1. Introduction
Diabetes has reached pandemic proportions worldwide. Complications of diabetes, such as impaired wound healing, represent a significant medical problem, with the annual cost of diabetic lower extremity ulcers alone exceeding 1.5 billion dollars [
1]. These chronic wounds result in significant morbidity for individuals, including long hospitalizations, prolonged exposure to antibiotics, acute and chronic pain, the need for cumbersome wound care, and restricted mobility. In addition, an ulcer of the lower extremity precedes 84% of all diabetic lower extremity amputations, and is the primary cause for hospitalization among diabetics [
2]. Despite the enormous impact of these chronic wounds on both individuals and society, effective therapies are lacking. Thus, the modification, correction, or prevention of diabetes impaired wound healing has far-reaching consequences, both on patient outcomes and on healthcare expenditures.
Normal wound repair follows an orderly and well-defined sequence of events that requires the interaction of many cell types, such as inflammatory cells, fibroblasts, keratinocytes, endothelial cells and progenitor cells, as well as the involvement of many growth factors, extracellular matrix (ECM) proteins, and enzymes. In diabetic wound healing, this complex orchestration of wound healing processes is disrupted. This impairment is associated with the significantly decreased production of granulation tissue and an increased epithelial gap, compared to non-diabetic wounds [
3,
4,
5,
6,
7]. The production of granulation tissue is dependent on the formation of new vessels in the wound bed, synthesis of extracellular matrix (ECM), and provides the substrate for epithelial cell migration and subsequent wound closure.
The stromal cell-derived factor1α (SDF-1α) is a CXC chemokine, important in the mobilization and recruitment of hematopoietic progenitor cells and other CXCR4+ cells to bone marrow and other tissues [
8]. SDF-1α secretion has also been shown to be upregulated by hypoxia inducible factor (HIF-1α) and its expression is increased in areas of tissue injury [
9,
10]. These observations have led to suggestions that SDF-1α may also have a central role in directing cells to injured tissues to facilitate tissue repair.
On analysis, we found that diabetic wounds produced significantly less SDF-1α both at an mRNA and protein level [
3] compared to normal wound tissue. We have previously shown that the local application of Mesenchymal Stem Cells (MSCs) enhances wound closure in a diabetic mouse model [
11]. We found that MSC-treated wounds had a decreased inflammation response, decreased epithelial gap, and increased vessel density [
11]. Furthermore, this effect appears to be due, in part, to the increased production of SDF-1α in wounds [
11].
In a gain of function experiment, we overexpressed SDF-1α by lentivirus vector, and found that SDF1 treatment resulted in greater granulation tissue, a smaller epithelial gap, and a smaller wound size in diabetic wounds [
3]. In a loss of function experiment, we injected a lentiviral vector that expresses a mutant form of SDF-1α that binds, but does not activate, CXC chemokine receptor type 4 (CXCR4) and measured its effect on granulation tissue formation, angiogenesis, inflammation, cell migration, and wound healing. We found that competitive inhibition of SDF-1α significantly impairs the rate of wound healing, decreases angiogenesis, and increases inflammation in the diabetic mouse [
12].
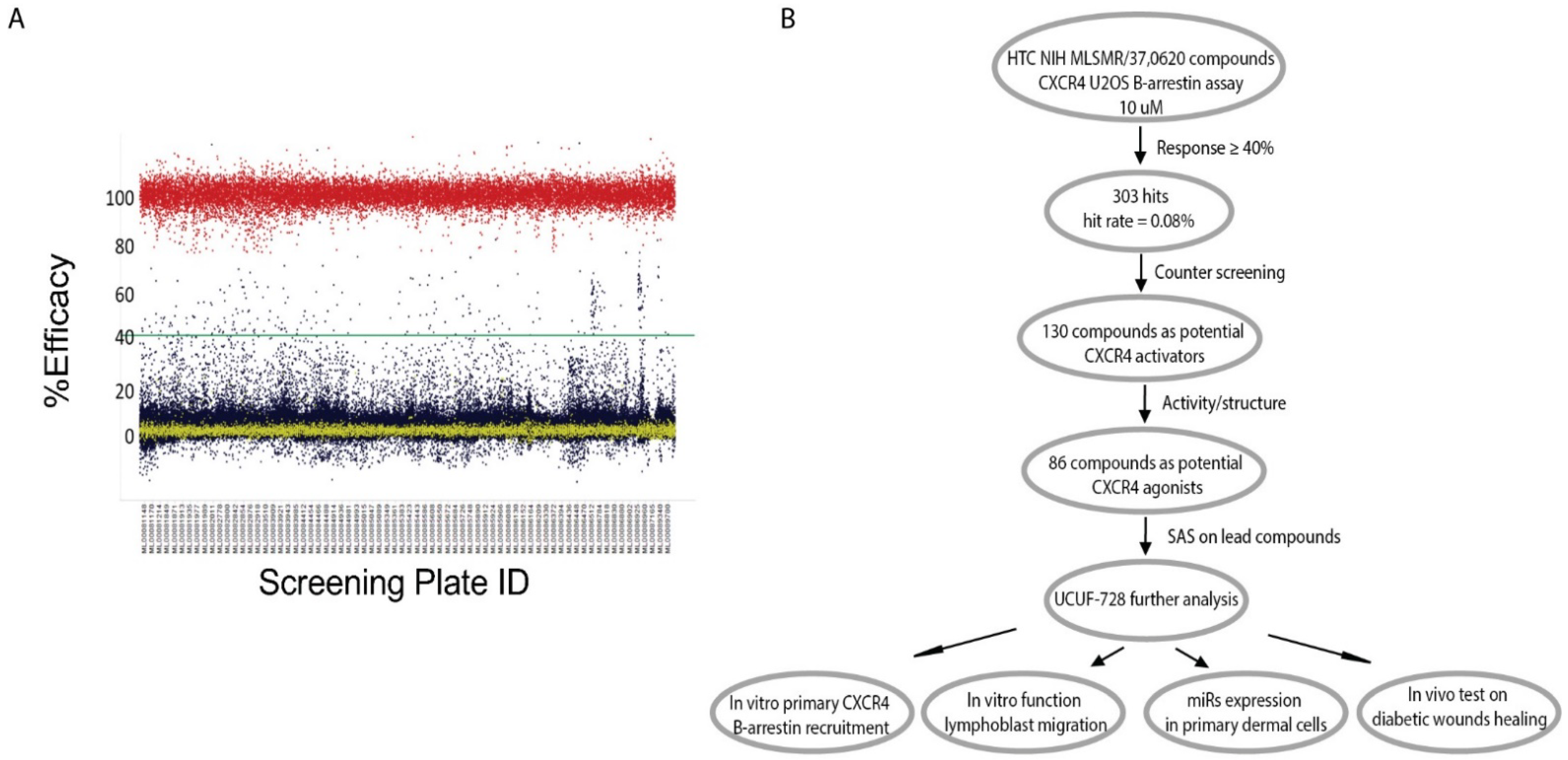
In summary, previous data indicated that SDF-1α is a key factor in the wound-healing process that could be targeted to correct the diabetic wound-healing defect. Therefore, screening for small molecule positive modulators that can activate the CXCR4 receptor and its downstream signaling pathway, thereby blocking SDF-1α response, which will provide a novel topical therapy for diabetic wound healing with a reduced risk for systemic receptor activation, and has a great potential for clinical application and commercialization.
3. Discussion
Secretion of the chemokine SDF-1α, with the subsequent activation of the CXCR4 receptor, is a critical component for effective wound healing [
3,
4,
17]. It is a chemokine that promotes the recruitment of hematopoietic progenitor cells to areas of tissue injury [
9,
10]. The expression of SDF-1α is decreased in diabetic wounds, which may underlie the wound healing impairment depicted by an increased wound closure time, decreased granulation tissue, and a larger epithelial gap [
3,
4,
5,
6,
7]. Reduced cellular migration resulting from decreased SDF-1a expression may contribute to observed healing impairment, as non-diabetic mouse wounds lacking lymphocytes recapitulated features of impaired wound healing, including preferential M1 polarization, increased basal ROS levels, and reduced angiogenesis [
18]. Theses mechanistic insights highlight the utility of exploring novel therapeutics that can circumvent deficits in CXCR4 receptor activation to correct healing impairment.
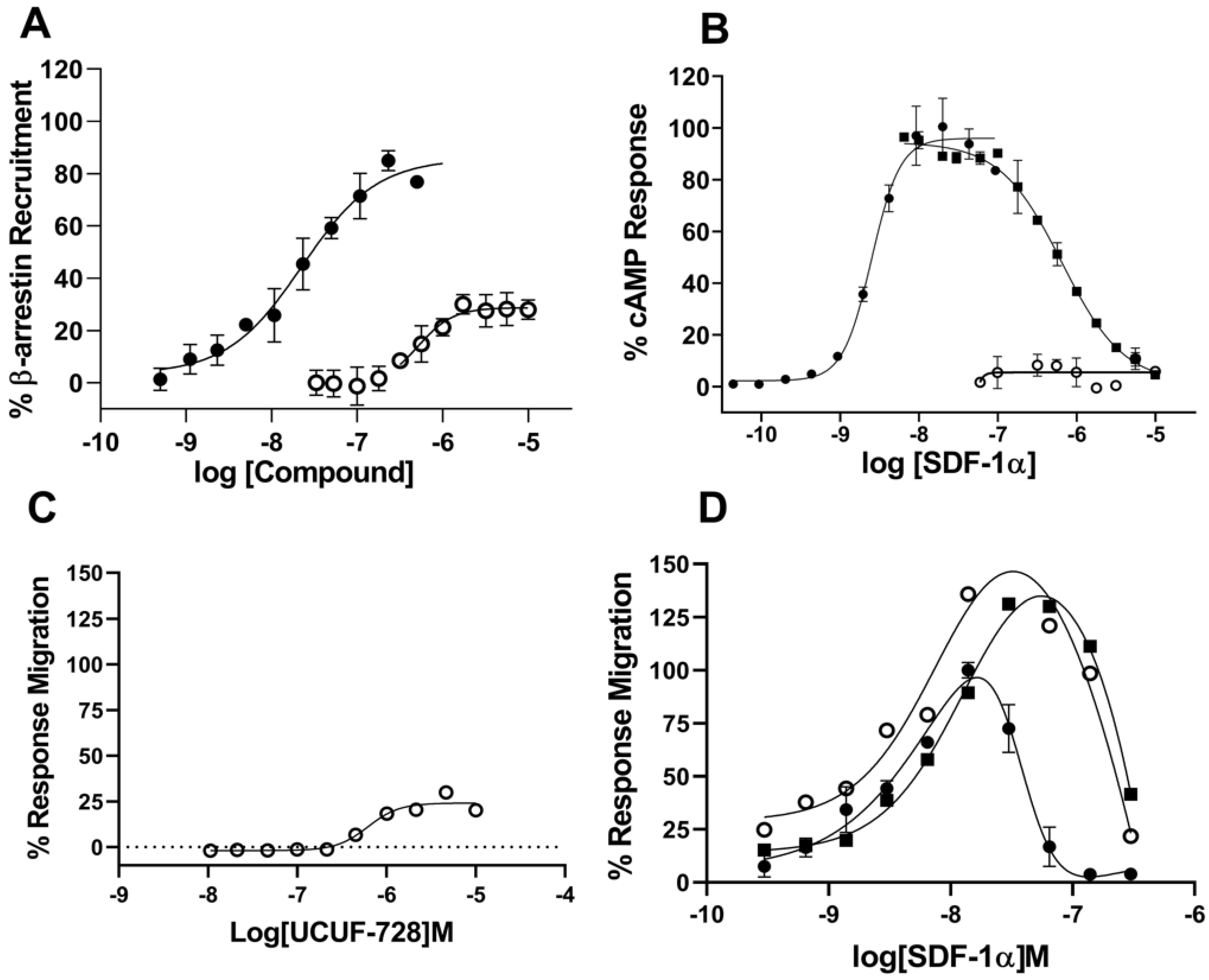
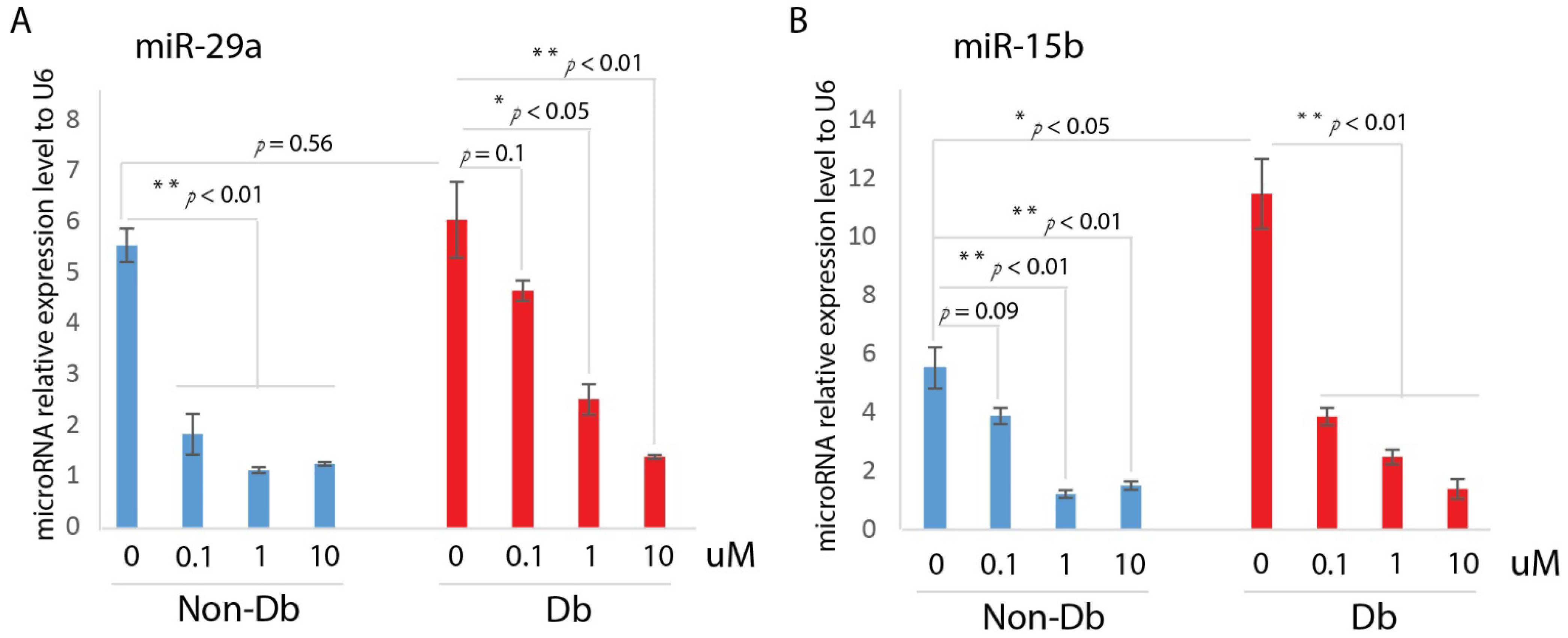
Here, we developed a high-throughput β-arrestin recruitment assay to screen compounds for potential utility as CXCR4 receptor activators. The subsequent structure–activity relationship (SAR) studies identified a chemical scaffold that functions as a CXCR4 agonist (UCUF-728), and further activity was confirmed with in vitro and in vivo validation studies. The treatment of human diabetic fibroblasts with UCUF-728 resulted in the potent suppression of miR-15b expression, an outcome observed with the lowest concentration (0.1 μM). MiR-15b is a negative modulator of angiogenesis [
19,
20] that is upregulated in diabetic wounds during the early phase of healing (Xu et al., 2014). An increased expression of miR-15b is associated with a decreased expression of proangiogenic target genes, including vascular endothelial growth factor (VEGFα), hypoxia inducible factor (HIF-1α), and B-cell lymphoma 2 (BCL2) [
7]. As previously demonstrated, the therapeutic suppression of miR-15b expression in diabetic wounds may contribute to accelerated wound closure by enhancing angiogenesis [
7]. UCUF-728 treatment also reduced miR-29a expression in diabetic fibroblasts, but relative suppression was less potent than that observed of miR-15b. MiR-29a is upregulated in human and murine diabetic skin [
16]. Evidence suggests that the dysregulation of miR-29a contributes to decreased collagen I protein content in diabetic wounds, leading to impaired biomechanical properties of skin that may underlie an increased susceptibility to injury [
16].
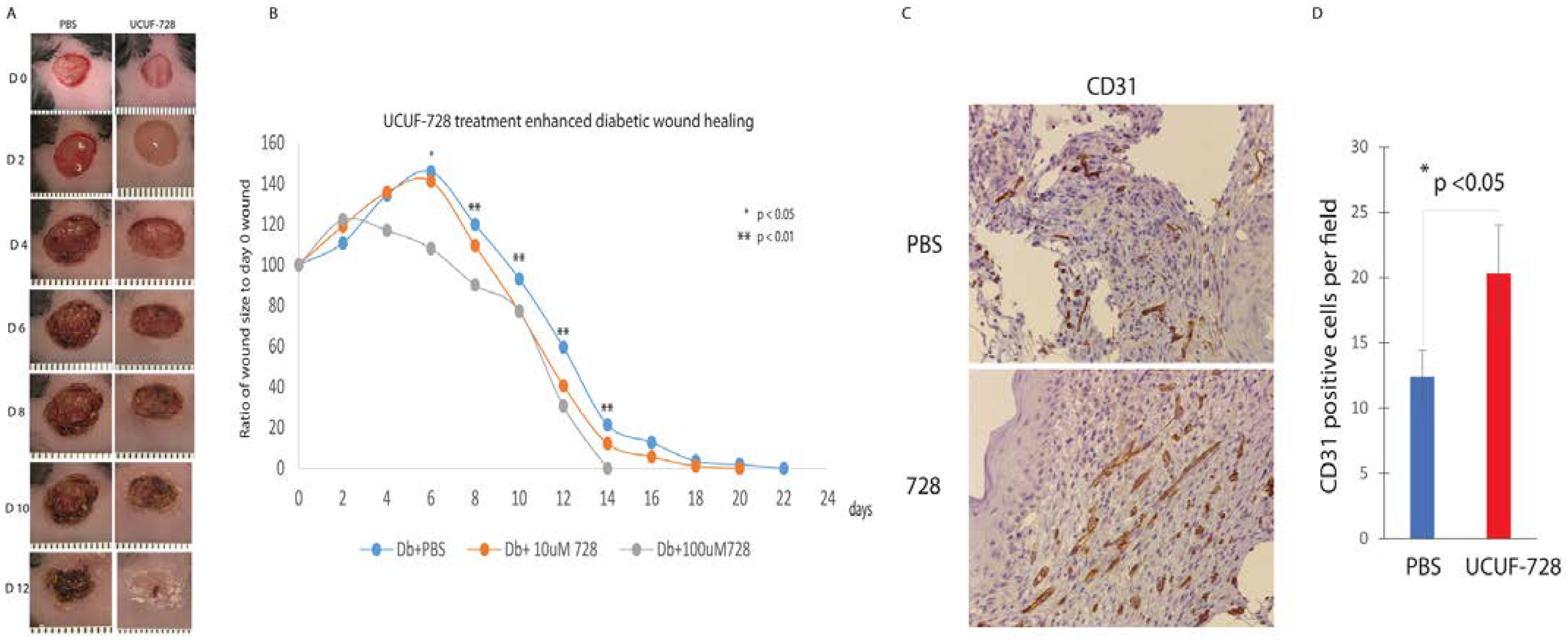
In vivo, UCUF-728 treatment reduced wound closure time by 36%, which was observed. This data indicated that the topical application of UCUF-728 can accelerate diabetic wound healing, limiting the risks with systemic CXCR4 activation. Enhanced angiogenesis was also found in treated diabetic wounds. Together, these studies suggest that the activation of CXCR4 receptors with UCUF-728 accelerates wound healing by favoring the promotion of angiogenesis via the suppression of miR-15b. Mild reductions in miR-29a were observed, with near normalization of miR-29a expression to nondiabetic control levels (no treatment). This suggests that the promotion of the collagen I protein content, via the suppression of miR-29a, could also be contributing to UCUF-728-mediated wound repair.
This initial appraisal of the small molecule CXCR4 agonist, UCUF-728, demonstrates the utility of our approach for the development of therapies for impaired wound healing. Future studies will aim to optimize UCUF-728 lead for the suppression of miR-15b and miR-29a dysregulation, which may promote angiogenesis and collagen production, leading to more effective repairs of pathologic wound healing and reducing the probability of reinjury.
4. Materials and Methods
4.1. Reagents and Drug Treatment Protocol
All compounds were maintained as 10 mM DMSO stocks. To determine effects of compounds screened in cell-based assays, selected compounds were added to respective plates using the TECAN D300e digital dispenser. DMSO concentration was constant across all assay wells and did not exceed 0.5% in any cell-based assay. Unless otherwise indicated, reagents were purchased from Thermo Fisher Scientific (Waltham, MA, USA).
4.2. Cell Culture
CEM-CCRF cells (ATCC CCL-119) were cultured in growth media consisting of RPMI 1640 (Corning; Corning NY, USA) supplemented with 10% Fetal Bovine Serum (Corning; Corning, NY, USA), and 100 IU penicillin, 100 mg/mL streptomycin sulfate (Corning; Corning, NY, USA). Cells were maintained in suspension and subcultured twice weekly to maintain cell densities between 0.8 × 10
6 cells/mL and 2.5 × 10
6 cells/mL to maintain exponential growth phase. Cells were harvested for use in migration and calcium flux assays at 2.0 × 10
6 cells/mL to 2.5 × 10
6 cells/mL and resuspended in assay media to desired density for respective experiments. Tango™ CXCR4-bla U2OS Cells were maintained as a monolayer culture in McCoy’s 5a (Corning; Corning, NY, USA) supplemented with 10% dialyzed Fetal Bovine Serum, 0.1 mM MEM Non-essential amino acids solution, 25 mM HEPES solution (Corning; Corning, NY, USA), 1 mM Sodium Pyruvate, 200 ug/mL Zeocin selection reagent, 50 ug/mL Hygromycin B, and 100 ug/mL Geneticin. U2OS cells were routinely subcultured twice weekly by 0.25% Trypsin-EDTA treatment (Corning; Corning, NY, USA) and passaged to continue exponential growth phase for cell signaling assays. All cell cultures were maintained in cell culture incubators at 37 °C with 5% CO
2. Human Dermal Fibroblasts were isolated from human skin biopsies according to the method [
21]. Human dermal fibroblasts were cultured in full medium comprising Dulbecco’s modified eagle high-glucose (DMEM, Sigma-Aldrich, St. Louis, MO, USA) supplemented with 10% fetal bovine serum (FBS) and maintained at 37 °C in a humidified atmosphere containing 5% CO
2. For further experiments, cells were seeded cultured for 12 h. Thereafter, cells were starved for 16 h and were stimulated with UCUF-728 at different dose.
4.3. B-Arrestin Recruitment
Compounds, SDF-1α (PeproTech; 30028A), and DMSO (Sigma Aldrich; St. Louis, MO, USA) normalization to 0.5% were dispensed using the TECAN D300e digital dispenser into a 384-well black wall clear-bottom microplate (Corning; Corning, NY, USA). Tango™ CXCR4-bla U2OS Cells were resuspended in assay media consisting of DMEM (Corning; Corning, NY, USA) supplemented with 1% dialyzed Fetal bovine serum, 0.1 mM MEM Non-essential amino acids solution, 25 mM HEPES solution (Corning; Corning, NY, USA), 1 mM Sodium Pyruvate and 100 IU penicillin, 100 mg/mL streptomycin sulfate (Corning; Corning, NY, USA) at a density of 3.0 × 105 cells/mL. Cells suspension was dispensed at 30 μL per well into assay plate and incubated overnight in a cell culture incubator at 37 °C with 5% CO2. Following overnight incubation, LiveBlazer FRET B/G Loading Kit working reagent was prepared according to supplier guidelines and 6 μL of solution was dispensed into each well. Assay plate was covered and incubated in a dark place for two hours and Fluorescence Intensity was measured using the BMG LABTECH CLARIOstar Plus (BMG LABTECH Inc., Cary, NC, USA) according to the LiveBlazer FRET B/G Loading Kit excitation and emission measurement guidelines.
4.4. cAMP Signaling Assay
AMD3100, UCUF-728 and SDF-1a (PeproTech; Cranbury, NJ, USA) were dispensed in concentration-response mode using the TECAN D300e digital dispenser into white 384-OptiPlates (Perkin Elmer, Waltham, MA, USA). Forskolin at final concentration of 0.5 mM was added to all wells using the TECAN dispenser. Ten milliliters of PathHunter CHO-K1 CXCR4 cells (DiscoverX, Fremont, CA, USA) were added to compounds at 3000 cells/well, centrifuged at 1000 rpm for 30 s and incubated for 30 min at room temperature. DMSO (Sigma Aldrich, St. Louis, MO, USA) was normalized to 0.5%. Five nanoliters of 4X Eu-cAMP tracer working solution were added to all wells, followed by addition of 5 μL of 4X Ulight-anti-cAMP working solution to all wells. Plates were centrifuged at 1000 rpm for 1 min and incubated for 1 h at room temperature, then read using CLARIOstar Plus (BMG LABTECH Inc., Cary, NC, USA) Lance CAMP protocol.
4.5. Migration Assay
CEM-CCRF (ATCC CCL-119) were suspended in RPMI 1640 (Corning; 10-040-CV) at 5.0 × 105 cells/mL and allowed to incubate in cell culture incubator at 37 °C with 5% CO2 for one hour. For agonist activity measurement, compounds, SDF-1α control ligand (PeproTech; 300-28A), and DMSO (Sigma Aldrich, St. Louis, MO, USA) normalized to 0.5% were dispensed into the compound receiver tray of Multi-Screen 96-well assay plate (Millipore Sigma, St. Louis, MO, USA) at desired concentration using the TECAN D300e digital dispenser. Following 1 h cell incubation and compound dispensing, 150 μL of RPMI 1640 supplemented with 2% Fetal Bovine Serum was added to each well of the compound receiver tray, and 50 μL of 5 × 105 cells/mL cell suspension in RPMI were added to each well of the top filter plate. Plates were reassembled and placed in a cell culture incubator at 37 °C with 5% CO2 for three hours. After incubation, Multi-Screen plate was disassembled and 100 μL of solution was collected from compound receiver tray and transferred to 96 well luminescent plate and an equal volume of ATPLite 1step luminescence reagent (Perkin Elmer, Waltham, MA, USA) was added to each well. Luminescence was measured using the CLARIOstar Plus (BMG LABTECH Inc., Cary, NC, USA).
4.6. Binding Activity
Binding activity was measured using the Cisbio Tag-Lite Chemokine CXCR4 system (CisBio, Bedford, MA, USA). This assay was conducted using ready-to-assay Tag-Lite Chemokine CXCR4 labeled cells (C1TT1CXCR4), red fluorescent labeled CXCR4 ligand, and 1X Tag-Lite buffer prepared according to supplier guidelines. Compounds and AMD3100 were dispensed into a 384-well microplate (Greiner Bio, Frickenhausen Germany) using the TECAN D300e digital dispenser. CXCR4 Tag-lite cells were thawed and washed in 5 mL 1X Tag-Lite buffer, then re-suspended in 2.7 mL of 1X Tag-Lite buffer. After resuspension, 10 μL of cell suspension was dispensed into each assay well, followed by 5 μL of Tag-lite buffer and 5 μL of fluorescent ligand and allowed to incubate at room temperature in a dark place for three hours. Saturation binding and competition binding experiments were conducted on the same plate, with estimated Kd value of 12.5 nM fluorescent ligand for competition binding experiments. Following three-hour incubation, HTRF measurements were taken using the BMG LABTECH CLARIOstar Plus (BMG LABTECH Inc., Cary, NC, USA) with emission signals set to the Cisbio Tag-Lite Binding Assay supplier guidelines. HTRF ratio calculations completed according to supplier guidelines.
4.7. Animal Studies
All animal experiments were approved by the Institutional Animal Care and Use Committee at the University of Colorado Denver—Anschutz Medical Campus, and experimental protocols followed the guidelines described in the NIH Guide for the Care and Use of Laboratory Animals. In these experiments, we used 10-week-old, female, genetically diabetic C57BKS.Cg-m/Leprdb/J (Db) mice and heterozygous, non-diabetic (non-Db), age-matched female controls from the Jackson Laboratory (Bar Harbor, ME, USA). Mice were anesthetized with inhaled isoflurane. Each mouse was shaved and depilated before wounding. The dorsal skin was swabbed with alcohol and Betadine (Purdue Pharma, Stamford, CT, USA). Each mouse received a single, full-thickness dorsal wound (including panniculus carnosum) with an 8-mm punch biopsy (Miltex Inc., York, PA, USA). After wounding, a Hamilton syringe was used to deliver 50 μL of either 10 uM CAG1 or PBS, as a control. Ten nanoliters were injected intradermally at 12, 3, 6 and 9 o’clock and at the wound base. All wounds were dressed with Tegaderm (3M, St. Paul, MN, USA), which was subsequently removed on postoperative day 2. Postoperatively, the mice received a subcutaneous injection of an analgesic, Banamine (Schering-Plough Animal Health Corp., Union, NJ, USA). A full-thickness skin sample, centered on the wound, was harvested 3 and 7 days after surgery (n = 5 per timepoint).
4.8. Real Time Quantitative PCR
Total RNA was extracted with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s established protocol. RNA was converted into cDNA using the SuperScript First-Strand Synthesis System (Invitrogen, Life Technologies). Primers and probes for mouse miR-15b and miR-29a were acquired from Applied Biosystems TaqMan gene expression assay (Applied Biosystems, Foster City, CA, USA). Quantitative PCR was performed on a BIO-RAD CFX96 according to the manufacturer’s instructions. Quantitative values of genes of interest are normalized based on U6. Samples (n = 5 per group) were amplified in triplicate and results were averaged for each individual sample. The ΔΔCT method was used to calculate relative gene expression. Results are reported as mean ± SD.
4.9. Statistical Analysis
Results are expressed as mean ± SD for 3 to 5 independent experiments. Statistically significant differences in gene expression between two groups were assessed by Student’s
t-test, ANOVA with an appropriate post hoc test was to be used for multiple comparisons.
p < 0.05 was considered to be statistically significant. All concentration response curves were analyzed to determine EC
50 and E
max using the following equation:


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
