
Major Histocompatibility Complex Class I Chain-Related α (MICA) STR Polymorphisms in COVID-19 Patients

 , and
, and
Abstract
:1. Introduction
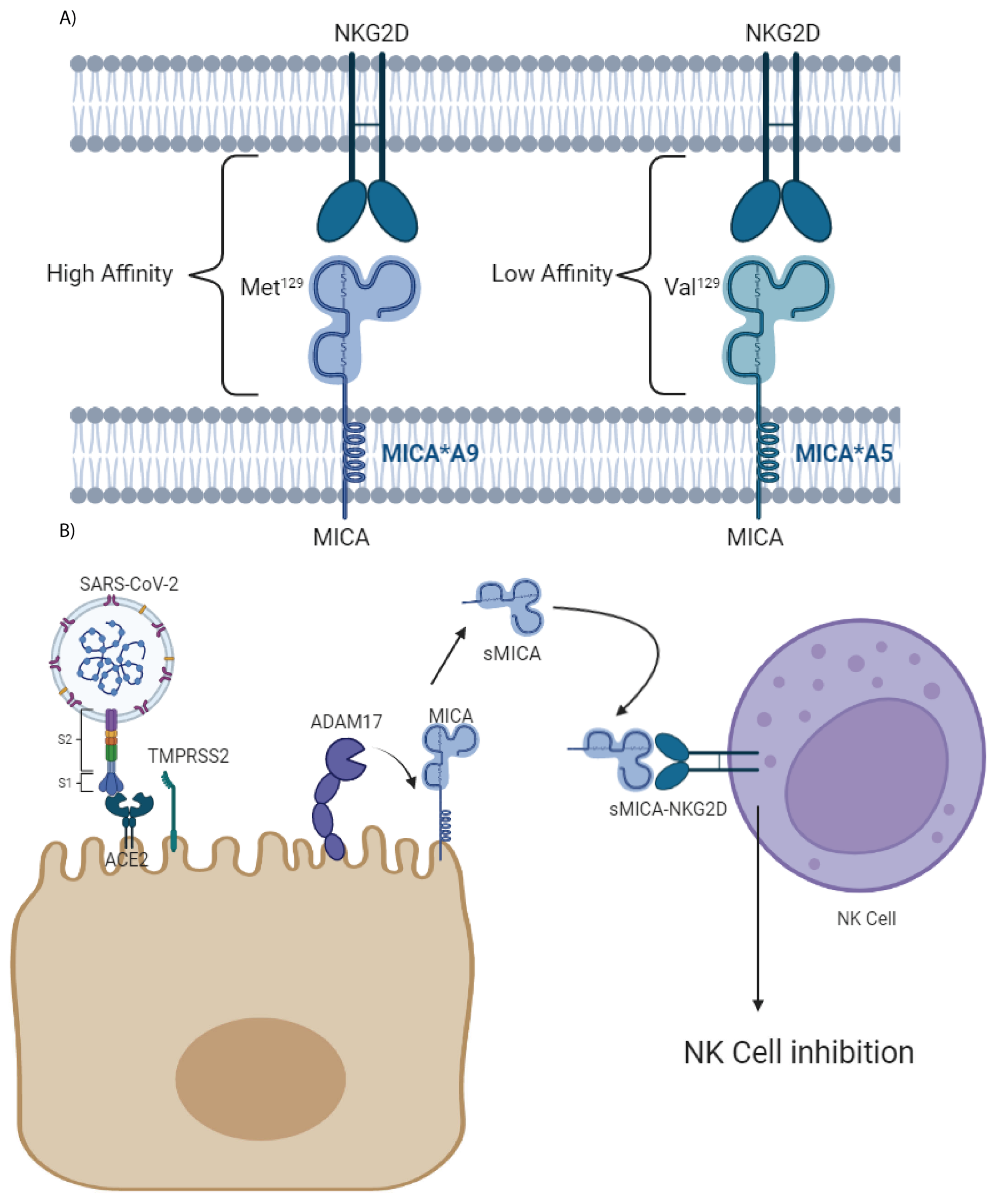
2. Results
3. Discussion
4. Materials and Methods
4.1. Samples
- Asymptomatic: 33 patients (23 women and 10 men); mean age of 42.6 years (26–63).
- Symptomatic:
- a.
- Moderate patients: 344 patients (158 women and 186 men); mean age of 63.6 years (25–98).
- b.
- Severe patients: 106 patients (38 women and 67 men); mean age of 61.8 years (26–86).
- (a)
- Asymptomatic: Individuals who test positive for SARS-CoV-2 using a virologic test but who have no symptoms that are consistent with COVID-19 [34].
- (b)
- Moderate Illness: Individuals who show evidence of lower respiratory disease during clinical assessment or imaging and who have an oxygen saturation (SpO2) ≥ 94% on room air at sea level [34].
- (c)
- Severe Illness: Individuals who have SpO2 < 94% on room air at sea level, a ratio of arterial partial pressure of oxygen to fraction of inspired oxygen (PaO2/FiO2) < 300 mm Hg, a respiratory rate > 30 breaths/min, or lung infiltrates > 50% [34].
4.2. Population Reference Group
4.3. DNA Extraction
4.4. HLA-B and MICA Genotyping
4.5. PCR Diagnosis of SARS-CoV-2 Infection
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goto, K.; Kato, N. MICA SNPs and the NKG2D system in virus-induced HCC. J. Gastroenterol. 2015, 50, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Waggoner, S.N.; Reighard, S.D.; Gyurova, I.E.; Cranert, S.A.; Mahl, S.E.; Karmele, E.P.; McNally, J.P.; Moran, M.T.; Brooks, T.R.; Yaqoob, F.; et al. Roles of natural killer cells in antiviral immunity. Curr. Opin. Virol. 2016, 16, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutiérrez-Bautista, J.F.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; Jiménez, P.; Garrido, F.; Anderson, P.; Ruiz-Cabello, F.; López-Ruz, M.Á. Negative Clinical Evolution in COVID-19 Patients Is Frequently Accompanied with an Increased Proportion of Undifferentiated Th Cells and a Strong Underrepresentation of the Th1 Subset. Front. Immunol. 2020, 11, 596553. [Google Scholar] [CrossRef] [PubMed]
- Marcus, A.; Gowen, B.G.; Thompson, T.W.; Iannello, A.; Ardolino, M.; Deng, W.; Wang, L.; Shifrin, N.; Raulet, D.H. Recognition of tumors by the innate immune system and natural killer cells. Adv. Immunol. 2014, 122, 91–128. [Google Scholar] [CrossRef] [Green Version]
- Shabani, Z.; Bagheri, M.; Zare-Bidaki, M.; Hassanshahi, G.; Arababadi, M.K.; Mohammadi Nejad, M.; Kennedy, D. NK cells in hepatitis B virus infection: A potent target for immunotherapy. Arch. Virol. 2014, 159, 1555–1565. [Google Scholar] [CrossRef]
- Eagle, R.A.; Traherne, J.A.; Ashiru, O.; Wills, M.R.; Trowsdale, J. Regulation of NKG2D ligand gene expression. Hum. Immunol. 2006, 67, 159–169. [Google Scholar] [CrossRef]
- Isernhagen, A.; Malzahn, D.; Bickeböller, H.; Dressel, R. Impact of the MICA-129Met/Val Dimorphism on NKG2D-Mediated Biological Functions and Disease Risks. Front. Immunol. 2016, 7, 588. [Google Scholar] [CrossRef] [Green Version]
- Jarduli, L.R.; Sell, A.M.; Reis, P.G.; Sippert, E.Â.; Ayo, C.M.; Mazini, P.S.; Alves, H.V.; Teixeira, J.J.V.; Visentainer, J.E.L. Role of HLA, KIR, MICA, and cytokines genes in leprosy. Biomed. Res. Int. 2013, 2013, 989837. [Google Scholar] [CrossRef] [Green Version]
- Ayo, C.M.; Bestetti, R.B.; de Campos Junior, E.; Ronchi, L.S.; Borim, A.A.; Brandão, C.C.; de Matttos, L.C. MICA and KIR: Immunogenetic Factors Influencing Left Ventricular Systolic Dysfunction and Digestive Clinical Form of Chronic Chagas Disease. Front. Immunol. 2021, 12, 714766. [Google Scholar] [CrossRef]
- Choy, M.-K.; Phipps, M.E. MICA polymorphism: Biology and importance in immunity and disease. Trends Mol. Med. 2010, 16, 97–106. [Google Scholar] [CrossRef]
- Suemizu, H.; Radosavljevic, M.; Kimura, M.; Sadahiro, S.; Yoshimura, S.; Bahram, S.; Inoko, H. A basolateral sorting motif in the MICA cytoplasmic tail. Proc. Natl. Acad. Sci. USA 2002, 99, 2971–2976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Messina, L.; Reyburn, H.T.; Valés-Gómez, M. Human NKG2D-ligands: Cell biology strategies to ensure immune recognition. Front. Immunol. 2012, 3, 299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hüe, S.; Mention, J.-J.; Monteiro, R.C.; Zhang, S.; Cellier, C.; Schmitz, J.; Verkarre, V.; Fodil, N.; Bahram, S.; Cerf-Bensussan, N.; et al. A direct role for NKG2D/MICA interaction in villous atrophy during celiac disease. Immunity 2004, 21, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Witkowski, M.; Tizian, C.; Ferreira-Gomes, M.; Niemeyer, D.; Jones, T.C.; Heinrich, F.; Frischbutter, S.; Angermair, S.; Hohnstein, T.; Mattiola, I.; et al. Untimely TGFβ responses in COVID-19 limit antiviral functions of NK cells. Nature 2021, 600, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Castelli, E.C.; de Castro, M.V.; Naslavsky, M.S.; Scliar, M.O.; Silva, N.S.B.; Andrade, H.S.; Souza, A.S.; Pereira, R.N.; Castro, C.F.B.; Mendes-Junior, C.T.; et al. MHC Variants Associated With Symptomatic Versus Asymptomatic SARS-CoV-2 Infection in Highly Exposed Individuals. Front. Immunol. 2021, 12, 742881. [Google Scholar] [CrossRef]
- Gutiérrez-Bautista, J.F.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; López-Ruz, M.Á.; Martín-Casares, A.M.; Fernández-Rubiales, A.; Anderson, P.; Garrido, F.; Ruiz-Cabello, F.; López-Nevot, M.Á. Study of HLA-A, -B, -C, -DRB1 and -DQB1 polymorphisms in COVID-19 patients. J. Microbiol. Immunol. Infect. 2021, 55, 421–427. [Google Scholar] [CrossRef]
- Jarduli, L.R.; Alves, H.V.; de Souza, V.H.; Uaska Sartori, P.V.; Fava, V.M.; de Souza, F.C.; Marcos, E.V.C.; Pereira, A.C.; Dias-Baptista, I.M.F.; Virmond, M.d.C.L.; et al. Association of MICA and HLA-B alleles with leprosy in two endemic populations in Brazil. Int. J. Immunogenet. 2021, 48, 25–35. [Google Scholar] [CrossRef]
- Tosh, K.; Ravikumar, M.; Bell, J.T.; Meisner, S.; Hill, A.V.S.; Pitchappan, R. Variation in MICA and MICB genes and enhanced susceptibility to paucibacillary leprosy in South India. Hum. Mol. Genet. 2006, 15, 2880–2887. [Google Scholar] [CrossRef] [Green Version]
- Steinle, A.; Li, P.; Morris, D.L.; Groh, V.; Lanier, L.L.; Strong, R.K.; Spies, T. Interactions of human NKG2D with its ligands MICA, MICB, and homologs of the mouse RAE-1 protein family. Immunogenetics 2001, 53, 279–287. [Google Scholar] [CrossRef]
- Kopp, R.; Glas, J.; Lau-Werner, U.; Albert, E.D.; Weiss, E.H. Association of MICA-TM and MICB C1_2_A microsatellite polymorphisms with tumor progression in patients with colorectal cancer. J. Clin. Immunol. 2009, 29, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Bouayad, A. Features of HLA class I expression and its clinical relevance in SARS-CoV-2: What do we know so far? Rev. Med. Virol. 2021, 31, e2236. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.; Boggs, D.A.; Uko, G.; Essiet, A.; Inyama, M.; Banjoko, B.; Adewole, T.; Ding, W.-Z.; Mohseni, M.; Fritz, R.; et al. MICA, HLA-B haplotypic variation in five population groups of sub-Saharan African ancestry. Genes Immun. 2003, 4, 500–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, W.; Cai, J.H.; Wang, F.; Li, L.X. MICA polymorphism in a northern Chinese Han population: The identification of a new MICA allele, MICA*059. Hum. Immunol. 2010, 71, 423–427. [Google Scholar] [CrossRef]
- Li, M.; Guo, W.; Dong, Y.; Wang, X.; Dai, D.; Liu, X.; Wu, Y.; Li, M.; Zhang, W.; Zhou, H. Elevated exhaustion levels of NK and CD8+ T cells as indicators for progression and prognosis of COVID-19 disease. Front. Immunol. 2020, 11, 580237. [Google Scholar] [CrossRef]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yaqinuddin, A.; Kashir, J. Innate immunity in COVID-19 patients mediated by NKG2A receptors, and potential treatment using Monalizumab, Cholroquine, and antiviral agents. Med. Hypotheses 2020, 140, 109777. [Google Scholar] [CrossRef]
- Vilches, C.; Parham, P. KIR: Diverse, rapidly evolving receptors of innate and adaptive immunity. Annu. Rev. Immunol. 2002, 20, 217–251. [Google Scholar] [CrossRef]
- Bernal, E.; Gimeno, L.; Alcaraz, M.J.; Quadeer, A.A.; Moreno, M.; Martínez-Sánchez, M.V.; Campillo, J.A.; Gomez, J.M.; Pelaez, A.; García, E.; et al. Activating Killer-Cell Immunoglobulin-Like Receptors Are Associated with the Severity of Coronavirus Disease 2019. J. Infect. Dis. 2021, 224, 229–240. [Google Scholar] [CrossRef]
- Hajeer, A.; Jawdat, D.; Massadeh, S.; Aljawini, N.; Abedalthagafi, M.S.; Arabi, Y.M.; Alaamery, M. Association of KIR gene polymorphisms with COVID-19 disease. Clin. Immunol. 2022, 234, 108911. [Google Scholar] [CrossRef]
- Van Eeden, C.; Khan, L.; Osman, M.S.; Cohen Tervaert, J.W. Natural Killer Cell Dysfunction and Its Role in COVID-19. Int. J. Mol. Sci. 2020, 21, 6351. [Google Scholar] [CrossRef]
- Groh, V.; Rhinehart, R.; Randolph-Habecker, J.; Topp, M.S.; Riddell, S.R.; Spies, T. Costimulation of CD8alphabeta T cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat. Immunol. 2001, 2, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L. NKG2D Receptor and Its Ligands in Host Defense. Cancer Immunol. Res. 2015, 3, 575–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badrinath, S.; Dellacherie, M.O.; Li, A.; Zheng, S.; Zhang, X.; Sobral, M.; Pyrdol, J.W.; Smith, K.L.; Lu, Y.; Haag, S.; et al. A vaccine targeting resistant tumours by dual T cell plus NK cell attack. Nature 2022, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Coronavirus Disease 2019 (COVID-19) Treatment Guidelines; National Institutes of Health (US): Bethesda, MD, USA, 21 April 2021.
- Hodcroft, E.B.; Zuber, M.; Nadeau, S.; Vaughan, T.G.; Crawford, K.H.D.; Althaus, C.L.; Reichmuth, M.L.; Bowen, J.E.; Walls, A.C.; Davide Corti, D.; et al. Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. medRxiv 2021. [Google Scholar] [CrossRef]


{kind=link}
| Allele | PRG (2n = 1234) n (%) | COVID-19 Patients (2n = 892) n (%) | Asymptomatic Patients (2n = 66) n (%) | Moderate Patients (2n = 686) n (%) | Severe Patients (2n = 206) n (%) | P1 (Pc) | P2 (Pc) | P3 (Pc) | P4 (Pc) | P5 (Pc) | P6 (Pc) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| MICA*A4 | 192 (15.6) | 119 (13.3) | 11 (16.7) | 95 (13.8) | 24 (11.7) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5 | 140 (11.3) | 102 (11.4) | 6 (9.1) | 81 (11.8) | 21 (10.2) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5.1 | 310 (25.1) | 196 (22.0) | 19 (28.8) | 158 (23) | 38 (18.4) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A6 | 423 (34.3) | 309 (34.6) | 22 (33.3) | 226 (32.9) | 83 (40.3) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A9 | 169 (13.7) | 166 (18.6) | 8 (12.1) | 126 (18.4) | 40 (19.4) | 0.004 (0.025) | n.s | 0.007 (0.035) | n.s | n.s | n.s |
| Genotype | PRG (n = 617) n (%) | COVID-19 Patients (n = 446) n (%) | Asymptomatic Patients (n = 33) n (%) | Moderate Patients (n = 343) n (%) | Severe Patients (n = 103) n (%) | P1 (Pc) | P2 (Pc) | P3 (Pc) | P4 (Pc) | P5 (Pc) | P6 (Pc) |
| MICA*A4,*A4 | 12 (1.9) | 4 (0.9) | 1 (3) | 4 (1.2) | 0 (0) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A4,*A5 | 18 (2.9) | 16 (3.6) | 3 (9.1) | 14 (4.1) | 2 (1.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A4,*A5.1 | 58 (9.4) | 33 (7.4) | 2 (6.1) | 28 (8.2) | 5 (4.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A4,*A6 | 67 (10.8) | 43 (9.6) | 3 (9.1) | 29 (8.5) | 14 (13.6) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A4,*A9 | 26 (4.2) | 19 (4.3) | 1 (3) | 16 (4.7) | 3 (2.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5,*A5 | 16 (2.6) | 9 (2.0) | 0 (0) | 7 (2) | 2 (1.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5,*A51 | 22 (3.6) | 20 (4.5) | 0 (0) | 18 (5.2) | 2 (1.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5,*A6 | 43 (7) | 29 (6.5) | 2 (6.1) | 21 (6.1) | 8 (7.8) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A5,*A9 | 23 (3.7) | 19 (4.3) | 1 (3) | 14 (4.1) | 5 (4.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A51,*A51 | 41 (6.6) | 20 (4.5) | 3 (9.1) | 17 (5) | 3 (2.9) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A51,*A6 | 107 (17.3) | 64 (14.3) | 9 (27.3) | 50 (14.6) | 14 (13.6) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A51,*A9 | 41 (6.6) | 39 (8.7) | 2 (6.1) | 28 (8.2) | 11 (10.7) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A6,*A6 | 77 (12.5) | 63 (14.1) | 3 (9.1) | 45 (13.1) | 18 (17.5) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A6,*A9 | 52 (8.4) | 47 (10.5) | 2 (6.1) | 36 (10.5) | 11 (10.7) | n.s | n.s | n.s | n.s | n.s | n.s |
| MICA*A9,*A9 | 14 (2.3) | 21 (4.7) | 1 (3) | 16 (4.7) | 5 (4.9) | n.s | n.s | 0.041 (n.s) | n.s | n.s | n.s |
| PRG | Asymptomatic | Moderate Patients | Severe Patients | COVID-19 Patients | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HLA-B/MICA | F | % | F | % | F | % | F | % | F | % |
| *07/*A51 | 105 | 8.5 | 9 | 13.6 | 58 | 8.5 | 16 | 7.8 | 83 | 8.8 |
| *41/*A6 | 14 | 1.1 | 2 | 3 | 10 | 1.5 | 3 | 1.5 | 15 | 1.6 |
| *14/*A5 | 14 | 1.1 | 2 | 3 | 8 | 1.2 | 1 | 0.5 | 11 | 1.2 |
| *35/*A9 | 56 | 4.5 | 2 | 3 | 38 | 5.5 | 12 | 5.8 | 51 | 5.4 |
| *50/*A6 | 38 | 3.1 | 1 | 1.5 | 14 | 2 | 10 | 4.9 | 25 | 2.6 |
| *40/*A5 | 20 | 1.6 | 1 | 1.5 | 20 | 2.9 | 3 | 1.5 | 24 | 2.5 |
| *27/*A4 | 41 | 3.3 | 3 | 4.5 | 14 | 2 | 8 | 3.9 | 25 | 2.6 |
| *18/*A4 | 125 | 10.1 | 5 | 7.6 | 58 | 8.5 | 13 | 6.3 | 75 | 7.9 |
| *57/*A9 | 25 | 2 | 1 | 1.5 | 22 | 3.2 | 11 | 5.3 | 34 | 3.6 |
| *44/*A51 | 57 | 4.6 | 4 | 6.1 | 29 | 4.2 | 10 | 4.9 | 43 | 4.6 |
| *51/*A6 | 102 | 8.3 | 3 | 4.5 | 39 | 5.7 | 17 | 8.3 | 58 | 6.1 |
| *15/*A5 | 33 | 2.7 | 3 | 4.5 | 19 | 2.8 | 9 | 4.4 | 30 | 3.2 |
| *44/*A6 | 124 | 10 | 8 | 12.1 | 78 | 11.4 | 29 | 14.1 | 114 | 12.1 |
| *35/*A6 | 20 | 1.6 | 2 | 3 | 9 | 1.3 | 2 | 1 | 13 | 1.4 |
| *40/*A51 | 17 | 1.4 | 2 | 3 | 8 | 1.2 | 2 | 1 | 12 | 1.3 |
| *08/*A51 | 53 | 4.3 | 3 | 4.5 | 36 | 5.2 | 4 | 1.9 | 42 | 4.4 |
| *14/*A6 | 50 | 4.1 | 2 | 3 | 37 | 5.4 | 10 | 4.9 | 49 | 5.2 |
| *39/*A9 | 15 | 1.2 | 1 | 1.5 | 12 | 1.7 | 2 | 1 | 16 | 1.7 |
| *38/*A9 | 28 | 2.3 | 1 | 1.5 | 22 | 3.2 | 10 | 4.9 | 33 | 3.5 |
| *53/*A9 | 15 | 1.2 | 1 | 1.5 | 15 | 2.2 | 3 | 1.5 | 19 | 2 |
| *55/*A4 | 9 | 0.7 | 1 | 1.5 | 13 | 1.9 | 2 | 1 | 16 | 1.7 |
| *13/*A51 | 19 | 1.5 | 1 | 1.5 | 9 | 1.3 | 3 | 1.5 | 13 | 1.4 |
| *49/*A6 | 31 | 2.5 | 1 | 1.5 | 20 | 2.9 | 7 | 3.4 | 28 | 3 |
| *52/*A6 | 14 | 1.1 | 1 | 1.5 | 10 | 1.5 | 5 | 2.4 | 16 | 1.7 |
| *35/*A5 | 41 | 3.3 | 0 | 0 | 27 | 3.9 | 6 | 2.9 | 32 | 3.4 |
| *58/*A9 | 10 | 1.06 | 0 | 0 | 8 | 1.2 | 2 | 1 | 16 | 1.3 |
| Characteristics | |||
|---|---|---|---|
| Moderate Patients (n = 344) | Severe Patients (n = 106) | p | |
| Age | 63.6 | 61.8 | n.s |
| Female | 159 (46.2%) | 38 (35.8%) | n.s |
| Male | 185 (53.1%) | 68 (64.2%) | |
| UCI | 23 (6.7%) | 103 (97.2%) | 3.72 × 10−74 |
| No UCI | 321 (93.3%) | 3 (2.8%) | |
| Mechanic Ventilation | 17 (4.9%) | 84 (79.2%) | 1.03× 10−50 |
| No Mechanic Ventilation | 327 (95.1%) | 22 (20.8%) | |
| Deceased | 28 (8.1%) | 41 (38.7%) | 3.02 × 10−11 |
| Survivors | 316 (91.9%) | 65 (61.3%) | |
| Comorbidities | |||
| Hypertension | 145 (42.2%) | 52 (49.1%) | n.s |
| DM | 65 (18.9%) | 36 (34%) | 0.01 |
| CKD | 27 (7.8%) | 5 (4.7%) | n.s |
| CVD | 20 (5.8%) | 2 (1.9%) | n.s |
| Overweight/Obesity | 41 (11.9%) | 30 (28.3%) | 8 × 10−4 |
| MI | 20 (5.8%) | 6 (5.7%) | n.s |
| HF | 16 (4.7%) | 9 (8.5%) | n.s |
| COPD | 20 (4.8%) | 13 (12.3%) | n.s |
| Asthma | 21 (6.1%) | 8 (7.5%) | n.s |
| PAD | 9 (2.6%) | 2 (1.9%) | n.s |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gutiérrez-Bautista, J.F.; Martinez-Chamorro, A.; Rodriguez-Nicolas, A.; Rosales-Castillo, A.; Jiménez, P.; Anderson, P.; López-Ruz, M.Á.; López-Nevot, M.Á.; Ruiz-Cabello, F. Major Histocompatibility Complex Class I Chain-Related α (MICA) STR Polymorphisms in COVID-19 Patients. Int. J. Mol. Sci. 2022, 23, 6979. https://doi.org/10.3390/ijms23136979
Gutiérrez-Bautista JF, Martinez-Chamorro A, Rodriguez-Nicolas A, Rosales-Castillo A, Jiménez P, Anderson P, López-Ruz MÁ, López-Nevot MÁ, Ruiz-Cabello F. Major Histocompatibility Complex Class I Chain-Related α (MICA) STR Polymorphisms in COVID-19 Patients. International Journal of Molecular Sciences. 2022; 23(13):6979. https://doi.org/10.3390/ijms23136979
Chicago/Turabian StyleGutiérrez-Bautista, Juan Francisco, Alba Martinez-Chamorro, Antonio Rodriguez-Nicolas, Antonio Rosales-Castillo, Pilar Jiménez, Per Anderson, Miguel Ángel López-Ruz, Miguel Ángel López-Nevot, and Francisco Ruiz-Cabello. 2022. "Major Histocompatibility Complex Class I Chain-Related α (MICA) STR Polymorphisms in COVID-19 Patients" International Journal of Molecular Sciences 23, no. 13: 6979. https://doi.org/10.3390/ijms23136979