y.; Ogawa, K.; Konno, S.; Sunagawa, K. Calorie restriction inhibits sympathetic nerve activity via anti-oxidant effect in the rostral ventrolateral medulla of obesity-induced hypertensive rats. Clin. Exp. Hypertens. 2011, 33, 240–245. [Google Scholar] [CrossRef]
- Chung, S.; Park, C.W.; Shin, S.J.; Lim, J.H.; Chung, H.W.; Youn, D.Y.; Kim, H.W.; Kim, B.S.; Lee, J.H.; Kim, G.H.; et al. Tempol or candesartan prevents high-fat diet-induced hypertension and renal damage in spontaneously hypertensive rats. Nephrol. Dial. Transplant. 2010, 25, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Nizar, J.M.; Bhalla, V. Molecular Mechanisms of Sodium-Sensitive Hypertension in the Metabolic Syndrome. Curr. Hypertens. Rep. 2017, 19, 60. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lai, W.T.; Lin, Y.J.; Tain, Y.L. Postnatal high-fat diet sex-specifically exacerbates prenatal dexamethasone-induced hypertension: Mass spectrometry-based quantitative proteomic approach. J. Nutr. Biochem. 2018, 57, 268–275. [Google Scholar] [CrossRef]
- Wu, J.; Lai, G.; Chen, F.; Zhang, B.; Zhao, Y. Renal NKCC2 Is Dual Regulated by the Synergy of 20-HETE and High-Fat Diet in CYP4F2 Transgenic Mice. Kidney Blood Press. Res. 2021, 46, 601–612. [Google Scholar] [CrossRef]
- Li, C.; Culver, S.A.; Quadri, S.; Ledford, K.L.; Al-Share, Q.Y.; Ghadieh, H.E.; Najjar, S.M.; Siragy, H.M. High-fat diet amplifies renal renin angiotensin system expression, blood pressure elevation, and renal dysfunction caused by Ceacam1 null deletion. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E802–E810. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Lin, Y.; Luo, R.; Chen, S.; Wang, F.; Zheng, P.; Levi, M.; Yang, T.; Wang, W. Intrarenal renin-angiotensin system mediates fatty acid-induced ER stress in the kidney. Am. J. Physiol. Ren. Physiol. 2016, 310, F351–F363. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.N.; Tain, Y.L. Targeting the renin–angiotensin–aldosterone system to prevent hypertension and kidney disease of developmental origins. Int. J. Mol. Sci. 2021, 22, 2298. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Hypertension of Developmental Origins: Consideration of Gut Microbiome in Animal Models. Biomedicines 2022, 10, 875. [Google Scholar] [CrossRef]
- Lin, X.H.; Gao, L.; Tian, S.; Klausen, C.; Guo, M.X.; Gao, Q.; Liu, M.E.; Wang, H.; Wu, D.D.; Zhou, C.L.; et al. Maternal high-fat-diet exposure is associated with elevated blood pressure and sustained increased leptin levels through epigenetic memory in offspring. Sci. Rep. 2021, 11, 316. [Google Scholar] [CrossRef]
- Resende, A.C.; Emiliano, A.F.; Cordeiro, V.S.; de Bem, G.F.; de Cavalho, L.C.; de Oliveira, P.R.; Neto, M.L.; Costa, C.A.; Boaventura, G.T.; de Moura, R.S. Grape skin extract protects against programmed changes in the adult rat offspring caused by maternal high-fat diet during lactation. J. Nutr. Biochem. 2013, 24, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.Y.; Taylor, P.D.; Dekou, V.; Seed, P.T.; Lakasing, L.; Graham, D.; Dominiczak, A.F.; Hanson, M.A.; Poston, L. Gender-linked hypertension in offspring of lard-fed pregnant rats. Hypertension 2003, 41, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Dekou, V.; Hanson, M.; Poston, L.; Taylor, P. Predictive adaptive responses to maternal high-fat diet prevent endothelial dysfunction but not hypertension in adult rat offspring. Circulation 2004, 110, 1097–1102. [Google Scholar] [CrossRef] [Green Version]
- Do Nascimento, L.C.P.; Neto, J.P.R.C.; de Andrade Braga, V.; Lagranha, C.J.; de Brito Alves, J.L. Maternal exposure to high-fat and high-cholesterol diet induces arterial hypertension and oxidative stress along the gut-kidney axis in rat offspring. Life Sci. 2020, 261, 118367. [Google Scholar] [CrossRef]
- Do Nascimento, L.C.P.; de Souza, E.L.; de Luna Freire, M.O.; de Andrade Braga, V.; de Albuqeurque, T.M.R.; Lagranha, C.J.; de Brito Alves, J.L. Limosilactobacillus fermentum prevent gut-kidney oxidative damage and the rise in blood pressure in male rat offspring exposed to a maternal high-fat diet. J. Dev. Orig. Health Dis. 2022, 19, 1–8. [Google Scholar]
- Gray, C.; Vickers, M.H.; Segovia, S.A.; Zhang, X.D.; Reynolds, C.M. A maternal high fat diet programmes endothelial function and cardiovascular status in adult male offspring independent of body weight, which is reversed by maternal conjugated linoleic acid (CLA) supplementation. PLoS ONE 2015, 10, e0115994. [Google Scholar]
- Torrens, C.; Ethirajan, P.; Bruce, K.D.; Cagampang, F.R.; Siow, R.C.; Hanson, M.A.; Byrne, C.D.; Mann, G.E.; Clough, G.F. Interaction between maternal and offspring diet to impair vascular function and oxidative balance in high fat fed male mice. PLoS ONE 2012, 7, e50671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Chan, J.Y.H.; Lee, C.T.; Tain, Y.L. Hypertension Programmed by Perinatal High-Fat Diet: Effect of Maternal Gut Microbiota-Targeted Therapy. Nutrients 2019, 11, E2908. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Lin, Y.J.; Sheen, J.M.; Lin, I.C.; Yu, H.R.; Huang, L.T.; Hsu, C.N. Resveratrol prevents the combined maternal plus postweaning high-fat-diets-induced hypertension in male offspring. J. Nutr. Biochem. 2017, 48, 120–127. [Google Scholar] [CrossRef]
- Hsu, C.N.; Lin, Y.J.; Tain, Y.L. Maternal Exposure to Bisphenol a Combined with High-Fat Diet-Induced Programmed Hypertension in Adult Male Rat Offspring: Effects of Resveratrol. Int. J. Mol. Sci. 2019, 20, 4382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.N.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Tain, Y.L. Maternal Garlic Oil Supplementation Prevents High-Fat Diet-Induced Hypertension in Adult Rat Offspring: Implications of H2S-Generating Pathway in the Gut and Kidneys. Mol. Nutr. Food Res. 2021, 65, e2001116. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Hou, C.Y.; Lee, C.T.; Chan, J.Y.H.; Tain, Y.L. The Interplay between Maternal and Post-Weaning High-Fat Diet and Gut Microbiota in the Developmental Programming of Hypertension. Nutrients 2019, 11, 1982. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Krege, J.H.; Hodgin, J.B.; Hagaman, J.R.; Smithies, O. A noninvasive computerized tail-cuff system for measuring blood pressure in mice. Hypertension 1995, 25, 1111–1115. [Google Scholar] [CrossRef]
- Mizuno, M.; Siddique, K.; Baum, M.; Smith, S.A. Prenatal programming of hypertension induces sympathetic overactivity in response to physical stress. Hypertension 2013, 61, 180–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messer, L.C.; Boone-Heinonen, J.; Mponwane, L.; Wallack, L.; Thornburg, K.L. Developmental Programming: Priming Disease Susceptibility for Subsequent Generations. Curr. Epidemiol. Rep. 2015, 2, 37–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.L.; Huang, L.T. Restoration of asymmetric dimethylarginine-nitric oxide balance to prevent the development of hypertension. Int. J. Mol. Sci. 2014, 15, 11773–11782. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N.; Chan, J.Y.; Huang, L.T. Renal Transcriptome Analysis of Programmed Hypertension Induced by Maternal Nutritional Insults. Int. J. Mol. Sci. 2015, 16, 17826–17837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherrer, U.; Rimoldi, S.F.; Sartori, C.; Messerli, F.H.; Rexhaj, E. Fetal programming and epigenetic mechanisms in arterial hypertension. Curr. Opin. Cardiol. 2015, 30, 393–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tain, Y.L.; Hsu, C.N. Interplay between Oxidative Stress and Nutrient Sensing Signaling in the Developmental Origins of Cardiovascular Disease. Int. J. Mol. Sci. 2017, 18, 841. [Google Scholar] [CrossRef]
- Dennery, P.A. Oxidative stress in development: Nature or nurture? Free Radic. Biol. Med. 2010, 49, 1147–1151. [Google Scholar] [CrossRef]
- Tain, Y.L.; Hsu, C.N. Oxidative Stress-Induced Hypertension of Developmental Origins: Preventive Aspects of Antioxidant Therapy. Antioxidants 2022, 11, 511. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qi, L.; Wu, J.; Xu, T.; Yang, C.; Chen, X.; Lv, J.; Xu, Z. Prenatal high-salt diet impaired vasodilatation with reprogrammed renin-angiotensin system in offspring rats. J. Hypertens. 2018, 36, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Svitok, P.; Okuliarova, M.; Varga, I.; Zeman, M. Renal impairment induced by prenatal exposure to angiotensin II in male rat offspring. Exp. Biol. Med. 2019, 244, 923–931. [Google Scholar] [CrossRef] [PubMed]
- Tsai, W.L.; Hsu, C.N.; Tain, Y.L. Whether AICAR in Pregnancy or Lactation Prevents Hypertension Programmed by High Saturated Fat Diet: A Pilot Study. Nutrients 2020, 12, 448. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Zadi, Z.H.; Zhang, J.; Scharf, S.M.; Pae, E.K. Intermittent hypoxia in utero damages postnatal growth and cardiovascular function in rats. J. Appl. Physiol. 2018, 124, 821–830. [Google Scholar] [CrossRef]
- Piecha, G.; Koleganova, N.; Ritz, E.; Müller, A.; Fedorova, O.V.; Bagrov, A.Y.; Lutz, D.; Schirmacher, P.; Gross-Weissmann, M.L. High salt intake causes adverse fetal programming--vascular effects beyond blood pressure. Nephrol. Dial. Transplant. 2012, 27, 3464–3476. [Google Scholar] [CrossRef] [Green Version]
- Marrocco, I.; Altieri, F.; Peluso, I. Measurement and Clinical Significance of Biomarkers of Oxidative Stress in Humans. Oxidative Med. Cell. Lolongevity 2017, 2017, 6501046. [Google Scholar] [CrossRef]
- Tain, Y.L.; Lee, C.T.; Chan, J.Y.; Hsu, C.N. Maternal melatonin or N-acetylcysteine therapy regulates hydrogen sulfide-generating pathway and renal transcriptome to prevent prenatal N(G)-Nitro-L-argininemethyl ester (L-NAME)-induced fetal programming of hypertension in adult male offspring. Am. J. Obstet. Gynecol. 2016, 215, 636. [Google Scholar] [CrossRef]
- Hsu, C.N.; Tain, Y.L. Regulation of nitric oxide production in the developmental programming of hypertension and kidney disease. Int. J. Mol. Sci. 2019, 20, 681. [Google Scholar] [CrossRef] [Green Version]
- Majzunova, M.; Dovinova, I.; Barancik, M.; Chan, J.Y. Redox signaling in pathophysiology of hypertension. J. Biomed. Sci. 2013, 20, 69. [Google Scholar] [CrossRef] [Green Version]
- Sheen, J.M.; Yu, H.R.; Tiao, M.M.; Chen, C.C.; Huang, L.T.; Chang, H.Y.; Tain, Y.L. Prenatal dexamethasone-induced programmed hypertension and renal programming. Life Sci. 2015, 132, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hsu, C.N. Toxic Dimethylarginines: Asymmetric Dimethylarginine (ADMA) and Symmetric Dimethylarginine (SDMA). Toxins (Basel) 2017, 9, E92. [Google Scholar] [CrossRef] [Green Version]
- Te Riet, L.; van Esch, J.H.; Roks, A.J.; van den Meiracker, A.H.; Danser, A.H. Hypertension: Renin-angiotensin-aldosterone system alterations. Circ. Res. 2015, 116, 960–975. [Google Scholar] [CrossRef]
- Vehaskari, V.M.; Stewart, T.; Lafont, D.; Soyez, C.; Seth, D.; Manning, J. Kidney angiotensin and angiotensin receptor expression in prenatally programmed hypertension. Am. J. Physiol.-Ren. Physiol. 2004, 287, F262–F267. [Google Scholar] [CrossRef] [PubMed]
- Jansson, T.; Powell, T. Role of Placental Nutrient Sensing in Developmental Programming. Clin. Obstet. Gynecol. 2013, 56, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Efeyan, A.; Comb, W.C.; Sabatini, D.M. Nutrient-sensing mechanisms and pathways. Nature 2015, 517, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N.; Chan, J.Y. PPARs link early life nutritional insults to later programmed hypertension and metabolic syndrome. Int. J. Mol. Sci. 2015, 17, 20. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N. AMP-Activated Protein Kinase as a Reprogramming Strategy for Hypertension and Kidney Disease of Developmental Origin. Int. J. Mol. Sci. 2018, 19, 1744. [Google Scholar] [CrossRef] [Green Version]
- Goyal, D.; Limesand, S.W.; Goyal, R. Epigenetic responses and the developmental origins of health and disease. J. Endocrinol. 2019, 242, T105–T119. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.J.; Huang, L.T.; Tsai, C.C.; Sheen, J.M.; Tiao, M.M.; Yu, H.R.; Lin, I.C.; Tain, Y.L. Maternal high-fat diet sex-specifically alters placental morphology and transcriptome in rats: Assessment by next-generation sequencing. Placenta 2019, 78, 44–53. [Google Scholar] [CrossRef]
- Gawlińska, K.; Gawliński, D.; Borczyk, M.; Korostyński, M.; Przegaliński, E.; Filip, M. A Maternal High-Fat Diet during Early Development Provokes Molecular Changes Related to Autism Spectrum Disorder in the Rat Offspring Brain. Nutrients 2021, 13, 3212. [Google Scholar] [CrossRef] [PubMed]
- Preston, C.C.; Larsen, T.D.; Eclov, J.A.; Louwagie, E.J.; Gandy, T.C.T.; Faustino, R.S.; Baack, M.L. Maternal High Fat Diet and Diabetes Disrupts Transcriptomic Pathways That Regulate Cardiac Metabolism and Cell Fate in Newborn Rat Hearts. Front. Endocrinol. 2020, 11, 570846. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C.; Akanji, A.O. Leptin, Obesity, and Hypertension: A Review of Pathogenetic Mechanisms. Metab. Syndr. Relat. Disord. 2020, 18, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [Green Version]
- Arrieta, M.C.; Stiemsma, L.T.; Amenyogbe, N.; Brown, E.M.; Finlay, B. The intestinal microbiome in early life: Health and disease. Front. Immunol. 2014, 5, 427. [Google Scholar] [CrossRef] [Green Version]
- Wolters, M.; Ahrens, J.; Romaní-Pérez, M.; Watkins, C.; Sanz, Y.; Benítez-Páez, A.; Stanton, C.; Günther, K. Dietary fat, the gut microbiota, and metabolic health—A systematic review conducted within the MyNewGut project. Clin. Nutr. 2019, 38, 2504–2520. [Google Scholar] [CrossRef] [Green Version]
- Guimarães, K.S.L.; Braga, V.A.; Noronha, S.I.S.R.; Costa, W.K.A.D.; Makki, K.; Cruz, J.C.; Brandão, L.R.; Chianca Junior, D.A.; Meugnier, E.; Leulier, F.; et al. Lactiplantibacillus plantarum WJL administration during pregnancy and lactation improves lipid profile, insulin sensitivity and gut microbiota diversity in dyslipidemic dams and protects male offspring against cardiovascular dysfunction in later life. Food Funct. 2020, 11, 8939–8950. [Google Scholar] [CrossRef]
- Mosca, A.; Leclerc, M.; Hugot, J.P. Gut Microbiota Diversity and Human Diseases: Should We Reintroduce Key Predators in Our Ecosystem? Front. Microbiol. 2016, 7, 455. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. The gut microbiota and the brain-gut-kidney axis in hypertension and chronic kidney disease. Nat. Rev. Nephrol. 2018, 14, 442–456. [Google Scholar] [CrossRef]
- DiRienzo, D.B. Effect of probiotics on biomarkers of cardiovascular disease: Implications for heart-healthy diets. Nutr. Rev. 2014, 72, 18–29. [Google Scholar] [CrossRef]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef]
- Pluznick, J.L. Microbial Short-Chain Fatty Acids and Blood Pressure Regulation. Curr. Hypertens. Rep. 2017, 19, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaworska, K.; Hering, D.; Mosieniak, G.; Bielak-Zmijewska, A.; Pilz, M.; Konwerski, M.; Gasecka, A.; Kapłon-Cie’slicka, A.; Filipiak, K.; Sikora, E.; et al. TMA, A Forgotten Uremic Toxin, but Not TMAO, Is Involved in Cardiovascular Pathology. Toxins 2019, 11, 490. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.; Zheng, L.; Zhuang, R.; Yu, P.; Xu, Z.; Liu, G.; Xi, X.; Zhou, X.; Fan, H. The Gut Microbial Metabolite Trimethylamine N-Oxide and Hypertension Risk: A Systematic Review and Dose-Response Meta-analysis. Adv. Nutr. 2019, 11, 66–76. [Google Scholar] [CrossRef]
- Szabo, C. A timeline of hydrogen sulfide (H2S) research: From environmental toxin to biological mediator. Biochem. Pharmacol. 2018, 149, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. Preventing developmental origins of cardiovascular disease: Hydrogen sulfide as a potential target? Antioxidants 2021, 10, 247. [Google Scholar] [CrossRef] [PubMed]
- Tomat, A.L.; Salazar, F.J. Mechanisms involved in developmental programming of hypertension and renal diseases. Gender differences. Horm. Mol. Biol. Clin. Investig. 2014, 18, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, N.B.; Intapad, S.; Alexander, B.T. Sex differences in the developmental programming of hypertension. Acta Physiol. 2014, 210, 307–316. [Google Scholar] [CrossRef] [Green Version]
- Harkins, C.P.; Kong, H.H.; Segre, J.A. Manipulating the Human Microbiome to Manage Disease. JAMA 2020, 323, 303–304. [Google Scholar] [CrossRef]
- Oniszczuk, A.; Oniszczuk, T.; Gancarz, M.; Szyma’nska, J. Role of gut microbiota, probiotics and prebiotics in the cardiovascular diseases. Molecules 2021, 26, 1172. [Google Scholar] [CrossRef]
- Chi, C.; Li, C.; Wu, D.; Buys, N.; Wang, W.; Fan, H.; Sun, J. Effects of Probiotics on Patients with Hypertension: A Systematic Review and Meta-Analysis. Curr. Hypertens. Rep. 2020, 22, 34. [Google Scholar] [CrossRef] [PubMed]
- Zółkiewicz, J.; Marzec, A.; Ruszczyn’ski, M.; Feleszko, W. Postbiotics—A step beyond pre- and probiotics. Nutrients 2020, 12, 2189. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.N.; Tain, Y.L. The Double-Edged Sword Effects of Maternal Nutrition in the Developmental Programming of Hypertension. Nutrients 2018, 10, 1917. [Google Scholar] [CrossRef] [Green Version]
- Tain, Y.L.; Hsu, C.N. Novel Insights on Dietary Polyphenols for Prevention in Early-Life Origins of Hypertension: A Review Focusing on Preclinical Animal Models. Int. J. Mol. Sci. 2022, 23, 6620. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic diagram summarizing the adverse offspring outcomes related to maternal high-fat diet.
Figure 1.
Schematic diagram summarizing the adverse offspring outcomes related to maternal high-fat diet.

Figure 2.
Schematic diagram of the common mechanisms linking maternal high-fat diet to offspring hypertension.
Figure 2.
Schematic diagram of the common mechanisms linking maternal high-fat diet to offspring hypertension.

Figure 3.
Schematic diagram of the potential reprogramming strategies for prevention of maternal high-fat-diet-induced offspring hypertension.
Figure 3.
Schematic diagram of the potential reprogramming strategies for prevention of maternal high-fat-diet-induced offspring hypertension.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Programming effects of maternal high-fat diet on offspring hypertension.
| Energy Percent from Fat in Maternal High-Fat Diet | Species/Sex | Intervention Period | Offspring Obesity | Programming Effects | Age at Measure | Ref. |
|---|---|---|---|---|---|---|
| 13.4% | Rabbit/M+F | Pregnancy and Lactation | No | Increased central leptin signaling and sympathetic responsivity | 20 weeks | [21] |
| 18.7% | SD rat/M | Pregnancy | No | Increased leptin expression and leptin promoter hypomethylation in adipose tissues | 1 year | [59] |
| 24% | Wistar rat/M | Lactation | Yes | Decreased plasma and mesenteric arteries antioxidant activities, and decreased NO | 22 weeks | [60] |
| 25.7% | SD rat/M+F | Lactation | Yes in females | Endothelial dysfunction | 25 weeks | [61] |
| 25.7% | SD rat/F | Pregnancy and Lactation | Yes | Endothelial dysfunction | 180 days | [62] |
| 31% | Wistar rat/M+F | Pregnancy and Lactation | ND | Reduced SOD activity and increased lipid peroxidation in the kidneys | 90 days | [63] |
| 31% | Wistar rat/M | Pregnancy and Lactation | ND | Increased oxidative stress in the kidneys | 100 days | [64] |
| 45% | SD rat/M | Pregnancy and Lactation | Yes | Endothelial dysfunction and reduced NO | 130 days | [65] |
| 45% | C57BL6J mice/M | Pregnancy and Lactation | Yes | Endothelial dysfunction, increased ROS, and reduced NO in femoral artery | 30 weeks | [66] |
| 58% | SD rat/M | Pregnancy and Lactation | No | Increased renal AT1R expression and shifts in gut microbiota | 16 weeks | [67] |
| 58% | SD rat/M | Pregnancy and Lactation | No | Increased renal oxidative stress, decreased urinary NO level, and decreased renal Ang-(1–7) level | 16 weeks | [68] |
| 58% | SD rat/M | Pregnancy and Lactation | No | Increased renal oxidative stress and decreased urinary NO level | 16 weeks | [69] |
| 58% | SD rat/M | Pregnancy and Lactation | Yes | Dysregulated H2S-generating pathway and shifts in gut microbiota | 16 weeks | [70] |
| 58% | SD rat/M | Pregnancy and Lactation | No | Dysregulated nutrient-sensing signals and shifts in gut microbiota | 16 weeks | [71] |
Studies tabulated according to energy percent from fat in maternal diet, species, and age at measure. SD = Sprague Dawley; M = male; F = female; ND = Not determined; NO = nitric oxide; ROS = reactive oxygen species; SOD = superoxide dismutase; AT1R = angiotensin II type 1 receptor; H2S = hydrogen sulfide.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tain, Y.-L.; Hsu, C.-N. Maternal High-Fat Diet and Offspring Hypertension. Int. J. Mol. Sci. 2022, 23, 8179. https://doi.org/10.3390/ijms23158179
AMA Style
Tain Y-L, Hsu C-N. Maternal High-Fat Diet and Offspring Hypertension. International Journal of Molecular Sciences. 2022; 23(15):8179. https://doi.org/10.3390/ijms23158179
Chicago/Turabian StyleTain, You-Lin, and Chien-Ning Hsu. 2022. "Maternal High-Fat Diet and Offspring Hypertension" International Journal of Molecular Sciences 23, no. 15: 8179. https://doi.org/10.3390/ijms23158179
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.