
Asian Sand Dust Particles Enhance the Development of Aspergillus fumigatus Biofilm on Nasal Epithelial Cells



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
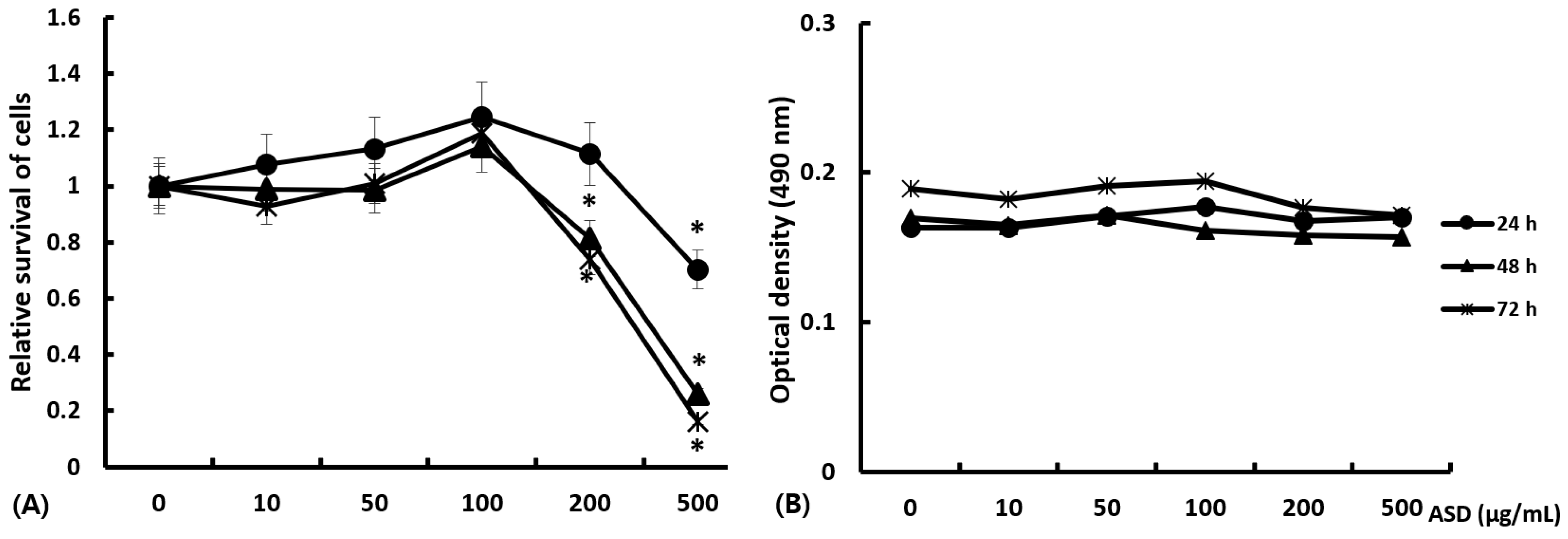
2.1. Cytotoxic Effects of ASD on Nasal Epithelial Cells
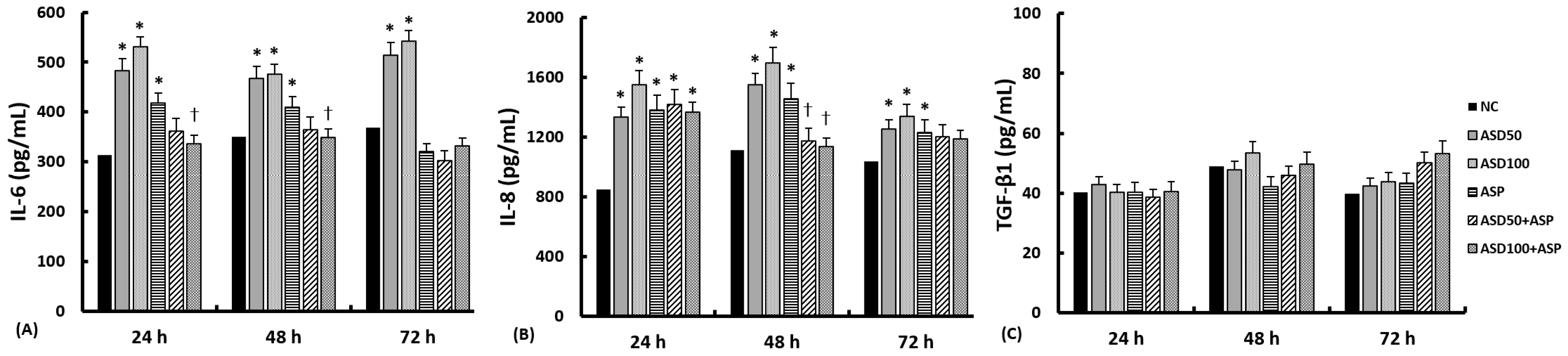
2.2. Effects of ASD and A. fumigatus on the Production of Inflammatory Chemical Mediators from Nasal Epithelial Cells
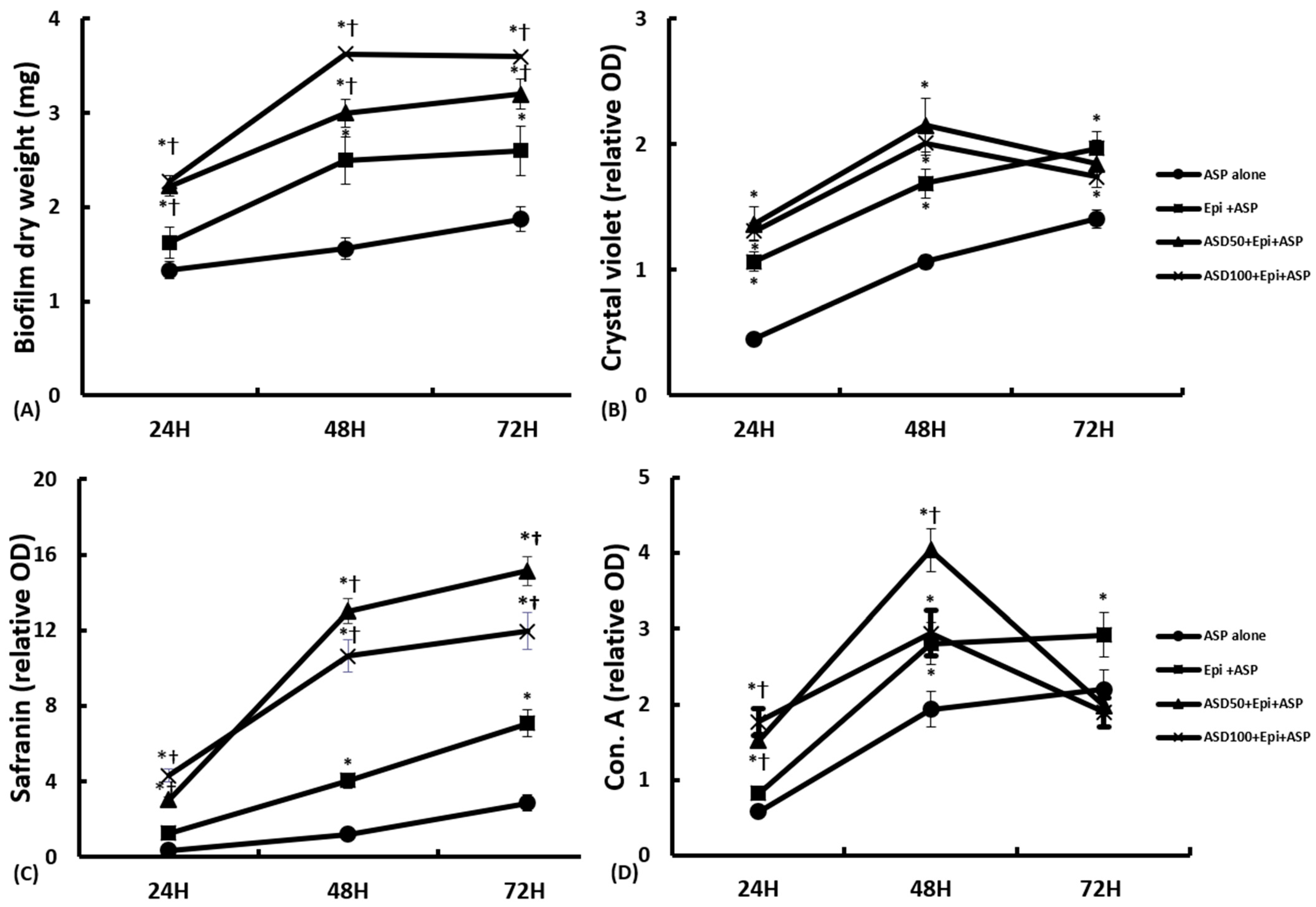
2.3. Effects of ASD on the Development of Biofilms on Nasal Epithelial Cells
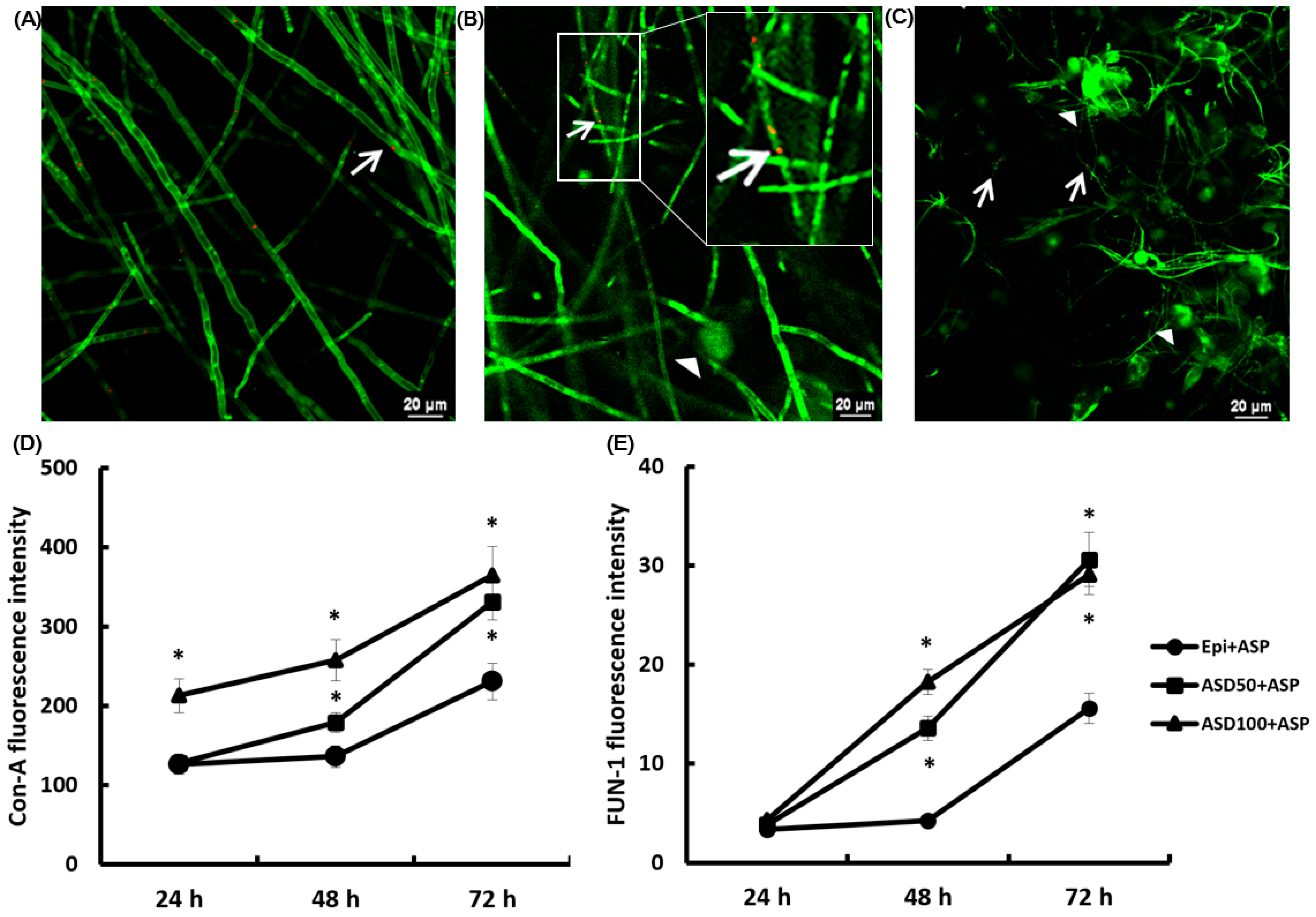
2.4. Confocal Laser Scanning Microscopic Findings
3. Discussion
4. Materials and Methods
4.1. Preparation of ASD and Aspergillus fumigatus Spores
4.2. Primary Nasal Epithelial Cell Culture and Cell Viability Assays
4.3. Analysis of Inflammatory Chemical Mediator Protein Production
4.4. Biofilm Dry Weight
4.5. Crystal Violet Staining
4.6. Safranin and Concanavalin A Staining
4.7. Confocal Laser Scanning Microscopy
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osherov, N. Interaction of the pathogenic mold Aspergillus fumigatus with lung epithelial cells. Front. Microbiol. 2012, 3, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldman, M.B.; Dutko, R.A.; Wood, M.A.; Ward, R.A.; Leung, H.M.; Snow, R.F.; De La Flor, D.J.; Yonker, L.M.; Reedy, J.L.; Tearney, G.J.; et al. Aspergillus fumigatus cell wall promotes apical airway epithelial recruitment of human neutrophils. Infect. Immun. 2020, 88, e00813-19. [Google Scholar] [CrossRef] [PubMed]
- Willinger, B.; Obradovic, A.; Selitsch, B.; Beck-Mannagetta, J.; Buzina, W.; Braun, H.; Apfalter, P.; Hirschl, A.M.; Makristathis, A.; Rotter, M. Detection and identification of fungi from fungus balls of the maxillary sinus by molecular techniques. J. Clin. Microbiol. 2003, 41, 581–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beauvais, A.; Fontaine, T.; Aimanianda, V.; Latge, J.P. Aspergillus cell wall and biofilm. Mycopathologia 2014, 178, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dong, D.; Cheng, J.; Fan, X.; Zhao, Y. Relationship between biofilms and clinical features in patients with sinus fungal ball. Eur. Arch. Otorhinolaryngol. 2015, 272, 2363–2369. [Google Scholar] [CrossRef]
- Foreman, A.; Psaltis, A.J.; Tan, L.W.; Wormald, P.J. Characterization of bacterial and fungal biofilms in chronic rhinosinusitis. Am. J. Rhinol. Allergy 2009, 23, 556–561. [Google Scholar] [CrossRef]
- Ichinose, T.; Yoshida, S.; Hiyoshi, K.; Sadakane, K.; Takano, H.; Nishikawa, M.; Mori, I.; Yanagisawa, R.; Kawazato, H.; Yasuda, A.; et al. The effects of microbial materials adhered to Asian sand dust on allergic lung inflammation. Arch. Environ. Contam. Toxicol. 2008, 55, 348–357. [Google Scholar] [CrossRef]
- Shin, S.H.; Ye, M.K.; Hwang, Y.J.; Kim, S.T. The effect of Asian sand dust-activated respiratory epithelial cells on activation and migration of eosinophils. Inhal. Toxicol. 2013, 25, 633–639. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhang, X.; Wang, J.; Dandekar, A.; Kim, H.; Qiu, Y.; Xu, X.; Cui, Y.; Wang, A.; Chen, L.C.; et al. Exposure to fine airborne particulate matters induces hepatic fibrosis in murine models. J. Hepatol. 2015, 63, 1397–1404. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Guo, Z.; Zhang, R.; Deng, C.; Xu, J.; Dong, W.; Hong, Z.; Yu, H.; Situ, H.; Liu, C.; et al. Nasal epithelial barrier disruption by particulate matter ≤2.5 um via tight junction protein degradation. J. Appl. Toxicol. 2018, 38, 678–687. [Google Scholar] [CrossRef]
- Lei, Y.C.; Chan, C.C.; Wang, P.Y.; Lee, C.T.; Cheng, T.J. Effects of Asian dust event particles on inflammation markers in peripheral blood and bronchoalveolar lavage in pulmonary hypertensive rats. Environ. Res. 2004, 95, 71–76. [Google Scholar] [CrossRef]
- Lee, J.S.; Shin, S.Y.; Lee, K.H.; Kim, S.W.; Cho, J.S. Change of prevalence and clinical aspects of fungal ball according to temporal difference. Eur. Arch. Otorhinolaryngol. 2013, 270, 1673–1677. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.W.; Kim, Y.M.; Min, J.Y.; Kim, J.W.; Kim, J.K.; Mo, J.H.; Shin, J.M.; Cho, K.S.; Kwak, S.G.; Shin, S.H. Clinicopathologic characteristics of paranasal sinus fungus ball: Retrospective, multicenter study in Korea. Eur. Arch. Otorhinolaryngol. 2020, 277, 761–765. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Go, Y.Y.; Chae, S.W.; Park, M.K.; Song, J.J. Asian Sand Dust Particles Increased Pneumococcal Biofilm Formation in vitro and Colonization in Human Middle Ear Epithelial Cells and Rat Middle Ear Mucosa. Front. Genet. 2020, 11, 323. [Google Scholar] [CrossRef]
- Doll, K.; Jongsthaphongpun, K.L.; Stumpp, N.S.; Winkel, A.; Stiesch, M. Quantifying implant-associated biofilms: Comparison of microscopic, microbiologic and biochemical methods. J. Microbiol. Methods 2016, 130, 61–68. [Google Scholar] [CrossRef]
- Stiefel, P.; Rosenberg, U.; Schneider, J.; Mauerhofer, S.; Maniura-Weber, K.; Ren, Q. Is biofilm removal properly assessed? Comparison of different quantification methods in a 96-well plate system. Appl. Microbiol. Biotechnol. 2016, 100, 4135–4145. [Google Scholar] [CrossRef] [Green Version]
- Tenorio, D.P.; Andrade, C.G.; Filho, P.E.C.; Sabino, C.P.; Kato, I.T.; Carvalho, L.B., Jr.; Alves, S., Jr.; Ribeiro, M.S.; Fontes, A.; Santos, B.S. CdTe quantum dots conjugated to concanavalin A as potential fluorescent molecular probes for saccharides detection in Candida albicans. J. Photochem. Photobiol. B 2015, 142, 237–243. [Google Scholar] [CrossRef]
- Honda, A.; Matsuda, Y.; Murayama, R.; Tsuji, K.; Nishikawa, M.; Koike, E.; Yoshida, S.; Ichinose, T.; Takano, H. Effects of Asian sand dust particles on the respiratory and immune system. J. Appl. Toxicol. 2014, 34, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Bautista, M.V.; Chen, Y.; Ivanova, V.S.; Rahimi, M.K.; Watson, A.M.; Rose, M.C. IL-8 regulates mucin gene expression at the posttranscriptional level in lung epithelial cells. J. Immunol. 2009, 183, 2159–2166. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.S.; Bae, C.H.; Song, S.Y.; Kim, Y.D. Asian sand dust increases MUC8 and MUC5B expressions via TLR4-dependent ERK2 and p38 MAPK in human airway epithelial cells. Am. J. Rhinol. Allergy 2015, 29, 161–165. [Google Scholar] [CrossRef]
- Gauthier, T.; Wang, X.; Sifuentes Dos Santos, J.; Fysikopoulos, A.; Tadrist, S.; Canlet, C.; Artigot, M.P.; Loiseau, N.; Oswald, I.P.; Puel, O. Trypacidin, a spore-borne toxin from Aspergillus fumigatus, is cytotoxic to lung cells. PLoS ONE 2012, 7, e29906. [Google Scholar] [CrossRef]
- Hohl, T.M.; Feldmesser, M. Aspergillus fumigatus: Principles of pathogenesis and host defense. Eukaryot. Cell 2007, 6, 1953–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidler, M.J.; Salvenmoser, S.; Muller, F.M. Aspergillus fumigatus forms biofilms with reduced antifungal drug susceptibility on bronchial epithelial cells. Antimicrob. Agents Chemother. 2008, 52, 4130–4136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuero, R.; Ouellet, T.; Yu, J.; Mogongwa, N. Metal ion enhancement of fungal growth, gene expression and aflatoxin synthesis in Aspergillus flavus: RT-PCR characterization. J. Appl. Microbiol. 2003, 94, 953–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, S.H.; Ye, M.K.; Lee, D.W.; Chae, M.H.; Han, B.D. Nasal Epithelial Cells Activated with Alternaria and House Dust Mite Induce Not Only Th2 but Also Th1 Immune Responses. Int. J. Mol. Sci. 2020, 21, 2693. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-H.; Ye, M.-K.; Lee, D.-W.; Chae, M.-H. Asian Sand Dust Particles Enhance the Development of Aspergillus fumigatus Biofilm on Nasal Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 3030. https://doi.org/10.3390/ijms23063030
Shin S-H, Ye M-K, Lee D-W, Chae M-H. Asian Sand Dust Particles Enhance the Development of Aspergillus fumigatus Biofilm on Nasal Epithelial Cells. International Journal of Molecular Sciences. 2022; 23(6):3030. https://doi.org/10.3390/ijms23063030
Chicago/Turabian StyleShin, Seung-Heon, Mi-Kyung Ye, Dong-Won Lee, and Mi-Hyun Chae. 2022. "Asian Sand Dust Particles Enhance the Development of Aspergillus fumigatus Biofilm on Nasal Epithelial Cells" International Journal of Molecular Sciences 23, no. 6: 3030. https://doi.org/10.3390/ijms23063030