
Neurogenesis as a Tool for Spinal Cord Injury


1
Institute of Experimental Medicine, Czech Academy of Sciences, Vídeňská 1083, 14220 Prague, Czech Republic
2
Department of Neuroscience, Second Faculty of Medicine, Charles University, 15006 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(7), 3728; https://doi.org/10.3390/ijms23073728
Submission received: 17 February 2022
/
Revised: 24 March 2022
/
Accepted: 25 March 2022
/
Published: 28 March 2022
(This article belongs to the Special Issue Neurogenesis and Neural Plasticity 2.0)
{kind=link}
{kind=link}
Abstract
:Spinal cord injury is a devastating medical condition with no effective treatment. One approach to SCI treatment may be provided by stem cells (SCs). Studies have mainly focused on the transplantation of exogenous SCs, but the induction of endogenous SCs has also been considered as an alternative. While the differentiation potential of neural stem cells in the brain neurogenic regions has been known for decades, there are ongoing debates regarding the multipotent differentiation potential of the ependymal cells of the central canal in the spinal cord (SCECs). Following spinal cord insult, SCECs start to proliferate and differentiate mostly into astrocytes and partly into oligodendrocytes, but not into neurons. However, there are several approaches concerning how to increase neurogenesis in the injured spinal cord, which are discussed in this review. The potential treatment approaches include drug administration, the reduction of neuroinflammation, neuromodulation with physical factors and in vivo reprogramming.
1. Introduction
A spinal cord injury (SCI) is an insult to the spinal cord resulting in a change, either temporary or permanent, in the cord’s normal motor, sensory or autonomic function. Patients with SCI usually have permanent and often devastating neurologic deficits and disability. The major consequences are tissue damage, the death of neurons and disruption of neuronal connections; all lead to the loss of mobility, sensation or autonomic function. The pathophysiology of traumatic SCI has two phases: primary and secondary injuries. Primary injury is the result of trauma in which spinal cord tissue is violently damaged. In this phase of injury, within a few minutes, cell death and damage to the vasculature and blood–spine barrier occurs [1]. Simultaneously, a cascade of events is initiated that leads to extensive secondary damage [2]. First, inflammation and hemorrhage develop in the tissue, leading to necrosis and ischemia [3]. Massive collections of inflammatory cells appear at the injury site within 12–24 h, where the first inflammatory cells are neutrophils, followed by lymphocytes and then macrophages [2]. Damage in the tissue continues 2–4 days after injury with the disruption of ion homeostasis [4], glutamate excitotoxicity [5], production of reactive oxygen species [6], lipid peroxidation [7], impaired autophagy [8], accumulation of nitric oxide flux [9], glial scar formation [10] and energy failure [11]. In the months and years after injury, the subacute phase transitions to a chronic phase in which central cavitation occurs, glial scar formation continues and changes in ion channels and receptors occur. Oligodendrocyte apoptosis leads to demyelination and damage to the surviving axons [12]. Regeneration in the form of sprouting axons [13] is prevented by the non-permissive environment formed by extracellular matrix molecules during the subacute and chronic phases of injury. The glial scar formed by reactive astrocytes around cystic cavities prevents damage to the adjacent tissue but, at the same time, together with extracellular matrix proteins, such as chondroitin sulfate proteoglycans, tenascin and NG2 proteoglycan, limits axon regeneration and plasticity [14].
To date, there is no effective treatment for SCI. One possible approach to SCI treatment could be provided by stem cells (SCs). Studies have mainly focused on the transplantation of exogenous SCs, but the induction of endogenous SCs has also been considered as an alternative. This approach would avoid the risks accompanying exogenous SC transplantations, such as immunoreactivity or the formation of tumors.
2. Endogenous Neural Stem Cells
Currently, there are two well-described main regions in the mammalian brain that contain neural stem cells (NSCs): the subgranular zone in the dentate gyrus and the subventricular zone of the lateral ventricles. Cells from the subventricular zone generate doublecortin positive neuroblasts, which migrate to the olfactory bulb and differentiate into olfactory neurons, tuning the fine plasticity of the olfactory system. The subgranular zone of the gyrus dentate is involved in learning and memory (reviewed in [15]). Thereafter, the striatum, along with other areas, were reported as an additional neurogenic niche in humans [16,17]. While the differentiation potential of neural stem cells in the brain neurogenic regions has been known for decades, there are ongoing debates regarding the multipotent differentiation potential of the ependymal cells of the central canal in the spinal cord (SCECs). SCECs form a continuous epithelial sheet lining the ventricles and the central canal of the spinal cord. These cells are of glial lineage but have many epithelial characteristics, including a basement membrane, cell–cell junctions and motile cilia [18].
The first reports regarding the renewal of ependymal cells in a mouse spinal cord appeared in 1962 using radioactivity experiments [19]. The authors reported newly dividing cells in SCI, which were identified as astrocytes and oligodendrocytes but not neurons. These findings were later confirmed by several authors supporting the statement that neurogenesis is not present in the rodent spinal cord [18,20,21]. New isolation, expansion and culturing of cells in the form of neurospheres brought new evidence that cells from the spinal cord can not only self-renew but can form neurospheres and generate in vitro neuronal cells [22]. Subsequently, it was necessary to identify these multipotent stem-cell-like cells. There were several possible candidates: astrocyte precursor cells, oligodendrocytes precursor cells and ependymal cells. Based on genetic fate mapping, it was shown that while oligodendrocyte progenitors can self-renew and give rise to new mature oligodendrocytes, only ependymal cells are multipotent and neural stem cell activity in the intact and injured adult mouse spinal cord is restricted to this cell population [18,21]. In contrast, oligodendrocyte progenitor cells expressing markers such as nerve/glial antigen-2 (NG2) and/or platelet-derived growth factor receptor alpha (PDGFRα), also known as NG2-glia, NG2-cells or polydendrocytes, are scattered in the white matter and gray matter throughout the central nervous system and represent the main proliferating cell population in the intact spinal cord; however, they do not display in vitro neural stem cell properties [18].
The discovery of NSCs in the spinal cord was prolonged due to their lack of activity under physiological conditions. SCECs can be activated during pathological conditions, such as SCI, inflammation or neurodegeneration. Spinal cord ependymal cells start to proliferate, migrate to the site of damage and differentiate. The phenotype of the differentiated cell depends on the surrounding environment. For example, in the model of multiple sclerosis as an inflammatory disease, SCECs differentiate into oligodendrocytes and possibly into neurons [23]. In a neurodegenerative disease, such as amyotrophic lateral sclerosis, SCECs mostly differentiate into astrocytes [24]. SCECs can also be activated by physical activity; proliferating nestin-positive cells were detected 4–7 days after running wheel training in rats [25].
SCECs in Spinal Cord Injury
Following spinal cord insult, SCECs start to proliferate and differentiate mostly into astrocytes and partly into oligodendrocytes, but not into neurons [18,21,25,26,27] (Figure 1). A glial scar has both beneficial and detrimental effects on recovery after spinal cord injury [28,29]. Therefore, the differentiation of SCECs to astrocytes plays a substantial role in SCI healing. It was shown that astrocytes originating from SCECs help to form a glial scar bordering the SCI [18]. Without these astrocytes, the damage would spread to the surrounding tissue and result in an enlarged lesion volume, neuronal degeneration and a worse functional outcome [30].
A glial scar is, however, not only formed from the astrocytes derived from SCECs but also from astrocytes derived from astrocyte progenitors. These two populations contain different properties. A few days following an SCI, the progenitor cells from the central canal region migrate in the direction of the injury center and downregulate ependymal cell markers, such as Sox2, Sox3 and FoxJ1. These cells further divide and differentiate into astrocytes, which form the core of a glial scar, whereas astrocytes from astrocyte progenitors migrate toward the periphery of the glial scar [18]. In addition, astrocytes from SCECs do not usually express GFAP, but they produce laminin, which helps axons to grow, whereas GFAP+ astrocytes from dividing astrocyte progenitors produce chondroitin sulfate proteoglycans (CSPGs) that inhibit the axon growth [21]. SCEC progenitors also exert a neurotrophic effect that is required for the survival of the surrounding neurons [30], as the progenitors express several growth factors and increase expression after differentiation when expanded in vitro [31].
Unfortunately, the beneficial effect of SCEC-differentiated astrocytes is not sufficient for spinal cord regeneration. For full recovery from an SCI, there is a need to replace destroyed neurons and also oligodendrocytes to myelinate them. Therefore, the activation of endogenous NSCs and their neuronal differentiation induction would be an interesting approach to generate the lost population of neurons. The first studies, which focused on overcoming the gliogenic environment in the spinal cord, were not successful. Spinal cord neural stem cells genetically modified to express neurogenin 2 (Ngn2) differentiated into neurons in vitro, but not after transplantation into an SCI [32]. In contrast, other studies have confirmed the ability of neural stem cells that expanded from the adult spinal cord to differentiate into neurons in the neurogenic niche in the dentate gyrus [33]. Therefore, different strategies to facilitate neuronal differentiation from SCEC have started to emerge (Figure 2).
3. Approaches to Promote Neurogenesis
3.1. The Differentiation of SCECs into Neurons
A small environmental change can induce NSCs to produce neurons instead of glial cells. The addition of valproic acid (VPA) to embryonic brain NSCs transplanted into the spinal cord led to the production of neurons instead of only astrocytes. These differentiated neurons were able to connect into the existing network and form synapses with endogenous neurons, which led to functional improvement in mice [34].
A similar effect was described for the endogenous SCECs. In vitro experiments clearly indicated that SCECs have an intrinsic capacity of producing neurons and their fate depends on the surrounding environment. SCECs cultured with VPA have increased neuronal induction and promote neuronal differentiation, while astrocytic differentiation is suppressed. Cell cycle regulator p21(Cip/WAF1) and proneural genes Ngn2 and NeuroD1 were increased in these two processes, respectively [35,36]. VPA has many pharmacological effects and is already used as a medicament to treat epilepsy and bipolar disorder. In rat SCI, delayed treatment with VPA led to in vivo increased neurogenesis; the evidence of which was based on the newborn neuron marker doublecortin and the mature neuron marker neuron-specific nuclear protein, which were enhanced in the epicenter of the SCI and neighboring tissue [36]. VPA-induced HDAC inhibition led to the activation of the Wnt/β-catenin pathway and, consequently, decreased GSK-3 activity. Reduced GSK-3 activity results in the upregulation of the substrate of GSK-3-cytoplasmic levels of the transcription factor β-catenin, that is, they were negatively regulated through phosphorylation-dependent degradation [37,38]. Moreover, neurogenesis induced using VPA can create a more protective environment in the tissue due to the expression of neurotrophic factor BDNF in newborn neurons. The upregulation of BDNF via BNDF-Trkb, leads to the subsequent activation of MAPK/Erk pathways [39,40]. The increase in Erk indirectly inhibits GSK-3; therefore, neuroprotection and neurogenesis are coupled in the VPA treatment (reviewed in [41]).
The inhibition of GSK-3 appears to be a potential target for increased neurogenesis, as shown in further studies. A recent study showed that the application of GSK-3 inhibitor Ro3303544 on SCECs isolated from mouse spinal cords and leads to the increased expression of early neuronal marker βIII-tubulin and late neuronal marker MAP2. Similarly, in mouse SCI, treatment with GSK-3 inhibitor Ro3303544 not only increased the survival of neurons but also more newborn neurons formed synapses close to the injury epicenter. This all led to improved motor recovery and decreased astrogliosis in the injury epicenter [42].
SCEC neuronal differentiation can be enhanced by substance P. This neuropeptide is involved in the synthesis of growth factors and cytokines and, therefore, in cell proliferation. The injection of substance P leads to functional improvement by activating SCECs. SCEC activation increases their proliferation and differentiation into neurons and decreases proliferation and differentiation into astrocytes in vivo. In vitro, it enhances neuronal differentiation by activating the Erk1/2 pathway [43]. These findings are consistent with previous studies with VPA and Ro3303544 since Erk1/2 inhibits GSK3 and vice versa [44,45,46].
VPA also synergizes with all-trans retinoic acid (RA), an important regulator during embryonic development, when it defines the anterior/posterior axis. RA increases the neurogenesis of SCECs in vitro and reduces their differentiation into astrocytes [35,47]. RA can be combined with the growth factors bFGF/EGF, which promote axon growth and improve the neuronal differentiation of the embryonic brain NSCs [48]. Neural differentiation induced by RA can be disrupted by knocking down the BAF45D protein [49]. This protein is present in the developing mouse cortex and the adult mouse hippocampus. Silencing its expression leads to the inhibited expression of Pax6, which is a neurogenic transcription factor contributing to neurogenesis. In the spinal cord, BAF45D is expressed in SCECs, neurons and oligodendrocytes, but not in astrocytes. After SCI, the expression of BAF45D in SCECs is decreased, and thus, proliferating SCECs mainly differentiate into astrocytes instead of neurons [50]. Therefore, the targeting of this peptide may be another potential way to affect neurogenesis after an SCI.
The important molecules that are also known from the developmental stage are connexins, which are proteins that form gap junctions. These molecules play a role not only in development but also in proliferation and differentiation [51,52]. The expression of Sox2, which is a neural progenitor marker participating in the conversion of endogenous glia into neurons [53], is regulated by connexin 50. Silencing connexin 50 resulted in the downregulation of connexin 50, while its overexpression led to more Sox2 cells in the SCEC population [52]. A subpopulation of SCECs lateral from the central canal is connected with gap junctions. This connection is downregulated at the end of development but upregulated again after an SCI. This increase in the gap junction coupling correlates with connexin 26 upregulation and leads to the recovery of SCEC proliferation. On the other hand, the blocking of this connexin decreases SCEC proliferation [51]. Therefore, connexins and gap junctions are important for SCEC proliferation and, thereby, a potential target.
Erythropoietin (Epo) is a hormone that is important for erythropoiesis. In spinal cord injury treatment, Epo signaling is involved in several neuroprotective processes, such as anti-apoptotic and anti-inflammatory functions and edema reduction [54,55]. Meanwhile, in the brain neurogenic regions, Epo application increases the number of newly generated neurons [56] in the healthy spinal cord and does not affect SCEC proliferation. However, after SCI, Epo treatment significantly promotes SCEC differentiation into neurons and oligodendrocytes [57].
Growth factors (GFs) are a family of proteins that are involved in the regulation of development and function, the survival of neurons, neurotransmitter release, recovery of synaptic function and axon regeneration [58]. However, various types of growth factors have different functions regarding repairing SCI [59]. Therefore, it is apparent that the use of growth factors is one of the most tested approaches to promote neurogenesis. The epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF, also known as FGF2) were shown to be able to jointly, but not alone, induce the proliferation of SCECs in vitro [22,60]. The same was even shown later in vivo [61,62], where the infusion of either EGF or bFGF alone into the spinal cord had no effect, but their combination led to the increased proliferation of SCECs. The same authors later reported that the infusion of EGF + bFGF into the injured spinal cord of rats led to the proliferation and migration of SCECs to the site of injury and functional improvement. However, the mechanism responsible for functional improvement remained unclear, as there were no new neurons or oligodendrocytes detected [63]. The effect of growth factors may remain behind the mechanisms for increased proliferation of SCECs after exercise, as physical activity increases the expression of GFs [25].
The effect of GFs can be facilitated by genetic manipulation. The combination of GF with transcription factor overexpression can support neurogenesis and/or oligodendrogliogenesis. The transcription factor Ngn2 induced neuronal differentiation in vitro but had almost no effect in vivo. However, in combination with the previously mentioned EGF and FGF2 (bFGF), the retroviral-induced overexpression of Ngn2 led to the production of new neurons in vivo [64]. The same viral vector was used to overexpress Mash1, which resulted in the production of new oligodendrocytes in the same experiment [64]. Due to the short half-life of GFs and the need for their sustained release, GFs are often delivered in combination with biomaterials. For example, a biodegradable chitosan scaffold was loaded with NT3 and inserted into a 5 mm gap in a completely transected spinal cord. Slowly released NT3 activated SCECs to migrate into the lesion area and differentiate into neurons, which formed a functional network that led to the functional recovery of rats with an SCI [65,66]. As this approach was successful it was also repeated in monkeys with a hemisection model of SCI, where it led to neuroregeneration, the growth of cortico-spinal tract (CST) axons through the lesion and functional recovery [67]. A sodium hyaluronate scaffold was combined with a ciliary neurotrophic factor [68]. In this study, a scaffold with neurotrophic factor was implanted into a 5 mm gap after the removal of a 5 mm T8 segment of a spinal cord in rats. The scaffold releasing ciliary neurotrophic factor (CNTF) led to the activation of endogenous SCECs, their migration to the injury site, differentiation into neurons and even the formation of functional synapses, followed by the improvement of motor and sensory functions [68]. Injectable hydrogels can fill the lesion cavities and, therefore, appropriately integrate into the tissue and modify the environment for better regeneration [69,70]. Hydrogels can support pro-regenerative macrophage polarization and angiogenesis, leading to axonal regeneration and neurogenesis. The injection of a functional ECM-resembling, self-assembling, peptide nanofiber hydrogel with CNTF, BDNF and NGF growth factors has led to the axonal regeneration, myelination, proliferation and neuronal differentiation of SCECs that connect to the CST, which resulted in the recovery of locomotion in rats with a SCI [70].
In addition to the direct administration of GFs, GF receptors are important targets. Myelin-associated inhibitors (MAIs) and chondroitin sulfate proteoglycans (CSPGs) are major components of the inhibitory microenvironment following an SCI. The activation of EGF receptor (EGFR) by MAIs suppresses the neuronal differentiation of NSCs [71,72]. Therefore, blocking this signaling pathway might lead to increased neuronal differentiation. This was demonstrated in studies using an EGFR antibody, such as the drug cetuximab, which is usually used for cancer treatment [73,74,75,76]. Cetuximab released from the collagen scaffold increased neuronal and decreased astrocyte differentiation in vitro [75]. Moreover, the transplantation of this scaffold into SCI induced neuronal differentiation, reduced astrogliosis and improved axonal regeneration in vivo [74,75,76]. Instead of the whole drug Cetuximab, only fragments of the antibody against EGFR, fused with a collagen-binding domain loaded on a collagen scaffold, can be engineered. When transplanted into an acute SCI, it can function in a similar way. The modified collagen scaffold facilitated the maturation of the newborn neurons, which differentiated from the endogenous neural stem cells. Synaptic connections detected in the lesion suggest the integration of newborn cells into the existing neuronal network [73].
The biggest limitation of the above section is the fact that the majority of experiments were performed in rodent cellular or animal models. More information on in vitro differentiation could be obtained from human iPS cells differentiated into ependymal cells to recapitulate some of the crucial experiments confirming neurogenesis described in Section 3. However, there are limited protocols describing the differentiation of iPS into ependymal cells [77]. Some studies even question the possibility of neurogenesis in the spinal cord by any means [78,79]. Ren et al. [78] performed large crush injuries across the spinal cord and found minimal SCEC migration with less than 2% contribution of SCECs to the total newly proliferated scar-forming astrocytes. Confirmation of neurogenesis in human spinal cord tissue requires a post mortem analysis of individuals with spinal cord injury compared to non-traumatic causes (controls). The study of Cawsey et al. [80] reported a significant increase in the percentage of SCECs that were nestin-positive (a marker of neural progenitor cell response) between the controls and trauma cases in human samples. Nestin-positive cells were seen in cervical, thoracic and lumbar levels of the spinal cord, suggesting that nestin reactivity is not just a localized reaction to injury. Further characterization of SCECs in the human spinal cord is, therefore, required to determine their role after injury and to confirm the character of neural progenitor cells.
3.2. The Reduction of Neuroinflammation
Primary injury in the spinal cord is followed by secondary injury, which is characterized by the creation of an inflammatory and inhibitory microenvironment that contains inhibitors for axon regeneration and repair, such as myelin-associated glycoproteins, reactive astrocytes, activated microglia and infiltrated macrophages [81]. These neuroinflammatory conditions inhibit axon regeneration and negatively influence the activated NSCs to differentiate into neurons [82,83]. Therefore, several treatments targeting neuroinflammation can also positively influence SCECs proliferation and/or differentiation, such as the aforementioned valproic acid. VPA exhibits neuroprotective benefits by reducing SCI-induced apoptosis, neurotoxicity, inflammation and autophagy during the secondary injury period. In addition, VPA upregulates pro-survival neurotrophic proteins, attenuating the inflammatory environment and protecting the remaining neural cells from secondary damage, as reviewed in [41]. Several anti-inflammatory strategies target activated microglia/microphages. One of the key regulators of microglial differentiation is the interaction between colony-stimulating factor 1 (CSF1) with its receptor CSF1R. Therefore, the reduction in activated CD68+ microglia/macrophages using an inhibitor of CSF1 receptor reduced inflammation and led to an increased number of neurons differentiated from SCECs [84]. Similar results can be obtained when shifting the polarization of the macrophage M1 pro-inflammatory phenotype toward the M2 pro-regenerative phenotype. The overexpression of Rictor (an important component of mTOR pathway that is responsible for axonal growth) in spinal cord injury shifted the macrophage polarization around the lesion from the M1 to the M2 phenotype and increased neurogenesis in the lesion epicenter [85].
Immunization with neural-derived peptides (INDP) shifts the inflammatory microenvironment toward a more permissive one, which is characterized by an increase in anti-inflammatory cytokines and the production of neurotrophic factors. These effects are carried out by stimulating an M2 macrophage phenotype. Moreover, a significant increase in neurogenesis, mainly at the central canal and at both the dorsal and ventral horns of INDP-treated animals, was even observed in animals in the chronic stage of SCI [86].
However, neuroinflammation has both beneficial and detrimental effects and there are several aspects that must be taken into consideration. A typical example is a methylprednisolone (MP) steroid, which is commonly used after spinal cord injury in patients for its effect on the attenuation of secondary injury. MP inhibits the activation and proliferation of various inflammatory cell types in animal models of SCI, by reducing the production of inflammatory cytokines/chemokines and free radicals, as well as inhibiting lipid peroxidation. However, it was shown that the application of MP inhibits the proliferation and migration of SCECs and oligodendrocytes after an SCI, not only in rodents but also in nonhuman primates [87].
3.3. Neuromodulation with Physical Factors
Neuromodulation is necessary for the participation of NSCs in neural repair [88]. Researchers recently found that physical factors, such as electric, magnetic and ultrasound effects, can stimulate the activation of stem cells in the CNS [89,90]. A robust increase in NSC proliferation in the adult mouse intact brain was reported after a 2-week application of repeated transcranial magnetic stimulation (rTMS) at both low (1 Hz) and high (30 Hz) frequencies [91]. These experiments were repeated in vitro with similar results, showing that the application of rTMS for 1 week with both 1 and 30 Hz also facilitated NSC proliferation and neuronal differentiation [91]. Furthermore, the very-low-frequency electromagnetic field could activate the excitability of neural progenitor cells and regulate T-type calcium channels, both of which are connected with electrical activity and have the potential formation of neural circuits [92,93]. To date, there is no evidence regarding the ability of rTMS to mobilize SCECs after an SCI. SCEC mobilization was achieved by extracorporeal shock waves, which were applied 4 weeks after an SCI in rats. In the treated animals, increased proliferation of SCECs was detected in the ependymal layer of the central canal and the injured posterior horn. Some limited differentiation into neuronal and glial phenotypes was also reported [94]. Physical factors can be relatively easily translated to human medicine since they are non-invasive and can serve in the future as part of rehabilitation or supportive treatment in patients with spinal cord injury.
3.4. In Vivo Reprogramming
Finally, a new approach regarding how to increase neurogenesis in the injured spinal cord emerged in the last few years with new advances in the reprogramming field. In vivo reprogramming techniques have the potential to convert non-neuronal cells into neurons via the forced expression of transcription factors. Recently, there have been several studies showing that the overexpression of different pro-neural transcription factors can convert endogenous glial cells into neurons (reviewed in [95]). The most often utilized TF is Sox2, a neural progenitor marker that keeps the balance between stem cell renewal and differentiation. The overexpression of Sox2 in astrocytes in the injured spinal cord resulted in the conversion into doublecortin positive neuroblasts. These cells can mature into neurons and connect with endogenous motoneurons. In combination with VPA administration, these cells can survive for 210 days, and the yield of neurons is increased twofold [53]. Interestingly, reprogramming with Sox2 does not skip the proliferating phase of neuroblasts; therefore, one astrocyte can give rise to several neurons. The Sox2 strategy was used not only for reprogramming astrocytes but also NG2 glia. Another advantage appears to be the fact that reprogrammed neurons are not only glutamatergic but also GABAergic [96]. NeuroD1 is another important TF that is needed for neuronal differentiation. The overexpression in astrocytes converts the astrocytes into glutamatergic neurons in a 1:1 ratio. As a pro-survival TF, it also reduces apoptosis in newly generated neurons [97]. Genes can be delivered using different viral vectors. Gene delivery with adeno-associated viruses (AAVs) have several advantages over lentiviruses or retroviruses. Transduction with AAVs does not require proliferating cells; therefore, AAVs can transduce astrocytes after the inflammation peak is over and the acute lesion is already closed. The use of AAVs is therefore suitable for the chronic stage of a SCI. It is easier to reprogram astrocytes in gray matter than in white matter, which is most likely due to neurotrophic support from the endogenous surrounding host neurons [98]. Recently, the type-II-clustered, regularly interspaced, short palindromic repeat and the Cas9 nuclease (CRISPR/Cas9) system from bacteria was utilized for genome editing. The overexpression of the TFs Islet-1 (Isl1), together with Ngn2, can convert astrocytes in the spinal cord gray matter into functional motoneurons [99]. These cells express motoneuron markers, such as CHAT and HB9, and were able to fire action potentials and project their axons into the sciatic nerve to innervate muscles. This technique was used in healthy spinal cords; how effectively these neurons will integrate into host neural circuits or replace damaged motoneurons after spinal cord injury and improve functional outcome needs to be investigated.
In vivo reprogramming has made major progress; however, there are several questions that remain unanswered. The cell population around the lesion is very heterogenous and using different reprogramming factors may not generate identical subtypes of neurons. Moreover, induced neurons may not have the developmental clues for axon guidance to make precise axonal projections within the injured tissue. Worse still, the induced neurons may disrupt host neural circuits or form abnormal ones.
4. Conclusions
The translation of methodology, resulting in the replacement of the depleted population of neurons after spinal cord injury, is an important issue in regenerative medicine. Recent science is currently not able to solve whether neurogenesis in the spinal cord can serve as a source of replacement neurons in human medicine. Some studies completely deny neurogenesis in the spinal cord [78,79]. Most likely, the type and severity of the spinal cord lesion, as well as the microenvironment, play a role in the induction of neurogenesis. Further studies highlight the fact that, in the human adult spinal cord, the spinal canal is not as prominent as in rodents and the remaining ependymal cells do not proliferate; therefore, they cannot serve as a pool for cell replacement [100]. Conversely, other researchers describe the increase in nestin-positive cells in the human spinal cord after injury [80]. In any event, most likely only subtle and advanced techniques that aim at an optimal interventional strategy focused on combined strategies can lead to the eventual success in promoting neurogenesis and/or reconstructing the damaged neuronal circuits and improving functional outcomes. Manipulating the intrinsic properties of ependymal cells in the central canal or gene editing the converting glial cells into neurons together with changing the nonpermissive environment could be the future of personalized medicine for patients with spinal cord injury.
Author Contributions
K.H. designed the review and wrote the manuscript; P.J. designed the review, wrote the manuscript and provided the funding. B.S. designed the figures and wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Czech Science Foundation GACR 19-10365S and by Operational Programme Research, Development and Education in the framework of the project “Center of Reconstructive Neuroscience”, registration number CZ.02.1.01/0.0./0.0/15_003/0000419.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rowland, J.W.; Hawryluk, G.W.; Kwon, B.; Fehlings, M.G. Current status of acute spinal cord injury pathophysiology and emerging therapies: Promise on the horizon. Neurosurg. Focus 2008, 25, E2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahuja, C.S.; Wilson, J.R.; Nori, S.; Kotter, M.R.N.; Druschel, C.; Curt, A.; Fehlings, M.G. Traumatic spinal cord injury. Nat. Rev. Dis. Primers 2017, 3, 17018. [Google Scholar] [CrossRef]
- Venkatesh, K.; Ghosh, S.K.; Mullick, M.; Manivasagam, G.; Sen, D. Spinal cord injury: Pathophysiology, treatment strategies, associated challenges, and future implications. Cell Tissue Res. 2019, 377, 125–151. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.M.; Wu, J.Y.; Li, F.C.; Chen, Q.X. Ion channel blockers and spinal cord injury. J. Neurosci. Res. 2011, 89, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Meldrum, B.S. Glutamate as a neurotransmitter in the brain: Review of physiology and pathology. J. Nutr. 2000, 130, 1007S–1015S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Z.; Zhu, H.; Li, J.; Wang, X.; Misra, H.; Li, Y. Oxidative stress in spinal cord injury and antioxidant-based intervention. Spinal Cord 2012, 50, 264–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christie, S.D.; Comeau, B.; Myers, T.; Sadi, D.; Purdy, M.; Mendez, I. Duration of lipid peroxidation after acute spinal cord injury in rats and the effect of methylprednisolone. Neurosurg. Focus 2008, 25, E5. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Sarkar, C.; Dinizo, M.; Faden, A.I.; Koh, E.Y.; Lipinski, M.M.; Wu, J. Disrupted autophagy after spinal cord injury is associated with er stress and neuronal cell death. Cell Death Dis. 2015, 6, e1582. [Google Scholar] [CrossRef]
- Marsala, J.; Orendacova, J.; Lukacova, N.; Vanicky, I. Traumatic injury of the spinal cord and nitric oxide. Prog. Brain Res. 2007, 161, 171–183. [Google Scholar]
- Bradbury, E.J.; Burnside, E.R. Moving beyond the glial scar for spinal cord repair. Nat. Commun. 2019, 10, 3879. [Google Scholar] [CrossRef]
- Han, Q.; Xie, Y.; Ordaz, J.D.; Huh, A.J.; Huang, N.; Wu, W.; Liu, N.; Chamberlain, K.A.; Sheng, Z.H.; Xu, X.M. Restoring cellular energetics promotes axonal regeneration and functional recovery after spinal cord injury. Cell Metab. 2020, 31, 623–641.e8. [Google Scholar] [CrossRef]
- Beattie, M.S.; Hermann, G.E.; Rogers, R.C.; Bresnahan, J.C. Cell death in models of spinal cord injury. Prog. Brain Res. 2002, 137, 37–47. [Google Scholar]
- Oyinbo, C.A. Secondary injury mechanisms in traumatic spinal cord injury: A nugget of this multiply cascade. Acta Neurobiol. Exp. 2011, 71, 281–299. [Google Scholar]
- Rowlands, D.; Sugahara, K.; Kwok, J.C. Glycosaminoglycans and glycomimetics in the central nervous system. Molecules 2015, 20, 3527–3548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leal-Galicia, P.; Chavez-Hernandez, M.E.; Mata, F.; Mata-Luevanos, J.; Rodriguez-Serrano, L.M.; Tapia-de-Jesus, A.; Buenrostro-Jauregui, M.H. Adult neurogenesis: A story ranging from controversial new neurogenic areas and human adult neurogenesis to molecular regulation. Int. J. Mol. Sci. 2021, 22, 11489. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, O.; Spalding, K.L.; Frisen, J. Adult neurogenesis in humans. Cold Spring Harb. Perspect. Biol. 2015, 7, a018994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nogueira, A.B.; Hoshino, H.S.R.; Ortega, N.C.; Dos Santos, B.G.S.; Teixeira, M.J. Adult human neurogenesis: Early studies clarify recent controversies and go further. Metab. Brain Dis. 2022, 37, 153–172. [Google Scholar] [CrossRef] [PubMed]
- Barnabe-Heider, F.; Goritz, C.; Sabelstrom, H.; Takebayashi, H.; Pfrieger, F.W.; Meletis, K.; Frisen, J. Origin of new glial cells in intact and injured adult spinal cord. Cell Stem Cell 2010, 7, 470–482. [Google Scholar] [CrossRef] [Green Version]
- Adrian, E.K., Jr.; Walker, B.E. Incorporation of thymidine-h3 by cells in normal and injured mouse spinal cord. J. Neuropathol. Exp. Neurol. 1962, 21, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Horner, P.J.; Power, A.E.; Kempermann, G.; Kuhn, H.G.; Palmer, T.D.; Winkler, J.; Thal, L.J.; Gage, F.H. Proliferation and differentiation of progenitor cells throughout the intact adult rat spinal cord. J. Neurosci. 2000, 20, 2218–2228. [Google Scholar] [CrossRef]
- Meletis, K.; Barnabe-Heider, F.; Carlen, M.; Evergren, E.; Tomilin, N.; Shupliakov, O.; Frisen, J. Spinal cord injury reveals multilineage differentiation of ependymal cells. PLoS Biol. 2008, 6, e182. [Google Scholar] [CrossRef] [PubMed]
- Weiss, S.; Dunne, C.; Hewson, J.; Wohl, C.; Wheatley, M.; Peterson, A.C.; Reynolds, B.A. Multipotent cns stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis. J. Neurosci. 1996, 16, 7599–7609. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Nakagomi, N.; Nakano-Doi, A.; Ishikawa, H.; Tatsumi, Y.; Bando, Y.; Yoshikawa, H.; Matsuyama, T.; Gomi, F.; Nakagomi, T. Potential of adult endogenous neural stem/progenitor cells in the spinal cord to contribute to remyelination in experimental autoimmune encephalomyelitis. Cells 2019, 8, 1025. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Lu, Y.; Fang, X.; Zhang, J.; Li, J.; Li, S.; Deng, X.; Yu, Y.; Xu, R. An astrocyte regenerative response from vimentin-containing cells in the spinal cord of amyotrophic lateral sclerosis’s disease-like transgenic (g93a sod1) mice. Neurodegener. Dis. 2015, 15, 1–12. [Google Scholar] [CrossRef]
- Cizkova, D.; Nagyova, M.; Slovinska, L.; Novotna, I.; Radonak, J.; Cizek, M.; Mechirova, E.; Tomori, Z.; Hlucilova, J.; Motlik, J.; et al. Response of ependymal progenitors to spinal cord injury or enhanced physical activity in adult rat. Cell Mol. Neurobiol. 2009, 29, 999–1013. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, B.; Wang, L.; Long, Z.; Li, Y.; Liao, W.; Wu, Y. Endogenous neural stem cells in central canal of adult rats acquired limited ability to differentiate into neurons following mild spinal cord injury. Int. J. Clin. Exp. Pathol. 2015, 8, 3835–3842. [Google Scholar]
- Yang, H.; Lu, P.; McKay, H.M.; Bernot, T.; Keirstead, H.; Steward, O.; Gage, F.H.; Edgerton, V.R.; Tuszynski, M.H. Endogenous neurogenesis replaces oligodendrocytes and astrocytes after primate spinal cord injury. J. Neurosci. 2006, 26, 2157–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhauser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Hamby, M.E.; Sofroniew, M.V. Reactive astrocytes as therapeutic targets for cns disorders. Neurotherapeutics 2010, 7, 494–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabelstrom, H.; Stenudd, M.; Reu, P.; Dias, D.O.; Elfineh, M.; Zdunek, S.; Damberg, P.; Goritz, C.; Frisen, J. Resident neural stem cells restrict tissue damage and neuronal loss after spinal cord injury in mice. Science 2013, 342, 637–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawryluk, G.W.; Mothe, A.J.; Chamankhah, M.; Wang, J.; Tator, C.; Fehlings, M.G. In vitro characterization of trophic factor expression in neural precursor cells. Stem Cells Dev. 2012, 21, 432–447. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, C.P.; Holmstrom, N.A.; Lilja, J.A.; Schweinhardt, P.; Hao, J.; Spenger, C.; Wiesenfeld-Hallin, Z.; Kurpad, S.N.; Frisen, J.; Olson, L. Allodynia limits the usefulness of intraspinal neural stem cell grafts; directed differentiation improves outcome. Nat. Neurosci. 2005, 8, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Shihabuddin, L.S.; Horner, P.J.; Ray, J.; Gage, F.H. Adult spinal cord stem cells generate neurons after transplantation in the adult dentate gyrus. J. Neurosci. 2000, 20, 8727–8735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abematsu, M.; Tsujimura, K.; Yamano, M.; Saito, M.; Kohno, K.; Kohyama, J.; Namihira, M.; Komiya, S.; Nakashima, K. Neurons derived from transplanted neural stem cells restore disrupted neuronal circuitry in a mouse model of spinal cord injury. J. Clin. Investig. 2010, 120, 3255–3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, T.; Zhou, H.; Wang, T.; Lu, L.; Li, F.; Liu, B.; Kong, X.; Feng, S. In vitro characteristics of valproic acid and all-trans-retinoic acid and their combined use in promoting neuronal differentiation while suppressing astrocytic differentiation in neural stem cells. Brain Res. 2015, 1596, 31–47. [Google Scholar] [CrossRef]
- Chu, W.; Yuan, J.; Huang, L.; Xiang, X.; Zhu, H.; Chen, F.; Chen, Y.; Lin, J.; Feng, H. Valproic acid arrests proliferation but promotes neuronal differentiation of adult spinal nspcs from sci rats. Neurochem. Res. 2015, 40, 1472–1486. [Google Scholar] [CrossRef] [PubMed]
- Jope, R.S.; Johnson, G.V. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem. Sci. 2004, 29, 95–102. [Google Scholar] [CrossRef]
- Takahashi-Yanaga, F.; Sasaguri, T. The wnt/beta-catenin signaling pathway as a target in drug discovery. J. Pharmacol. Sci. 2007, 104, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.T.; Wang, Z.; Hunsberger, J.G.; Chuang, D.M. Therapeutic potential of mood stabilizers lithium and valproic acid: Beyond bipolar disorder. Pharmacol. Rev. 2013, 65, 105–142. [Google Scholar] [CrossRef] [Green Version]
- Kostrouchova, M.; Kostrouch, Z.; Kostrouchova, M. Valproic acid, a molecular lead to multiple regulatory pathways. Folia Biol. 2007, 53, 37–49. [Google Scholar]
- Chu, T.; Zhou, H.; Lu, L.; Kong, X.; Wang, T.; Pan, B.; Feng, S. Valproic acid-mediated neuroprotection and neurogenesis after spinal cord injury: From mechanism to clinical potential. Regen. Med. 2015, 10, 193–209. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Jimenez, F.J.; Vilches, A.; Perez-Arago, M.A.; Clemente, E.; Roman, R.; Leal, J.; Castro, A.A.; Fustero, S.; Moreno-Manzano, V.; Jendelova, P.; et al. Activation of neurogenesis in multipotent stem cells cultured in vitro and in the spinal cord tissue after severe injury by inhibition of glycogen synthase kinase-3. Neurotherapeutics 2021, 18, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Li, G.; Ye, J.; Lu, D.; Chen, Z.; Xiang, A.P.; Jiang, M.H. Substance p enhances endogenous neurogenesis to improve functional recovery after spinal cord injury. Int. J. Biochem. Cell Biol. 2017, 89, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Pal, R.; Bondar, V.V.; Adamski, C.J.; Rodney, G.G.; Sardiello, M. Inhibition of erk1/2 restores gsk3beta activity and protein synthesis levels in a model of tuberous sclerosis. Sci. Rep. 2017, 7, 4174. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, Y.; Wang, X.; Evers, B.M. Glycogen synthase kinase-3 is a negative regulator of extracellular signal-regulated kinase. Oncogene 2006, 25, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Chao, X.J.; Wu, J.F.; Cheng, B.C.; Su, T.; Fu, X.Q.; Li, T.; Guo, H.; Tse, A.K.; Kwan, H.Y.; et al. Erk/gsk3beta signaling is involved in atractylenolide i-induced apoptosis and cell cycle arrest in melanoma cells. Oncol. Rep. 2015, 34, 1543–1548. [Google Scholar] [CrossRef]
- Tan, B.T.; Wang, L.; Li, S.; Long, Z.Y.; Wu, Y.M.; Liu, Y. Retinoic acid induced the differentiation of neural stem cells from embryonic spinal cord into functional neurons in vitro. Int. J. Clin. Exp. Pathol. 2015, 8, 8129–8135. [Google Scholar]
- Zhao, H.; Zuo, X.; Ren, L.; Li, Y.; Tai, H.; Du, J.; Xie, X.; Zhang, X.; Han, Y.; Wu, Y.; et al. Combined use of bfgf/egf and all-trans-retinoic acid cooperatively promotes neuronal differentiation and neurite outgrowth in neural stem cells. Neurosci. Lett 2019, 690, 61–68. [Google Scholar] [CrossRef]
- Liu, C.; Sun, R.; Huang, J.; Zhang, D.; Huang, D.; Qi, W.; Wang, S.; Xie, F.; Shen, Y.; Shen, C. The baf45d protein is preferentially expressed in adult neurogenic zones and in neurons and may be required for retinoid acid induced pax6 expression. Front. Neuroanat. 2017, 11, 94. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, J.; Liu, C.; Liu, L.; Shen, Y.; Shen, C.; Liu, C. Baf45d downregulation in spinal cord ependymal cells following spinal cord injury in adult rats and its potential role in the development of neuronal lesions. Front. Neurosci. 2019, 13, 1151. [Google Scholar] [CrossRef] [PubMed]
- Fabbiani, G.; Reali, C.; Valentin-Kahan, A.; Rehermann, M.I.; Fagetti, J.; Falco, M.V.; Russo, R.E. Connexin signaling is involved in the reactivation of a latent stem cell niche after spinal cord injury. J. Neurosci. 2020, 40, 2246–2258. [Google Scholar] [CrossRef]
- Rodriguez-Jimenez, F.J.; Alastrue, A.; Stojkovic, M.; Erceg, S.; Moreno-Manzano, V. Connexin 50 modulates sox2 expression in spinal-cord-derived ependymal stem/progenitor cells. Cell Tissue Res. 2016, 365, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Niu, W.; Liu, M.L.; Zou, Y.; Zhang, C.L. In vivo conversion of astrocytes to neurons in the injured adult spinal cord. Nat Commun. 2014, 5, 3338. [Google Scholar] [CrossRef] [PubMed]
- Nekoui, A.; Blaise, G. Erythropoietin and nonhematopoietic effects. Am. J. Med. Sci. 2017, 353, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Simon, F.H.; Erhart, P.; Vcelar, B.; Scheuerle, A.; Schelzig, H.; Oberhuber, A. Erythropoietin preconditioning improves clinical and histologic outcome in an acute spinal cord ischemia and reperfusion rabbit model. J. Vasc Surg. 2016, 64, 1797–1804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassouna, I.; Ott, C.; Wustefeld, L.; Offen, N.; Neher, R.A.; Mitkovski, M.; Winkler, D.; Sperling, S.; Fries, L.; Goebbels, S.; et al. Revisiting adult neurogenesis and the role of erythropoietin for neuronal and oligodendroglial differentiation in the hippocampus. Mol. Psychiatry 2016, 21, 1752–1767. [Google Scholar] [CrossRef]
- Zhang, H.; Fang, X.; Huang, D.; Luo, Q.; Zheng, M.; Wang, K.; Cao, L.; Yin, Z. Erythropoietin signaling increases neurogenesis and oligodendrogenesis of endogenous neural stem cells following spinal cord injury both in vivo and in vitro. Mol. Med. Rep. 2018, 17, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Keefe, K.M.; Sheikh, I.S.; Smith, G.M. Targeting neurotrophins to specific populations of neurons: Ngf, bdnf, and nt-3 and their relevance for treatment of spinal cord injury. Int. J. Mol. Sci. 2017, 18, 548. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, F.; Cheng, F.; Ying, L.; Wang, C.; Shi, K.; Wang, J.; Xia, K.; Gong, Z.; Huang, X.; et al. Strategies and prospects of effective neural circuits reconstruction after spinal cord injury. Cell Death Dis. 2020, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Kanakasabai, S.; Pestereva, E.; Chearwae, W.; Gupta, S.K.; Ansari, S.; Bright, J.J. Ppargamma agonists promote oligodendrocyte differentiation of neural stem cells by modulating stemness and differentiation genes. PLoS ONE 2012, 7, e50500. [Google Scholar] [CrossRef] [PubMed]
- Jimenez Hamann, M.C.; Tator, C.H.; Shoichet, M.S. Injectable intrathecal delivery system for localized administration of egf and fgf-2 to the injured rat spinal cord. Exp. Neurol. 2005, 194, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Kojima, A.; Tator, C.H. Epidermal growth factor and fibroblast growth factor 2 cause proliferation of ependymal precursor cells in the adult rat spinal cord in vivo. J. Neuropathol. Exp. Neurol. 2000, 59, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Kojima, A.; Tator, C.H. Intrathecal administration of epidermal growth factor and fibroblast growth factor 2 promotes ependymal proliferation and functional recovery after spinal cord injury in adult rats. J. Neurotrauma 2002, 19, 223–238. [Google Scholar] [CrossRef]
- Ohori, Y.; Yamamoto, S.; Nagao, M.; Sugimori, M.; Yamamoto, N.; Nakamura, K.; Nakafuku, M. Growth factor treatment and genetic manipulation stimulate neurogenesis and oligodendrogenesis by endogenous neural progenitors in the injured adult spinal cord. J. Neurosci. 2006, 26, 11948–11960. [Google Scholar] [CrossRef] [Green Version]
- Oudega, M.; Hao, P.; Shang, J.; Haggerty, A.E.; Wang, Z.; Sun, J.; Liebl, D.J.; Shi, Y.; Cheng, L.; Duan, H.; et al. Validation study of neurotrophin-3-releasing chitosan facilitation of neural tissue generation in the severely injured adult rat spinal cord. Exp. Neurol. 2019, 312, 51–62. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, A.; Duan, H.; Zhang, S.; Hao, P.; Ye, K.; Sun, Y.E.; Li, X. Nt3-chitosan elicits robust endogenous neurogenesis to enable functional recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2015, 112, 13354–13359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, J.S.; Zhao, C.; Zhang, A.; Duan, H.; Hao, P.; Wei, R.H.; Shang, J.; Zhao, W.; Liu, Z.; Yu, J.; et al. Nt3-chitosan enables de novo regeneration and functional recovery in monkeys after spinal cord injury. Proc. Natl. Acad. Sci. USA 2018, 115, E5595–E5604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Song, W.; Zhao, W.; Gao, Y.; Shang, J.; Hao, P.; Yang, Z.; Duan, H.; Li, X. Application of the sodium hyaluronate-cntf scaffolds in repairing adult rat spinal cord injury and facilitating neural network formation. Sci. China Life Sci. 2018, 61, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, C.; Haggerty, A.E.; Yan, J.; Lan, M.; Seu, M.; Yang, M.; Marlow, M.M.; Maldonado-Lasuncion, I.; Cho, B.; et al. The effect of a nanofiber-hydrogel composite on neural tissue repair and regeneration in the contused spinal cord. Biomaterials 2020, 245, 119978. [Google Scholar] [CrossRef]
- Liu, H.; Xu, X.; Tu, Y.; Chen, K.; Song, L.; Zhai, J.; Chen, S.; Rong, L.; Zhou, L.; Wu, W.; et al. Engineering microenvironment for endogenous neural regeneration after spinal cord injury by reassembling extracellular matrix. ACS Appl. Mater. Interfaces 2020, 12, 17207–17219. [Google Scholar] [CrossRef] [PubMed]
- Koprivica, V.; Cho, K.S.; Park, J.B.; Yiu, G.; Atwal, J.; Gore, B.; Kim, J.A.; Lin, E.; Tessier-Lavigne, M.; Chen, D.F.; et al. Egfr activation mediates inhibition of axon regeneration by myelin and chondroitin sulfate proteoglycans. Science 2005, 310, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xiao, Z.; Chen, B.; Dai, J. The neuronal differentiation microenvironment is essential for spinal cord injury repair. Organogenesis 2017, 13, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, C.; Li, X.; Xiao, Z.; Zhao, Y.; Liang, H.; Wang, B.; Han, S.; Li, X.; Xu, B.; Wang, N.; et al. A modified collagen scaffold facilitates endogenous neurogenesis for acute spinal cord injury repair. Acta Biomater. 2017, 51, 304–316. [Google Scholar] [CrossRef]
- Fan, C.; Li, X.; Zhao, Y.; Xiao, Z.; Xue, W.; Sun, J.; Li, X.; Zhuang, Y.; Chen, Y.; Dai, J. Cetuximab and taxol co-modified collagen scaffolds show combination effects for the repair of acute spinal cord injury. Biomater. Sci. 2018, 6, 1723–1734. [Google Scholar] [CrossRef]
- Li, X.; Xiao, Z.; Han, J.; Chen, L.; Xiao, H.; Ma, F.; Hou, X.; Li, X.; Sun, J.; Ding, W.; et al. Promotion of neuronal differentiation of neural progenitor cells by using egfr antibody functionalized collagen scaffolds for spinal cord injury repair. Biomaterials 2013, 34, 5107–5116. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, Y.; Cheng, S.; Han, S.; Shu, M.; Chen, B.; Chen, X.; Tang, F.; Wang, N.; Tu, Y.; et al. Cetuximab modified collagen scaffold directs neurogenesis of injury-activated endogenous neural stem cells for acute spinal cord injury repair. Biomaterials 2017, 137, 73–86. [Google Scholar] [CrossRef]
- Lu, T.M.; Houghton, S.; Magdeldin, T.; Durán, J.G.B.; Minotti, A.P.; Snead, A.; Sproul, A.; Nguyen, D.T.; Xiang, J.; Fine, H.A.; et al. Pluripotent stem cell-derived epithelium misidentified as brain microvascular endothelium requires ETS factors to acquire vascular fate. Proc Natl Acad Sci USA 2021, 118, e2016950118. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Ao, Y.; O’Shea, T.M.; Burda, J.E.; Bernstein, A.M.; Brumm, A.J.; Muthusamy, N.; Ghashghaei, H.T.; Carmichael, S.T.; Cheng, L.; et al. Ependymal cell contribution to scar formation after spinal cord injury is minimal, local and dependent on direct ependymal injury. Sci. Rep. 2017, 7, 41122. [Google Scholar] [CrossRef] [Green Version]
- Shah, P.T.; Stratton, J.A.; Stykel, M.G.; Abbasi, S.; Sharma, S.; Mayr, K.A.; Koblinger, K.; Whelan, P.J.; Biernaskie, J. Single-cell transcriptomics and fate mapping of ependymal cells reveals an absence of neural stem cell function. Cell 2018, 173, 1045–1057.e1049. [Google Scholar] [CrossRef] [Green Version]
- Cawsey, T.; Duflou, J.; Weickert, C.S.; Gorrie, C.A. Nestin-positive ependymal cells are increased in the human spinal cord after traumatic central nervous system injury. J. Neurotrauma 2015, 32, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.P.; Coulter, M.; Miotke, J.; Meyer, R.L.; Takemaru, K.; Levine, J.M. Abrogation of beta-catenin signaling in oligodendrocyte precursor cells reduces glial scarring and promotes axon regeneration after cns injury. J. Neurosci. 2014, 34, 10285–10297. [Google Scholar] [CrossRef] [Green Version]
- Lukovic, D.; Stojkovic, M.; Moreno-Manzano, V.; Jendelova, P.; Sykova, E.; Bhattacharya, S.S.; Erceg, S. Concise review: Reactive astrocytes and stem cells in spinal cord injury: Good guys or bad guys? Stem Cells 2015, 33, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, W.S.; Abdel-Hamid, G.A.; Al-Karim, S.; Zakar, N.; Elassouli, M.Z. Neuroectodermal stem cells: A remyelinating potential in acute compressed spinal cord injury in rat model. J. Biosci. 2018, 43, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Deng, M.; Zhao, X.Q.; Liu, M. Alternatively polarized macrophages regulate the growth and differentiation of ependymal stem cells through the sirt2 pathway. Exp. Neurobiol. 2020, 29, 150–163. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, P.; Liu, X.; Li, J.; Wan, Y.; Liu, S.; Wei, F. Overexpression of rictor in the injured spinal cord promotes functional recovery in a rat model of spinal cord injury. FASEB J. 2020, 34, 6984–6998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Barrera, R.; Flores-Romero, A.; Garcia, E.; Fernandez-Presas, A.M.; Incontri-Abraham, D.; Navarro-Torres, L.; Garcia-Sanchez, J.; Juarez-Vignon Whaley, J.J.; Madrazo, I.; Ibarra, A. Immunization with neural-derived peptides increases neurogenesis in rats with chronic spinal cord injury. CNS Neurosci. Ther. 2020, 26, 650–658. [Google Scholar] [CrossRef]
- Ye, J.; Qin, Y.; Tang, Y.; Ma, M.; Wang, P.; Huang, L.; Yang, R.; Chen, K.; Chai, C.; Wu, Y.; et al. Methylprednisolone inhibits the proliferation of endogenous neural stem cells in nonhuman primates with spinal cord injury. J. Neurosurg. Spine 2018, 29, 199–207. [Google Scholar] [CrossRef]
- Zheng, Y.; Mao, Y.R.; Yuan, T.F.; Xu, D.S.; Cheng, L.M. Multimodal treatment for spinal cord injury: A sword of neuroregeneration upon neuromodulation. Neural Regen. Res. 2020, 15, 1437–1450. [Google Scholar] [PubMed]
- Blackmore, J.; Shrivastava, S.; Sallet, J.; Butler, C.R.; Cleveland, R.O. Ultrasound neuromodulation: A review of results, mechanisms and safety. Ultrasound Med. Biol. 2019, 45, 1509–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, M.; Ge, H.; Zhao, H.; Zou, Y.; Chen, Y.; Feng, H. Electromagnetic fields for the regulation of neural stem cells. Stem Cells Int. 2017, 2017, 9898439. [Google Scholar] [CrossRef] [Green Version]
- Abbasnia, K.; Ghanbari, A.; Abedian, M.; Ghanbari, A.; Sharififar, S.; Azari, H. The effects of repetitive transcranial magnetic stimulation on proliferation and differentiation of neural stem cells. Anat. Cell Biol. 2015, 48, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Liu, X.; Yang, T.; Mei, Y.A.; Hu, C. Exposure to extremely low-frequency electromagnetic fields inhibits t-type calcium channels via aa/lte4 signaling pathway. Cell Calcium 2014, 55, 48–58. [Google Scholar] [CrossRef] [Green Version]
- He, Y.L.; Liu, D.D.; Fang, Y.J.; Zhan, X.Q.; Yao, J.J.; Mei, Y.A. Exposure to extremely low-frequency electromagnetic fields modulates na+ currents in rat cerebellar granule cells through increase of aa/pge2 and ep receptor-mediated camp/pka pathway. PLoS ONE 2013, 8, e54376. [Google Scholar] [CrossRef] [Green Version]
- Shin, D.C.; Ha, K.Y.; Kim, Y.H.; Kim, J.W.; Cho, Y.K.; Kim, S.I. Induction of endogenous neural stem cells by extracorporeal shock waves after spinal cord injury. Spine 2018, 43, E200–E207. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, H. Neuronal reprogramming in treating spinal cord injury. Neural Regen. Res. 2022, 17, 1440–1445. [Google Scholar] [PubMed]
- Tai, W.; Wu, W.; Wang, L.L.; Ni, H.; Chen, C.; Yang, J.; Zang, T.; Zou, Y.; Xu, X.M.; Zhang, C.L. In vivo reprogramming of ng2 glia enables adult neurogenesis and functional recovery following spinal cord injury. Cell Stem Cell 2021, 28, 923–937.e4. [Google Scholar] [CrossRef] [PubMed]
- Puls, B.; Ding, Y.; Zhang, F.; Pan, M.; Lei, Z.; Pei, Z.; Jiang, M.; Bai, Y.; Forsyth, C.; Metzger, M.; et al. Regeneration of functional neurons after spinal cord injury via in situ neurod1-mediated astrocyte-to-neuron conversion. Front. Cell Dev. Biol. 2020, 8, 591883. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.L.; Su, Z.; Tai, W.; Zou, Y.; Xu, X.M.; Zhang, C.L. The p53 pathway controls sox2-mediated reprogramming in the adult mouse spinal cord. Cell Rep. 2016, 17, 891–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Tao, X.; Sui, M.; Cui, M.; Liu, D.; Wang, B.; Wang, T.; Zheng, Y.; Luo, J.; Mu, Y.; et al. Reprogramming astrocytes to motor neurons by activation of endogenous ngn2 and isl1. Stem Cell Rep. 2021, 16, 1777–1791. [Google Scholar] [CrossRef]
- Paniagua-Torija, B.; Norenberg, M.; Arevalo-Martin, A.; Carballosa-Gautam, M.M.; Campos-Martin, Y.; Molina-Holgado, E.; Garcia-Ovejero, D. Cells in the adult human spinal cord ependymal region do not proliferate after injury. J. Pathol. 2018, 246, 415–421. [Google Scholar] [CrossRef]
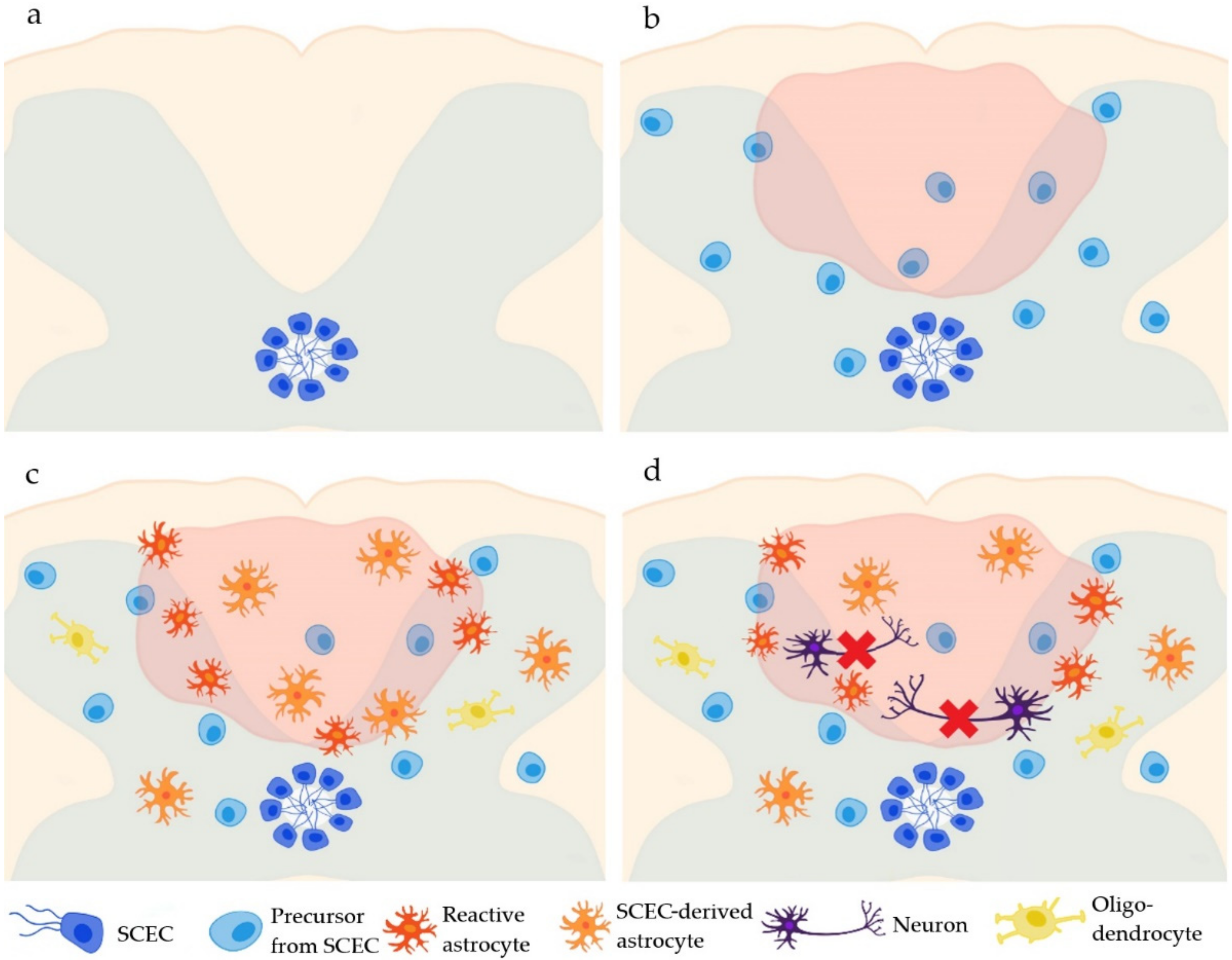
Figure 1.
The fate of the ependymal cells of the central canal in the spinal cord (SCECs). Under physiological conditions, SCECs border the central canal of the spinal cord (a). After an SCI, SCECs start to proliferate (b) and differentiate mostly into astrocytes (2 weeks post-injury) and partly into oligodendrocytes (4 months post-injury) (c), but not into neurons (d).
Figure 1.
The fate of the ependymal cells of the central canal in the spinal cord (SCECs). Under physiological conditions, SCECs border the central canal of the spinal cord (a). After an SCI, SCECs start to proliferate (b) and differentiate mostly into astrocytes (2 weeks post-injury) and partly into oligodendrocytes (4 months post-injury) (c), but not into neurons (d).

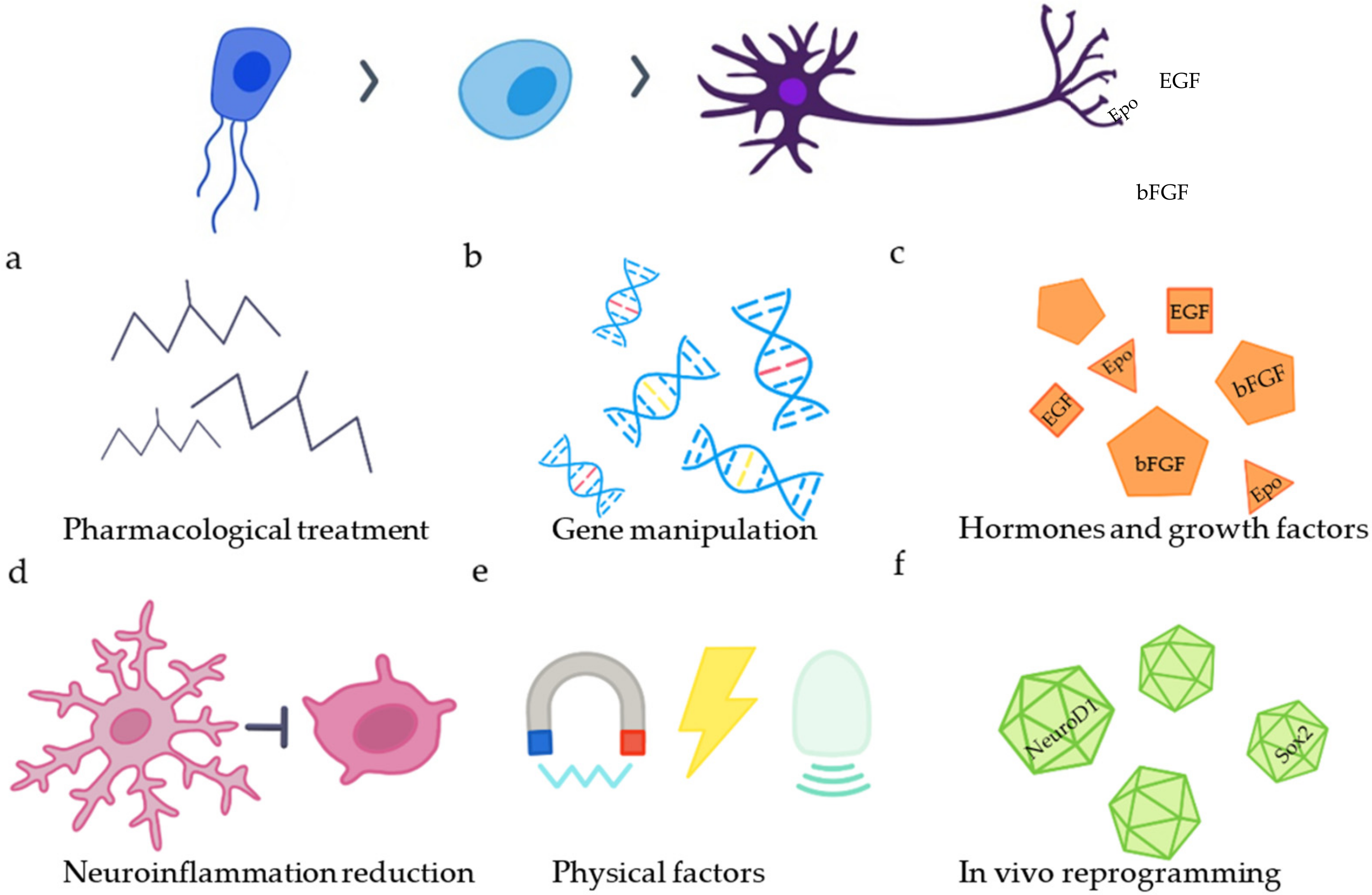
Figure 2.
Strategies to increase endogenous neurogenesis. The differentiation of SCECs into neurons can be enhanced by (a) application of different drugs (VPA, RA, Ro3303544), (b) manipulation of genes (BAF45D peptide, connexin 50), (c) application of hormones or growth factors (EGF, bFGF, Epo, substance P), (d) reduction in neuroinflammation, (e) application of physical factors and (f) in vivo reprogramming of reactive astrocytes and NG2 glia.
Figure 2.
Strategies to increase endogenous neurogenesis. The differentiation of SCECs into neurons can be enhanced by (a) application of different drugs (VPA, RA, Ro3303544), (b) manipulation of genes (BAF45D peptide, connexin 50), (c) application of hormones or growth factors (EGF, bFGF, Epo, substance P), (d) reduction in neuroinflammation, (e) application of physical factors and (f) in vivo reprogramming of reactive astrocytes and NG2 glia.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Havelikova, K.; Smejkalova, B.; Jendelova, P. Neurogenesis as a Tool for Spinal Cord Injury. Int. J. Mol. Sci. 2022, 23, 3728. https://doi.org/10.3390/ijms23073728
AMA Style
Havelikova K, Smejkalova B, Jendelova P. Neurogenesis as a Tool for Spinal Cord Injury. International Journal of Molecular Sciences. 2022; 23(7):3728. https://doi.org/10.3390/ijms23073728
Chicago/Turabian StyleHavelikova, Katerina, Barbora Smejkalova, and Pavla Jendelova. 2022. "Neurogenesis as a Tool for Spinal Cord Injury" International Journal of Molecular Sciences 23, no. 7: 3728. https://doi.org/10.3390/ijms23073728
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.