
The Effect of Leptin on the Blood Hormonal Profile (Cortisol, Insulin, Thyroid Hormones) of the Ewe in Acute Inflammation in Two Different Photoperiodical Conditions

 , , and
, , and
Abstract
:1. Introduction
2. Results
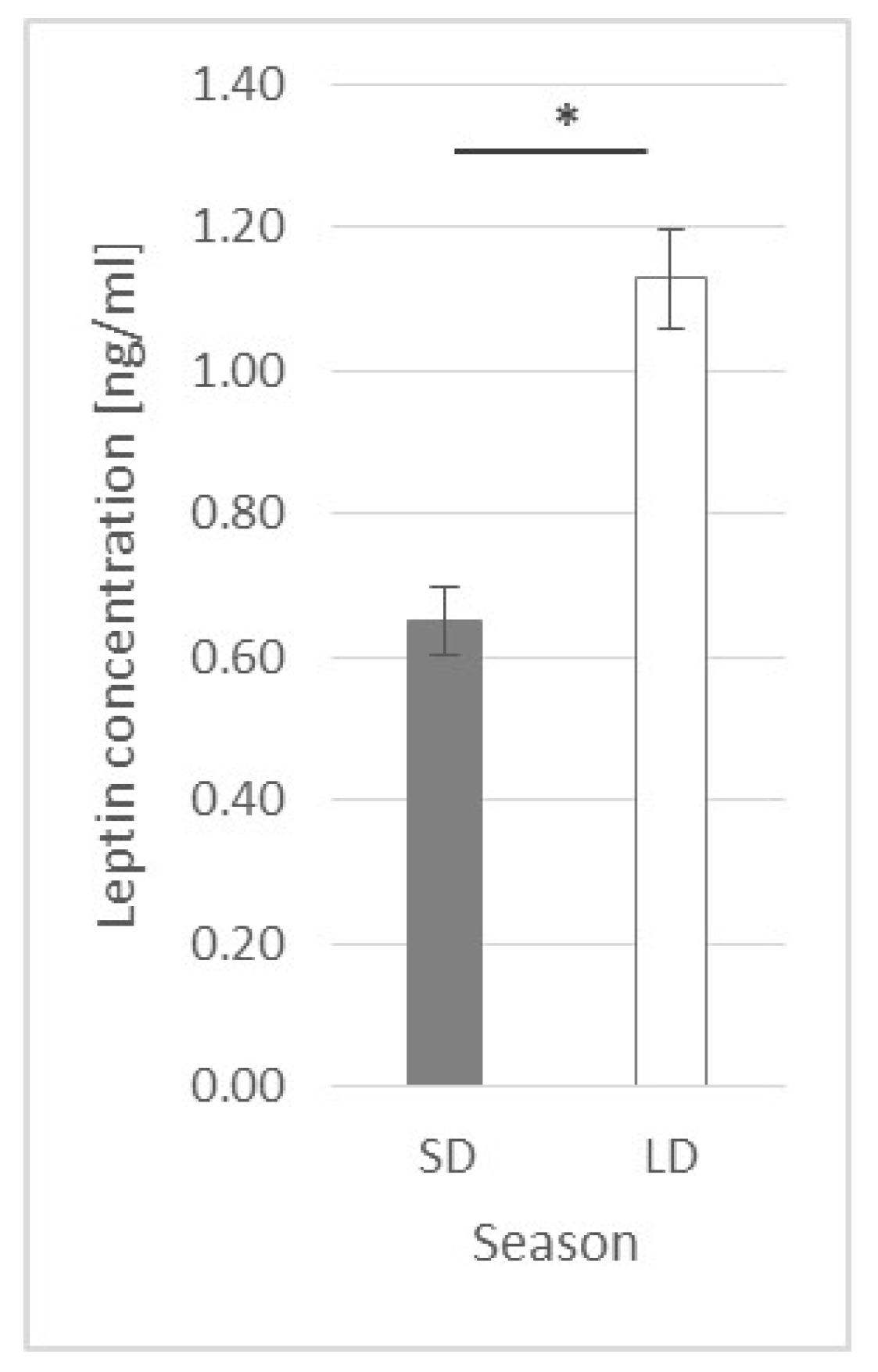
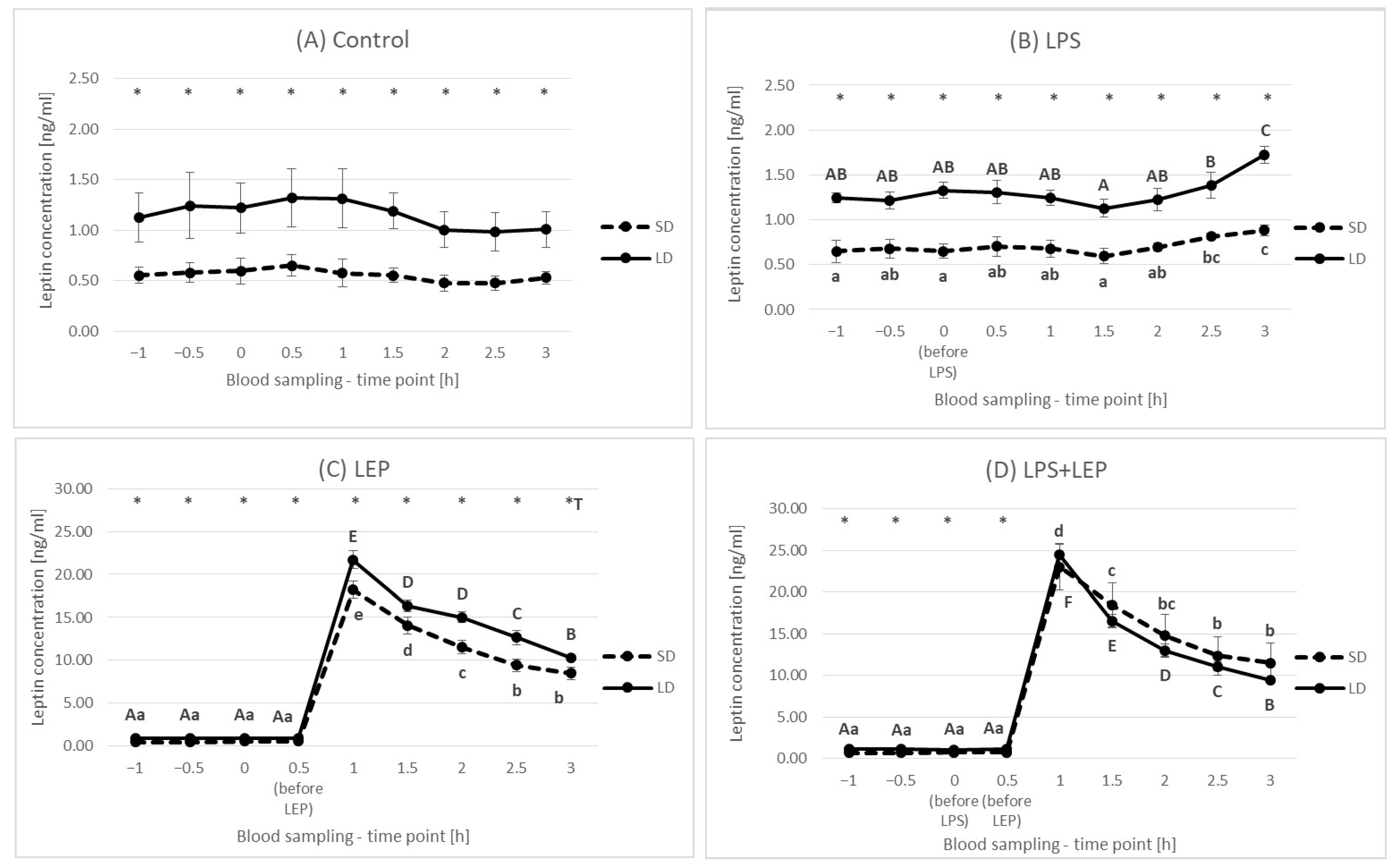
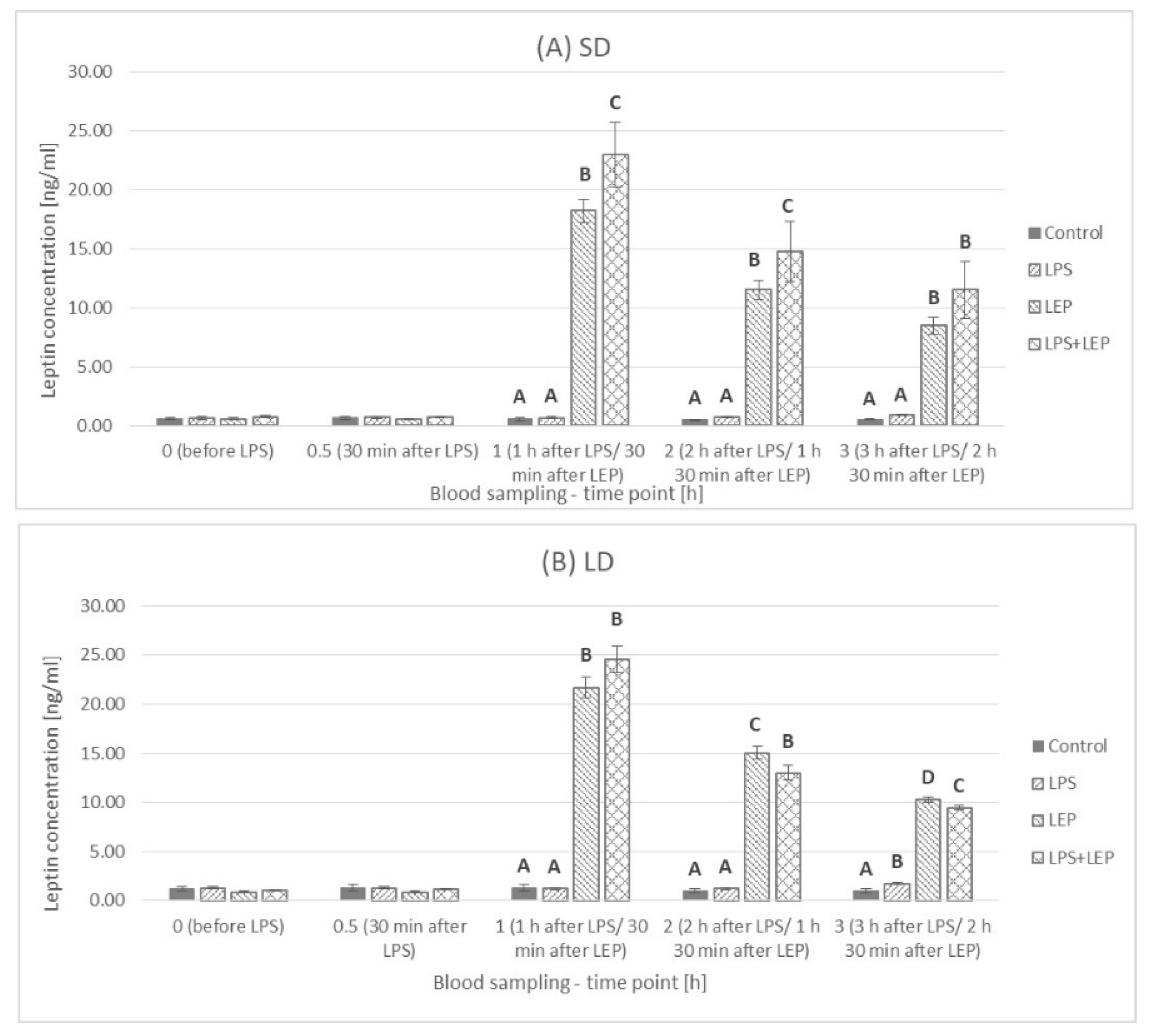
2.1. Leptin Plasma Concentration
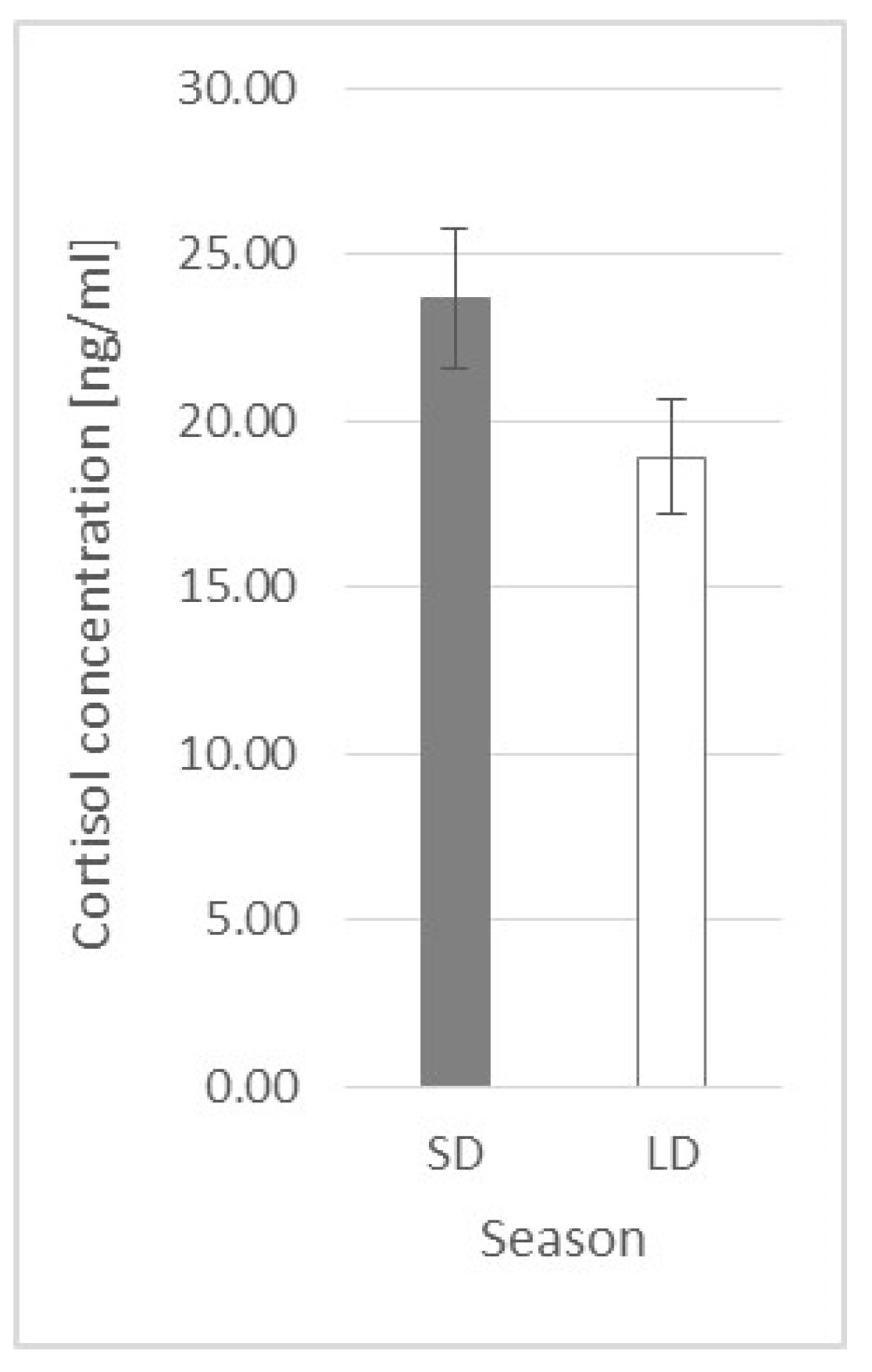
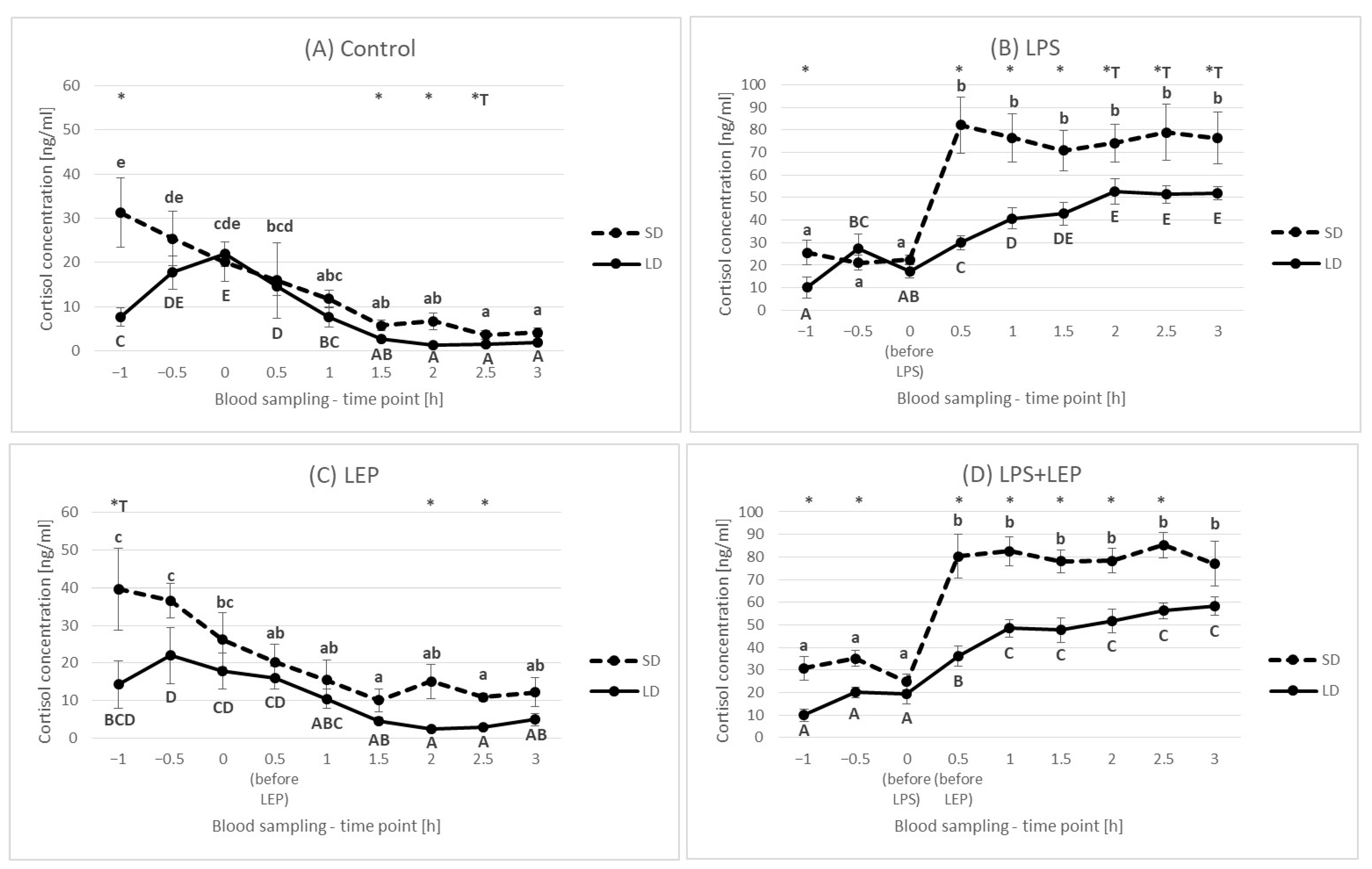
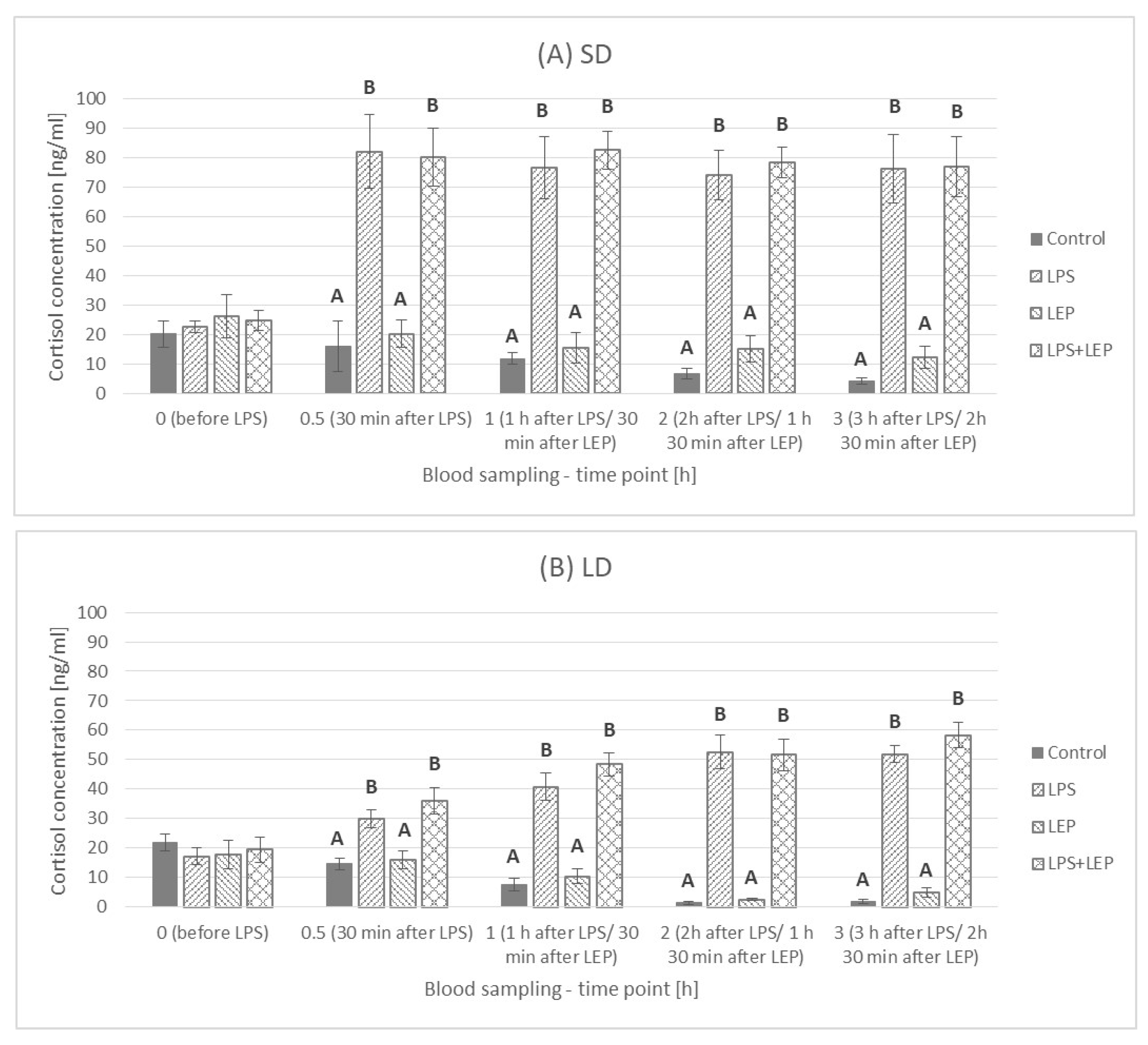
2.2. Cortisol Plasma Concentration
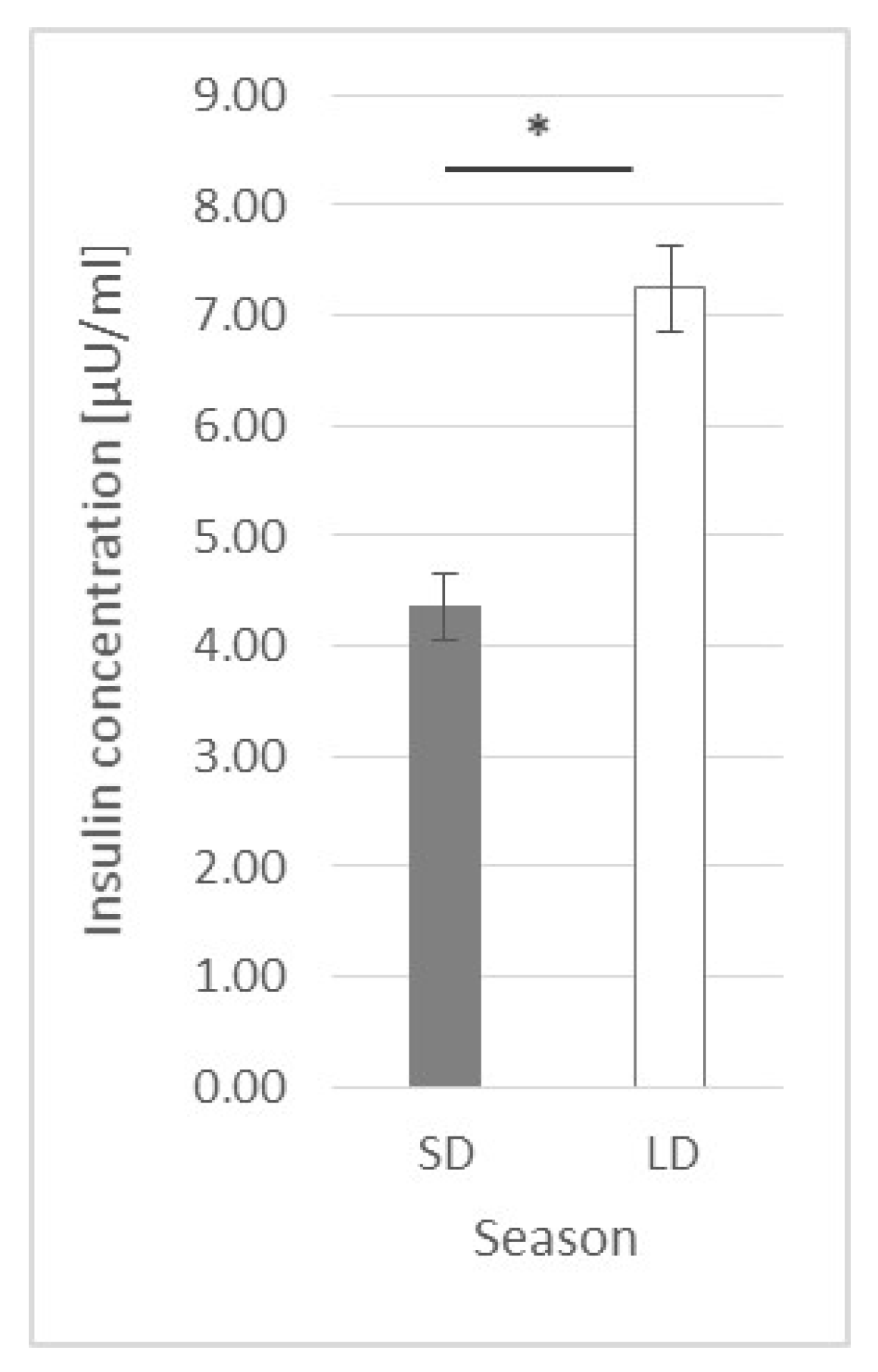
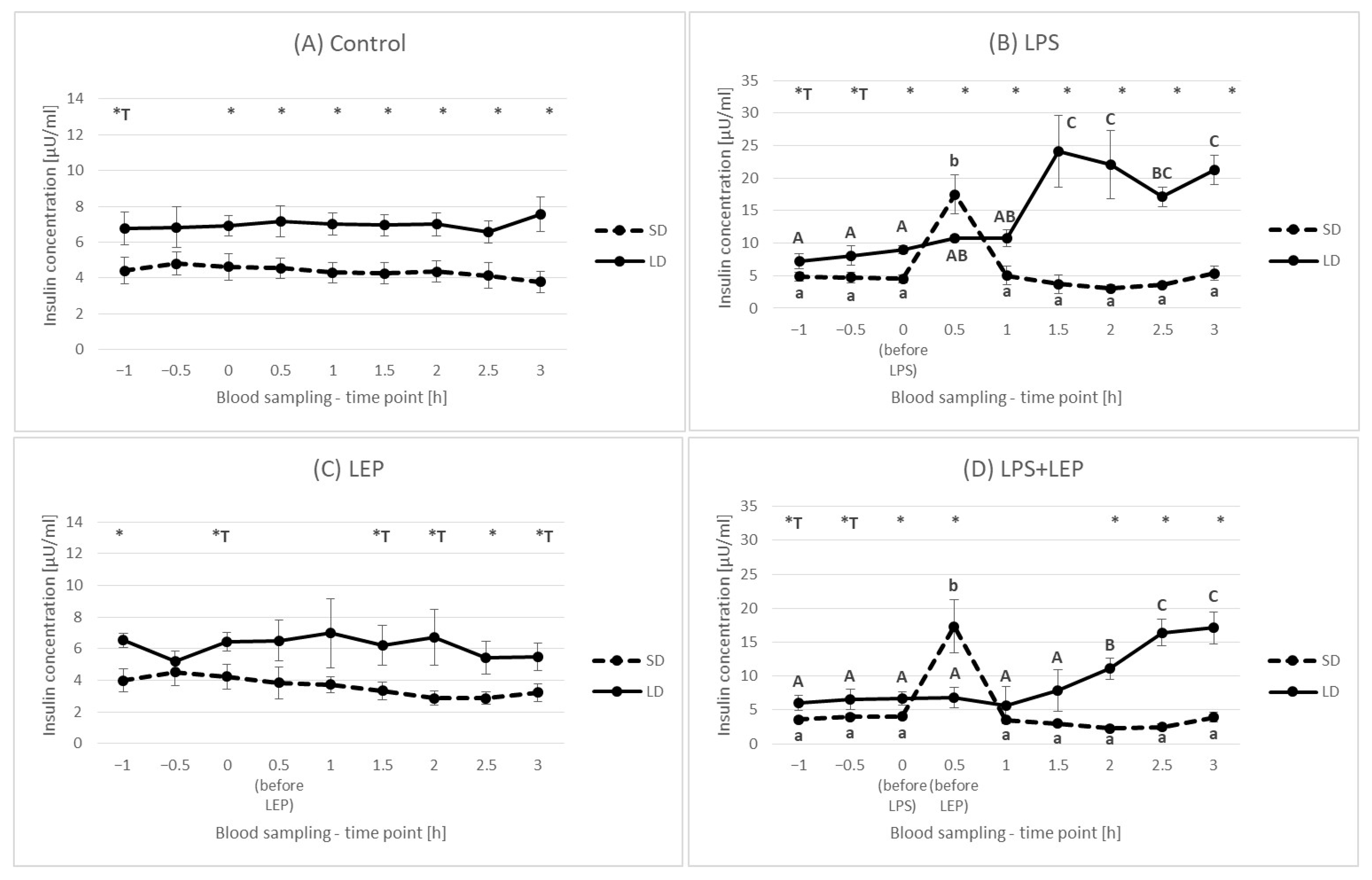
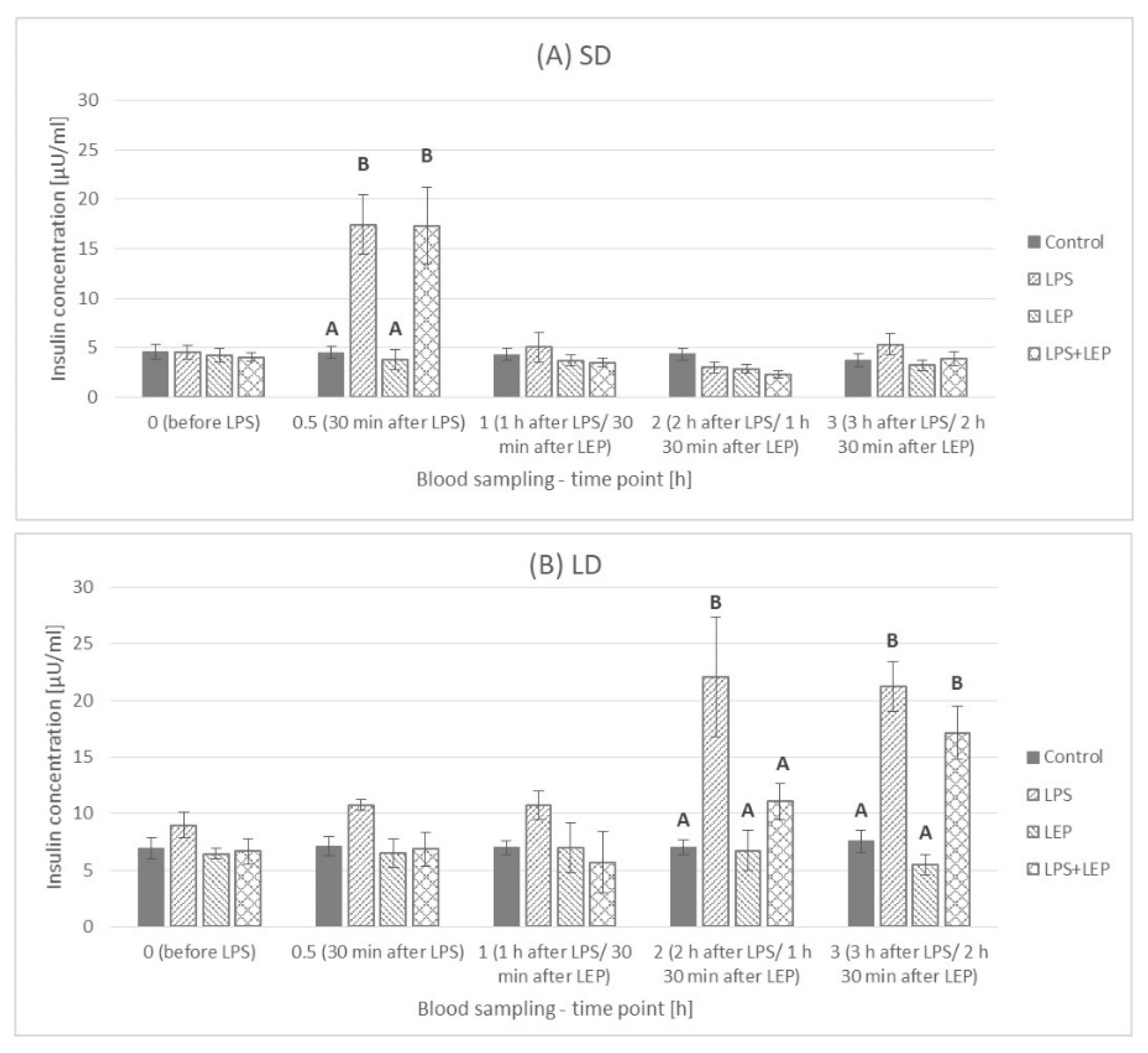
2.3. Insulin Plasma Concentration
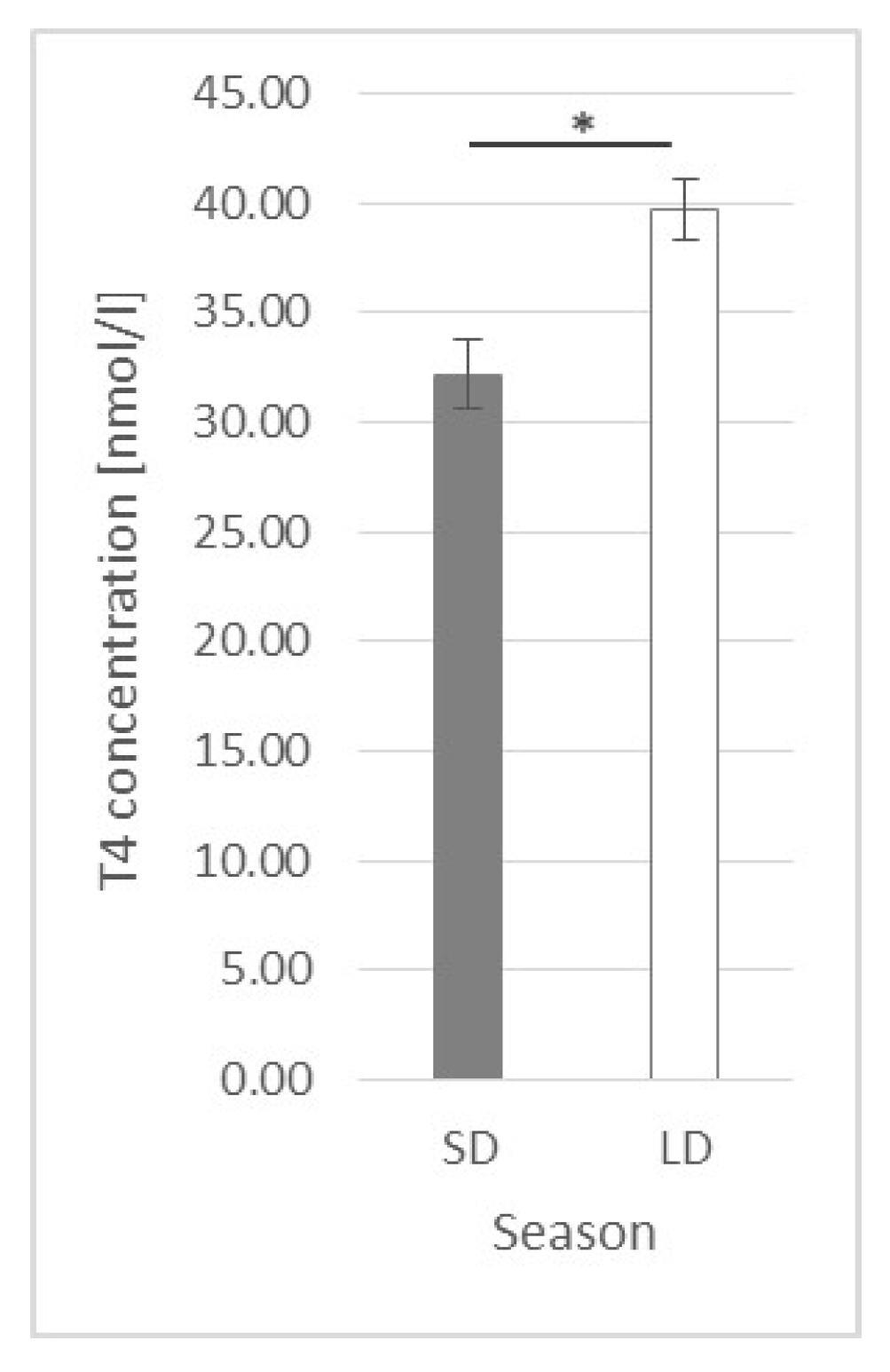
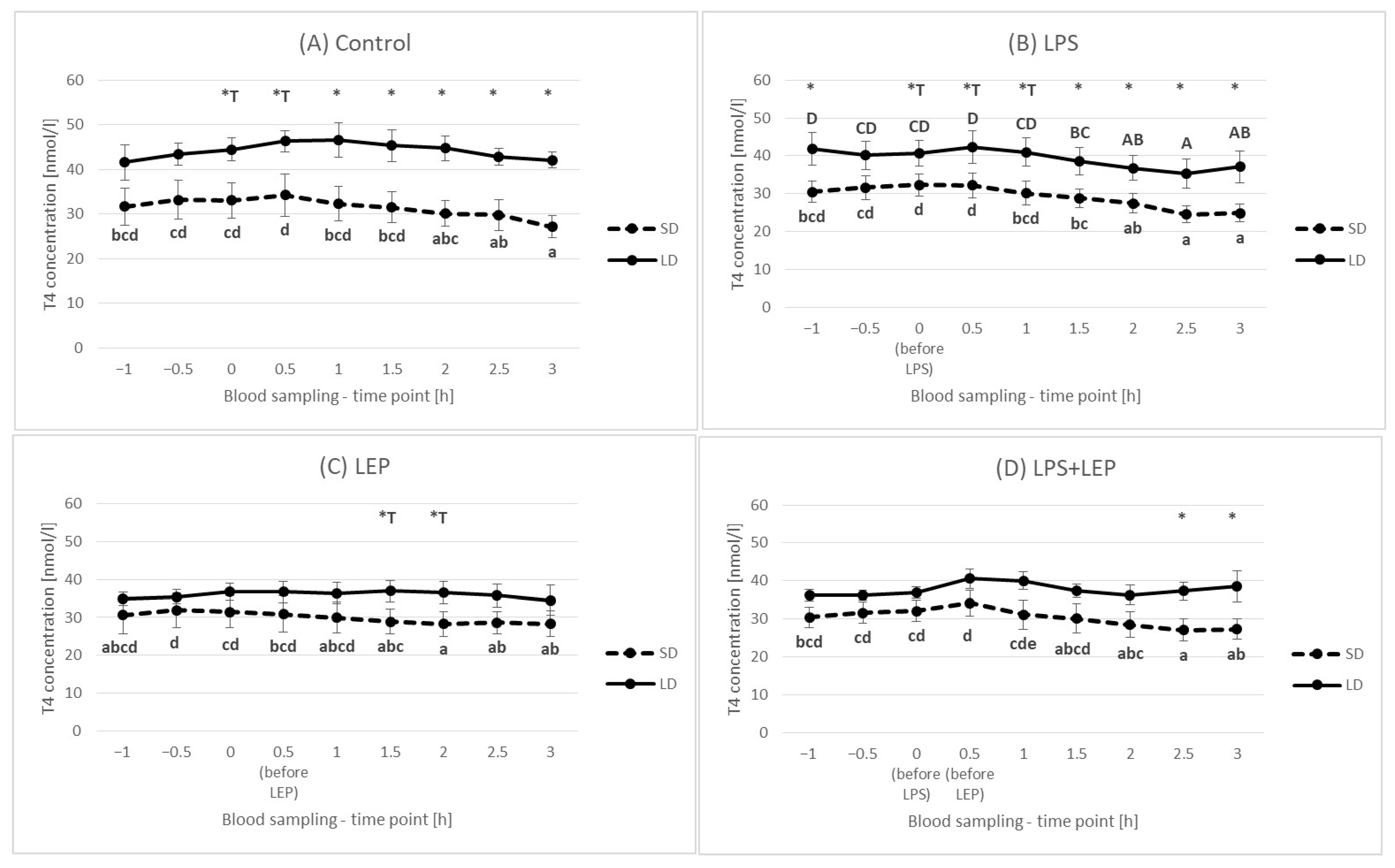
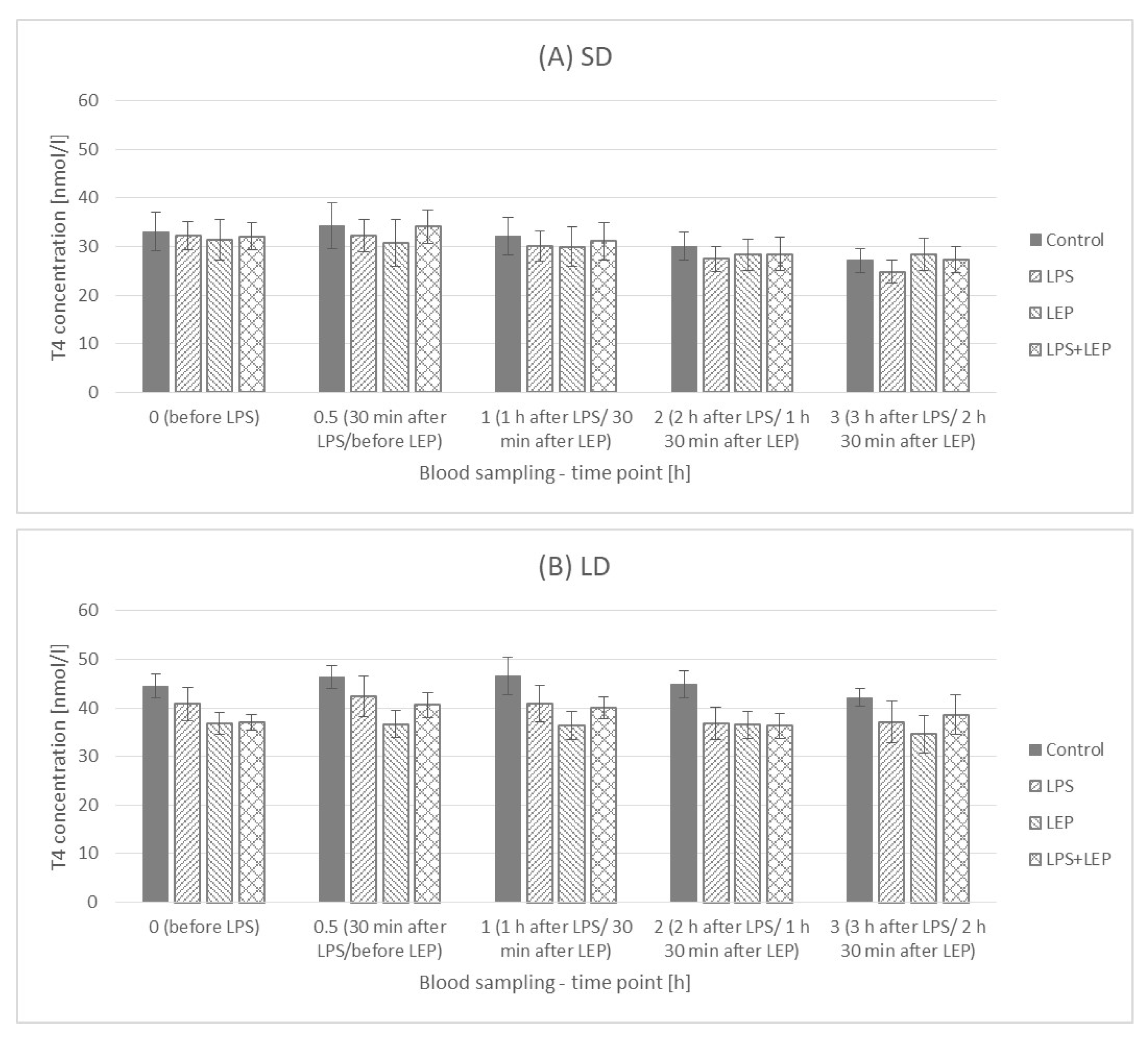
2.4. Thyroxine (T4) Plasma Concentration
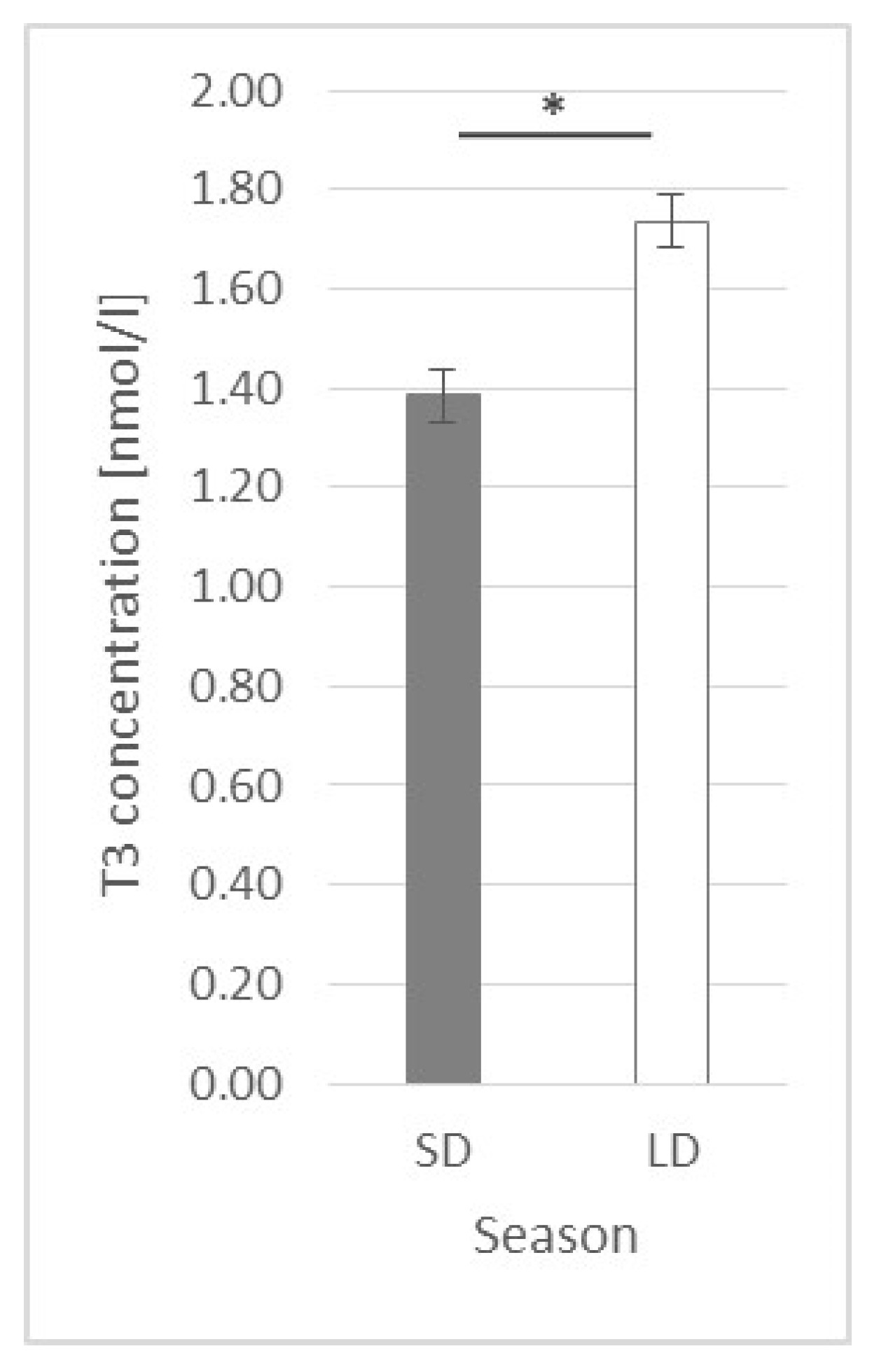
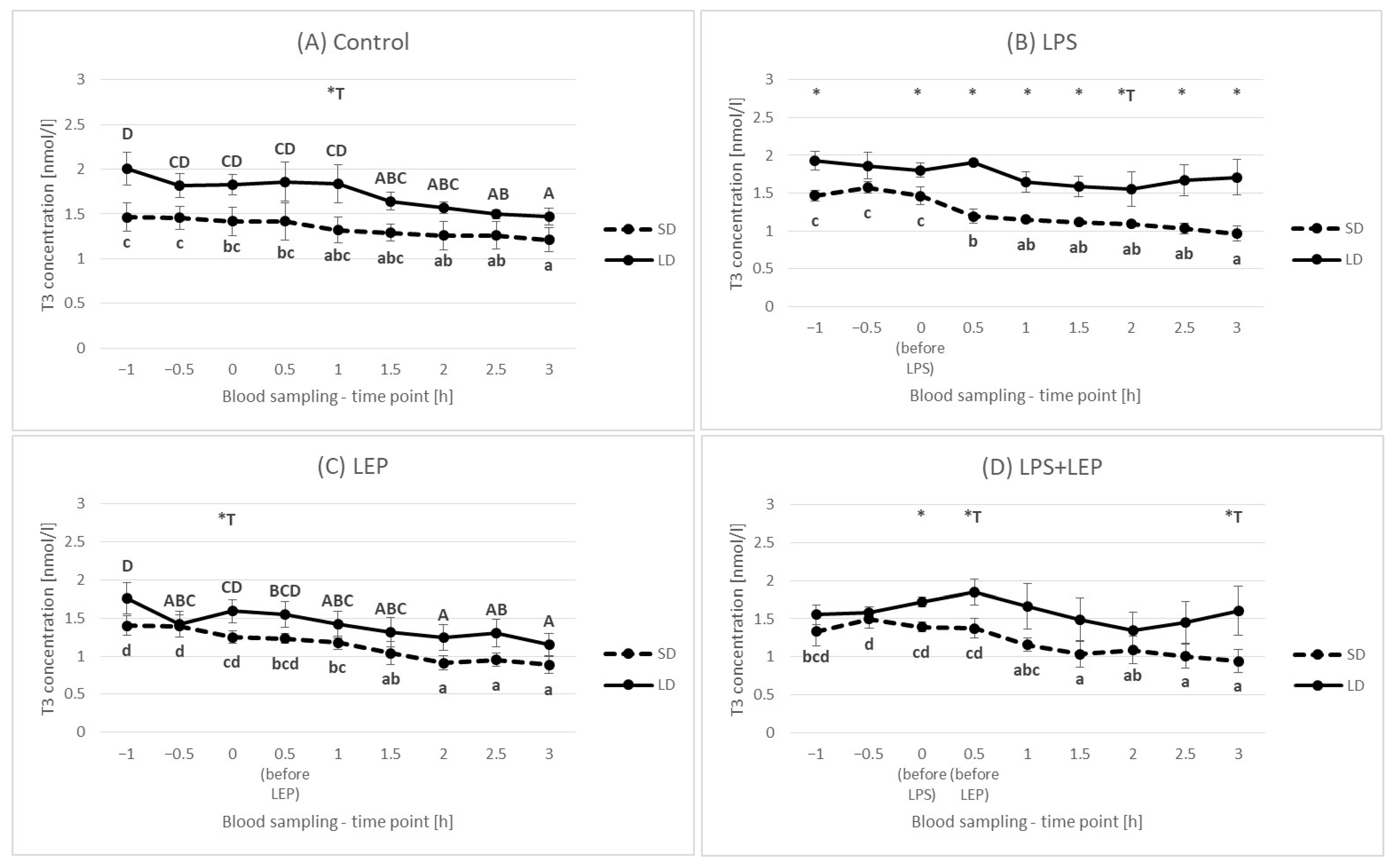
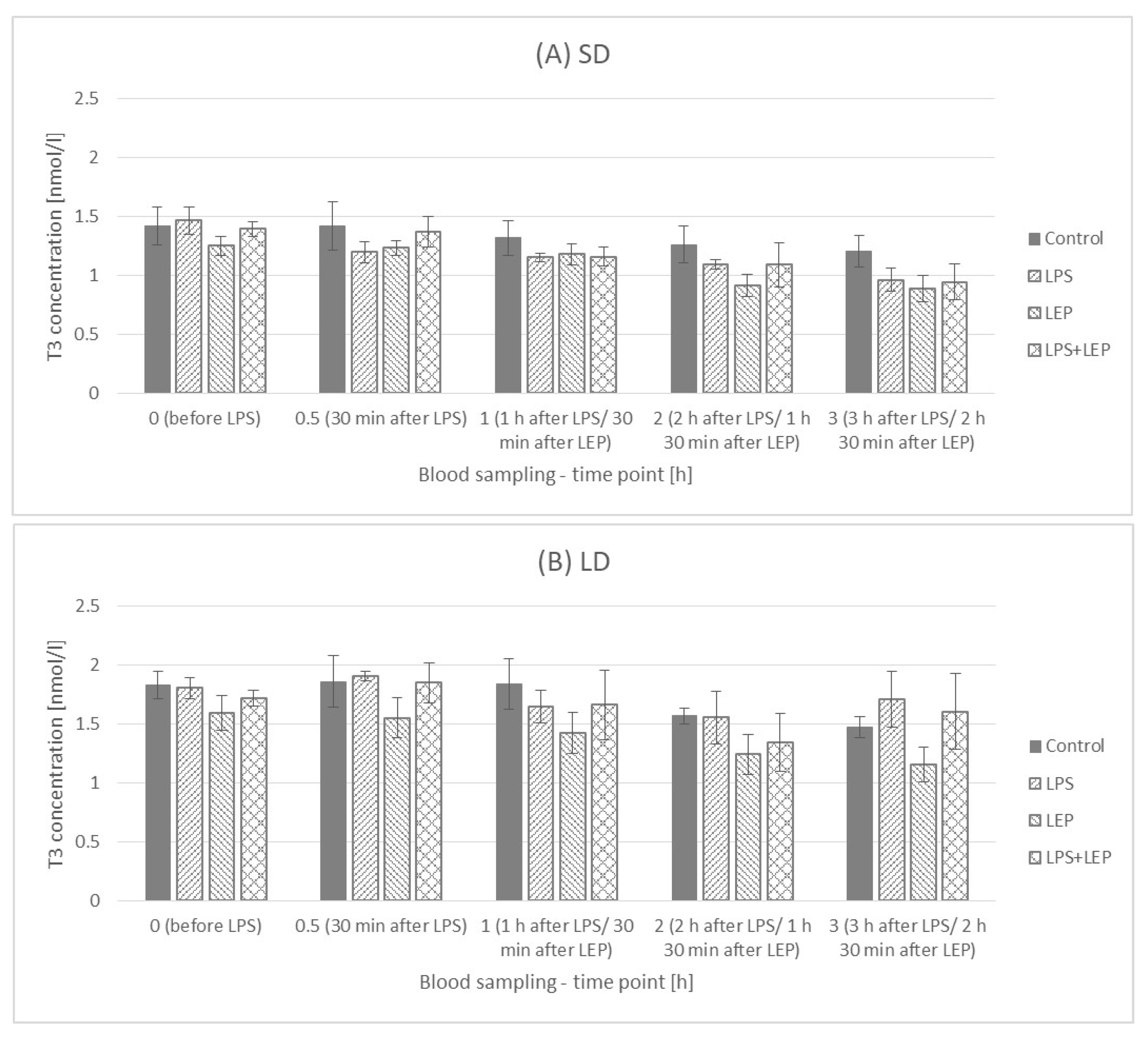
2.5. Triiodothyronine (T3) Plasma Concentration
3. Discussion
3.1. Leptin Plasma Concentration
3.2. Cortisol Plasma Concentration
3.3. Insulin Plasma Concentration
3.4. Thyroid Hormones Plasma Concentrations
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Blood Hormones Concentration Assay
4.2.1. Leptin
4.2.2. Insulin
4.2.3. Thyroid Hormones
4.2.4. Cortisol
4.3. Statistical Analysis
- Repeated measured analysis of variance (ANOVA) followed by a post-hoc Fisher’s test, with the animal as a dependent factor for time point (nine time points: −1 h, −0.5 h, 0 h, 0.5 h, 1 h, 1.5 h, 2 h, 2.5 h and 3 h) comparisons for each experimental group separately (n = 6 for each time point, as there were six animals in each group);
- Student’s t-test for the comparison between groups treated with the same factor (C, LPS, LEP and LPS + LEP) but in different seasons (SD vs. LD) for each time point separately (n = 6);
- Student’s t-test for a general comparison between seasons (SD vs. LD) at a 0 h time point—after a one-hour adaptation but before any factor injection (n = 24, as there were 24 animals in each season);
- Two-way ANOVA, with LPS and leptin injections as the main factors, followed by a post-hoc Fisher’s test for a comparison between the experimental groups (C vs. LPS vs. LEP vs. LPS + LEP) at selected time points (0 h, 0.5 h, 1 h, 2 h and 3 h) for each season separately (n = 6).
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Faggioni, R.; Fantuzzi, G.; Fuller, J.; Dinarello, C.A.; Feingold, K.R.; Grunfeld, C. IL-1 Beta Mediates Leptin Induction during Inflammation. Am. J. Physiol. 1998, 274, R204–R208. [Google Scholar] [PubMed]
- Francis, J.; MohanKumar, S.M.; MohanKumar, P.S. Correlations of Norepinephrine Release in the Paraventricular Nucleus with Plasma Corticosterone and Leptin after Systemic Lipopolysaccharide: Blockade by Soluble IL-1 Receptor. Brain Res. 2000, 867, 180–187. [Google Scholar] [CrossRef]
- Wang, Z.; Nakayama, T. Inflammation, a Link between Obesity and Cardiovascular Disease. Mediat. Inflamm. 2010, 2010, 535918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fain, J.N.; Leffler, C.W.; Cowan, G.S.M.; Buffington, C.; Pouncey, L.; Bahouth, S.W. Stimulation of Leptin Release by Arachidonic Acid and Prostaglandin E2 in Adipose Tissue from Obese Humans. Metabolism 2001, 50, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Sachot, C.; Poole, S.; Luheshi, G.N. Circulating Leptin Mediates Lipopolysaccharide-Induced Anorexia and Fever in Rats: Leptin, a Circulating Mediator of Inflammation. J. Physiol. 2004, 561, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Harden, L.M.; du Plessis, I.; Poole, S.; Laburn, H.P. Interleukin-6 and Leptin Mediate Lipopolysaccharide-Induced Fever and Sickness Behavior. Physiol. Behav. 2006, 89, 146–155. [Google Scholar] [CrossRef]
- Faggioni, R.; Fuller, J.; Moser, A.; Feingold, K.R.; Grunfeld, C. LPS-Induced Anorexia in Leptin-Deficient (Ob/Ob) and Leptin Receptor-Deficient (Db/Db) Mice. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1997, 273, R181–R186. [Google Scholar] [CrossRef]
- Kim, Y.-W.; Kim, K.-H.; Ahn, D.-K.; Kim, H.-S.; Kim, J.-Y.; Lee, D.C.; Park, S.-Y. Time-Course Changes of Hormones and Cytokines by Lipopolysaccharide and Its Relation with Anorexia. J. Physiol. Sci. 2007, 57, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Soliman, M.; Abdelhady, S.; Fattouh, I.; Ishioka, K.; Kitamura, H.; Kimura, K.; Saito, M. No Alteration in Serum Leptin Levels during Acute Endotoxemia in Sheep. J. Vet. Med. Sci. 2001, 63, 1143–1145. [Google Scholar] [CrossRef] [Green Version]
- Skipor, J.; Misztal, T.; Kaczmarek, M.M. Independent Changes of Thyroid Hormones in Blood Plasma and Cerebrospinal Fluid after Melatonin Treatment in Ewes. Theriogenology 2010, 74, 236–245. [Google Scholar] [CrossRef]
- Marie, M.; Findlay, P.; Thomas, L.; Adam, C. Daily Patterns of Plasma Leptin in Sheep: Effects of Photoperiod and Food Intake. J. Endocrinol. 2001, 170, 277–286. [Google Scholar] [CrossRef] [Green Version]
- Friedman, J.M.; Halaas, J.L. Leptin and the Regulation of Body Weight in Mammals. Nature 1998, 395, 763–770. [Google Scholar] [CrossRef] [PubMed]
- Adam, C.; Archer, Z.; Miller, D. Leptin Actions on the Reproductive Neuroendocrine Axis in Sheep. Reprod. (Camb. Engl.) Suppl. 2003, 61, 283–297. [Google Scholar] [CrossRef]
- Chilliard, Y.; Delavaud, C.; Bonnet, M. Leptin Expression in Ruminants: Nutritional and Physiological Regulations in Relation with Energy Metabolism. Domest. Anim. Endocrinol. 2005, 29, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Zieba, D.A.; Szczesna, M.; Klocek-Gorka, B.; Williams, G.L. Leptin as a Nutritional Signal Regulating Appetite and Reproductive Processes in Seasonally-Breeding Ruminants. J. Physiol. Pharmacol. 2008, 59 (Suppl. S9), 7–18. [Google Scholar]
- Zieba, D.A.; Szczesna, M.; Klocek-Gorka, B.; Molik, E.; Misztal, T.; Williams, G.L.; Romanowicz, K.; Stepien, E.; Keisler, D.H.; Murawski, M. Seasonal Effects of Central Leptin Infusion on Secretion of Melatonin and Prolactin and on SOCS-3 Gene Expression in Ewes. J. Endocrinol. 2008, 198, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Szczesna, M.; Zieba, D.A.; Klocek-Gorka, B.; Misztal, T.; Stepien, E. Seasonal Effects of Central Leptin Infusion and Prolactin Treatment on Pituitary SOCS-3 Gene Expression in Ewes. J. Endocrinol. 2011, 208, 81–88. [Google Scholar] [CrossRef] [Green Version]
- Krawczyńska, A.; Antushevich, H.; Bochenek, J.; Wojtulewicz, K.; Pawlina, B.; Herman, A.; Zięba, D. Photoperiodic Conditions as a Factor Modulating Leptin Influenceon Pro-Inflammatory Cytokinesand Their Receptors Gene Expression in Ewe’s Aorta. J. Anim. Feed Sci. 2019, 28, 128–137. [Google Scholar] [CrossRef]
- Krawczyńska, A.; Herman, A.P.; Antushevich, H.; Bochenek, J.; Wojtulewicz, K.; Zięba, D.A. The Influence of Photoperiod on the Action of Exogenous Leptin on Gene Expression of Proinflammatory Cytokines and Their Receptors in the Thoracic Perivascular Adipose Tissue (PVAT) in Ewes. Mediat. Inflamm. 2019, 2019, 7129476. [Google Scholar] [CrossRef]
- Szczepkowska, A.; Kowalewska, M.; Krawczyńska, A.; Herman, A.P.; Skipor, J. Photoperiod Affects Leptin Action on the Choroid Plexus in Ewes Challenged with Lipopolysaccharide—Study on the MRNA Level. Int. J. Mol. Sci. 2020, 21, 7647. [Google Scholar] [CrossRef]
- Wójcik, M.; Herman, A.P.; Zieba, D.A.; Krawczyńska, A. The Impact of Photoperiod on the Leptin Sensitivity and Course of Inflammation in the Anterior Pituitary. Int. J. Mol. Sci. 2020, 21, 4153. [Google Scholar] [CrossRef] [PubMed]
- Morrison, A.E.; Fleming, S.; Levy, M.J. A Review of the Pathophysiology of Functional Hypothalamic Amenorrhoea in Women Subject to Psychological Stress, Disordered Eating, Excessive Exercise or a Combination of These Factors. Clin. Endocrinol. 2021, 95, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Phylactou, M.; Clarke, S.A.; Patel, B.; Baggaley, C.; Jayasena, C.N.; Kelsey, T.W.; Comninos, A.N.; Dhillo, W.S.; Abbara, A. Clinical and Biochemical Discriminants between Functional Hypothalamic Amenorrhoea (FHA) and Polycystic Ovary Syndrome (PCOS). Clin. Endocrinol. 2021, 95, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Qiao, J. Association of Insulin Resistance and Elevated Androgen Levels with Polycystic Ovarian Syndrome (PCOS): A Review of Literature. J. Healthc. Eng. 2022, 2022, 9240569. [Google Scholar] [CrossRef] [PubMed]
- Enkhbaatar, P.; Nelson, C.; Salsbury, J.R.; Carmical, J.R.; Torres, K.E.O.; Herndon, D.; Prough, D.S.; Luan, L.; Sherwood, E.R. Comparison of Gene Expression by Sheep and Human Blood Stimulated with the TLR4 Agonists Lipopolysaccharide and Monophosphoryl Lipid A. PLoS ONE 2015, 10, e0144345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warren, H.S.; Fitting, C.; Hoff, E.; Adib-Conquy, M.; Beasley-Topliffe, L.; Tesini, B.; Liang, X.; Valentine, C.; Hellman, J.; Hayden, D.; et al. Resilience to Bacterial Infection: Difference between Species Could Be Due to Proteins in Serum. J. Infect. Dis. 2010, 201, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Zieba, D.A.; Biernat, W.; Barć, J. Roles of Leptin and Resistin in Metabolism, Reproduction, and Leptin Resistance. Domest. Anim. Endocrinol. 2020, 73, 106472. [Google Scholar] [CrossRef]
- Soliman, M.; Ishioka, K.; Kimura, K.; Kushibiki, S.; Saito, M. Plasma Leptin Responses to Lipopolysaccharide and Tumor Necrosis Factor Alpha in Cows. Jpn. J. Vet. Res. 2002, 50, 107–114. [Google Scholar]
- Barrichon, M.; Hadi, T.; Wendremaire, M.; Ptasinski, C.; Seigneuric, R.; Marcion, G.; Delignette, M.; Marchet, J.; Dumas, M.; Sagot, P.; et al. Dose-Dependent Biphasic Leptin-Induced Proliferation Is Caused by Non-Specific IL-6/NF-ΚB Pathway Activation in Human Myometrial Cells: Leptin Induces Human Myometrial Cell Proliferation. Br. J. Pharmacol. 2015, 172, 2974–2990. [Google Scholar] [CrossRef] [Green Version]
- Herman, A.P.; Tomaszewska-Zaremba, D.; Kowalewska, M.; Szczepkowska, A.; Oleszkiewicz, M.; Krawczyńska, A.; Wójcik, M.; Antushevich, H.; Skipor, J. Neostigmine Attenuates Proinflammatory Cytokine Expression in Preoptic Area but Not Choroid Plexus during Lipopolysaccharide-Induced Systemic Inflammation. Mediat. Inflamm. 2018, 2018, 9150207. [Google Scholar] [CrossRef]
- Wojtulewicz, K.; Krawczyńska, A.; Tomaszewska-Zaremba, D.; Wójcik, M.; Herman, A.P. Effect of Acute and Prolonged Inflammation on the Gene Expression of Proinflammatory Cytokines and Their Receptors in the Anterior Pituitary Gland of Ewes. Int. J. Mol. Sci. 2020, 21, 6939. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.P.; Krawczyńska, A.; Bochenek, J.; Haziak, K.; Romanowicz, K.; Misztal, T.; Antushevich, H.; Herman, A.; Tomaszewska-Zaremba, D. The Effect of Rivastigmine on the LPS-Induced Suppression of GnRH/LH Secretion during the Follicular Phase of the Estrous Cycle in Ewes. Anim. Reprod. Sci. 2013, 138, 203–212. [Google Scholar] [CrossRef]
- Herman, A.; Romanowicz, K.; Tomaszewska-Zaremba, D. Effect of LPS on Reproductive System at the Level of the Pituitary of Anestrous Ewes: Effect of LPS on Reproductive System at the Pituitary Level. Reprod. Domest. Anim. 2010, 45, e351–e359. [Google Scholar] [CrossRef] [PubMed]
- Persson, R.; Garde, A.H.; Hansen, Å.M.; Österberg, K.; Larsson, B.; Ørbæk, P.; Karlson, B. Seasonal Variation in Human Salivary Cortisol Concentration. Chronobiol. Int. 2008, 25, 923–937. [Google Scholar] [CrossRef] [PubMed]
- Chergui, N.; Mormede, P.; Foury, A.; Khammar, F.; Amirat, Z. Seasonal Effects on Plasma Cortisol Concentrations in the Bedouin Buck: Circadian Studies and Response to ACTH. Animal 2017, 11, 445–451. [Google Scholar] [CrossRef]
- Lightman, S.L.; Birnie, M.T.; Conway-Campbell, B.L. Dynamics of ACTH and Cortisol Secretion and Implications for Disease. Endocr. Rev. 2020, 41, bnaa002. [Google Scholar] [CrossRef] [Green Version]
- Chrousos, G.P. The Hypothalamic–Pituitary–Adrenal Axis and Immune-Mediated Inflammation. N. Engl. J. Med. 1995, 332, 1351–1363. [Google Scholar] [CrossRef]
- Cadaret, C.N.; Abebe, M.D.; Barnes, T.L.; Posont, R.J.; Yates, D.T. Lipopolysaccharide Endotoxin Injections Elevated Salivary TNFα and Corneal Temperatures and Induced Dynamic Changes in Circulating Leukocytes, Inflammatory Cytokines, and Metabolic Indicators in Wether Lambs. J. Anim. Sci. 2021, 99, skab120. [Google Scholar] [CrossRef]
- Li, L.; Messina, J.L. Acute Insulin Resistance Following Injury. Trends Endocrinol. Metab. 2009, 20, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Brunkhorst, F.M.; Engel, C.; Bloos, F.; Meier-Hellmann, A.; Ragaller, M.; Weiler, N.; Moerer, O.; Gruendling, M.; Oppert, M.; Grond, S.; et al. Intensive Insulin Therapy and Pentastarch Resuscitation in Severe Sepsis. N. Engl. J. Med. 2008, 358, 125–139. [Google Scholar] [CrossRef] [Green Version]
- Chalmeh, A.; Badiei, K.; Pourjafar, M.; Nazifi, S. Modulation of Inflammatory Responses Following Insulin Therapy in Experimentally Bolus Intravenous Escherichia Coli Lipopolysaccharide Serotype O55:B5 Induced Endotoxemia in Iranian Fat-Tailed Sheep. Small Rumin. Res. 2013, 113, 283–289. [Google Scholar] [CrossRef]
- Ghasemi-Panahi, B.; Rafat, S.; Ebrahimi, M.; Akbarzadeh, M.; Hajializadeh Valiloo, R. New Technique for Activating Reproductive System during Non-Breeding Season in Ghezel Ewes. Iran. J. Appl. Anim. Sci. 2016, 6, 357–361. [Google Scholar]
- Zieba, D.A.; Amstalden, M.; Maciel, M.N.; Keisler, D.H.; Raver, N.; Gertler, A.; Williams, G.L. Divergent Effects of Leptin on Luteinizing Hormone and Insulin Secretion Are Dose Dependent. Exp. Biol. Med. 2003, 228, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Seufert, J. Leptin Suppression of Insulin Secretion and Gene Expression in Human Pancreatic Islets: Implications for the Development of Adipogenic Diabetes Mellitus. J. Clin. Endocrinol. Metab. 1999, 84, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Seufert, J.; Kieffer, T.J.; Habener, J.F. Leptin Inhibits Insulin Gene Transcription and Reverses Hyperinsulinemia in Leptin-Deficient Ob/Ob Mice. Proc. Natl. Acad. Sci. USA 1999, 96, 674–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, R.N.; Wang, Z.L.; Wang, R.M.; Hurley, J.D.; Smith, D.M.; Ghatei, M.A.; Withers, D.J.; Gardiner, J.V.; Bailey, C.J.; Bloom, S.R. Leptin Rapidly Suppresses Insulin Release from Insulinoma Cells, Rat and Human Islets and, in Vivo, in Mice. J. Clin. Investig. 1997, 100, 2729–2736. [Google Scholar] [CrossRef]
- Marroquí, L.; Gonzalez, A.; Ñeco, P.; Caballero-Garrido, E.; Vieira, E.; Ripoll, C.; Nadal, A.; Quesada, I. Role of Leptin in the Pancreatic β-Cell: Effects and Signaling Pathways. J. Mol. Endocrinol. 2012, 49, R9–R17. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, V.; Shyng, S.-L. Leptin-Induced Trafficking of KATP Channels: A Mechanism to Regulate Pancreatic β-Cell Excitability and Insulin Secretion. Int. J. Mol. Sci. 2019, 20, 2660. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, A.; Murakami, T.; Otani, S.; Kuwajima, M.; Shima, K. Leptin Affects Pancreatic Endocrine Functions through the Sympathetic Nervous System 1. Endocrinology 1998, 139, 3863–3870. [Google Scholar] [CrossRef]
- Vives-Pi, M.; Somoza, N.; Fernández-Alvarez, J.; Vargas, F.; Caro, P.; Alba, A.; Gomis, R.; Labeta, M.O.; Pujol-Borrell, R. Evidence of Expression of Endotoxin Receptors CD14, Toll-like Receptors TLR4 and TLR2 and Associated Molecule MD-2 and of Sensitivity to Endotoxin (LPS) in Islet Beta Cells. Clin. Exp. Immunol. 2003, 133, 208–218. [Google Scholar] [CrossRef]
- Jiang, M.; He, J.; Sun, Y.; Dong, X.; Yao, J.; Gu, H.; Liu, L. Leptin Induced TLR4 Expression via the JAK2-STAT3 Pathway in Obesity-Related Osteoarthritis. Oxidative Med. Cell. Longev. 2021, 2021, 7385160. [Google Scholar] [CrossRef] [PubMed]
- Dardente, H.; Hazlerigg, D.G.; Ebling, F.J.P. Thyroid Hormone and Seasonal Rhythmicity. Front. Endocrinol. 2014, 5, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebling, F.J.P. On the Value of Seasonal Mammals for Identifying Mechanisms Underlying the Control of Food Intake and Body Weight. Horm. Behav. 2014, 66, 56–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fekete, C.; Gereben, B.; Doleschall, M.; Harney, J.W.; Dora, J.M.; Bianco, A.C.; Sarkar, S.; Liposits, Z.; Rand, W.; Emerson, C.; et al. Lipopolysaccharide Induces Type 2 Iodothyronine Deiodinase in the Mediobasal Hypothalamus: Implications for the Nonthyroidal Illness Syndrome. Endocrinology 2004, 145, 1649–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klocek-Gorka, B.; Szczesna, M.; Molik, E.; Zieba, D.A. The Interactions of Season, Leptin and Melatonin Levels with Thyroid Hormone Secretion, Using an in Vitro Approach. Small Rumin. Res. 2010, 91, 231–235. [Google Scholar] [CrossRef]
- Stoney, P.N.; Rodrigues, D.; Helfer, G.; Khatib, T.; Ashton, A.; Hay, E.A.; Starr, R.; Kociszewska, D.; Morgan, P.; McCaffery, P. A Seasonal Switch in Histone Deacetylase Gene Expression in the Hypothalamus and Their Capacity to Modulate Nuclear Signaling Pathways. Brain Behav. Immun. 2017, 61, 340–352. [Google Scholar] [CrossRef] [Green Version]
- Aung, H.T.; Schroder, K.; Himes, S.R.; Brion, K.; Van Zuylen, W.; Trieu, A.; Suzuki, H.; Hayashizaki, Y.; Hume, D.A.; Sweet, M.J.; et al. LPS Regulates Proinflammatory Gene Expression in Macrophages by Altering Histone Deacetylase Expression. FASEB J. 2006, 20, 1315–1327. [Google Scholar] [CrossRef]
- Wu, C.; Li, A.; Hu, J.; Kang, J. Histone Deacetylase 2 Is Essential for LPS-Induced Inflammatory Responses in Macrophages. Immunol. Cell Biol. 2019, 97, 72–84. [Google Scholar] [CrossRef]
- Sun, L.-H.; Pi, D.-A.; Zhao, L.; Wang, X.-Y.; Zhu, L.-Y.; Qi, D.-S.; Liu, Y.-L. Response of Selenium and Selenogenome in Immune Tissues to LPS-Induced Inflammatory Reactions in Pigs. Biol. Trace Elem. Res. 2017, 177, 90–96. [Google Scholar] [CrossRef]
- Strzetelski, J.A.; Brzóska, F.; Kowalski, Z.M.; Osięgłowski, S. Zalecenia Żywieniowe Dla Przeżuwaczy i Tabele Wartości Pokarmowej Pasz (Feeding Recommendation for Ruminants and Feed Tables); National Research Institute of Animal Production: Krakow, Poland, 2014. [Google Scholar]
- Przybył, B.; Wójcik-Gładysz, A.; Gajewska, A.; Szlis, M. Brain-Derived Neurotrophic Factor (BDNF) Affects Somatotrophicaxis Activity in Sheep. J. Anim. Feed Sci. 2021, 30, 329–339. [Google Scholar] [CrossRef]
- Herman, A.P.; Tomaszewska-Zaremba, D. Effect of Endotoxin on the Expression of GnRH and GnRHR Genes in the Hypothalamus and Anterior Pituitary Gland of Anestrous Ewes. Anim. Reprod. Sci. 2010, 120, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Maciel, M.N.; Zieba, D.A.; Amstalden, M.; Keisler, D.H.; Neves, J.P.; Williams, G.L. Chronic Administration of Recombinant Ovine Leptin in Growing Beef Heifers: Effects on Secretion of LH, Metabolic Hormones, and Timing of Puberty1. J. Anim. Sci. 2004, 82, 2930–2936. [Google Scholar] [CrossRef] [PubMed]
- Delavaud, C.; Bocquier, F.; Chilliard, Y.; Keisler, D.; Gertler, A.; Kann, G. Plasma Leptin Determination in Ruminants: Effect of Nutritional Status and Body Fatness on Plasma Leptin Concentration Assessed by a Specific RIA in Sheep. J. Endocrinol. 2000, 165, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Catunda, A.G.V.; Lima, I.C.S.; Bandeira, G.C.; Gadelha, C.R.F.; Pereira, E.S.; Salmito-Vanderley, C.S.B.; Araújo, A.A.; Martins, G.A.; Campos, A.C.N. Blood Leptin, Insulin and Glucose Concentrations in Hair Sheep Raised in a Tropical Climate. Small Rumin. Res. 2013, 114, 272–279. [Google Scholar] [CrossRef] [Green Version]
- Bivolarski, B.; Vachkova, E.; Laleva, L.; Slavova, P.; Ivanov, I. Comparative Studies on Some Parameters of Innate Resistance and Metabolic Profile of Sheep and Their Offspring Depending on the Ration. Agric. Sci. Technol. 2011, 3, 103–106. [Google Scholar]
- Kokot, F.; Stupnicki, R. Metody Radioimmunologiczne i Radiokompetyncyjne Stosowane w Klinice, 2nd ed.; PZWL: Warszawa, Poland, 1985. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Group | Number of Animals | Experimental Factor | LPS Dose [ng/kg of Body Mass] | Leptin Dose [μg /kg of Body Mass] |
|---|---|---|---|---|---|
| Experiment 1—Short-Day Season | |||||
| 1. | Control | 6 | NaCl | 0 | 0 |
| 2. | LPS | 6 | LPS | 400 | 0 |
| 3. | LEP | 6 | Leptin | 0 | 20 |
| 4. | LPS + LEP | 6 | LPS + Leptin injected 30 min after LPS | 400 | 20 |
| Experiment 2—Long-Day Season | |||||
| 5. | Control | 6 | NaCl | 0 | 0 |
| 6. | LPS | 6 | LPS | 400 | 0 |
| 7. | LEP | 6 | Leptin | 0 | 20 |
| 8. | LPS + LEP | 6 | LPS + Leptin injected 30 min after LPS | 400 | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krawczyńska, A.; Herman, A.P.; Antushevich, H.; Bochenek, J.; Wojtulewicz, K.; Zieba, D.A. The Effect of Leptin on the Blood Hormonal Profile (Cortisol, Insulin, Thyroid Hormones) of the Ewe in Acute Inflammation in Two Different Photoperiodical Conditions. Int. J. Mol. Sci. 2022, 23, 8109. https://doi.org/10.3390/ijms23158109
Krawczyńska A, Herman AP, Antushevich H, Bochenek J, Wojtulewicz K, Zieba DA. The Effect of Leptin on the Blood Hormonal Profile (Cortisol, Insulin, Thyroid Hormones) of the Ewe in Acute Inflammation in Two Different Photoperiodical Conditions. International Journal of Molecular Sciences. 2022; 23(15):8109. https://doi.org/10.3390/ijms23158109
Chicago/Turabian StyleKrawczyńska, Agata, Andrzej Przemysław Herman, Hanna Antushevich, Joanna Bochenek, Karolina Wojtulewicz, and Dorota Anna Zieba. 2022. "The Effect of Leptin on the Blood Hormonal Profile (Cortisol, Insulin, Thyroid Hormones) of the Ewe in Acute Inflammation in Two Different Photoperiodical Conditions" International Journal of Molecular Sciences 23, no. 15: 8109. https://doi.org/10.3390/ijms23158109