
The Proteolysis of ECM in Intervertebral Disc Degeneration


Department of Orthopaedics, Union Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430022, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2022, 23(3), 1715; https://doi.org/10.3390/ijms23031715
Submission received: 13 December 2021
/
Revised: 28 January 2022
/
Accepted: 29 January 2022
/
Published: 2 February 2022
(This article belongs to the Special Issue Proteolysis of Extracellular Matrix in Human Disease 2.0)
Abstract
:Intervertebral disc (IVD) degeneration (IDD) is a pathological process that commonly occurs throughout the human life span and is a major cause of lower back pain. Better elucidation of the molecular mechanisms involved in disc degeneration could provide a theoretical basis for the development of lumbar disc intervention strategies. In recent years, extracellular matrix (ECM) homeostasis has received much attention due to its relevance to the mechanical properties of IVDs. ECM proteolysis mediated by a variety of proteases is involved in the pathological process of disc degeneration. Here, we discuss in detail the relationship between the IVD as well as the ECM and the role of ECM proteolysis in the degenerative process of the IVD. Targeting ECM proteolysis-associated proteases may be an effective means of intervention in IDD.
1. Introduction
Lower back pain is a chronic and widespread musculoskeletal disorder that occurs in approximately more than 85% of people worldwide during their lifetime [1,2]. Disc degeneration has been implicated as a major cause of lower back pain [3,4]. Current clinical treatments for IDD include conservative treatment and surgical intervention. However, these treatment strategies all tend to relief symptoms rather than eliminate the underlying cause [1]. Further exploration of the molecular mechanisms involved in disc degeneration will provide new therapeutic targets to intervene in lower back pain.
The IVD is a complex structure consisting of three parts: the nucleus pulposus (NP) at its core, the annulus fibrosus (AF) surrounding it, and the endplates anchored to the upper and lower sides [5]. Many studies have shown that a variety of genetic or environmental etiologies that damage the nucleus pulposus, annulus fibrosus, or endplate can lead to disc degeneration, with the nucleus pulposus bearing the brunt of the degeneration [6,7,8]. Currently, the nucleus pulposus cells are considered to bear the highest degree of remolding during IDD and maintain the homeostasis of the ECM. In normal IVDs, the anabolism and catabolism of the ECM are in a dynamic balance. When the homeostatic balance of the ECM is disturbed by various stimuli, disc degeneration usually occurs [9]. As disc degeneration progresses, proteoglycans and collagen type II content decrease significantly, while collagen type I content increases significantly, resulting in decreased water absorption, a dysfunctional mechanical microenvironment, and decreased resistance to the loading of the ECM [10,11,12]. Mounting evidence indicate that various proteases play key roles in ECM proteolysis in disc degeneration. Abnormal expression of diverse protease in degenerated disc tissue and its role in ECM proteolysis have been reported. In this review, we systematically describe the role of diverse proteases in IDD. In addition, recent advances in the treatment of disc degeneration by maintaining ECM homeostasis are detailed in this article. A better understanding of the precise mechanisms by which proteases are associated with disc degeneration may provide guidance for the development of therapeutic strategies for disc degeneration.
2. ECM in IVD
The ECM is a non-cellular structure composed of about three hundred proteins [13]. ECM is present in the extracellular environment of all tissues and is involved in numerous cellular processes [14]. The major ECM components of the IVD include collagens, proteoglycan, and non-collagenous proteins [15]. Fine-tuned ECM dynamics are essential for the normal physiological functioning of the IVD [16]. NP cells are dispersed in a complex network of interwoven polysaccharides and collagen, which continuously interact with the surrounding ECM in a bidirectional manner to maintain tissue homeostasis [15].
2.1. Collagens
Collagen is a key component of the ECM and provides mechanical support for the disc cells. By acting on cell membrane receptors, collagen regulates a variety of cellular phenotypes including migration, proliferation, and differentiation [17]. Collagens I and II constitute the bulk of fibrillar matrix of the IVD, with other collagen subtypes including III, V, VI, IX, XI, XII, and XIV [18,19]. As the IVD degenerates, there is a shift from collagen type II to collagen type I [10]. In addition, the distribution of collagens changes, with more collagen II accumulating on the outer annulus and type I collagen accumulating in the nucleus pulposus and inner annulus [20].
2.2. Proteoglycans
Proteoglycans are macromolecules composed of core proteins and glycosaminoglycan (GAG) side chains. Aggrecan, the most abundant proteoglycan in IVD, is responsible for much of the water-attracting properties due to its high fixed charge density [10]. As age and degeneration progress, aggrecan is cleaved into non-aggregating fractions, leading to reduced hydration of the IVD [15]. Decorin is a member of small leucine-rich proteoglycans (SLRPs), which are widely expressed in connective tissue [21]. Decorin binds to a variety of ECM proteins such as collagen type I, II, III, and VI, which contribute to ECM assembly [22,23]. In addition, decorin binds to various signaling factors to regulate cell proliferation and differentiation and ECM turnover [24,25]. Decorin has been shown to be highly expressed in degenerating discs and may be involved in tissue repair [21]. Versican is a large extracellular proteoglycan that plays important roles in cell division, adhesion, migration, differentiation, and ECM turnover [26,27]. Previous studies have shown that versican is more highly expressed in IVD tissue than in articular cartilage and has a higher abundance in the nucleus pulposus [28]. In addition, in vitro studies have shown that versican exerts a protective effect by promoting the proliferation and adhesion of NP cells [29].
2.3. Non-Collagenous Proteins
Elastin is cross-linked by the soluble precursor tropoelastin [30]. Mature Elastin is present in various soft tissue ECM and is the most hydrolytically resistant ECM macromolecule [31]. Elastin constitutes approximately 10% of the IVD matrix and its abundance is higher in the AF compared to the NP [32,33]. Elastin, together with collagen, is thought to play an important role in IVD biomechanics [34,35]. Fibronectin, a ubiquitous secretory glycoprotein, plays a key role in the adhesion of many cell types by interacting with cell surface specific receptors and ECM components [36]. In addition, Anderson et al. showed that all five Fibronectin splice variants are present in IVD tissues [37]. Laminin is a heterotrimeric complex consisting of three polypeptide chains (α, β, and γ) [38]. At least 15 Laminin isoforms are formed by the combination of three different peptide chains, and multiple Laminin isoforms and their receptors are expressed in IVD tissues [38]. In addition, integrin-mediated interaction with Laminin regulates the phenotype of NP cells [39].
3. ECM Dysregulation in IDD
Mounting evidence has shown that various genetic and environmental factors, such as reduction in nutrition, overloading biomechanical stress, diabetes mellitus, smoking and infection, can damage NP cells and, thus, cause the dysregulation of ECM homeostasis [6,7,8].
First, the process of disc degeneration is accompanied by the progressive loss of notochord-like NP cells and their transformation into chondrocyte-like NP cells, partly attributed to non-physiological loading, inflammation, metabolic dysregulation, and hypoxia [40,41,42,43]. Notochord-like NP cells are essential for ECM homeostasis, and the loss or phenotypic alteration of notochord-like NP cells can lead to ECM dysregulation. Chondrocyte-like NP cells exhibit a different expression profile of ECM components compared to notochord-like NP cells. Chondrocyte-like NP cells tend to express collagen type I, which is capable of forming stronger fibers, accompanied by a reduced amount of more absorbent proteoglycans and collagen type II [44,45]. In addition, the shift from anabolic to catabolic metabolism occurs in IDD. A variety of cytokines expressed by degenerating nucleus pulposus cells upregulate the expression of proteases involved in the remodeling of the ECM [46,47,48,49]. Altogether, there is a decrease in proteoglycans and collagen type II and an increase in collagen type I during the degeneration process.
Second, changes in the ECM in the process of IDD impact the behavior of NP cells. The degenerative process of the IVD is accompanied by an increase in fibrotic-like collagen type I and a decrease in water-attracting proteoglycans and collagen type II [10,11,12]. The altered ECM composition increases its stiffness and regulates the phenotype and function of NP cells [50]. Available evidence indicates that a stiff substrate induces phenotypic changes including proliferation, apoptosis, and senescence in nucleus pulposus cells and is closely associated with disc degeneration [51]. In addition, ECM remodeling alters the mechanical microenvironment of the IVD. Inappropriate forces imposed on the nucleus pulposus, such as compression, stretch, and shear, impair the functioning of the nucleus pulposus cells [52,53,54,55]. Available evidence suggests that the response of nucleus pulposus cells to sustained mechanical stimuli is closely related to disc degeneration [16,56]. Furthermore, alterations in ECM composition modulate NP cell behavior by directly affecting cell–ECM interactions without altering the mechanical environment [39,57]. Dysregulation of ECM homeostasis is closely related to the process of disc degeneration, and proteolysis of the ECM occurs at the beginning of this process. A better understanding of the role played by ECM proteolysis in the degenerative process of the IVD will contribute to the development of new therapeutics.
4. The Proteolysis of ECM in IVD Pathogenesis
4.1. Matrix Metalloproteinases (MMPs)
MMPs, a class of Zn+-dependent endopeptidases, were first described in 1962 and are mainly responsible for the degradation of the ECM. To date, a total of 23 human MMPs have been identified [58]. Under normal human physiological conditions, MMPs are in a low expression state. Injury remodeling processes and an inflammatory environment increase their expression [59]. MMPs are first secreted as zymogens and are activated in the extracellular space by the hydrolytic cleavage of inactive structural domains or oxidative modification of thiol groups [60]. Importantly, MMPs are involved in a wide range of biological processes and are closely associated with disease development [61,62,63].
MMP expression is low in normal IVD tissue but increased in degenerative IVD tissue [20]. Studies using human IVD specimens show that MMPs increase gradually with an increasing grade of IVD degeneration [64]. In addition, highly expressed MMPs have been described in multiple animal models of degenerative discs [65,66,67,68]. Collectively, this evidence indicates that the level of MMPs expression is positively correlated with the grade of IVD degeneration. It also has been demonstrated that highly expressed MMPs are involved in IDD pathogenesis. Under pressure overload conditions, NP cells exhibit abnormal MMPs expression and corresponding ECM degeneration. Omlor et al. demonstrated a progressive elevation of MMP-13 with sustained applied pressure, which resulted in enhanced catabolism of the ECM [69]. Using a rat tail static stress model, Yurube found an imbalance in the expression of MMPs and anti-catabolic proteins [70]. Similarly, Yan et al. showed that compression loading altered the expression of multiple MMPs [71]. In addition, the inflammatory microenvironment demonstrated a significant effect on the expression of MMPs. Zhang et al. found disc degeneration accompanied by elevated expression of inflammatory cytokines and catabolic enzymes in a goat disc degeneration model [72]. It has been proven that inflammatory cytokines indirectly induce the hydrolytic activity of MMP-2 by elevating the expression of MT-MMP [73]. In addition, much evidence suggests that oxidative stress is also closely associated with over-activated MMPs. Reactive oxygen species (ROS) are involved in the transduction of multiple intracellular signaling pathways, but excessive ROS production can impair cellular function. In an earlier study, Nasto and colleagues showed that mitochondria-derived ROS degenerate in myeloid cells by upregulating the expression of MMP-1 and MMP-3 [74]. Dimozi showed that exogenous ROS addition can also induce the expression of ECM enzymes including MMP-1, MMP-3, and MMP-9 in hydrogen peroxide-treated NP cells [75]. There is increasing evidence that high glucose is closely associated with excessive ROS production, and Cheng et al. showed that oxidative stress impairs ECM metabolic homeostasis through the activation of p38/MAPK in high glucose-induced degenerating NP cells [76]. In a recent study, Liu et al. showed that oxidative stress resulted in decreased expression of the antioxidant protein FOXOs and upregulated expression of MMP-13 [77]. Furthermore, a range of therapies targeting MMPs has been explored to treat disc degeneration. Cheng et al. used hydrogel delivery of ferulic acid targeting MMP-3 to improve ECM homeostasis for the treatment of IDD [78]. Using a bioresponsive hydrogel loaded with miR-29a, Feng et al. proposed a strategy for targeting MMP-2 to treat disc degeneration [79]. Further exploration of the precise mechanisms of MMPs is needed to develop promising therapies for IDD.
4.2. A Disintegrin and Metalloprotease with Thrombospondin Motifs (ADAMTSs)
ADAMTS is a secreted protease of the metalloproteinase family that contains a thrombospondin structural domain that allows it to bind to ECM components, which can further lead to ECM proteolysis [80,81]. ADAMTS can be further classified into four categories based on their function and preferential ECM substrates. The hyalectanases (ADAMTS-1, ADAMTS-4, ADAMTS-5, ADAMTS-8, ADAMTS-9, ADAMTS-15, and ADAMTS-20) mainly target proteoglycans, while pro-collagen N-propeptidases (ADAMTS-2, ADAMTS-3, and ADAMTS-14) mainly target collagens I, II, and III. ADAMTS-13 is involved in the cleavage of the von Willebrand factor, which contributes to the pathophysiology of coagulation and thrombotic thrombocytopenic purpura. The function of the remaining ADAMTS (ADAMTS-6, ADAMTS-7, ADAMTS-10, ADAMTS-12, ADAMTS-16, ADAMTS-17, ADAMTS-18, and ADAMTS-19) is unclear.
Degenerative aggrecan is an important biochemical feature of IDD. The hyalectanases are responsible for aggrecan degradation in disc degeneration [82]. Pockert et al. demonstrated that ADAMTS-1, 4, 5, 9, and 15 accumulate in degenerating disc tissue and may contribute to the alteration of the ECM during disc degeneration [83]. Wang et al. found that inflammatory cytokines regulate ADAMTS-5 expression through SDC4, which is involved in the pathophysiology of disc degeneration [84]. Using a rat tail static compression model, high expression of ADAMTS-4 and ADAMTS-5 was observed [70]. In addition, the direct degradation effect of ADAMTS-4 on the ECM of the nucleus pulposus was assessed. It was shown that intradiscal injection with ADAMTS-4 impaired cell activity and collagen content [85]. It also has been demonstrated that sIL-13Rα2-Fc is able to rescue ECM degradation by targeting ADAMTS-8 [86]. Collectively, these studies indicated that the hyalectanases play key roles in the pathophysiology of disc degeneration. In addition to hyalectanases, other ADAMTSs have been reported to be associated with disc degenerative processes. For example, McCann et al. found that TNF-α and ADAMTS-7 were highly expressed in IL-21-treated NP cells, and further studies showed that inflammatory factor-induced expression of multiple catabolic enzymes was ADAMTS-7-mediated [87,88]. In addition, Yu et al. found elevated expression of ADAMTS-7 and ADAMTS-12 in degenerated disc tissue accompanied by degradation of the cartilage oligomeric matrix protein [89]. Further investigation of the role of ADAMTSs in IDD etiology is critical for IDD intervention.
4.3. Cathepsins Proteases
Cathepsins belong to the peptidases family, which are primarily involved in the endosomes and lysosomes for protein degradation [61]. There is increasing evidence indicating that cathepsins are also expressed in the extracellular space and are involved in the degradation of the ECM [90]. Cathepsins are composed of different protease families: cysteine, serine, and aspartyl proteases. Cathepsins play an important role in a variety of diseases, such as bone and joint degenerative diseases, and are receiving widespread attention [61].
Dando et al. described a rabbit IVD model in which intradisc injection of Cathepsins B resulted in proteoglycan breakdown and histological and imaging alterations of the IVD [62]. Further studies demonstrated that Cathepsins B was highly expressed in degenerating discs [63]. Ariga et al. found that cathepsins D, K, and L accumulate at the site of degenerated discs and that this specific localization may be associated with ECM disorders during disc degeneration [91]. In addition, Gruber et al. found that cathepsins K expression was significantly elevated in degenerated discs and positively correlated with receptor activator of the NF-κB ligand [92]. In a zebrafish model, the impact of high bone mineral density induced by mutant cathepsins K on IVDs was examined [93]. It was found that high bone mineral density was significantly associated with disc degeneration. Additionally, Konttinen et al. demonstrated a significant increase in cathepsins K-positive cells in degenerating IVDs [94]. These studies suggest that cathepsins play key roles in IDD.
4.4. Other Proteases
High temperature requirement A1 (HTRA1) belongs to the serine protease family and is capable of degrading a variety of ECM components. Tiaden et al. found that HTRA1 expression was elevated in degenerating discs, and further studies showed that HTRA1 could upregulate various catabolic enzymes [95]. Hepsin (HPN) is also known as transmembrane Serine Protease 1 (TMPRSS1), whose overexpression leads to degradation of the IVD ECM [96]. Plasmin is involved in the activation of several MMPs [97]. Salo et al. showed that plasmin was present in degenerating disc tissue with high expression of multiple catabolic enzymes, suggesting that plasmin may be associated with anabolic imbalance during disc degeneration. In addition, Campos et al. demonstrated that heparanase isoforms (HPSE1 and HPSE2) are highly expressed in degenerated disc tissues [98].
5. Therapeutic Targeting of Proteases for ID
Current clinical treatments for IDD are aimed at relieving symptoms rather than directly targeting disc degeneration. Given the critical role of ECM proteolysis driven by proteases in the process of disc degeneration, targeting proteases in disc degeneration is an alternative option. Here we review recent advances in modulating ECM homeostasis for the treatment of IDD (Table 1).
5.1. Anti-Inflammation
Inflammatory stimulation is one of the most important etiologies of disc degeneration, emphasizing its ability to upregulate multiple proteases. Anti-inflammatory therapy demonstrates the potential for treating disc degeneration. The NF-κB pathway has been reported to mediate the transcriptional activation of multiple hydrolases in response to inflammatory stimuli. In cytokine-induced inflammatory models, multiple bioactive compounds were shown to downregulate the expression of multiple proteases by inhibiting NF-κB, which in turn blocked the ECM destruction process [99,100,101]. In addition, Li et al. found that Crocin and Sesamin exerted anti-inflammatory effects by inhibiting the activation of the JNK pathway and slowed down the degradation of the degenerating IVD ECM [102,103]. It has also been reported that Platelet-Rich Plasma (PRP) to treat disc degeneration, and the results showed that PRP treatment promoted the production of major components of the ECM and inhibited the expression of MMP-1/3 [104,105]. These studies suggest that inhibition of inflammation represents a new class of treatment for disc degeneration (Table 1).
5.2. Anti-Oxidation
In recent years, there has been increasing evidence that oxidative stress is closely related to the imbalance of ECM homeostasis in degenerated discs. Using a rat model of IDD, Qin et al. found that Danshen effectively attenuated the disc degeneration process by targeting the oxidative reaction [106]. Wang et al. demonstrated that Genistein (GES) treatment rescued the expression levels of aggrecan and type II collagen in TBHP-treated NP cells. The role of GES in restoring ECM homeostasis was further demonstrated to be achieved through Nrf2-mediated anti-proteases [107]. 1,4-dihydropyridine (DHP), a specific sirt1 agonist, was shown to be a potential therapeutic agent for IDD due to its antioxidant effect. The administration of DHP significantly alleviated the degeneration of the ECM by reducing the expression of MMP-3 and ADAMTS-5 in degenerating NP cells [108]. Aspirin is widely used as an anti-inflammatory agent for the treatment of low back pain. Liu et al. found that aspirin exerts antioxidant and anti-inflammatory effects in an AMPK-dependent manner. Aspirin treatment protected NP cells from ECM degradation by inhibiting the expression of multiple hydrolases, including MMP-3/13 and ADAMTS-5. Furthermore, aspirin intervertebral injections significantly hindered the degenerative process of the IVD in a rat model [126]. In addition, melatonin and N-acetyl cysteine (NAC) were reported to maintain ECM homeostasis by regulating redox homeostasis [127,128]. These studies suggest that antioxidant drugs may be a promising therapeutic strategy for disc degeneration, emphasizing their modulation of ECM homeostasis (Table 1).
5.3. Stem Cells Therapy
Recently, many studies have focused on stem cell therapy for disc degeneration by the injection or transplantation of stem cells, which aim to reconstitute the ECM of the IVD [129]. Previous studies have reported that the co-incubation of nucleus pulposus cells with stem cells can induce stem cell differentiation, leading to regenerative repair of degenerated discs [110]. Further, Yang and his colleagues used bone marrow derived mesenchymal stem cells (BMSCs) injections to treat disc degeneration. They showed that BMSCs transplantation alleviated disc degeneration by stimulating endogenous ECM regeneration in addition to autonomous differentiation [109]. Yi et al. used human tissue inhibitor of metalloproteinase 1 (TIMP-1) overexpression modified BMSCs to treat degenerated discs by directly targeting the ECM anabolic catabolic homeostasis [111]. In addition, Wang et al. found that BMSCs promoted ECM regeneration in degenerated NP cells via the miR-101-3p/EIF4G2 axis [130]. However, stem cell injections or transplants alone often fail to achieve the desired effect due to their lack of protection and support by the ECM. In a recent study, Feng et al. used an injectable microsphere scaffold for the delivery of stem cells, which provided a suitable environment for stem cell implantation, proliferation, and differentiation, and significantly improved the efficacy of stem cell therapy for degenerated discs [131]. In a similar study, Ukeba et al. used ultra-purified alginate gels loaded with BMSCs to treat degenerated discs, which significantly promoted ECM synthesis with significantly higher efficacy than BMSCs injection alone [132]. In addition, Frapin et al. treated disc degeneration by modulating the distribution of endogenous disc stem/progenitor cells. Chemokine CCL-5 was delivered to the nucleus pulposus via pullulan microbeads (PMBs) to recruit disc stem/progenitor cells, which reversed the phenotype of degenerating NP cells [112]. Collectively, substantial evidence suggests that stem cell injection or transplantation is an effective treatment for targeting the degenerating ECM, but further studies are needed to assess their safety and efficacy (Table 1).
5.4. Metabolic Modulation
Metabolic diseases such as the hyperglycemic microenvironment produced by diabetes mellitus and estrogen deficiency-induced bone loss have been shown to be closely associated with disc degeneration [77,133,134]. Marein, a plant active ingredient with antidiabetic effects, was shown to alleviate degeneration of the ECM of the degenerating NP cells due to high glucose exposure [113]. In addition, anti-inflammatory combined with anti-advanced-glycation-end-products (AGEs) treatment significantly improved disc degeneration in a mouse model of diabetes mellitus [114]. Using an ovariectomized rat model, Song et al. found that alendronate (ALN) treatment significantly retarded the progression of disc degeneration. ALN not only reduced the expression of multiple hydrolases and type I collagen, but also promoted the expression of aggrecan and type II collagen, and the underlying mechanism may be related to the maintenance of disc structural integrity [115,116,117]. In conclusion, these data suggest that the imbalance in metabolic homeostasis may be a potential target for the degeneration of the IVD ECM (Table 1).
5.5. Biomaterials
In recent years, novel bioactive materials based on bioengineering have been widely used to treat disc degeneration. Cheng et al. constructed an injectable thermosensitive chitosan–gelatin–glycerol phosphate (C/G/GP) hydrogel for the treatment of disc degeneration by the controlled release of ferredoxin acid. This study demonstrated that C/G/GP gels are well adapted to ferulic acid and achieve controlled release through their thermosensitive properties to target oxidative stress-induced ECM degeneration [78,118]. Using a bioresponsive hydrogel loaded with miR-29a, Feng et al. proposed a strategy for targeting MMP-2 to treat disc degeneration. In this study, the investigators exploited the hyperactivation of proteases in degenerating discs to prepare a hydrolase-responsive miRNA delivery system to regulate ECM homeostasis [79]. Moreover, Larrañaga et al. prepared tannic acid (TA)-functionalized polymer capsules to target MMP-3 and ADAMTS-5 for the treatment of IVDs. The antioxidant functionalized polymer capsules prepared in this study represent the application of polymer capsules for the treatment of oxidative stress-induced ECM degeneration [119]. More recently, a controllable release hydrogel was prepared for the delivery of aspirin. This study demonstrated that controlled release gels carrying aspirin significantly alleviated the ECM degeneration of the IVDs and had superior mechanical properties [120]. Additional efforts to treat degenerating discs include injectable microspheres loaded with tumor necrosis factor (TNF) receptor type II. This antagonist microsphere significantly slows disc degeneration by targeting disorders of ECM metabolism [121]. Similarly, a GelMA microsphere coupled with the active peptide APETx2 was shown to modulate local inflammation overactivation in the degenerated IVD. This study suggests that modulation of the inflammatory microenvironment is a potential means of targeting ECM degradation [122]. In recent studies, caveolae associated protein 2 (Cavin-2) modified engineered extracellular vesicles were used to treat disc degeneration. Cavin-2 modification significantly increased the extracellular vesicle uptake efficacy in degenerating NP cells compared to stem cell-derived extracellular vesicles alone [135]. Collectively, these studies demonstrate the great potential of bioactive materials in targeting IVD ECM degeneration, but more studies are needed to evaluate their long-term safety and efficacy (Table 1).
5.6. Gene Therapy
Recently, gene therapy has received increasing attention. There is evidence that gene therapy has the potential to treat IDD [136,137]. Given the important role of multiple proteases in the imbalance of ECM metabolism, targeting proteases through gene therapy may provide a new strategy for the treatment of IDD. Using a rabbit model of IDD, Leckie et al. showed that AAV2-TIMP1 effectively alleviated disc degeneration by increasing collagen type II levels [123]. Similarly, another study showed that TIMP1 co-transduction with transforming growth factor (TGF-β3) and connective tissue growth factor (CTGF) promoted the synthesis of ECM in degenerated discs [124]. In addition, ADAMTS-5 siRNA was shown to retard disc degeneration by directly targeting the protease ADAMTS-5 [125]. Thus, transduction of endogenous proteases inhibitors and direct targeting of proteases have important potential applications in the treatment of IDD (Table 1).
Despite the great progress made in exploring IDD therapeutic strategies, many challenges remain in its translational future. The avascularity of the nucleus pulposus results in a lack of nutrient supply to the core region of the disc. The poor microenvironment of the IVD is a great obstacle to the treatment of IDD [133,134]. Proteases activation is thought to be a driver of ECM degradation in the disc; however, IDD treatment strategies targeting proteases have also failed to meet expectations due to transient effects [138]. Overall, further studies in developing therapeutic intervention for IDD should take these factors into account.
6. Conclusions
The imbalance of ECM homeostasis is closely related to the pathological process of disc degeneration. ECM proteolysis, mediated by multiple hydrolases in the extracellular space, plays an initiating role in this process (Figure 1). Here, we describe in detail the role of the major proteases in disc degeneration and list strategies for targeting the ECM to treat degenerative discs.
Given the paucity of therapeutic options for disc degeneration and the critical role of proteases in this regard, targeting one or more proteases demonstrates potential for treating disc degeneration. However, the regulation and precise mechanisms of key proteases in disc degeneration are still largely unknown. Further research is needed that focuses on the molecular mechanisms of proteases regulation and to develop effective treatments for disc degeneration that target proteases.
Author Contributions
H.L., Y.S., R.L., G.L., W.Z. and C.Y. contributed to the manuscript’s design and writing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant 638 Nos. 81902261 and 81772401), the Application Foundation and Advanced Program of 639 Wuhan Science and Technology Bureau (2019020701011457), the Fundamental 640 Research Funds for the Central Universities (2019kfyXMBZ063), and the National 641 Key Research and Development Program of China (2018YFB1105700).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest in relation to this review.
References
- Deyo, R.A.; Mirza, S.K. CLINICAL PRACTICE. Herniated Lumbar Intervertebral Disk. N. Engl. J. Med. 2016, 374, 1763–1772. [Google Scholar] [CrossRef]
- Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [CrossRef] [Green Version]
- Vlaeyen, J.W.S.; Maher, C.G.; Wiech, K.; Van Zundert, J.; Meloto, C.B.; Diatchenko, L.; Battié, M.C.; Goossens, M.; Koes, B.; Linton, S.J. Low back pain. Nat. Rev. Dis. Primers 2018, 4, 52. [Google Scholar] [CrossRef]
- Risbud, M.V.; Shapiro, I.M. Role of cytokines in intervertebral disc degeneration: Pain and disc content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef]
- Roughley, P.J. Biology of intervertebral disc aging and degeneration: Involvement of the extracellular matrix. Spine 2004, 29, 2691–2699. [Google Scholar] [CrossRef]
- Song, Y.; Li, S.; Geng, W.; Luo, R.; Liu, W.; Tu, J.; Wang, K.; Kang, L.; Yin, H.; Wu, X.; et al. Sirtuin 3-dependent mitochondrial redox homeostasis protects against AGEs-induced intervertebral disc degeneration. Redox Biol. 2018, 19, 339–353. [Google Scholar] [CrossRef]
- Kang, L.; Xiang, Q.; Zhan, S.; Song, Y.; Wang, K.; Zhao, K.; Li, S.; Shao, Z.; Yang, C.; Zhang, Y. Restoration of Autophagic Flux Rescues Oxidative Damage and Mitochondrial Dysfunction to Protect against Intervertebral Disc Degeneration. Oxidative Med. Cell. Longev. 2019, 2019, 7810320. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Wang, Y.; Zhang, Y.; Geng, W.; Liu, W.; Gao, Y.; Li, S.; Wang, K.; Wu, X.; Kang, L.; et al. Advanced glycation end products regulate anabolic and catabolic activities via NLRP3-inflammasome activation in human nucleus pulposus cells. J. Cell. Mol. Med. 2017, 21, 1373–1387. [Google Scholar] [CrossRef]
- Le Maitre, C.L.; Pockert, A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35, 652–655. [Google Scholar] [CrossRef] [Green Version]
- Antoniou, J.; Steffen, T.; Nelson, F.; Winterbottom, N.; Hollander, A.P.; Poole, R.A.; Aebi, M.; Alini, M. The human lumbar intervertebral disc: Evidence for changes in the biosynthesis and denaturation of the extracellular matrix with growth, maturation, ageing, and degeneration. J. Clin. Investig. 1996, 98, 996–1003. [Google Scholar] [CrossRef]
- Weiler, C.; Nerlich, A.G.; Zipperer, J.; Bachmeier, B.E.; Boos, N. 2002 SSE Award Competition in Basic Science: Expression of major matrix metalloproteinases is associated with intervertebral disc degradation and resorption. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2002, 11, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, C.; Schietzsch, M.; Kirchner, T.; Nerlich, A.G.; Boos, N.; Wuertz, K. Age-related changes in human cervical, thoracal and lumbar intervertebral disc exhibit a strong intra-individual correlation. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2012, 21 (Suppl. S6), S810–S818. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O.; Naba, A. Overview of the matrisome--an inventory of extracellular matrix constituents and functions. Cold Spring Harb. Perspect. Biol. 2012, 4, a004903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamanos, N.K.; Piperigkou, Z.; Passi, A.; Götte, M.; Rousselle, P.; Vlodavsky, I. Extracellular matrix-based cancer targeting. Trends Mol. Med. 2021, 27, 1000–1013. [Google Scholar] [CrossRef] [PubMed]
- Roughley, P.J.; Melching, L.I.; Heathfield, T.F.; Pearce, R.H.; Mort, J.S. The structure and degradation of aggrecan in human intervertebral disc. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2006, 15 (Suppl. S3), S326–S332. [Google Scholar] [CrossRef] [Green Version]
- Vergroesen, P.P.; Kingma, I.; Emanuel, K.S.; Hoogendoorn, R.J.; Welting, T.J.; van Royen, B.J.; van Dieën, J.H.; Smit, T.H. Mechanics and biology in intervertebral disc degeneration: A vicious circle. Osteoarthr. Cartil. 2015, 23, 1057–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.L.; Walker, R.A. Integrins: A role as cell signalling molecules. Mol. Pathol. 1999, 52, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Eyre, D.R.; Matsui, Y.; Wu, J.J. Collagen polymorphisms of the intervertebral disc. Biochem. Soc. Trans. 2002, 30, 844–848. [Google Scholar] [CrossRef] [Green Version]
- Sivan, S.S.; Hayes, A.J.; Wachtel, E.; Caterson, B.; Merkher, Y.; Maroudas, A.; Brown, S.; Roberts, S. Biochemical composition and turnover of the extracellular matrix of the normal and degenerate intervertebral disc. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2014, 23 (Suppl. S3), S344–S353. [Google Scholar] [CrossRef]
- Wang, W.J.; Yu, X.H.; Wang, C.; Yang, W.; He, W.S.; Zhang, S.J.; Yan, Y.G.; Zhang, J. MMPs and ADAMTSs in intervertebral disc degeneration. Clin. Chim. Acta Int. J. Clin. Chem. 2015, 448, 238–246. [Google Scholar] [CrossRef]
- Zhang, T.W.; Dong, J.; Jiang, L.B.; Li, Z.F.; Jiang, L.B. Decorin Inhibits Nucleus Pulposus Apoptosis by Matrix-Induced Autophagy via mTOR Pathway. J. Orthop. Res. 2020. [Google Scholar] [CrossRef] [PubMed]
- Kinsella, M.G. Retrovirally Mediated Expression of Decorin by Macrovascular Endothelial Cells Effects on Cellular Migration and Fibronectin Fibrillogenesisin Vitro. J. Biol. Chem. 2000, 275, 13924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neill, T.; Schaefer, L.; Iozza, R.V. Decorin: A Guardian from the Matrix - ScienceDirect. Am. J. Pathol. 2012, 181, 380–387. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.R.; Rudisill, J.A.; Gallo, R.L. Structural and sequence motifs in dermatan sulfate for promoting FGF-2 and FGF-7 activity. J. Biol. Chem. 2004, 280, 5300–5306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidler, D.G.; Dreier, R. Decorin and its galactosaminoglycan chain: Extracellular regulator of cellular function? Iubmb Life 2010, 60, 729–733. [Google Scholar] [CrossRef]
- Andersson-Sjoland, A.; Hallgren, O.; Rolandsson, S.; Weitoft, M.; Tykesson, E.; Larsson-Callerfelt, A.K.; Rydell-Tormanen, K.; Bjermer, L.; Malmstrom, A.; Karlsson, J.C. Versican in inflammation and tissue remodeling: The impact on lung disorders. Glycobiology 2014. [Google Scholar] [CrossRef] [Green Version]
- Wight, T.N. Versican: A versatile extracellular matrix proteoglycan in cell biology. Curr. Opin. Cell Biol. 2002, 14, 617–623. [Google Scholar] [CrossRef]
- Sztrolovics, R.; Grover, J.; Cs-Szabo, G.; Shi, S.L.; Roughley, P.J. The characterization of versican and its message in human articular cartilage and intervertebral disc. J. Orthop. Res. 2002, 20, 257–266. [Google Scholar] [CrossRef]
- Bing, L.Y.; Yang, B.B.; Erwin, M.; Ang, L.C.; Finkelstein, J.; Yee, A. Versican G3 domain enhances cellular adhesion and proliferation of bovine intervertebral disc cells cultured in vitro. Life Sci. 2003, 73, 3399–3413. [Google Scholar]
- Rosenbloom, J. Elastin: Relation of protein and gene structure to disease. Lab. Investig. 1985, 51, 605–623. [Google Scholar]
- Debelle, L.; Tamburro, A.M. Elastin: Molecular description and function. Int. J. Biochem. Cell Biol. 1999, 31, 261–272. [Google Scholar] [CrossRef]
- Olczyk, K. Age-related change of elastin content in human intervertebral discs. Folia Histochem. Cytobiol 1994, 32. [Google Scholar]
- Johnson, E.F.; Caldwell, R.W.; Berryman, H.E.; Miller, A.; Chetty, K. Elastic fibers in the anulus fibrosus of the dog intervertebral disc. Acta Anat. 1984, 118, 238–242. [Google Scholar] [CrossRef]
- Smith, L.J.; Fazzalari, N.L. Regional variations in the density and arrangement of elastic fibres in the anulus fibrosus of the human lumbar disc. J. Anat. 2010, 209, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Fairbank, J.; Roberts, S.; Urban, J. The Elastic Fiber Network of the Anulus Fibrosus of the Normal and Scoliotic Human Intervertebral Disc. Spine 2005, 30, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.G.; Markova, D.; Adams, S.L.; Pacifici, M.; An, H.S.; Zhang, Y. Fibronectin splicing variants in human intervertebral disc and association with disc degeneration. Spine 2010, 35, 1581. [Google Scholar] [CrossRef]
- Chen, J.; Jing, L.; Gilchrist, C.L.; Richardson, W.J.; Fitch, R.D.; Setton, L.A. Expression of Laminin Isoforms, Receptors and Binding Proteins Unique to Nucleus Pulposus Cells of Immature Intervertebral Disc. Connect. Tissue Res. 2009, 50, 294–306. [Google Scholar] [CrossRef]
- Speer, J.; Barcellona, M.; Jing, L.; Liu, B.; Lu, M.; Kelly, M.; Buchowski, J.; Zebala, L.; Luhmann, S.; Gupta, M.; et al. Integrin-mediated interactions with a laminin-presenting substrate modulate biosynthesis and phenotypic expression for cells of the human nucleus pulposus. Eur. Cells Mater. 2021, 41, 793–810. [Google Scholar] [CrossRef]
- Zhao, C.Q.; Wang, L.M.; Jiang, L.S.; Dai, L.Y. The cell biology of intervertebral disc aging and degeneration. Ageing Res. Rev. 2007, 6, 247–261. [Google Scholar] [CrossRef]
- Guehring, T.; Nerlich, A.; Kroeber, M.; Richter, W.; Omlor, G.W. Sensitivity of notochordal disc cells to mechanical loading: An experimental animal study. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2010, 19, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guehring, T.; Wilde, G.; Sumner, M.; Grünhagen, T.; Karney, G.B.; Tirlapur, U.K.; Urban, J.P. Notochordal intervertebral disc cells: Sensitivity to nutrient deprivation. Arthritis Rheum. 2009, 60, 1026–1034. [Google Scholar] [CrossRef] [PubMed]
- Yurube, T.; Hirata, H.; Kakutani, K.; Maeno, K.; Takada, T.; Zhang, Z.; Takayama, K.; Matsushita, T.; Kuroda, R.; Kurosaka, M.; et al. Notochordal cell disappearance and modes of apoptotic cell death in a rat tail static compression-induced disc degeneration model. Arthritis Res. Ther. 2014, 16, R31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, J.P.G.; Roberts, S.; Ralphs, J.R. The Nucleus of the Intervertebral Disc from Development to Degeneration1. Am. Zool. 2015, 40, 53–061. [Google Scholar] [CrossRef]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.; et al. Exhaustion of nucleus pulposus progenitor cells with ageing and degeneration of the intervertebral disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, S.M.; Doyle, P.; Minogue, B.M.; Gnanalingham, K.; Hoyland, J.A. Increased expression of matrix metalloproteinase-10, nerve growth factor and substance P in the painful degenerate intervertebral disc. Arthritis Res. Ther. 2009, 11, R126. [Google Scholar] [CrossRef] [Green Version]
- Purmessur, D.; Walter, B.A.; Roughley, P.J.; Laudier, D.M.; Hecht, A.C.; Iatridis, J. A role for TNFα in intervertebral disc degeneration: A non-recoverable catabolic shift. Biochem. Biophys. Res. Commun. 2013, 433, 151–156. [Google Scholar] [CrossRef] [Green Version]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration. Arthritis Res. Ther. 2005, 7, R732–R745. [Google Scholar] [CrossRef] [Green Version]
- Binch, A.L.; Cole, A.A.; Breakwell, L.M.; Michael, A.L.; Chiverton, N.; Cross, A.K.; Le Maitre, C.L. Expression and regulation of neurotrophic and angiogenic factors during human intervertebral disc degeneration. Arthritis Res. Ther. 2014, 16, 416. [Google Scholar] [CrossRef] [Green Version]
- Iatridis, J.C.; Setton, L.A.; Weidenbaum, M.; Mow, V.C. Alterations in the mechanical behavior of the human lumbar nucleus pulposus with degeneration and aging. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 1997, 15, 318–322. [Google Scholar] [CrossRef]
- Fearing, B.V.; Jing, L.; Barcellona, M.N.; Witte, S.E.; Buchowski, J.M.; Zebala, L.P.; Kelly, M.P.; Luhmann, S.; Gupta, M.C.; Pathak, A.; et al. Mechanosensitive transcriptional coactivators MRTF-A and YAP/TAZ regulate nucleus pulposus cell phenotype through cell shape. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 14022–14035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gullbrand, S.E.; Peterson, J.; Mastropolo, R.; Roberts, T.T.; Lawrence, J.P.; Glennon, J.C.; DiRisio, D.J.; Ledet, E.H. Low rate loading-induced convection enhances net transport into the intervertebral disc in vivo. Spine J. Off. J. North Am. Spine Soc. 2015, 15, 1028–1033. [Google Scholar] [CrossRef] [PubMed]
- Neidlinger-Wilke, C.; Galbusera, F.; Pratsinis, H.; Mavrogonatou, E.; Mietsch, A.; Kletsas, D.; Wilke, H.J. Mechanical loading of the intervertebral disc: From the macroscopic to the cellular level. Eur. Spine J. Off. Publ. Eur. Spine Soc. Eur. Spinal Deform. Soc. Eur. Sect. Cerv. Spine Res. Soc. 2014, 23 (Suppl. S3), S333–S343. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.R.; Yuan, T.Y.; Huang, C.Y.; Brown, M.D.; Gu, W.Y. Nutrient transport in human annulus fibrosus is affected by compressive strain and anisotropy. Ann. Biomed. Eng. 2012, 40, 2551–2558. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, H.T.; Hoyland, J.A.; Millward-Sadler, S.J. The response of human anulus fibrosus cells to cyclic tensile strain is frequency-dependent and altered with disc degeneration. Arthritis Rheum. 2010, 62, 3385–3394. [Google Scholar] [CrossRef]
- Setton, L.A.; Chen, J. Mechanobiology of the intervertebral disc and relevance to disc degeneration. J. Bone Jt. Surg. Am. Vol. 2006, 88 (Suppl. S2), 52–57. [Google Scholar] [CrossRef]
- Bridgen, D.T.; Gilchrist, C.L.; Richardson, W.J.; Isaacs, R.E.; Brown, C.R.; Yang, K.L.; Chen, J.; Setton, L.A. Integrin-mediated interactions with extracellular matrix proteins for nucleus pulposus cells of the human intervertebral disc. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2013, 31, 1661–1667. [Google Scholar] [CrossRef] [Green Version]
- Vacek, T.P.; Rehman, S.; Neamtu, D.; Yu, S.; Givimani, S.; Tyagi, S.C. Matrix metalloproteinases in atherosclerosis: Role of nitric oxide, hydrogen sulfide, homocysteine, and polymorphisms. Vasc. Health Risk Manag. 2015, 11, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef]
- Lutgens, S.P.; Cleutjens, K.B.; Daemen, M.J.; Heeneman, S. Cathepsin cysteine proteases in cardiovascular disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2007, 21, 3029–3041. [Google Scholar] [CrossRef]
- Dando, P.M.; Morton, D.B.; Buttle, D.J.; Barrett, A.J. Quantitative assessment of human proteinases as agents for chemonucleolysis. Spine 1988, 13, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.E.; Hoelscher, G.L.; Ingram, J.A.; Bethea, S.; Hanley, E.N., Jr. Autophagy in the Degenerating Human Intervertebral Disc: In Vivo Molecular and Morphological Evidence, and Induction of Autophagy in Cultured Annulus Cells Exposed to Proinflammatory Cytokines-Implications for Disc Degeneration. Spine 2015, 40, 773–782. [Google Scholar] [CrossRef]
- Xu, H.; Mei, Q.; He, J.; Liu, G.; Zhao, J.; Xu, B. Correlation of matrix metalloproteinases-1 and tissue inhibitor of metalloproteinases-1 with patient age and grade of lumbar disk herniation. Cell Biochem. Biophys. 2014, 69, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Sobajima, S.; Shimer, A.L.; Chadderdon, R.C.; Kompel, J.F.; Kim, J.S.; Gilbertson, L.G.; Kang, J.D. Quantitative analysis of gene expression in a rabbit model of intervertebral disc degeneration by real-time polymerase chain reaction. Spine J. Off. J. North Am. Spine Society 2005, 5, 14–23. [Google Scholar] [CrossRef]
- Wei, F.; Zhong, R.; Zhou, Z.; Wang, L.; Pan, X.; Cui, S.; Zou, X.; Gao, M.; Sun, H.; Chen, W.; et al. In vivo experimental intervertebral disc degeneration induced by bleomycin in the rhesus monkey. BMC Musculoskelet. Disord. 2014, 15, 340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, K.L.; Jordan-Mahy, N.; Nicklin, M.J.; Le Maitre, C.L. Interleukin-1 receptor antagonist deficient mice provide insights into pathogenesis of human intervertebral disc degeneration. Ann. Rheum. Dis. 2013, 72, 1860–1867. [Google Scholar] [CrossRef]
- Iwata, M.; Aikawa, T.; Hakozaki, T.; Arai, K.; Ochi, H.; Haro, H.; Tagawa, M.; Asou, Y.; Hara, Y. Enhancement of Runx2 expression is potentially linked to β-catenin accumulation in canine intervertebral disc degeneration. J. Cell. Physiol. 2015, 230, 180–190. [Google Scholar] [CrossRef]
- Omlor, G.W.; Lorenz, H.; Engelleiter, K.; Richter, W.; Carstens, C.; Kroeber, M.W.; Guehring, T. Changes in gene expression and protein distribution at different stages of mechanically induced disc degeneration--An in vivo study on the New Zealand white rabbit. J. Orthop.Res. Off. Publ. Orthop. Res. Soc. 2006, 24, 385–392. [Google Scholar] [CrossRef]
- Yurube, T.; Takada, T.; Suzuki, T.; Kakutani, K.; Maeno, K.; Doita, M.; Kurosaka, M.; Nishida, K. Rat tail static compression model mimics extracellular matrix metabolic imbalances of matrix metalloproteinases, aggrecanases, and tissue inhibitors of metalloproteinases in intervertebral disc degeneration. Arthritis Res. Ther. 2012, 14, R51. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Pan, Y.; Wang, S.; Cheng, M.; Kong, H.; Sun, C.; Hu, K.; Chen, T.; Dong, Q.; Chen, J. Static Compression Induces ECM Remodeling and Integrin α2β1 Expression and Signaling in a Rat Tail Caudal Intervertebral Disc Degeneration Model. Spine 2017, 42, E448–E458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Gullbrand, S.E.; Schaer, T.P.; Lau, Y.K.; Jiang, Z.; Dodge, G.R.; Elliott, D.M.; Mauck, R.L.; Malhotra, N.R.; Smith, L.J. Inflammatory cytokine and catabolic enzyme expression in a goat model of intervertebral disc degeneration. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2020, 38, 2521–2531. [Google Scholar] [CrossRef] [PubMed]
- Séguin, C.A.; Pilliar, R.M.; Madri, J.A.; Kandel, R.A. TNF-alpha induces MMP2 gelatinase activity and MT1-MMP expression in an in vitro model of nucleus pulposus tissue degeneration. Spine 2008, 33, 356–365. [Google Scholar] [CrossRef] [PubMed]
- Nasto, L.A.; Robinson, A.R.; Ngo, K.; Clauson, C.L.; Dong, Q.; St Croix, C.; Sowa, G.; Pola, E.; Robbins, P.D.; Kang, J.; et al. Mitochondrial-derived reactive oxygen species (ROS) play a causal role in aging-related intervertebral disc degeneration. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2013, 31, 1150–1157. [Google Scholar] [CrossRef] [Green Version]
- Dimozi, A.; Mavrogonatou, E.; Sklirou, A.; Kletsas, D. Oxidative stress inhibits the proliferation, induces premature senescence and promotes a catabolic phenotype in human nucleus pulposus intervertebral disc cells. Eur. Cells Mater. 2015, 30, 89–102. [Google Scholar] [CrossRef]
- Cheng, X.; Ni, B.; Zhang, F.; Hu, Y.; Zhao, J. High Glucose-Induced Oxidative Stress Mediates Apoptosis and Extracellular Matrix Metabolic Imbalances Possibly via p38 MAPK Activation in Rat Nucleus Pulposus Cells. J. Diabetes Res. 2016, 2016, 3765173. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Tan, Z.; Xie, C.; Ling, L.; Hu, H. Oxidative stress as a critical factor might involve in intervertebral disc degeneration via regulating NOXs/FOXOs. J. Orthop. Sci. Off. J. Jpn. Orthop. Assoc. 2021. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Yang, S.H.; Lin, F.H. Thermosensitive chitosan-gelatin-glycerol phosphate hydrogel as a controlled release system of ferulic acid for nucleus pulposus regeneration. Biomaterials 2011, 32, 6953–6961. [Google Scholar] [CrossRef]
- Feng, G.; Zha, Z.; Huang, Y.; Li, J.; Wang, Y.; Ke, W.; Chen, H.; Liu, L.; Song, Y.; Ge, Z. Sustained and Bioresponsive Two-Stage Delivery of Therapeutic miRNA via Polyplex Micelle-Loaded Injectable Hydrogels for Inhibition of Intervertebral Disc Fibrosis. Adv. Healthc. Mater. 2018, 7, e1800623. [Google Scholar] [CrossRef]
- Apte, S.S. A disintegrin-like and metalloprotease (reprolysin-type) with thrombospondin type 1 motif (ADAMTS) superfamily: Functions and mechanisms. J. Biol. Chem. 2009, 284, 31493–31497. [Google Scholar] [CrossRef] [Green Version]
- Dubail, J.; Apte, S.S. Insights on ADAMTS proteases and ADAMTS-like proteins from mammalian genetics. Matrix Biol. J. Int. Soc. Matrix Biol. 2015, 44-46, 24–37. [Google Scholar] [CrossRef] [PubMed]
- Gendron, C.; Kashiwagi, M.; Lim, N.H.; Enghild, J.J.; Thøgersen, I.B.; Hughes, C.; Caterson, B.; Nagase, H. Proteolytic activities of human ADAMTS-5: Comparative studies with ADAMTS-4. J. Biol. Chem. 2007, 282, 18294–18306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pockert, A.J.; Richardson, S.M.; Le Maitre, C.L.; Lyon, M.; Deakin, J.A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Modified expression of the ADAMTS enzymes and tissue inhibitor of metalloproteinases 3 during human intervertebral disc degeneration. Arthritis Rheum. 2009, 60, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Markova, D.; Anderson, D.G.; Zheng, Z.; Shapiro, I.M.; Risbud, M.V. TNF-α and IL-1β promote a disintegrin-like and metalloprotease with thrombospondin type I motif-5-mediated aggrecan degradation through syndecan-4 in intervertebral disc. J. Biol. Chem. 2011, 286, 39738–39749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtwängler, T.; Chan, S.C.; Bahrenberg, G.; Richards, P.J.; Gantenbein-Ritter, B. Assessment of the matrix degenerative effects of MMP-3, ADAMTS-4, and HTRA1, injected into a bovine intervertebral disc organ culture model. Spine 2013, 38, E1377–E1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Tan, J.; Sun, J.; Fang, P.; Chen, J.; Yuan, W.; Chen, H.; Liu, Y. Transcriptomics Study to Determine the Molecular Mechanism by which sIL-13Rα2-Fc Inhibits Caudal Intervertebral Disc Degeneration in Rats. BioMed Res. Int. 2020, 2020, 7645989. [Google Scholar] [CrossRef]
- Chen, B.; Liu, Y.; Zhang, Y.; Li, J.; Cheng, K.; Cheng, L. IL-21 Is Positively Associated with Intervertebral Disc Degeneration by Interaction with TNF-α Through the JAK-STAT Signaling Pathway. Inflammation 2017, 40, 612–622. [Google Scholar] [CrossRef]
- Wang, X.; Li, C.; Liang, A.; Peng, Y.; Sun, J.; Huang, D.; Xu, K.; Ye, W. Regulation of a disintegrins and metalloproteinase with thrombospondin motifs 7 during inflammation in nucleus pulposus (NP) cells: Role of AP-1, Sp1 and NF-κB signaling. Inflamm. Res. 2016, 65, 951–962. [Google Scholar] [CrossRef]
- Yu, H.; Zhu, Y. Expression of ADAMTS-7 and ADAMTS-12 in the nucleus pulposus during degeneration of rat caudal intervetebral disc. J. Vet. Med Sci. 2012, 74, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Menou, A.; Duitman, J.; Crestani, B. The impaired proteases and anti-proteases balance in Idiopathic Pulmonary Fibrosis. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 68-69, 382–403. [Google Scholar] [CrossRef]
- Ariga, K.; Yonenobu, K.; Nakase, T.; Kaneko, M.; Okuda, S.; Uchiyama, Y.; Yoshikawa, H. Localization of cathepsins D, K, and L in degenerated human intervertebral discs. Spine 2001, 26, 2666–2672. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.E.; Ingram, J.A.; Hoelscher, G.L.; Zinchenko, N.; Norton, H.J.; Hanley, E.N., Jr. Constitutive expression of cathepsin K in the human intervertebral disc: New insight into disc extracellular matrix remodeling via cathepsin K and receptor activator of nuclear factor-κB ligand. Arthritis Res. Ther. 2011, 13, R140. [Google Scholar] [CrossRef] [Green Version]
- Kague, E.; Turci, F.; Newman, E.; Yang, Y.; Brown, K.R.; Aglan, M.S.; Otaify, G.A.; Temtamy, S.A.; Ruiz-Perez, V.L.; Cross, S.; et al. 3D assessment of intervertebral disc degeneration in zebrafish identifies changes in bone density that prime disc disease. Bone Res. 2021, 9, 39. [Google Scholar] [CrossRef]
- Konttinen, Y.T.; Kääpä, E.; Hukkanen, M.; Gu, X.H.; Takagi, M.; Santavirta, S.; Alaranta, H.; Li, T.F.; Suda, A. Cathepsin G in degenerating and healthy discal tissue. Clin. Exp. Rheumatol. 1999, 17, 197–204. [Google Scholar] [PubMed]
- Tiaden, A.N.; Klawitter, M.; Lux, V.; Mirsaidi, A.; Bahrenberg, G.; Glanz, S.; Quero, L.; Liebscher, T.; Wuertz, K.; Ehrmann, M.; et al. Detrimental role for human high temperature requirement serine protease A1 (HTRA1) in the pathogenesis of intervertebral disc (IVD) degeneration. J. Biol. Chem. 2012, 287, 21335–21345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Li, D.; Wu, H.; Liu, F.; Liu, F.; Zhang, Q.; Li, J. LncRNA TRPC7-AS1 regulates nucleus pulposus cellular senescence and ECM synthesis via competing with HPN for miR-4769-5p binding. Mech. Ageing Dev. 2020, 190, 111293. [Google Scholar] [CrossRef] [PubMed]
- Salo, J.; Mackiewicz, Z.; Indahl, A.; Konttinen, Y.T.; Holm, A.K.; Sukura, A.; Holm, S. Plasmin-matrix metalloproteinase cascades in spinal response to an experimental disc lesion in pig. Spine 2008, 33, 839–844. [Google Scholar] [CrossRef]
- Rodrigues, L.M.; Theodoro, T.R.; Matos, L.L.; Mader, A.M.; Milani, C.; Pinhal, M.A. Heparanase isoform expression and extracellular matrix remodeling in intervertebral disc degenerative disease. Clinics 2011, 66, 903–909. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.M.; Lu, C.C.; Shen, P.C.; Chou, S.H.; Shih, C.L.; Chen, J.C.; Tien, Y.C. Suramin attenuates intervertebral disc degeneration by inhibiting NF-κB signalling pathway. Bone Jt. Res. 2021, 10, 498–513. [Google Scholar] [CrossRef]
- Yao, Z.; Nie, L.; Zhao, Y.; Zhang, Y.; Liu, Y.; Li, J.; Cheng, L. Salubrinal Suppresses IL-17-Induced Upregulation of MMP-13 and Extracellular Matrix Degradation Through the NF-kB Pathway in Human Nucleus Pulposus Cells. Inflammation 2016, 39, 1997–2007. [Google Scholar] [CrossRef]
- Xie, C.; Ma, H.; Shi, Y.; Li, J.; Wu, H.; Wang, B.; Shao, Z.; Huang, C.; Chen, J.; Sun, L.; et al. Cardamonin protects nucleus pulposus cells against IL-1β-induced inflammation and catabolism via Nrf2/NF-κB axis. Food Funct. 2021, 12, 2703–2714. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Li, Y.; Ma, Z.; Zhao, J. Crocin exerts anti-inflammatory and anti-catabolic effects on rat intervertebral discs by suppressing the activation of JNK. Int. J. Mol. Med. 2015, 36, 1291–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K.; Li, Y.; Xu, B.; Mao, L.; Zhao, J. Sesamin inhibits lipopolysaccharide-induced inflammation and extracellular matrix catabolism in rat intervertebral disc. Connect. Tissue Res. 2016, 57, 347–359. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Holt, D.C., 3rd; Smith, R.; Kim, S.J.; Gardocki, R.J.; Hasty, K.A. The Effects of Platelet-Rich Plasma on Halting the Progression in Porcine Intervertebral Disc Degeneration. Artif. Organs 2016, 40, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Yeom, J.S.; Koh, Y.G.; Yeo, J.E.; Kang, K.T.; Kang, Y.M.; Chang, B.S.; Lee, C.K. Anti-inflammatory effect of platelet-rich plasma on nucleus pulposus cells with response of TNF-α and IL-1. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2014, 32, 551–556. [Google Scholar] [CrossRef]
- Qin, R.; Dai, S.; Zhang, X.; Liu, H.; Zhou, B.; Zhou, P.; Hu, C. Danshen Attenuates Intervertebral Disc Degeneration via Antioxidation in SD Rats. Oxidative Med. Cell. Longev. 2020, 2020, 6660429. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Hu, S.; Wang, B.; Wang, J.; Wang, X.; Xu, C. Genistein protects intervertebral discs from degeneration via Nrf2-mediated antioxidant defense system: An in vitro and in vivo study. J. Cell. Physiol. 2019. [Google Scholar] [CrossRef]
- Song, Y.; Wang, Z.; Liu, L.; Zhang, S.; Zhang, H.; Qian, Y. 1,4-Dihydropyridine (DHP) suppresses against oxidative stress in nucleus pulposus via activating sirtuin-1. Biomed. Pharmacother. Biomed. Pharmacother. 2020, 121, 109592. [Google Scholar] [CrossRef]
- Yang, F.; Leung, V.Y.; Luk, K.D.; Chan, D.; Cheung, K.M. Mesenchymal stem cells arrest intervertebral disc degeneration through chondrocytic differentiation and stimulation of endogenous cells. Mol. Ther. J. Am. Soc. Gene Ther. 2009, 17, 1959–1966. [Google Scholar] [CrossRef]
- Richardson, S.M.; Walker, R.V.; Parker, S.; Rhodes, N.P.; Hunt, J.A.; Freemont, A.J.; Hoyland, J.A. Intervertebral disc cell-mediated mesenchymal stem cell differentiation. Stem Cells 2006, 24, 707–716. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Guanjun, T.; Lin, C.; Zifeng, P. Effects of Transplantation of hTIMP-1-Expressing Bone Marrow Mesenchymal Stem Cells on the Extracellular Matrix of Degenerative Intervertebral Discs in an In Vivo Rabbit Model. Spine 2014, 39, E669–E675. [Google Scholar] [CrossRef] [PubMed]
- Frapin, L.; Clouet, J.; Chédeville, C.; Moraru, C.; Samarut, E.; Henry, N.; André, M.; Bord, E.; Halgand, B.; Lesoeur, J.; et al. Controlled release of biological factors for endogenous progenitor cell migration and intervertebral disc extracellular matrix remodelling. Biomaterials 2020, 253, 120107. [Google Scholar] [CrossRef] [PubMed]
- Yao, M.; Zhang, J.; Li, Z.; Guo, S.; Zhou, X.; Zhang, W. Marein protects human nucleus pulposus cells against high glucose-induced injury and extracellular matrix degradation at least partly by inhibition of ROS/NF-κB pathway. Int. Immunopharmacol. 2020, 80, 106126. [Google Scholar] [CrossRef] [PubMed]
- Illien-Junger, S.; Grosjean, F.; Laudier, D.M.; Vlassara, H.; Striker, G.E.; Iatridis, J.C. Combined anti-inflammatory and anti-AGE drug treatments have a protective effect on intervertebral discs in mice with diabetes. PLoS ONE 2013, 8, e64302. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Zhang, L.; Wang, W.Y.; Hu, Q.F.; Song, H.P.; Su, Y.L.; Zhang, Y.Z. Alendronate retards the progression of lumbar intervertebral disc degeneration in ovariectomized rats. Bone 2013, 55, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Tian, F.M.; Wang, P.; Gou, Y.; Zhang, H.; Song, H.P.; Wang, W.Y.; Zhang, L. Alendronate Prevents Intervertebral Disc Degeneration Adjacent to a Lumbar Fusion in Ovariectomized Rats. Spine 2015, 40, E1073–E1083. [Google Scholar] [CrossRef] [Green Version]
- Song, H.; Luo, Y.; Wang, W.; Li, S.; Yang, K.; Dai, M.; Shen, Y.; Zhang, Y.; Zhang, L. Effects of alendronate on lumbar intervertebral disc degeneration with bone loss in ovariectomized rats. Spine J. Off. J. North Am. Spine Soc. 2017, 17, 995–1003. [Google Scholar] [CrossRef]
- Cheng, Y.H.; Yang, S.H.; Liu, C.C.; Gefen, A.; Lin, F.H. Thermosensitive hydrogel made of ferulic acid-gelatin and chitosan glycerophosphate. Carbohydr. Polym. 2013, 92, 1512–1519. [Google Scholar] [CrossRef]
- Larrañaga, A.; Isa, I.L.M.; Patil, V.; Thamboo, S.; Lomora, M.; Fernández-Yague, M.A.; Sarasua, J.R.; Palivan, C.G.; Pandit, A. Antioxidant functionalized polymer capsules to prevent oxidative stress. Acta Biomater. 2018, 67, 21–31. [Google Scholar] [CrossRef]
- Liu, Y.; Du, J.; Peng, P.; Cheng, R.; Lin, J.; Xu, C.; Yang, H.; Cui, W.; Mao, H.; Li, Y.; et al. Regulation of the inflammatory cycle by a controllable release hydrogel for eliminating postoperative inflammation after discectomy. Bioact. Mater. 2021, 6, 146–157. [Google Scholar] [CrossRef]
- Xu, Y.; Gu, Y.; Cai, F.; Xi, K.; Xin, T.; Tang, J.; Wu, L.; Wang, Z.; Wang, F.; Deng, L.; et al. Metabolism Balance Regulation via Antagonist-Functionalized Injectable Microsphere for Nucleus Pulposus Regeneration. Adv. Funct. Mater. 2020, 30, 2006333. [Google Scholar] [CrossRef]
- Bian, J.; Cai, F.; Chen, H.; Tang, Z.; Xi, K.; Tang, J.; Wu, L.; Xu, Y.; Deng, L.; Gu, Y.; et al. Modulation of Local Overactive Inflammation via Injectable Hydrogel Microspheres. Nano Lett. 2021, 21, 2690–2698. [Google Scholar] [CrossRef] [PubMed]
- Leckie, S.K.; Bechara, B.P.; Hartman, R.A.; Sowa, G.A.; Woods, B.I.; Coelho, J.P.; Witt, W.T.; Dong, Q.D.; Bowman, B.W.; Bell, K.M.; et al. Injection of AAV2-BMP2 and AAV2-TIMP1 into the nucleus pulposus slows the course of intervertebral disc degeneration in an in vivo rabbit model. Spine J. Off. J. North Am. Spine Soc. 2012, 12, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Yu, T.; Ma, X.X.; Xiang, H.F.; Hu, Y.G.; Chen, B.H. Lentivirus-mediated TGF-β3, CTGF and TIMP1 gene transduction as a gene therapy for intervertebral disc degeneration in an in vivo rabbit model. Exp. Ther. Med. 2016, 11, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Seki, S.; Asanuma-Abe, Y.; Masuda, K.; Kawaguchi, Y.; Asanuma, K.; Muehleman, C.; Iwai, A.; Kimura, T. Effect of small interference RNA (siRNA) for ADAMTS5 on intervertebral disc degeneration in the rabbit anular needle-puncture model. Arthritis Res. Ther. 2009, 11, R166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Lin, J.; Wu, X.; Guo, X.; Sun, H.; Yu, B.; Shen, J.; Bai, J.; Chen, Z.; Yang, H.; et al. Aspirin-Mediated Attenuation of Intervertebral Disc Degeneration by Ameliorating Reactive Oxygen Species In Vivo and In Vitro. Oxidative Med. Cell. Longev. 2019, 2019, 7189854. [Google Scholar] [CrossRef] [Green Version]
- He, R.; Cui, M.; Lin, H.; Zhao, L.; Wang, J.; Chen, S.; Shao, Z. Melatonin resists oxidative stress-induced apoptosis in nucleus pulposus cells. Life Sci. 2018, 199, 122–130. [Google Scholar] [CrossRef]
- Suzuki, S.; Fujita, N.; Hosogane, N.; Watanabe, K.; Ishii, K.; Toyama, Y.; Takubo, K.; Horiuchi, K.; Miyamoto, T.; Nakamura, M.; et al. Excessive reactive oxygen species are therapeutic targets for intervertebral disc degeneration. Arthritis Res. Ther. 2015, 17, 316. [Google Scholar] [CrossRef] [Green Version]
- Hiyama, A.; Mochida, J.; Iwashina, T.; Omi, H.; Watanabe, T.; Serigano, K.; Tamura, F.; Sakai, D. Transplantation of mesenchymal stem cells in a canine disc degeneration model. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2008, 26, 589–600. [Google Scholar] [CrossRef]
- Wang, Z.; Ding, X.; Cao, F.; Zhang, X.; Wu, J. Bone Mesenchymal Stem Cells Promote Extracellular Matrix Remodeling of Degenerated Nucleus Pulposus Cells via the miR-101-3p/EIF4G2 Axis. Front. Bioeng. Biotechnol. 2021, 9, 642502. [Google Scholar] [CrossRef]
- Feng, G.; Zhang, Z.; Dang, M.; Rambhia, K.J.; Ma, P.X. Nanofibrous spongy microspheres to deliver rabbit mesenchymal stem cells and anti-miR-199a to regenerate nucleus pulposus and prevent calcification. Biomaterials 2020, 256, 120213. [Google Scholar] [CrossRef] [PubMed]
- Ukeba, D.; Sudo, H.; Tsujimoto, T.; Ura, K.; Yamada, K.; Iwasaki, N. Bone marrow mesenchymal stem cells combined with ultra-purified alginate gel as a regenerative therapeutic strategy after discectomy for degenerated intervertebral discs. EBioMedicine 2020, 53, 102698. [Google Scholar] [CrossRef] [PubMed]
- Horner, H.A.; Urban, J.P. 2001 Volvo Award Winner in Basic Science Studies: Effect of nutrient supply on the viability of cells from the nucleus pulposus of the intervertebral disc. Spine 2001, 26, 2543–2549. [Google Scholar] [CrossRef]
- Urban, J.P.; Smith, S.; Fairbank, J.C. Nutrition of the intervertebral disc. Spine 2004, 29, 2700–2709. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Liu, H.; Ma, L.; Lei, J.; Tong, B.; Li, G.; Ke, W.; Wang, K.; Feng, X.; Hua, W.; et al. Engineering Extracellular Vesicles Restore the Impaired Cellular Uptake and Attenuate Intervertebral Disc Degeneration. ACS Nano 2021, 15, 14709–14724. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Qu, J.N.; Zhang, Y.G. Animal models of disc degeneration and major genetic strategies. Pain Physician 2013, 16, E267–E275. [Google Scholar] [PubMed]
- Woods, B.I.; Vo, N.; Sowa, G.; Kang, J.D. Gene therapy for intervertebral disk degeneration. Orthop. Clin. North Am. 2011, 42, 563–574. [Google Scholar] [CrossRef]
- Binch, A.L.A.; Fitzgerald, J.C.; Growney, E.A.; Barry, F. Cell-based strategies for IVD repair: Clinical progress and translational obstacles. Nat. Rev. Rheumatol. 2021, 17, 158–175. [Google Scholar] [CrossRef]
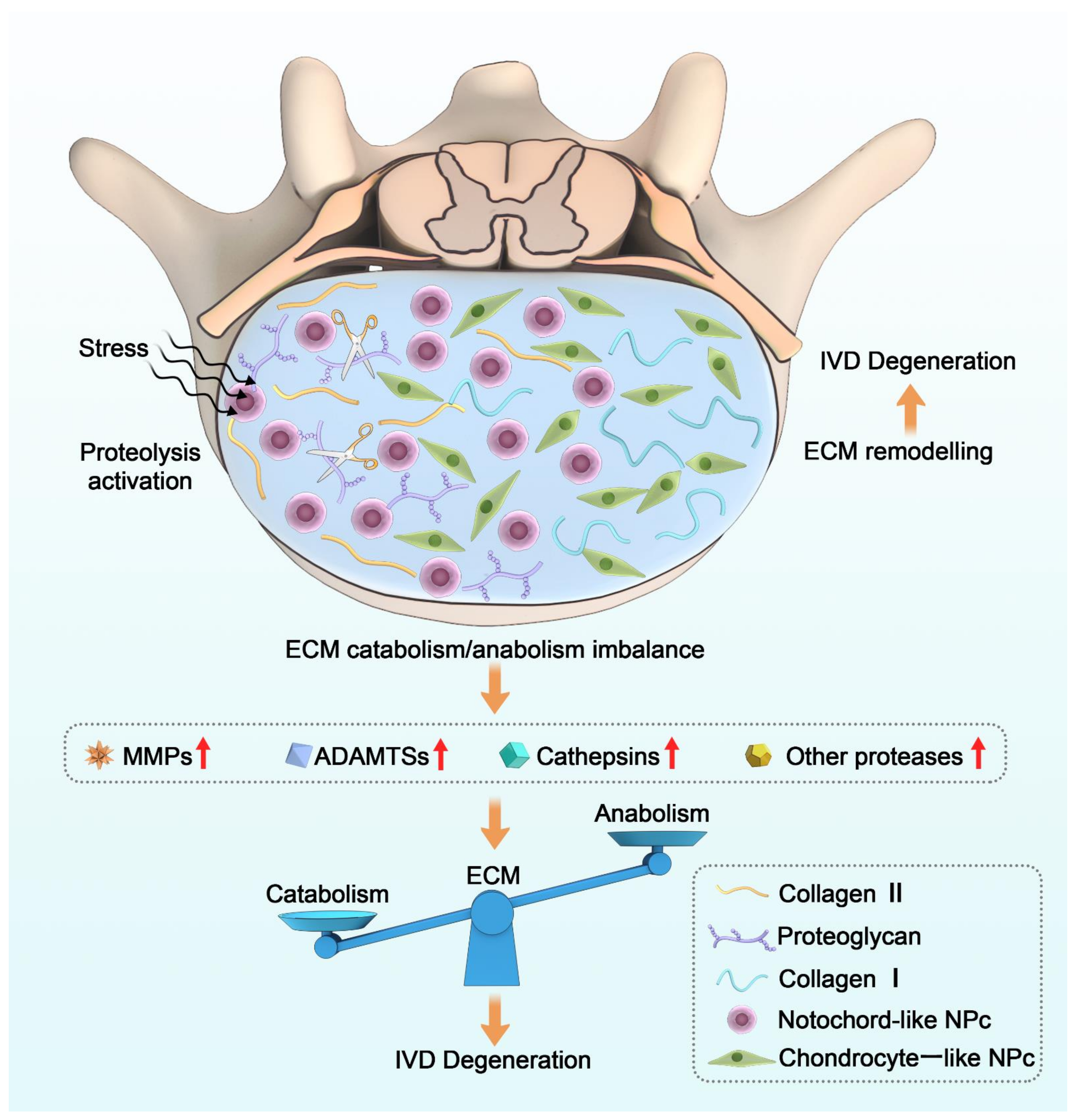
Figure 1.
General mechanisms of the pathological process of IDD and the role of proteases in the imbalance of ECM metabolism. Proteolysis activation leads to an imbalance in ECM homeostasis when the IVD is subjected to various stimuli, in which proteases play a key role.
Figure 1.
General mechanisms of the pathological process of IDD and the role of proteases in the imbalance of ECM metabolism. Proteolysis activation leads to an imbalance in ECM homeostasis when the IVD is subjected to various stimuli, in which proteases play a key role.


{kind=link}
Table 1.
Recent advances in modulating ECM homeostasis for the treatment of IDD.
| Strategies | Interventions | Effects | Reference(s) |
|---|---|---|---|
| Anti-inflammation | Cardamonin, Salubrinal, Suramin | Downregulate multiple proteases by inhibiting NF-κB | [99,100,101] |
| Crocin, Sesamin | Downregulate multiple proteases by inhibiting JNK | [102,103] | |
| PRP | Downregulate multiple MMPs and increase levels of several beneficial growth factors | [104,105] | |
| Anti-oxidation | Danshen | Downregulate MMP-3 | [106] |
| Genistein | Downregulate MMP-13 via Nrf2-mediated antioxidant system | [107] | |
| DHP | Downregulate MMP-3 and ADAMTS-5 via activating sirtuin-1 | [108] | |
| Stem cells therapy | Stem cell implantation | Induce differentiation of stem cells into nucleus pulposus cells and stimulate endogenous ECM regeneration | [109,110] |
| TIMP-1 overexpression modified BMSCs | Modulate ECM anabolic catabolic homeostasis | [111] | |
| Chemokine CCL-5 | Recruit disc stem/progenitor cells to nucleus pulposus | [112] | |
| Metabolic modulation | Marein | Downregulate MMP-3 and MMP-13 | [113] |
| Pyridoxamine | Downregulate MMP-13 and ADAMTS-5 by antagonizing AGE | [114] | |
| ALN | Downregulate MMP-1, MMP-3 and MMP-13 | [115,116,117] | |
| Biomaterials | FA-G/C/GP hydrogel | Downregulate MMP-3 and upregulate aggrecan and type II collagen | [78,118] |
| TA-functionalized polymer capsules | Downregulate MMP-3 and ADAMTS-5 | [119] | |
| Aspirin controllable release hydrogel | Downregulate MMP-3/13 and ADAMTS-4/5 | [120] | |
| Injectable microspheres load with TNFRII or APETx2 | Downregulate MMP-3 and ADAMTS-5 via modulate local inflammation microenvironment | [121,122] | |
| Gene therapy | AAV2-TIMP1 | promote synthesis of type II collagen | [123] |
| TGF-β3, CTGF and TIMP1 co-transduction | Promote synthesis of aggrecan and type II collagen | [124] | |
| ADAMTS-5 siRNA | Downregulate ADAMTS-5 | [125] |
PRP: Platelet-Rich Plasma, DHP: 1,4-dihydropyridine, TIMP-1: tissue inhibitor of metalloproteinase 1, CCL-5: C-C motif chemokine ligand 5, ALN: alendronate, TNFRII: tumor necrosis factor receptor type II. TGF-β3: transforming growth factor, CTGF: connective tissue growth factor, ADAMTS: A disintegrin and metalloprotease with thrombospondin motifs, MMP: Matrix metalloproteinase.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liang, H.; Luo, R.; Li, G.; Zhang, W.; Song, Y.; Yang, C. The Proteolysis of ECM in Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2022, 23, 1715. https://doi.org/10.3390/ijms23031715
AMA Style
Liang H, Luo R, Li G, Zhang W, Song Y, Yang C. The Proteolysis of ECM in Intervertebral Disc Degeneration. International Journal of Molecular Sciences. 2022; 23(3):1715. https://doi.org/10.3390/ijms23031715
Chicago/Turabian StyleLiang, Huaizhen, Rongjin Luo, Gaocai Li, Weifeng Zhang, Yu Song, and Cao Yang. 2022. "The Proteolysis of ECM in Intervertebral Disc Degeneration" International Journal of Molecular Sciences 23, no. 3: 1715. https://doi.org/10.3390/ijms23031715
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.