
Identification and Characterization of PSEUDO-RESPONSE REGULATOR (PRR) 1a and 1b Genes by CRISPR/Cas9-Targeted Mutagenesis in Chinese Cabbage (Brassica rapa L.)


 and
and
Abstract
:1. Introduction
2. Results
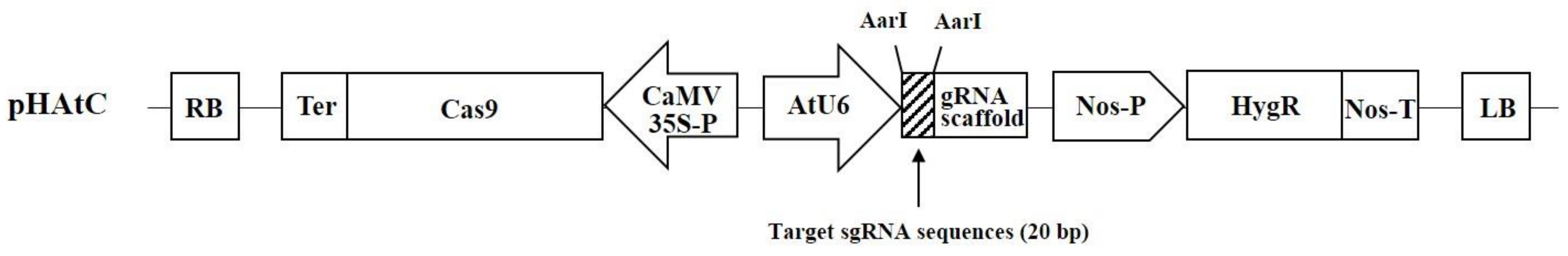
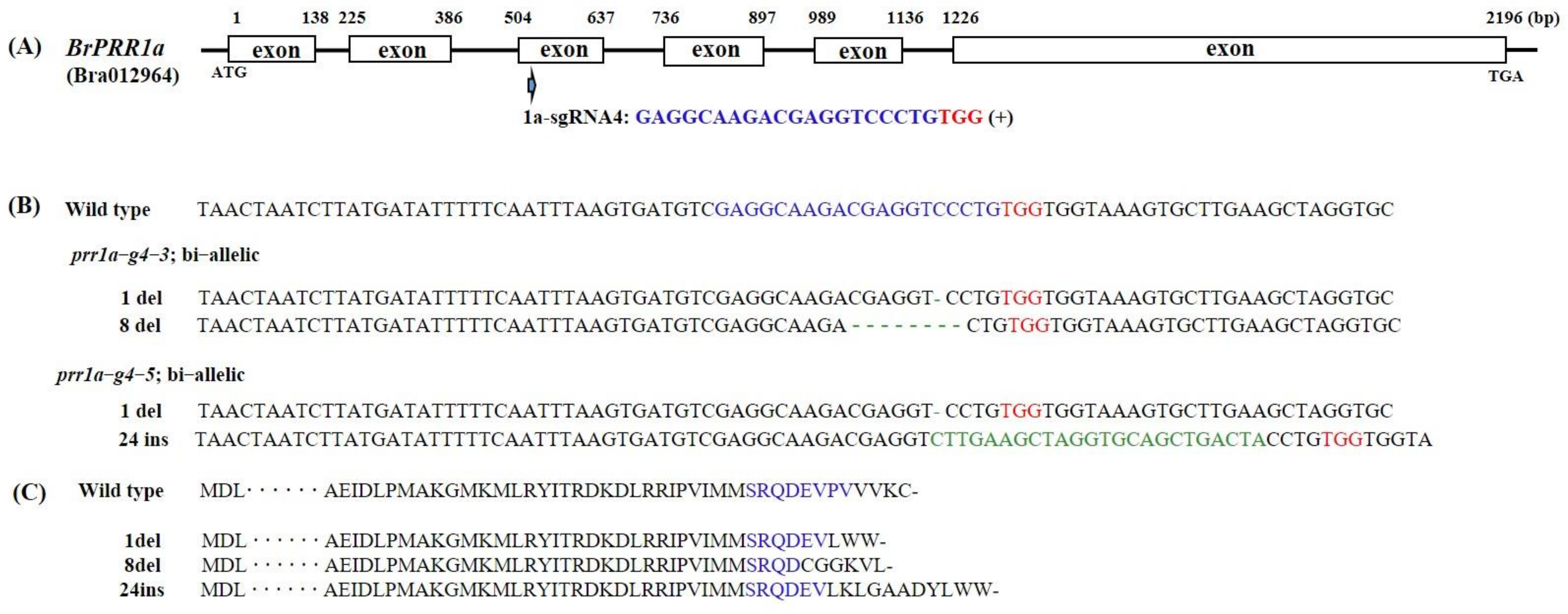
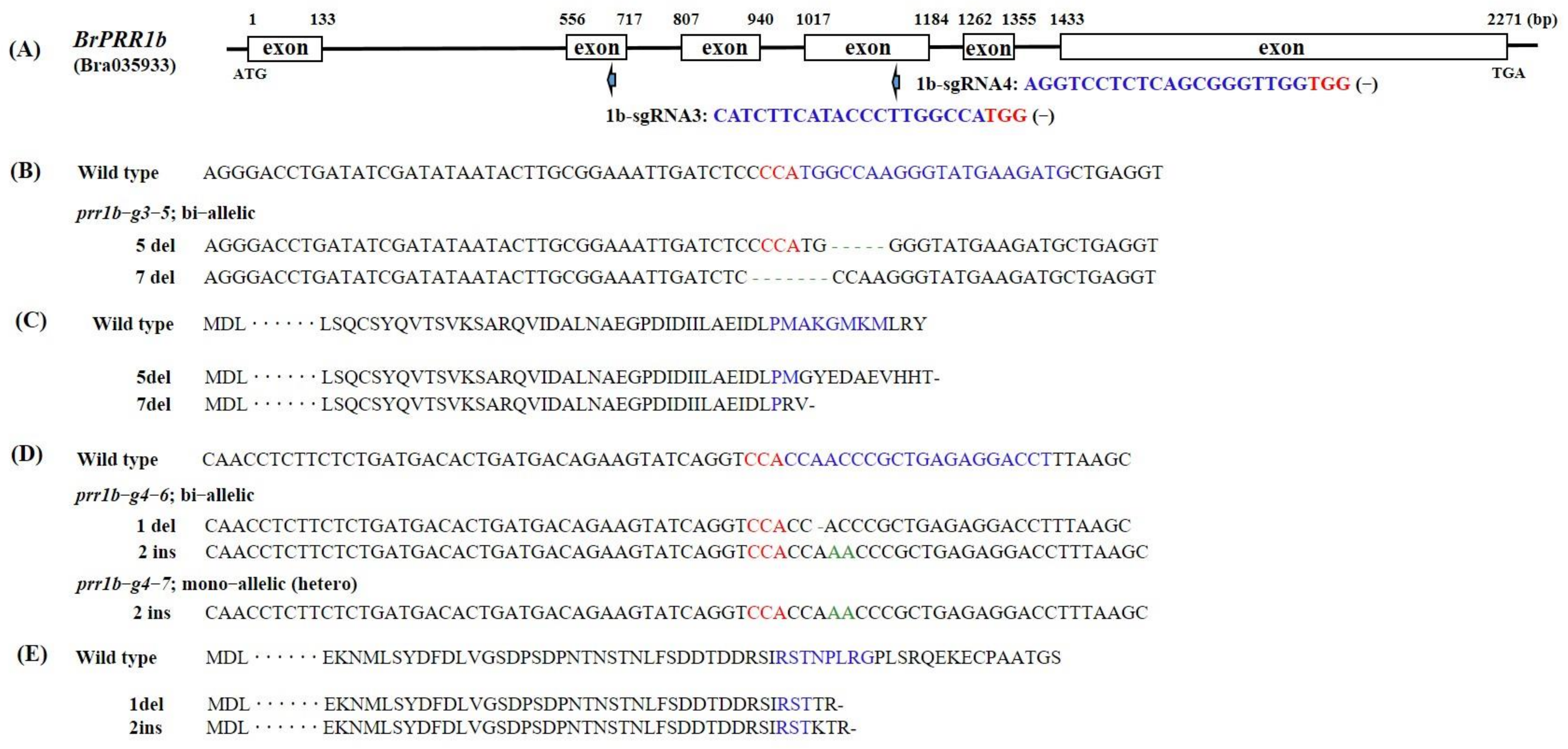
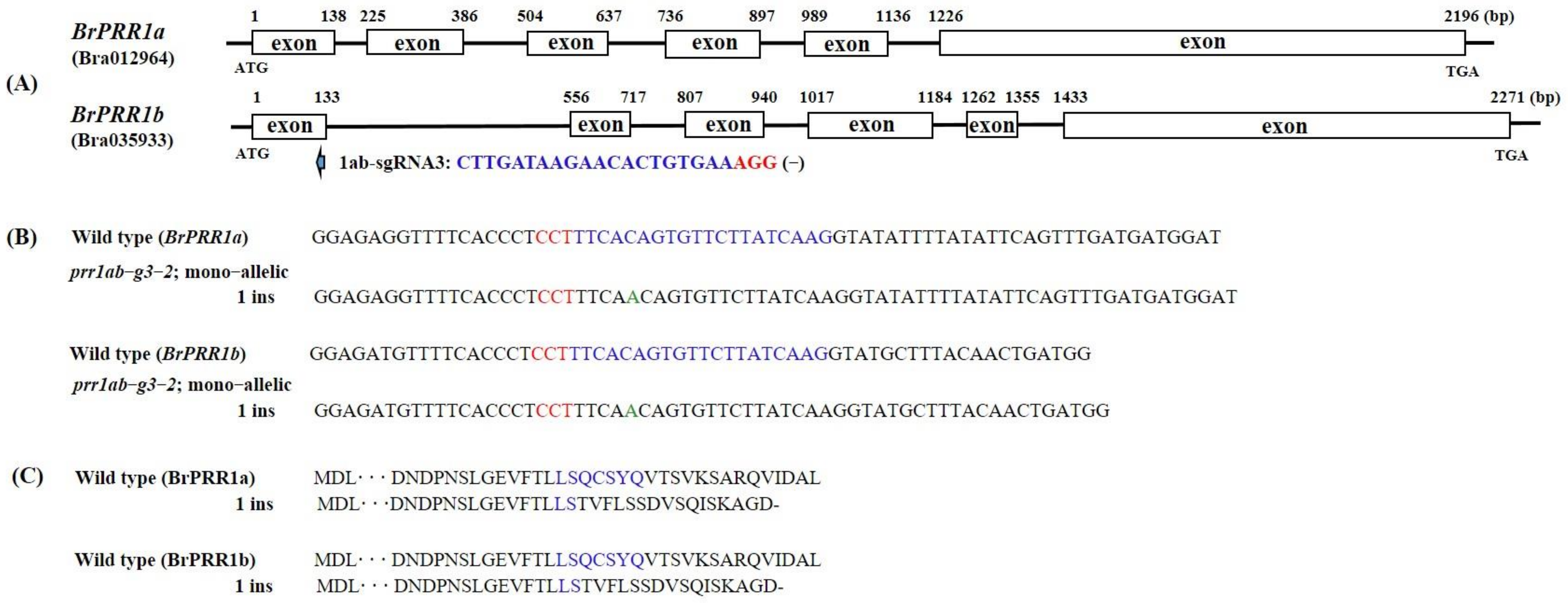
2.1. Selection of Target Genes and Generation of BrPRR1a, 1b, and 1ab Double-Knockout Mutants Using CRISPR/Cas9
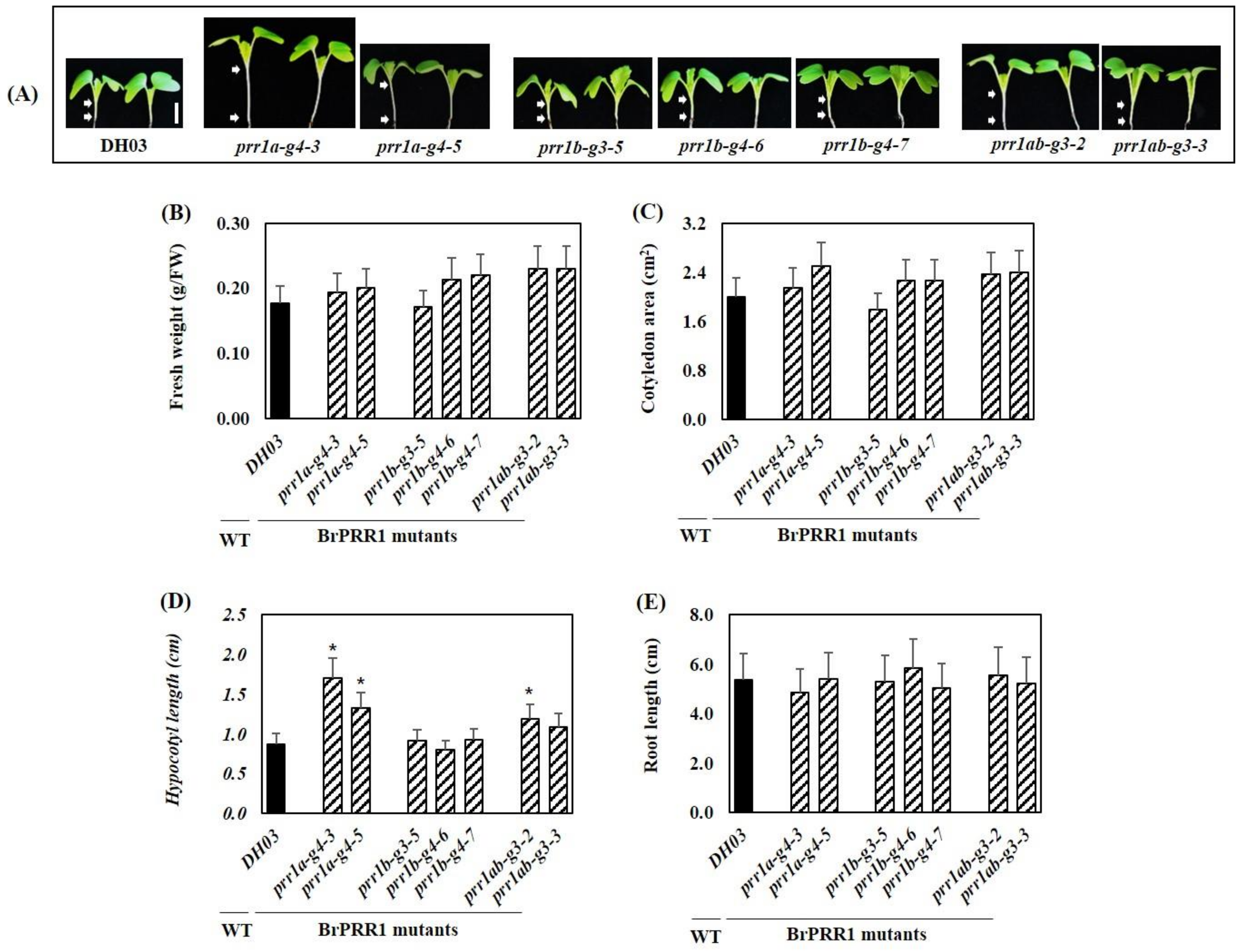
2.2. Morphological Effects of BrPRR1a, 1b, and 1ab Mutants in Chinese Cabbage Seedlings
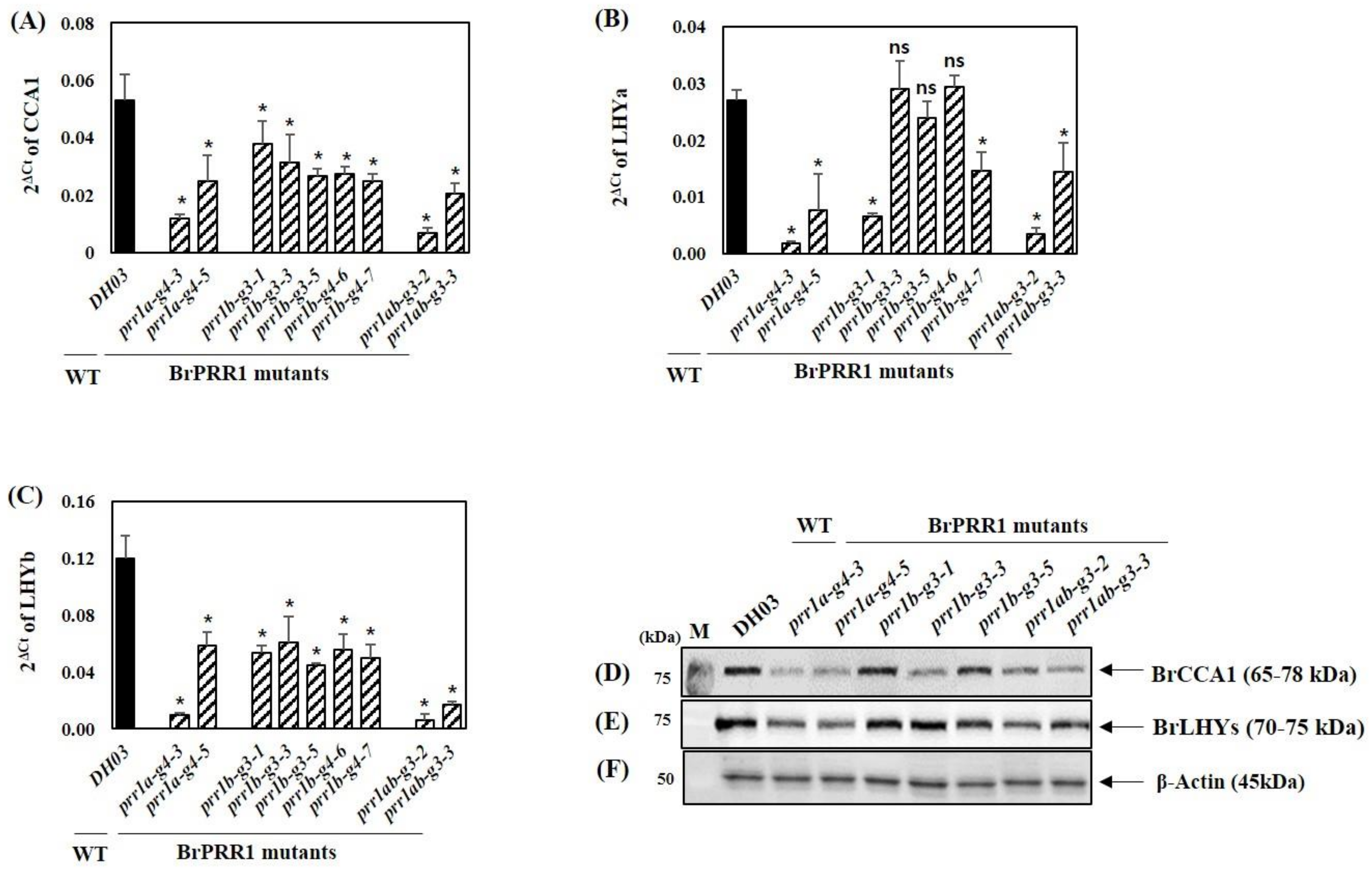
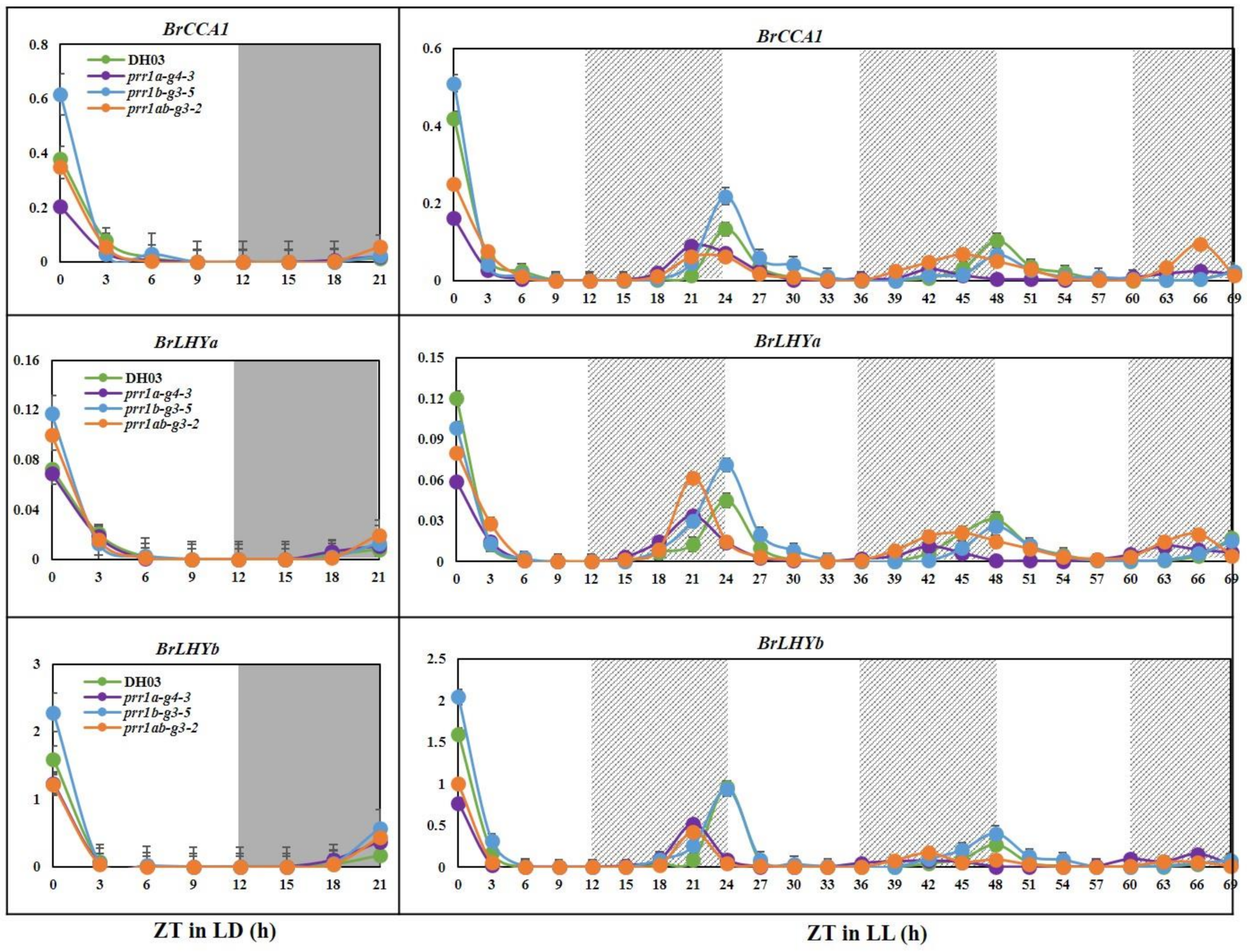
2.3. Expression of Circadian Clock Genes in BrPRR1a, 1b, and 1ab Mutants in Chinese Cabbage Seedlings
2.4. BrPRR1 Mutant Changed the Regulation of BrCCA1 and BrLHYa and b Expression in Chinese Cabbage
3. Discussion
3.1. Conservation of Genes through Evolution and Obtaining New Functions
3.2. PRR Genes in Plants
3.3. Possibility of Using PRR Genes in Crop Breeding
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. CRISPR/Cas9 Target Site Selection and Vector Construction
4.3. Chinese Cabbage Transformation and Acclimation to the Greenhouse
4.4. Targeted Deep Sequencing and Mutation Analysis
4.5. Quantitative Real-Time PCR Expression Analysis in B. rapa
4.6. Western Blotting Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Imaizumi, T. Arabidopsis circadian clock and photoperiodism: Time to think about location. Curr. Opin. Plant Biol. 2010, 13, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panter, P.E.; Muranaka, T.; Cuitun-Coronado, D.; Graham, C.A.; Yochikawa, A.; Kudoh, H.; Dodd, A.N. Circadian regulation of the plant transcriptome under natural conditions. Front Genet. 2019, 10, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locke, J.C.; Kozma-Bognar, I.; Gould, P.D.; Feher, B.; Kevei, E.; Nagy, F.; Turner, M.S.; Hall, A.; Millar, A.J. Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol. Syst. Biol. 2006, 2, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunlap, J.C. Genetic and molecular analysis of circadian rhythms. Annu. Rev. Genet. 1996, 30, 579–601. [Google Scholar] [CrossRef]
- Gendron, J.M.; Pruneda-Paz, J.L.; Doherty, C.J.; Gross, A.M.; Kang, S.E.; Kay, S.A. Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. USA 2012, 109, 3167–3172. [Google Scholar] [CrossRef] [Green Version]
- Pokhilko, A.; Fernandez, A.P.; Edwards, K.D.; Southern, M.M.; Halliday, K.J.; Illar, A.J. The clock gene circuit in Arabidopsis includes a repressilator with additional feedback loops. Mol. Syst. Biol. 2012, 8, 574. [Google Scholar] [CrossRef]
- Millar, A.J.; Carre, I.A.; Strayer, C.A.; Chua, N.H.; Kay, S.A. Circadian clock mutants in Arabidopsis identified by luciferase imaging. Science 1995, 267, 1161–1163. [Google Scholar] [CrossRef]
- Matsushika, A.; Makino, S.; Kojima, M.; Mizuno, T. Circadian waves of expression of the APRR1/TOC1 family of pseudo-response regulators in Arabidopsis thaliana: Insight into the plant circadian clock. Plant Cell Physiol. 2000, 41, 1002–1012. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Fujiwara, S.; Somers, D.E. PRR5 regulates phosphorylation, nuclear import and subnuclear localization of TOC1 in the Arabidopsis circadian clock. EMBO J. 2010, 29, 1903–1915. [Google Scholar] [CrossRef] [Green Version]
- Para, A.; Farre, E.M.; Imaizumi, T.; Pruneda-Paz, J.L.; Harmon, F.G.; Kay, S.A. PRR3 is a vascular regulator of TOC1 stability in the Arabidopsis circadian clock. Plant Cell 2007, 19, 3462–3473. [Google Scholar]
- Mas, P.; Kim, W.Y.; Somers, D.E.; Kay, S.A. Targeted degradation of TOC1 by ZTL modulates circadian function in Arabidopsis thaliana. Nature 2003, 426, 567–570. [Google Scholar] [CrossRef]
- Perales, M.; Mas, P. A functional link between rhythmic changes in chromatin structure and the Arabidopsis biological clock. Plant Cell 2007, 19, 2111–2123. [Google Scholar] [CrossRef] [Green Version]
- Alabadi, D.; Oyama, T.; Yanovsky, M.J.; Harmon, F.G.; Mas, P.; Kay, S.A. Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 2001, 293, 880–883. [Google Scholar]
- Legnaioli, T.; Mas, J. TOC1 functions as a molecular switch connecting the circadian clock with plant response to drought. EMBO J. 2009, 28, 3745–3757. [Google Scholar] [CrossRef] [Green Version]
- Strayer, C.; Oyama, T.; Schultz, T.F.; Raman, R.; Somers, D.E.; Mas, P.; Panda, S.; Kreps, J.A.; Kay, S.A. Cloning of the Arabidopsis clock gene TOC1, and autoregulatory response regulator homolog. Science 2000, 289, 768–771. [Google Scholar]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Kiu, J.; Xi, J.J.; Qiu, J.L.; et al. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686–688. [Google Scholar]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 2019, 70, 667–697. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar]
- Ma, X.; Zhang, Q.; Zhu, Q.; Liu, W.; Chen, Y.; Qiu, R.; Wang, B.; Yang, Z.; Li, H.; Lin, Y.; et al. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicot plants. Mol. Plant 2015, 8, 1274–1278. [Google Scholar]
- Yang, H.; Wu, J.J.; Tang, T.; Liu, K.D.; Dai, C. CRISPR/Cas9-mediated genome editing efficiently creates specific mutations at multiple loci using one sgRNA in Brassica napus. Sci. Rep. 2017, 7, 7489. [Google Scholar] [CrossRef] [Green Version]
- Braatz, J.; Harloff, H.J.; Mascher, M.; Stein, N.; Immelbach, A.; Jung, C. CRISPR-Cas9 targeted mutagenesis leads to simultaneous modification of different homoeologous gene copies in polyploidy oilseed rape (Brassica napus). Plant Physiol. 2017, 174, 935–942. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.F.; Liu, D.; Wu, D.; Fang, Y.; Wu, J.; Wang, Y. CRISPR/Cas9-mediated multiplex genome editing of the BnWRKY11 and BnWRKY70 genes in Brassica napus L. Int. J. Mol. Sci. 2018, 19, 2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.; Cai, S.; Yang, Y.; Amoo, O.; Fan, C.; Zhou, Y. CRISPR/Cas9-mediated genome editing reveals differences in the contribution of INDEHISCENT homologues to pod shatter resistance in Brassica napus L. Theor. Appl. Genet. 2019, 132, 2111–2123. [Google Scholar] [CrossRef]
- Chang, T.; Guan, M.; Zhou, B.; Peng, Z.; Xing, M.; Wang, X.; Guan, C. Progress of CRISPR/Cas9 technology in breeding of Brassica napus. Oil Crop Sci. 2021, 6, 53–57. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Makhmoudova, A.; Nitschke, F.; Tetlow, J.J.; Emes, M.J. CRISPR-Cas9-mediated editing of starch branching enzymes results in altered starch structure in Brassica napus. Plant Physiol. 2022, 188, 1866–1886. [Google Scholar] [CrossRef]
- Ma, C.; Zhu, C.; Zheng, M.; Liu, M.; Zhang, D.; Liu, B.; Li, Q.; Si, J.; Ren, X.; Song, H.Y. CRISPR/Cas9-mediated multiple gene editing in Brassica oleracea var. capitate using the endogenous tRNA-processing system. Hortic. Res. 2019, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neequaye, M.; Stavnstrup, S.; Harwood, W.; Lawrenson, T.; Hundleby, P.; Irwin, J.; Troncoso-Rey, P.; Saha, S.; Traka, M.H.; Mithen, R.; et al. CRISPR-Cas9-mediated gene editing of MYB28 genes impair glucoraphanin accumulation of Brassica oleracea in the field. CRISPR J. 2021, 4, 416–426. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, J.S.; Hong, J.K.; Lee, Y.H.; Choi, B.S.; Seol, Y.J.; Jeon, C.H. Comparative mapping, genomic structure, and expression analysis of eight pseudo-response regulator genes in Brassica rapa. Mol. Genet. Genom. 2012, 278, 373–388. [Google Scholar] [CrossRef]
- Kim, J.A.; Kim, J.S.; Hong, J.K.; Lee, Y.H.; Lee, S.I.; Jeong, M.J. Development of a marker system to discern the flowering type in Brassica rapa crops. J. Plant Biotechnol. 2017, 44, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.Y.; Oh, E.; Wang, T.; Wang, Z.Y. TOC1–PIF4 interaction mediates the circadian gating of thermoresponsive growth in Arabidopsis. Nat. Commun. 2016, 7, 13692. [Google Scholar] [CrossRef]
- Kim, J.A.; Jung, H.E.; Hong, J.K.; Hermand, V.; McClung, R.; Lee, Y.H.; Kim, J.Y.; Lee, S.I.; Jeong, M.J.; Kim, J.S.; et al. Reduction of GIGANTEA expression in transgenic Brassica rapa enhances salt tolerance. Plant Cell Rep. 2016, 35, 1943–1954. [Google Scholar] [CrossRef]
- Mao, Y.; Zhang, Z.; Feng, Z.; Wei, P.; Zhang, H.; Botella, J.R.; Zhu, J.K. Development of germ-line-specific CRISPR-Cas9 systems to improve the production of heritable gene modifications in Arabidopsis. Plant Biotech. J. 2016, 14, 519–532. [Google Scholar] [CrossRef] [Green Version]
- Lawrenson, T.; Shorinola, O.; Stacey, N.; Li, C.; Østergaard, L.; Patron, N.; Uauy, C.; Harwood, W. Induction of targeted, heritable mutations in barley and Brassica oleracea using RNA-guided Cas9 nuclease. Genome Biol. 2015, 16, 258. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Shim, D.H.; Kumari, S.; Jung, H.E.; Jung, K.H.; Jeong, H.S.; Kim, W.Y.; Lee, S.I.; Jeong, M.J. Transcriptome analysis of diurnal gene expression in Chinese cabbage. Genes 2019, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Moore, R.C.; Purugganan, M.D. The evolutionary dynamics of plant duplicate genes. Curr. Opin. Plant Biol. 2005, 8, 122–128. [Google Scholar] [CrossRef]
- Mena, M.; Ambrose, B.A.; Meeley, R.B.; Briggs, S.P.; Yanofsky, M.F.; Schmidt, R.J. Diversification of C-function activity in maize flower development. Science 1996, 274, 1537–1540. [Google Scholar] [CrossRef]
- Kroymann, J.; Donnerhacke, S.; Schnabelrauch, D.; Mitchell-Olds, T. Evolutionary dynamics of an Arabidopsis insect resistance quantitative trait locus. Proc. Natl. Acad. Sci. USA 2003, 100, 14587–14592. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, T.; Yamashino, T. Comparative transcriptome of diurnally oscillating genes and hormone-responsive genes in Arabidopsis thaliana: Insight into circadian clock-controlled daily responses to common ambient stresses in plants. Plant Cell Physiol. 2008, 49, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Lou, P.; Wu, J.; Cheng, F.; Cressman, L.G.; Wang, X.; McClung, C.R. Preferential retention of circadian clock genes during diploidization following whole genome triplication in Brassica rapa. Plant Cell 2012, 24, 2415–2426. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.A.; Yang, T.J.; Kim, J.S.; Park, J.Y.; Kwon, S.J.; Lim, M.H.; Jin, M.; Lee, S.C.; Lee, S.I.; Choi, B.S.; et al. Isolation of circadian-associated genes in Brassica rapa by comparative genomics with Arabidopsis thaliana. Mol. Cells 2007, 23, 145–153. [Google Scholar]
- Harmer, S.L.; Hogenesch, J.B.; Straume, M.; Chang, H.S.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.A.; Kay, S.A. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Salathia, N.; Hall, A.; Kevei, E.; Toth, R.; Nagy, F.; Hibberd, J.M.; Millar, A.J.; Webb, A.A.R. Plant circadian clocks increase photosynthesis, growth, survival, and competitive advantage. Science 2005, 309, 630–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bieniawska, Z.; Espinoza, C.; Schlereth, A.; Sulpice, R.; Hincha, D.K.; Hannah, M.A. Disruption of the Arabidopsis circadian clock is responsible for extensive variation in the cold-responsive transcriptome. Plant Physiol. 2008, 147, 263–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamichi, N.; Kusano, M.; Fukushima, A.; Kita, M.; Ito, S.; Yamashino, T.; Saito, K.; Sakakibara, H.; Mizuno, T. Transcript profiling of an Arabidopsis PSEUDO RESPONSE REGULATOR arrhythmic triple mutant reveals a role for the circadian clock in cold stress response. Plant Cell Physiol. 2009, 50, 447–462. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.Y.; Ali, Z.; Park, H.J.; Park, S.J.; Cha, J.Y.; Perez-Hormaeche, J.; Quintero, F.J.; Shin, G.; Kim, M.R.; Qiang, Z.; et al. Release of SOS2 kinase from sequestration with GIGANTEA determines salt tolerance in Arabidopsis. Nat. Commun. 2013, 4, 1352. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Xie, Q.; Anderson, R.G.; Ng, G.; Seitz, N.C.; Peterson, T.; McClung, C.R.; McDowell, J.M.; Kong, D.; Kwak, J.M.; et al. Crosstalk between the circadian clock and innate immunity in Arabidopsis. PLoS Pathog. 2013, 9, e1003370. [Google Scholar] [CrossRef] [Green Version]
- Baek, D.; Kim, W.Y.; Cha, J.Y.; Park, H.J.; Shin, G.; Park, J.; Lim, C.J.; Chun, H.J.; Li, N.; Kim, D.H.; et al. The GIGANTEA-ENHANCED EM LEVEL complex enhances drought tolerance via regulation of abscisic acid synthesis. Plant Physiol. 2020, 184, 443–458. [Google Scholar] [CrossRef]
- Marcolino-Gomes, J.; Rodrigues, F.A.; Fuganti-Pagliarini, R.; Bendix, C.; Nakayama, T.J.; Celaya, B.; Molinari, H.B.C.; Neves de Oliveira, M.C.; Harmon, F.G.; Nepomuceno, A. Diurnal oscillations of soybean circadian clock and drought responsive genes. PLoS ONE 2014, 9, e86402. [Google Scholar]
- Izawa, T.; Mihara, M.; Suzuki, Y.; Gupta, M.; Itoh, H.; Nagano, A.J.; Motoyama, R.; Sawada, Y.; Yano, M.; Hirai, M.Y.; et al. Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell 2011, 23, 1741–1755. [Google Scholar] [CrossRef] [Green Version]
- Brandoli, C.; Petri, C.; Egea-Cortines, M.; Weiss, J. The clock gene Gigantea 1 from Petunia hybrida coordinates vegetative growth and inflorescence architecture. Sci. Rep. 2020, 10, 275. [Google Scholar] [CrossRef]
- Ke, Q.; Kim, H.S.; Wang, Z.; Ji, C.Y.; Jeong, J.C.; Lee, H.S.; Choi, Y.I.; Xu, B.; Deng, X.; Yun, D.J.; et al. Down-regulation of GIGANTEA-like genes increases plant growth and salt stress tolerance in poplar. Plant Biotechnol. J. 2017, 15, 331–343. [Google Scholar] [CrossRef]
- Cervela-Cardona, L.; Yoshida, T.; Zhang, Y.; Okada, M.; Fernie, A.; Mas, P. Circadian control of metabolism by the clock component TOC1. Front. Plant Sci. 2021, 12, 683516. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.T.; Ryu, J.; Choi, M.K.; Kweon, J.; Kang, B.C.; Ahn, H.M.; Bae, S.; Kim, J.; Kim, J.S.; et al. A simple, flexible and high-throughput cloning system for plant genome editing via CRISPR-Cas system. J. Integr. Plant Biol. 2016, 58, 705–712. [Google Scholar] [CrossRef]
- Park, J.; Lim, K.; Kim, J.S.; Bae, S. Cas-analyzer: An online tool for assessing genome editing results using NGS data. Bioinformatics 2017, 33, 286–288. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | sgRNA | sgRNA Target | Direction | GC Contents (%) | Out-of Frame Score | Mismatches | ||
|---|---|---|---|---|---|---|---|---|
| 0 bp | 1 bp | 2 bp | ||||||
| BrPRR1a | 1a-sgRNA1 | TGATCCCAATAGCTTGGGAGAGG | + | 50 | 70.2 | 1 | 0 | 0 |
| 1a-sgRNA2 | AAAACCTCTCCCAAGCTATTGGG | − | 40 | 73.4 | 1 | 0 | 0 | |
| 1a-sgRNA3 | GCCTGACATCGATATAATACTGG | + | 40 | 56.4 | 1 | 0 | 0 | |
| 1a-sgRNA4 | GAGGCAAGACGAGGTCCCTGTGG | + | 65 | 52.3 | 1 | 0 | 0 | |
| 1a-sgRNA5 | TCAAGCACTTTACCACCACAGGG | + | 45 | 60.1 | 1 | 0 | 0 | |
| BrPRR1b | 1b-sgRNA1 | GGAGGGTGAAAACATCTCCCAGG | − | 55 | 56.5 | 1 | 0 | 0 |
| 1b-sgRNA2 | TGCGGAAATTGATCTCCCCATGG | + | 50 | 72.1 | 1 | 0 | 0 | |
| 1b-sgRNA3 | CATCTTCATACCCTTGGCCATGG | − | 50 | 57.4 | 1 | 0 | 0 | |
| 1b-sgRNA4 | AGGTCCTCTCAGCGGGTTGGTGG | − | 65 | 80.8 | 1 | 0 | 0 | |
| 1b-sgRNA5 | CGGCTTAAAGGTCCTCTCAGCGG | − | 55 | 72.6 | 1 | 0 | 0 | |
| BrPRR1a and BrPRR1b (1ab) | 1ab-sgRNA1 | TTATTGACAGAAGCAGAGTTAGG | + | 35 | 60.5 | 2 | 0 | 0 |
| 1ab-sgRNA2 | ATAAGAACACTGTGAAAGGAGGG | − | 35 | 85.4 | 2 | 0 | 0 | |
| 1ab-sgRNA3 | CTTGATAAGAACACTGTGAAAGG | − | 35 | 78.3 | 2 | 0 | 0 | |
| 1ab-sgRNA4 | TCACACGTGACAAAGATCTTCGG | + | 40 | 61.7 | 2 | 0 | 0 | |
| 1ab-sgRNA5 | CCAACGAGCTTCTCAACTTGTGG | + | 50 | 76.5 | 2 | 0 | 0 | |
| Gene | sgRNA | Target Region | Sample | Total | Insertion | Deletion | Mutated Count | Mutated Ratio |
|---|---|---|---|---|---|---|---|---|
| BrPRR1a | 1a-sgRNA1 | TGATCCCAATAGCTTGGGAGAGG | prr1a-g1 | 30,969 | 1505 | 397 | 1902 | 6.14% |
| 1a-sgRNA2 | AAAACCTCTCCCAAGCTATTGGG | prr1a-g2 | 30,205 | 7706 | 421 | 8127 | 26.91% | |
| 1a-sgRNA3 | GCCTGACATCGATATAATACTGG | prr1a-g3 | 8436 | 0 | 8428 | 8428 | 99.91% | |
| 1a-sgRNA4 | GAGGCAAGACGAGGTCCCTGTGG | prr1a-g4 | 11,652 | 0 | 11,652 | 11,652 | 100.00% | |
| 1a-sgRNA5 | TCAAGCACTTTACCACCACAGGG | prr1a-g5 | 37,980 | 463 | 537 | 1000 | 2.63% | |
| BrPRR1b | 1b-sgRNA1 | GGAGGGTGAAAACATCTCCCAGG | prr1b-g1 | 25,547 | 80 | 46 | 126 | 0.49% |
| 1b-sgRNA2 | TGCGGAAATTGATCTCCCCATGG | prr1b-g2 | 41,371 | 2424 | 1315 | 3739 | 9.04% | |
| 1b-sgRNA3 | CATCTTCATACCCTTGGCCATGG | prr1b-g3 | 9903 | 3 | 9898 | 9901 | 99.98% | |
| 1b-sgRNA4 | AGGTCCTCTCAGCGGGTTGGTGG | prr1b-g4 | 6633 | 9 | 3432 | 3441 | 51.88% | |
| 1b-sgRNA5 | CGGCTTAAAGGTCCTCTCAGCGG | prr1b-g5 | 41,905 | 431 | 231 | 662 | 1.58% | |
| BrPRR1ab | BrPRR1a region | |||||||
| 1ab-sgRNA1 | TTATTGACAGAAGCAGAGTTAGG | prr1ab-g1 | 12,299 | 2978 | 210 | 3188 | 25.92% | |
| 1ab-sgRNA2 | ATAAGAACACTGTGAAAGGAGGG | prr1ab-g2 | 16,666 | 7618 | 8945 | 16,563 | 99.38% | |
| 1ab-sgRNA3 | CTTGATAAGAACACTGTGAAAGG | prr1ab-g3 | 16,353 | 3858 | 3480 | 7338 | 44.87% | |
| 1ab-sgRNA4 | TCACACGTGACAAAGATCTTCGG | prr1ab-g4 | 35,791 | 0 | 0 | 0 | 0.00% | |
| 1ab-sgRNA5 | CCAACGAGCTTCTCAACTTGTGG | prr1ab-g5 | 25,968 | 40 | 75 | 115 | 0.44% | |
| BrPRR1b region | ||||||||
| 1ab-gRNA1 | TTATTGACAGAAGCAGAGTTAGG | prr1ab-g1 | 13,192 | 3299 | 9887 | 13,186 | 99.95% | |
| 1ab-sgRNA2 | ATAAGAACACTGTGAAAGGAGGG | prr1ab-g2 | 18,464 | 8607 | 9818 | 18,425 | 99.79% | |
| 1ab-sgRNA3 | CTTGATAAGAACACTGTGAAAGG | prr1ab-g3 | 15,966 | 8329 | 6763 | 15,092 | 94.53% | |
| 1ab-sgRNA4 | TCACACGTGACAAAGATCTTCGG | prr1ab-g4 | 37,652 | 0 | 3 | 3 | 0.01% | |
| 1ab-sgRNA5 | CCAACGAGCTTCTCAACTTGTGG | prr1ab-g5 | 31,533 | 69 | 136 | 205 | 0.65% |
| Sample | No. of Genome Mutated Plants | Indel Frequency (%) | Genotypes (No.) | ||||
|---|---|---|---|---|---|---|---|
| Homo Allelic | Hetero Allelic | Bi Allelic | Multiple Allelic | Mosaic Allelic | |||
| prr1a-g3 | 13 | 99.8–100 | 1 | 12 | |||
| prr1a-g4 | 16 | 99.9–100 | 14 | 2 | |||
| prr1b-g3 | 11 | 98.3–100 | 2 | 5 | 2 | 2 | |
| prr1b-g4 | 13 | 44.9–100 | 2 | 8 | 1 | 2 | |
| prr1ab-g2 | 9 | 51–99.9 | 1 | 2 | 5 | 1 | |
| prr1ab-g3 | 13 | 39–99.9 | 5 | 2 | 6 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.-S.; Yu, J.; Bae, S.; Kim, H.S.; Park, S.; Lee, K.; Lee, S.I.; Kim, J.A. Identification and Characterization of PSEUDO-RESPONSE REGULATOR (PRR) 1a and 1b Genes by CRISPR/Cas9-Targeted Mutagenesis in Chinese Cabbage (Brassica rapa L.). Int. J. Mol. Sci. 2022, 23, 6963. https://doi.org/10.3390/ijms23136963
Kim N-S, Yu J, Bae S, Kim HS, Park S, Lee K, Lee SI, Kim JA. Identification and Characterization of PSEUDO-RESPONSE REGULATOR (PRR) 1a and 1b Genes by CRISPR/Cas9-Targeted Mutagenesis in Chinese Cabbage (Brassica rapa L.). International Journal of Molecular Sciences. 2022; 23(13):6963. https://doi.org/10.3390/ijms23136963
Chicago/Turabian StyleKim, Nan-Sun, Jihyeon Yu, Sangsu Bae, Hyang Suk Kim, Soyoung Park, Kijong Lee, Soo In Lee, and Jin A. Kim. 2022. "Identification and Characterization of PSEUDO-RESPONSE REGULATOR (PRR) 1a and 1b Genes by CRISPR/Cas9-Targeted Mutagenesis in Chinese Cabbage (Brassica rapa L.)" International Journal of Molecular Sciences 23, no. 13: 6963. https://doi.org/10.3390/ijms23136963