
The Non-Gastric H+/K+ ATPase (ATP12A) Is Expressed in Mammalian Spermatozoa

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
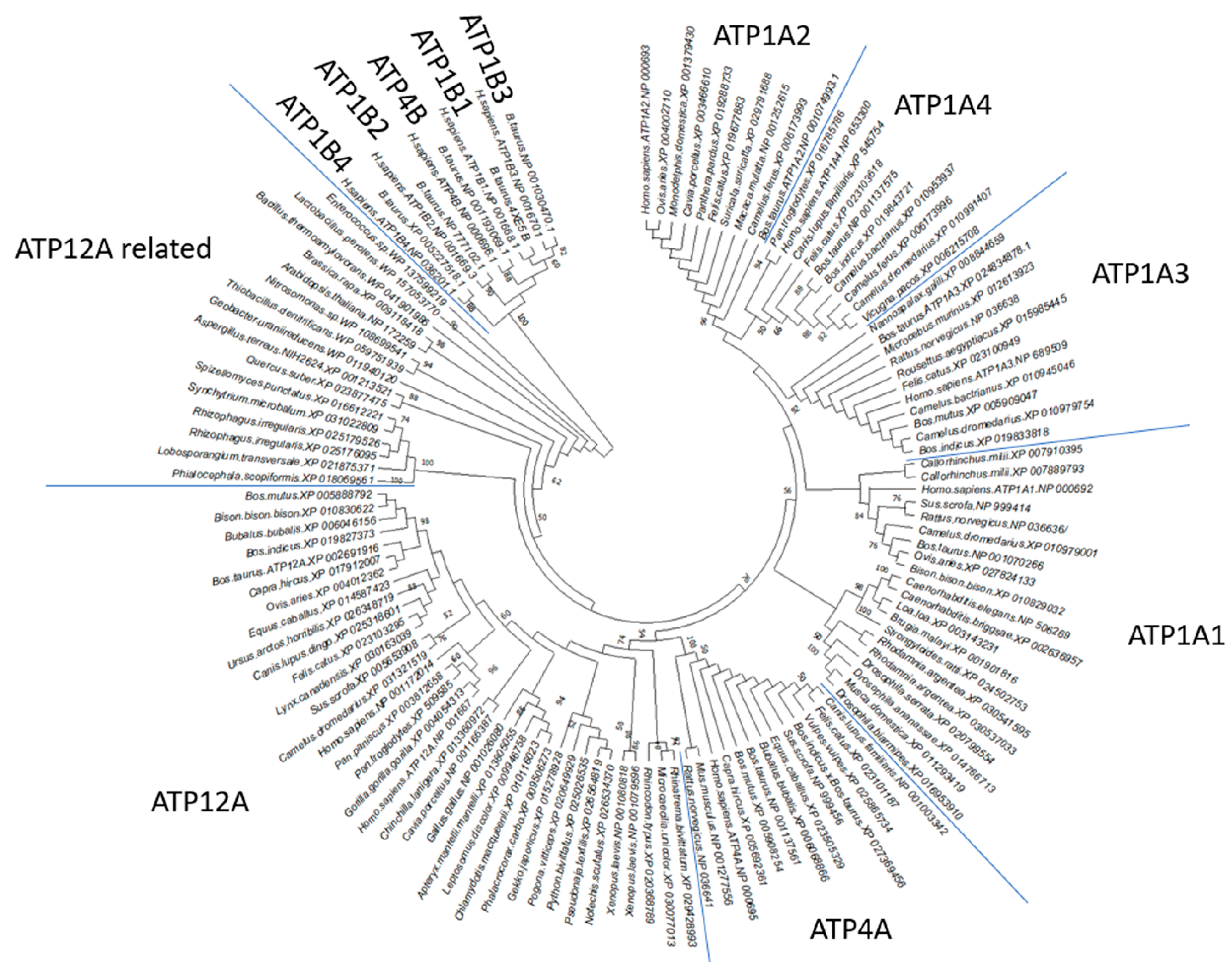
2.1. ATP12A Shows Conserved Sequence Features Detectable along Phylogenetically Distant Species That Determine a Dedicated Cluster in Phylogenetic Analysis When Compared to Other P2C-ATPase Family Members
2.2. Multiple Sequence Alignment (MSA) Highlights ATP12A Specific Sequence Features in the Targeted Epitopes despite the High Percentage of Identical Amino Acids Shared with Other α Subunits of P2C-ATPases Members
2.3. MSA Highlights ATP1B1-Specific Sequence Features in the Targeted Epitopes and a Low Percentage of Identical Amino Acids Shared with Others β Subunits of P2C-ATPases Members
2.4. MSA Highlights ATP1A1 and ATP1A4 Specific Sequence Features in Targeted Epitopes and a Low Percentage of Identical Amino Acids Shared with Other α Subunits of P2C-ATPases Members
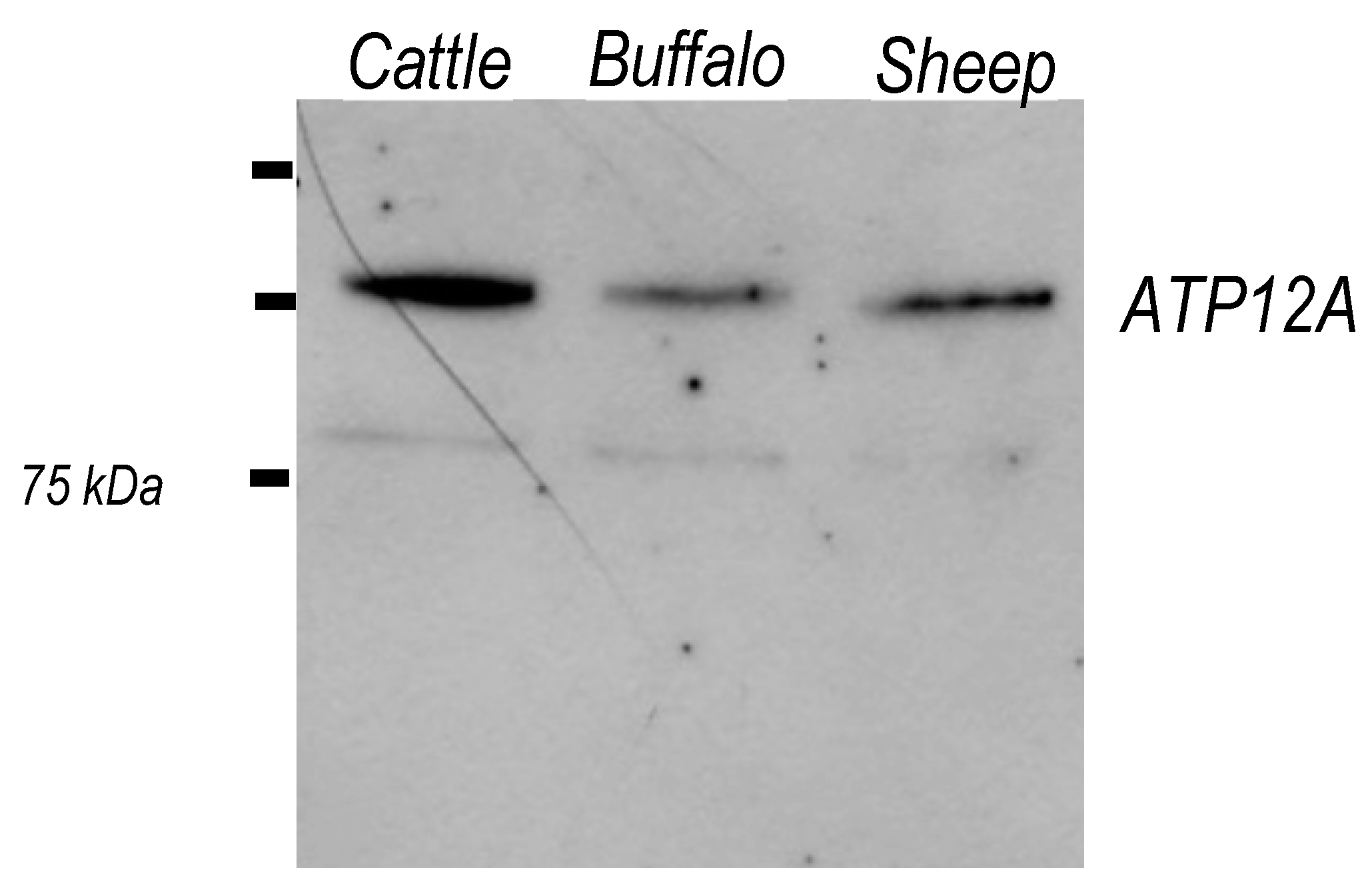
2.5. ATP12A Is Expressed in Frozen/Thawed Sperm Cells
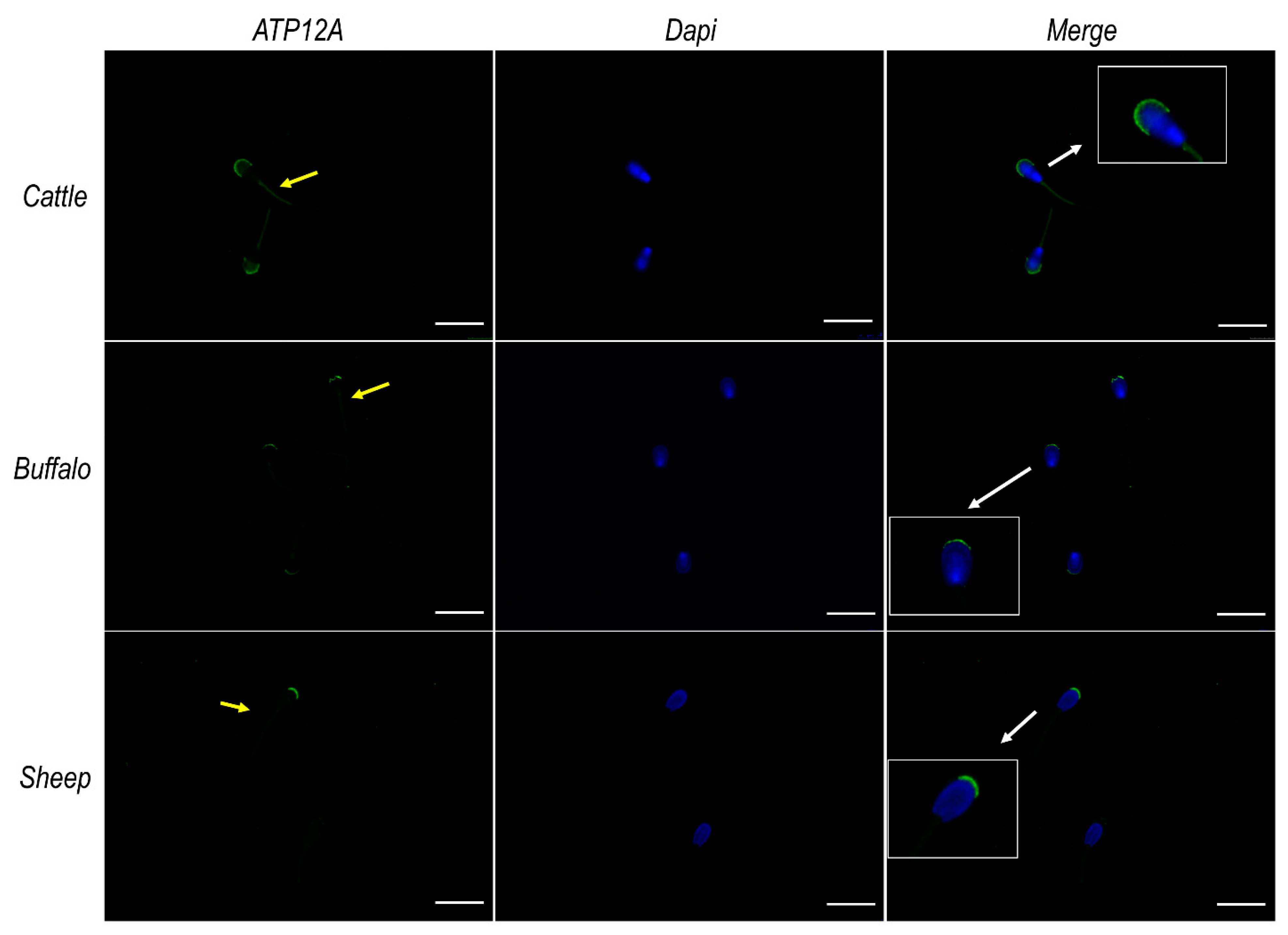
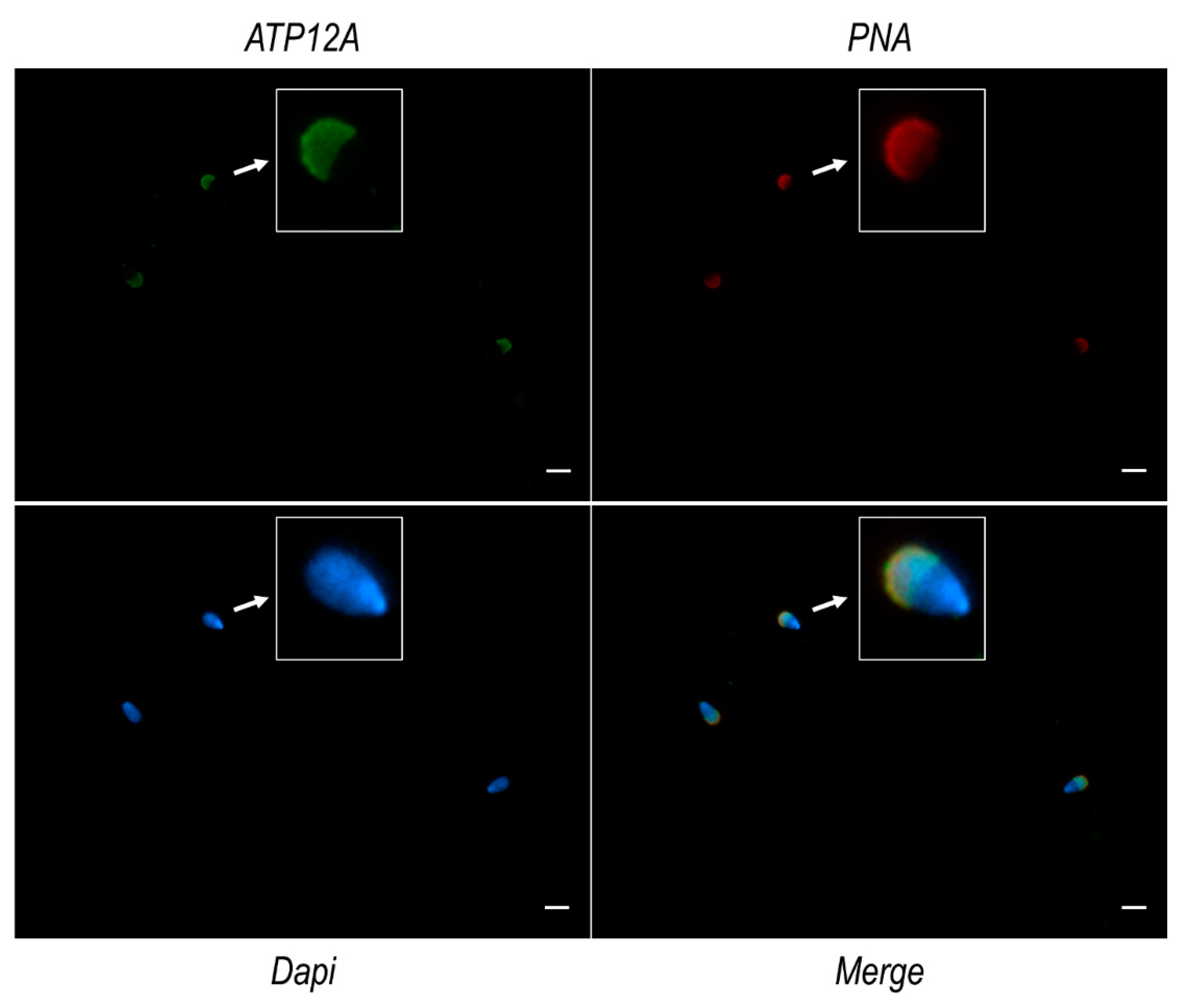
2.6. ATP12A Localizes at the Acrosomal Region in Frozen/Thawed Sperm Cells
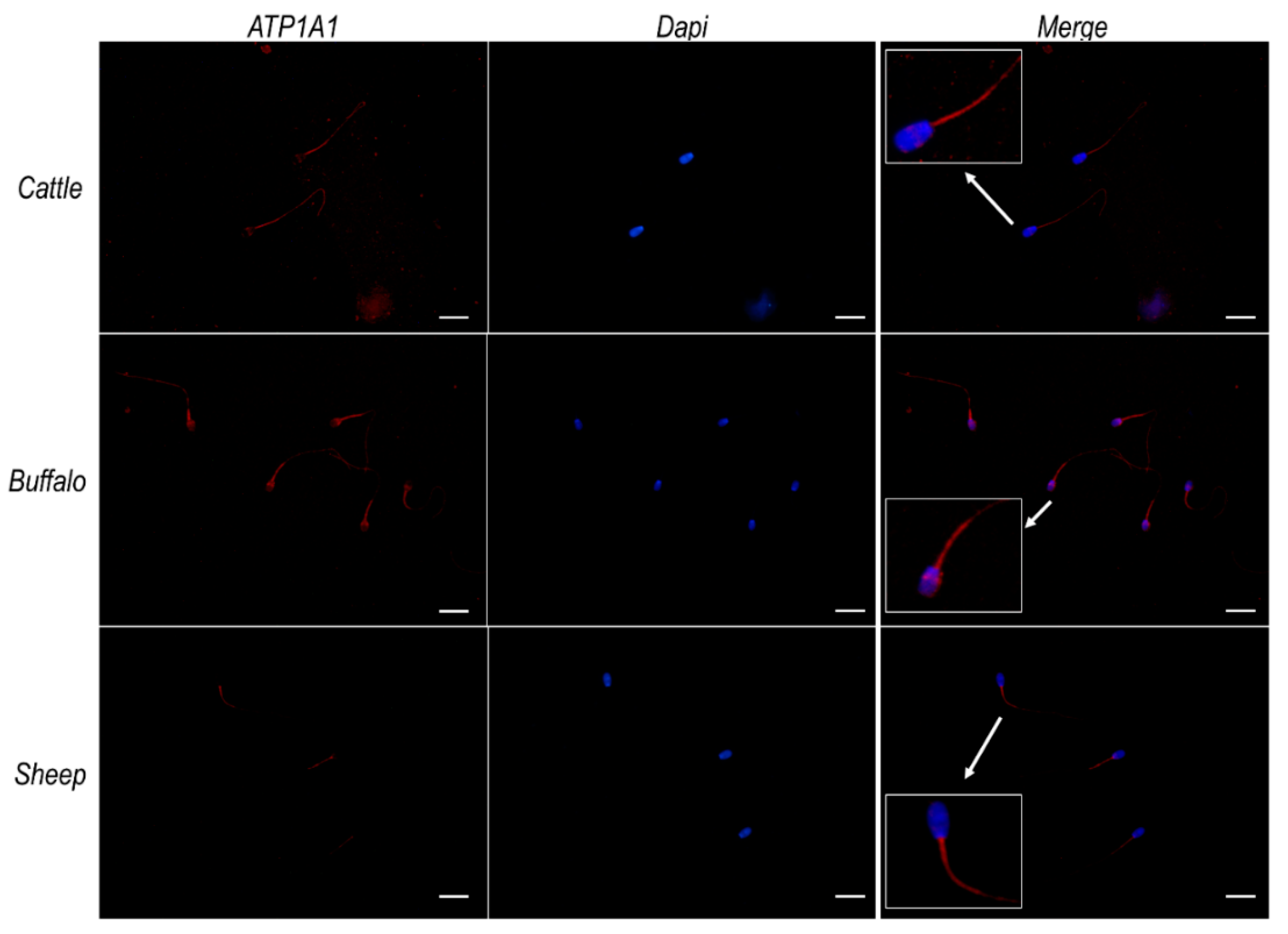
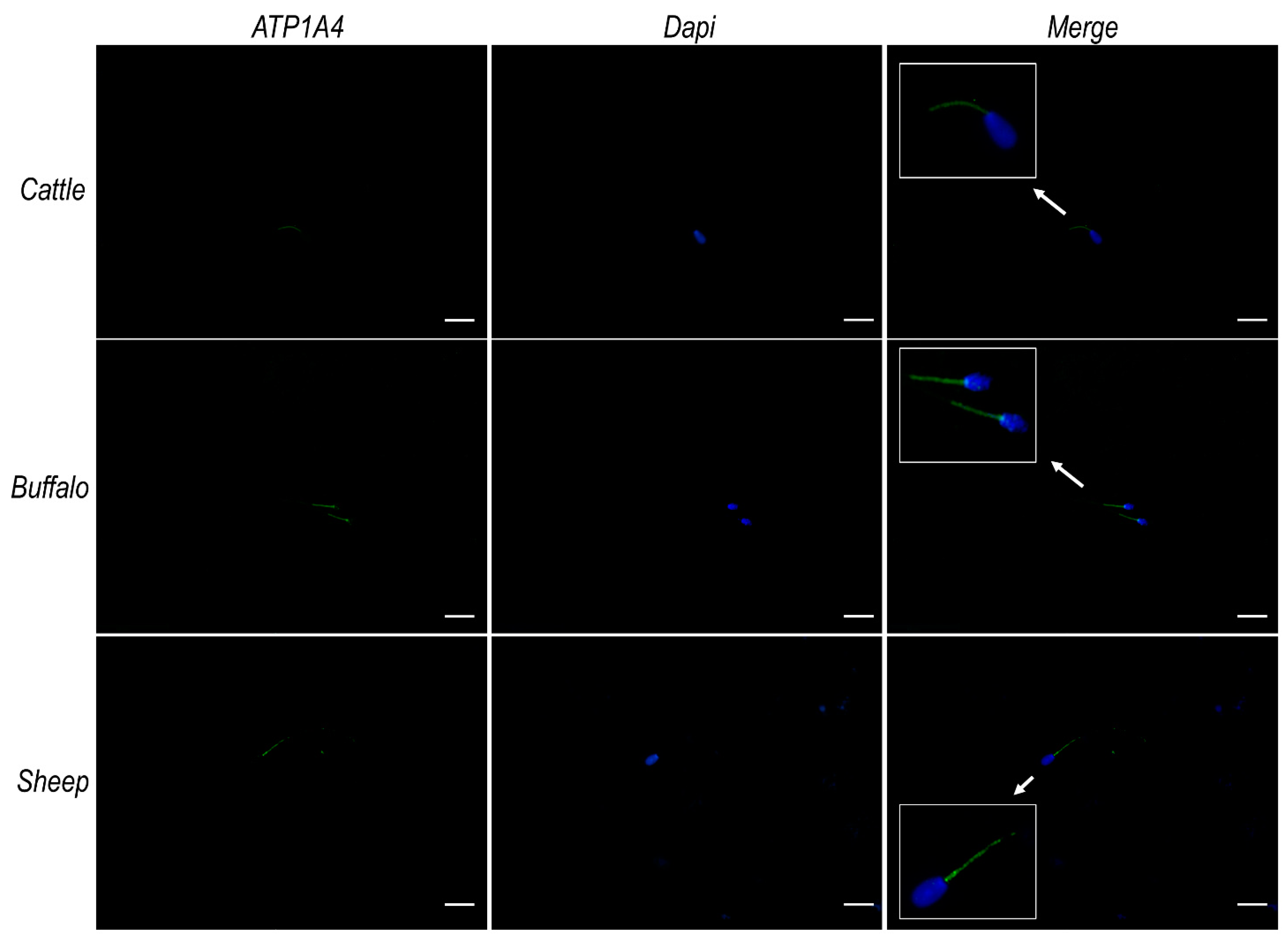
2.7. Na+/K+ ATPase Subunits α1 and α4 Do Not Colocalize with ATP12A at the Acrosomal Region in Frozen/Thawed Sperm Cells
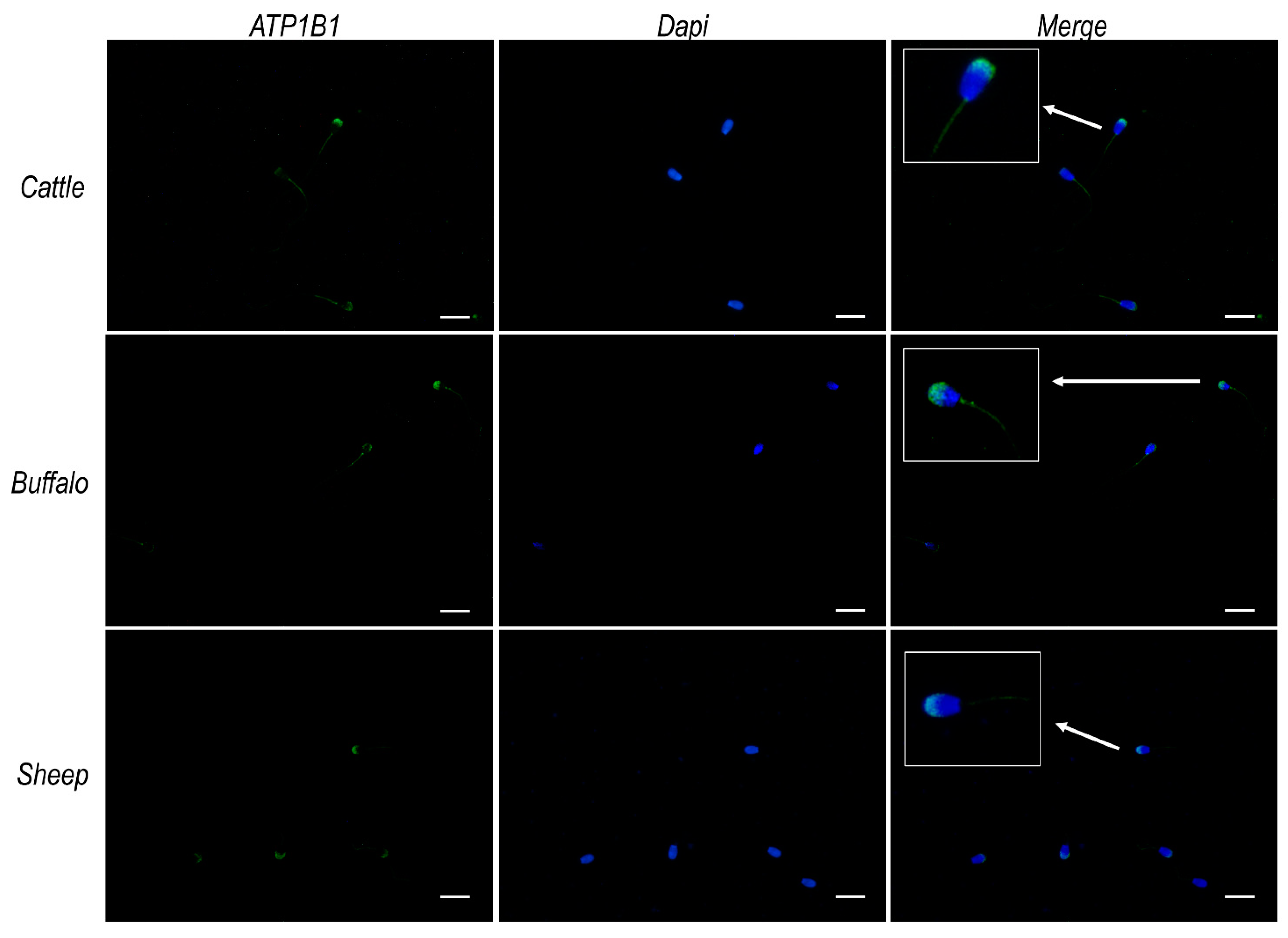
2.8. ATP12A Colocalizes with Na+/K+ ATPase Subunit β1 at the Acrosomal Region in Frozen/Thawed Sperm Cells
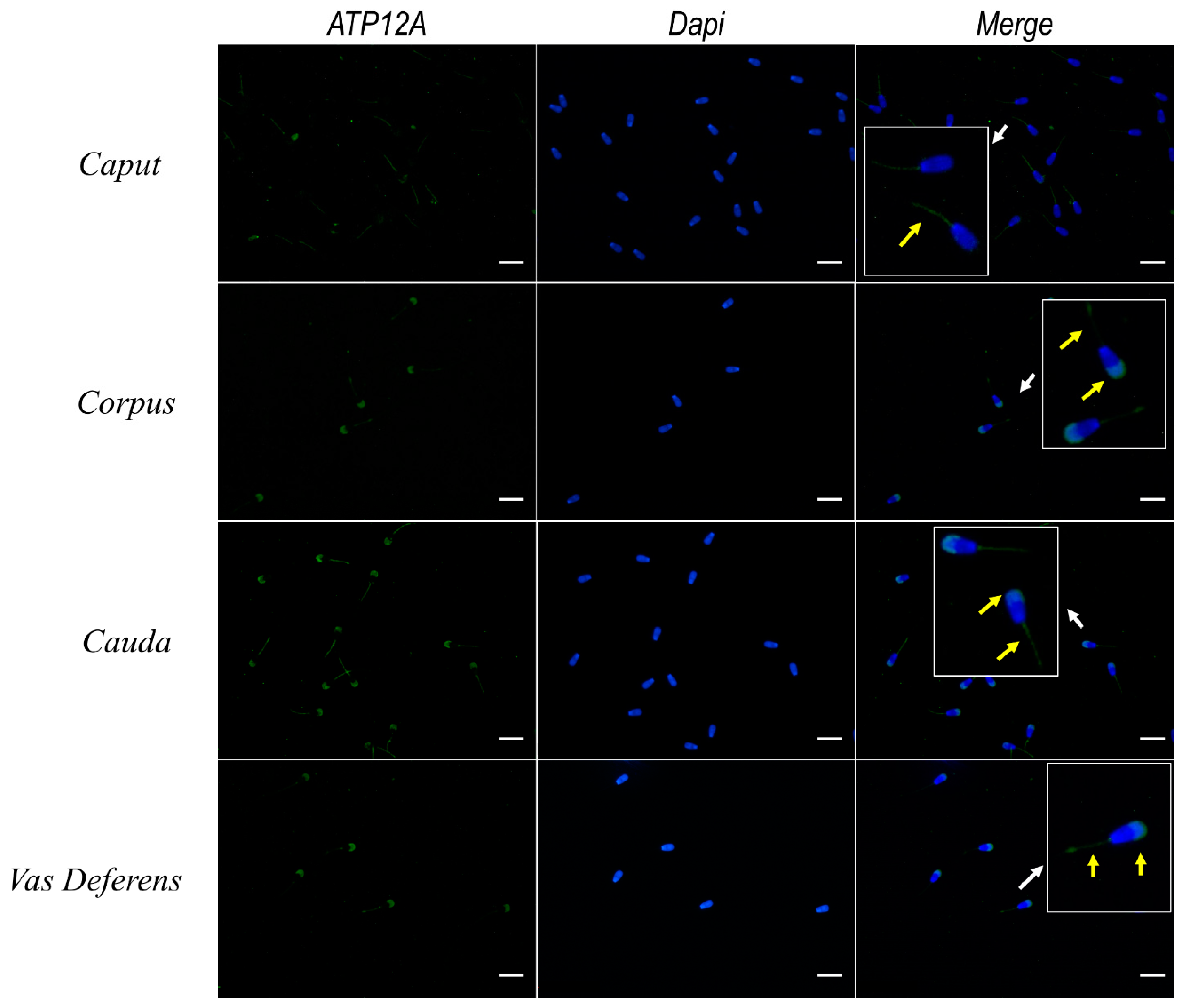
2.9. Expression of ATP12A Changes during Sperm Maturation
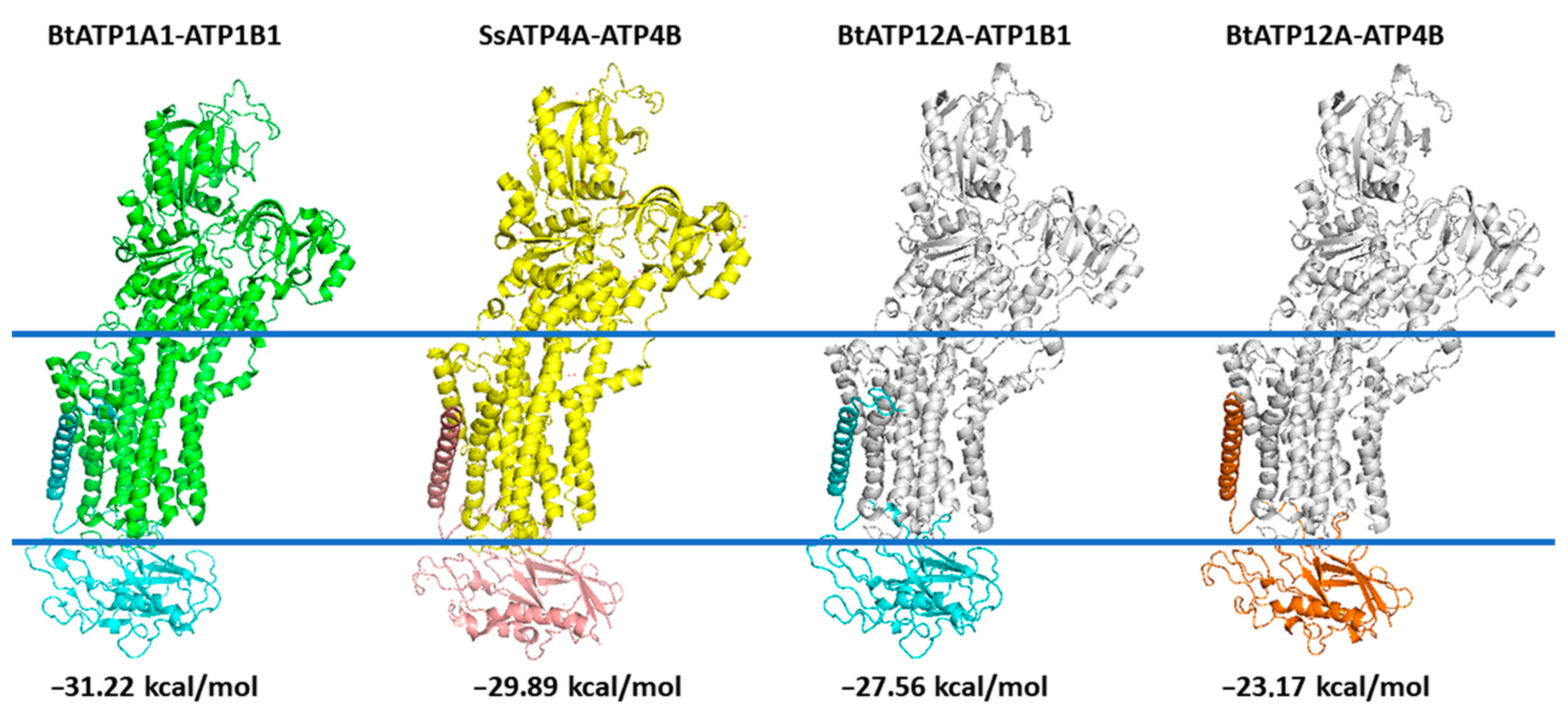
2.10. 3D Comparative Modeling of the Bovine ATP12A.ATP1B1 and ATP12A.ATP4B
2.11. Interaction Energy Calculation
3. Discussion
4. Materials and Methods
4.1. Semen Samples and Ethics Statement
4.2. Swim-Up
4.3. Recovery of Spermatozoa from Testis and Epididymis
4.4. Polyacrylamide Gel Electrophoresis and Immunoblot Analysis
4.5. Immunofluorescence Analysis
4.6. Protein Sequence Sampling and Multiple Sequence Alignment (MSA)
4.7. Phylogenetic Analysis
4.8. 3D Comparative Modeling of Bovine ATP12A, Bovine ATP4B, Bovine ATP12A.ATP1B1 and Bovine ATP12A.ATP4B Protein Complexes
4.9. Interaction Energy Calculations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Crambert, G. H-K-ATPase type 2: Relevance for renal physiology and beyond. Am. J. Physiol. Ren. Physiol. 2014, 306, F693–F700. [Google Scholar] [CrossRef] [Green Version]
- Sverdlov, V.E.; Kostina, M.B.; Modyanov, N.N. Genomic Organization of the Human ATP1AL1 Gene Encoding a Ouabain-Sensitive H,K-ATPase. Genomics 1996, 32, 317–327. [Google Scholar] [CrossRef]
- Scudieri, P.; Musante, I.; Caci, E.; Venturini, A.; Morelli, P.; Walter, C.; Tosi, D.; Palleschi, A.; Martin-Vasallo, P.; Sermet-Gaudelus, I.; et al. Increased expression of ATP12A proton pump in cystic fibrosis airways. JCI Insight 2018, 3, e123616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestov, N.B.; Korneenko, T.V.; Radkov, R.; Zhao, H.; Shakhparonov, M.I.; Modyanov, N.N. Identification of the β-subunit for nongastric H-K-ATPase in rat anterior prostate. Am. J. Physiol. Physiol. 2004, 286, C1229–C1237. [Google Scholar] [CrossRef] [PubMed]
- Pestov, N.B.; Korneenko, T.V.; Shakhparonov, M.I.; Shull, G.E.; Modyanov, N.N. Loss of acidification of anterior prostate fluids inAtp12a-null mutant mice indicates that nongastric H-K-ATPase functions as proton pump in vivo. Am. J. Physiol. Physiol. 2006, 291, C366–C374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, G.; Tillekeratne, M.; Yu, C.; Pestov, N.B.; Modyanov, N.N. Catalytic Function of Nongastric H,K-ATPase Expressed in Sf-21 Insect Cells. Biochemistry 2001, 40, 5765–5776. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, J.; Grishin, A.V.; Oberleithner, H.; Caplan, M.J. Differential localization of human nongastric H+-K+-ATPase ATP1AL1 in polarized renal epithelial cells. Am. J. Physiol. Physiol. 2000, 279, F417–F425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Codina, J.; Delmas-Mata, J.T.; DuBose, T.D.J. The α-Subunit of the Colonic H+,K+-ATPase Assembles with β1-Na+,K+-ATPase in Kidney and Distal Colon. J. Biol. Chem. 1998, 273, 7894–7899. [Google Scholar] [CrossRef] [Green Version]
- Kraut, J.A.; Hiura, J.; Shin, J.M.; Smolka, A.; Sachs, G.; Scott, D. The Na+-K+-ATPase β1 subunit is associated with the HKα2 protein in the rat kidney. Kidney Int. 1998, 53, 958–962. [Google Scholar] [CrossRef]
- Chan, H.; Babayan, V.; Blyumin, E.; Gandhi, C.; Hak, K.; Harake, D.; Kumar, K.; Lee, P.; Li, T.T.; Liu, H.Y.; et al. The P-Type ATPase Superfamily. J. Mol. Microbiol. Biotechnol. 2010, 19, 5–104. [Google Scholar] [CrossRef] [PubMed]
- van Veen, S.; Sørensen, D.M.; Holemans, T.; Holen, H.W.; Palmgren, M.G.; Vangheluwe, P. Cellular Function and Pathological Role of ATP13A2 and Related P-Type Transport ATPases in Parkinson’s Disease and Other Neurological Disorders. Front. Mol. Neurosci. 2014, 7, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belisario, D.C.; Rocafull, M.A.; del Castillo, J.R. Purification and Characterization of the Ouabain-Sensitive H+/K+-ATPase from Guinea-Pig Distal Colon. Arch. Biochem. Biophys. 2010, 496, 21–32. [Google Scholar] [CrossRef]
- Gumz, M.L.; Lynch, I.J.; Greenlee, M.M.; Cain, B.D.; Wingo, C.S. The Renal H+-K+-ATPases: Physiology, Regulation, and Structure. Am. J. Physiol. Ren. Physiol. 2010, 298, F12–F21. [Google Scholar] [CrossRef] [Green Version]
- Johansson, M.; Jansson, T.; Pestov, N.B.; Powell, T.L. Non-Gastric H+/K+ ATPase Is Present in the Microvillous Membrane of the Human Placental Syncytiotrophoblast. Placenta 2004, 25, 505–511. [Google Scholar] [CrossRef]
- Shah, V.S.; Meyerholz, D.K.; Tang, X.X.; Reznikov, L.; Abou Alaiwa, M.; Ernst, S.E.; Karp, P.H.; Wohlford-Lenane, C.L.; Heilmann, K.P.; Leidinger, M.R.; et al. Airway Acidification Initiates Host Defense Abnormalities in Cystic Fibrosis Mice. Science 2016, 351, 503–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorrieri, G.; Scudieri, P.; Caci, E.; Schiavon, M.; Tomati, V.; Sirci, F.; Napolitano, F.; Carrella, D.; Gianotti, A.; Musante, I.; et al. Goblet Cell Hyperplasia Requires High Bicarbonate Transport To Support Mucin Release. Sci. Rep. 2016, 6, 36016. [Google Scholar] [CrossRef] [Green Version]
- Wandall, J.H. Effects of Omeprazole on Neutrophil Chemotaxis, Super Oxide Production, Degranulation, and Translocation of Cytochrome b-245. Gut 1992, 33, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Ritter, M.; Schratzberger, P.; Rossmann, H.; Wöll, E.; Seiler, K.; Seidler, U.; Reinisch, N.; Kähler, C.M.; Zwierzina, H.; Lang, H.J.; et al. Effect of Inhibitors of Na+/H+-Exchange and Gastric H+/K+ ATPase on Cell Volume, Intracellular PH and Migration of Human Polymorphonuclear Leucocytes. Br. J. Pharmacol. 1998, 124, 627–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins de Oliveira, R.; Antunes, E.; Pedrazzoli, J.J.; Gambero, A. The Inhibitory Effects of H+ K+ ATPase Inhibitors on Human Neutrophils in Vitro: Restoration by a K+ Ionophore. Inflamm. Res. 2007, 56, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Barbuskaite, D.; Tozzi, M.; Giannuzzo, A.; Sørensen, C.E.; Novak, I. Proton Pump Inhibitors Inhibit Pancreatic Secretion: Role of Gastric and Non-Gastric H+/K+-ATPases. PLoS ONE 2015, 10, e0126432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, I.; Wang, J.; Henriksen, K.L.; Haanes, K.A.; Krabbe, S.; Nitschke, R.; Hede, S.E. Pancreatic Bicarbonate Secretion Involves Two Proton Pumps. J. Biol. Chem. 2011, 286, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Yeo, M.; Kim, D.-K.; Kim, Y.-B.; Oh, T.Y.; Lee, J.-E.; Cho, S.W.; Kim, H.C.; Hahm, K.-B. Selective Induction of Apoptosis with Proton Pump Inhibitor in Gastric Cancer Cells. Clin. Cancer Res. 2004, 10, 8687–8696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakab, M.; Hofer, S.; Ravasio, A.; Huber, F.; Schmidt, S.; Hitzl, W.; Geibel, J.P.; Fürst, J.; Ritter, M. The Putative Role of the Non-Gastric H+/K+-ATPase ATP12A (ATP1AL1) as Anti-Apoptotic Ion Transporter: Effect of the H+/K+ATPase Inhibitor SCH28080 on Butyrate-Stimulated Myelomonocytic HL-60 Cells. Cell. Physiol. Biochem. 2014, 34, 1507–1526. [Google Scholar] [CrossRef] [PubMed]
- Jakab, M.; Ketterl, N.; Fürst, J.; Beyreis, M.; Kittl, M.; Kiesslich, T.; Hauser-Kronberger, C.; Gaisberger, M.; Ritter, M. The H+/K+ ATPase Inhibitor SCH-28080 Inhibits Insulin Secretion and Induces Cell Death in INS-1E Rat Insulinoma Cells. Cell. Physiol. Biochem. 2017, 43, 1037–1051. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Sakai, H.; Kuragari, M.; Suzuki, T.; Tauchi, K.; Minamimura, T.; Tsukada, K.; Asano, S.; Takeguchi, N. Expression of ATP1AL1, a Non-Gastric Proton Pump, in Human Colorectum. Jpn. J. Physiol. 2002, 52, 317–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streif, D.; Iglseder, E.; Hauser-Kronberger, C.; Fink, K.G.; Jakab, M.; Ritter, M. Expression of the Non-gastric H+/K+ ATPase ATP12A in Normal and Pathological Human Prostate Tissue. Cell. Physiol. Biochem. 2011, 28, 1287–1294. [Google Scholar] [CrossRef]
- Huttlin, E.L.; Jedrychowski, M.P.; Elias, J.E.; Goswami, T.; Rad, R.; Beausoleil, S.A.; Villen, J.; Haas, W.; Sowa, M.E.; Gygi, S.P. A Tissue-Specific Atlas of Mouse Protein Phosphorylation and Expression. Cell 2010, 143, 1174–1189. [Google Scholar] [CrossRef] [Green Version]
- Meierhofer, D.; Halbach, M.; Sen, N.E.; Gispert, S.; Auburger, G. Ataxin-2 (Atxn2)-Knock-Out Mice Show Branched Chain Amino Acids and Fatty Acids Pathway Alterations. Mol. Cell. Proteom. 2016, 15, 1728–1739. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Guo, Y.; Zhou, T.; Shi, X.; Yu, J.; Yang, Y.; Wu, Y.; Wang, J.; Liu, M.; Chen, X.; et al. In-depth proteomic analysis of the human sperm reveals complex protein compositions. J. Proteom. 2013, 79, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Rajamanickam, G.D.; Kastelic, J.; Thundathil, J.C. Na/K-ATPase regulates bovine sperm capacitation through raft- and non-raft-mediated signaling mechanisms. Mol. Reprod. Dev. 2017, 84, 1168–1182. [Google Scholar] [CrossRef]
- Rajamanickam, G.D.; Kastelic, J.P.; Thundathil, J.C. Testis-Specific Isoform of Na/K-ATPase (ATP1A4) Interactome in Raft and Non-Raft Membrane Fractions from Capacitated Bovine Sperm. Int. J. Mol. Sci. 2019, 20, 3159. [Google Scholar] [CrossRef] [Green Version]
- Lestari, S.; Miati, D.N.; Seoharso, P.; Sugiyanto, R.; Pujianto, D.A. Sperm Na+, K+-ATPase α4 and plasma membrane Ca2+-ATPase (PMCA) 4 regulation in asthenozoospermia. Syst. Biol. Reprod. Med. 2017, 63, 294–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, T.; Sánchez, G.; Blanco, G. Activity of the Na,K-ATPase 4 Isoform Is Regulated During Sperm Capacitation to Support Sperm Motility. J. Androl. 2012, 33, 1047–1057. [Google Scholar] [CrossRef]
- Newton, L.D.; Krishnakumar, S.; Menon, A.G.; Kastelic, J.P.; van der Hoorn, F.A.; Thundathil, J.C. Na+/K+ATPase regulates sperm capacitation through a mechanism involving kinases and redistribution of its testis-specific isoform. Mol. Reprod. Dev. 2010, 77, 136–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hlivko, J.T.; Chakraborty, S.; Hlivko, T.J.; Sengupta, A.; James, P.F. The human Na,K-ATPase alpha4 isoform is a ouabain-sensitive alpha isoform that is expressed in sperm. Mol. Reprod. Dev. 2006, 73, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, G.; Nguyen, A.-N.T.; Timmerberg, B.; Tash, J.S.; Blanco, G. The Na,K-ATPase α4 isoform from humans has distinct enzymatic properties and is important for sperm motility. Mol. Hum. Reprod. 2006, 12, 565–576. [Google Scholar] [CrossRef]
- Wagoner, K.; Sánchez, G.; Nguyen, A.-N.; Enders, G.C.; Blanco, G. Different expression and activity of the α1 and α4 isoforms of the Na,K-ATPase during rat male germ cell ontogeny. Reproduction 2005, 130, 627–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koçak-Toker, N.; Aktan, G.; Aykaç-Toker, G. The role of Na,K-ATPase in human sperm motility. Int. J. Androl. 2002, 25, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Avila, J.; Cozar-Castellano, I.; Brownleader, M.D.; Trevan, M.; Francis, M.J.; Lamb, J.F.; Martín-Vasallo, P. Na+, K+-ATPase Isozyme Diversity; Comparative Biochemistry and Physiological Implications of Novel Functional Interactions. Biosci. Rep. 2000, 20, 51–91. [Google Scholar] [CrossRef]
- Woo, A.L.; James, P.F.; Lingrel, J.B. Sperm Motility Is Dependent on a Unique Isoform of the Na,K-ATPase. J. Biol. Chem. 2000, 275, 20693–20699. [Google Scholar] [CrossRef] [Green Version]
- Thundathil, J.C.; Rajamanickam, G.D.; Kastelic, J.P. Na/K-ATPase and regulation of sperm function. In Proceedings of the 10th International Ruminant Reproduction Symposium (IRRS 2018), Foz do Iguaçu, PR, Brazil, 16–20 September 2018. [Google Scholar] [CrossRef]
- McDermott, J.P.; Numata, S.; Blanco, G. Na,K-ATPase Atp1a4 isoform is important for maintaining sperm flagellar shape. J. Assist. Reprod. Genet. 2021, 38, 1493–1505. [Google Scholar] [CrossRef]
- McDermott, J.P.; Sánchez, G.; Mitra, A.; Numata, S.; Liu, L.C.; Blanco, G. Na,K-ATPase α4, and Not Na,K-ATPase α1, is the Main Contributor to Sperm Motility, But its High Ouabain Binding Affinity Site is Not Required for Male Fertility in Mice. J. Membr. Biol. 2021, 254, 549–561. [Google Scholar] [CrossRef]
- Syeda, S.S.; Sánchez, G.; McDermott, J.P.; Hong, K.H.; Blanco, G.; Georg, G. The Na+ and K+ transport system of sperm (ATP1A4) is essential for male fertility and an attractive target for male contraception†. Biol. Reprod. 2020, 103, 343–356. [Google Scholar] [CrossRef]
- Thundathil, J.C.; Anzar, M.; Buhr, M.M. Na+/K+ATPase as a Signaling Molecule During Bovine Sperm Capacitation1. Biol. Reprod. 2006, 75, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Hickey, K.D.; Buhr, M.M. Characterization of Na+K+-ATPase in bovine sperm. Theriogenology 2012, 77, 1369–1380. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef]
- Bublin, M.; Breiteneder, H. Cross-reactivities of non-homologous allergens. Allergy 2019, 75, 1019–1022. [Google Scholar] [CrossRef]
- Lennox, A.T.; Coburn, S.L.; Leech, J.A.; Heidrich, E.M.; Kleyman, T.R.; Wenzel, S.E.; Pilewski, J.M.; Corcoran, T.E.; Myerburg, M.M. ATP12A promotes mucus dysfunction during Type 2 airway inflammation. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Min, J.-Y.; Ocampo, C.J.; Stevens, W.W.; Price, C.P.; Thompson, C.F.; Homma, T.; Huang, J.H.; Norton, J.E.; Suh, L.A.; Pothoven, K.L.; et al. Proton pump inhibitors decrease eotaxin-3/CCL26 expression in patients with chronic rhinosinusitis with nasal polyps: Possible role of the nongastric H,K-ATPase. J. Allergy Clin. Immunol. 2017, 139, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Swarts, H.G.P.; Koenderink, J.B.; Willems, P.H.G.M.; De Pont, J.J.H.H.M. The Human Non-Gastric H,K-ATPase Has a Different Cation Specificity than the Rat Enzyme. Biochim. Biophys. Acta 2007, 1768, 580–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pestov, N.B.; Korneenko, T.V.; Adams, G.; Tillekeratne, M.; Shakhparonov, M.I.; Modyanov, N.N. Nongastric H-K-ATPase in Rodent Prostate: Lobe-Specific Expression and Apical Localization. Am. J. Physiol. Cell Physiol. 2002, 282, C907–C916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, D.; Curtis, E.F.; Miller, R.G. Specific Labelling by Peanut Agglutinin of the Outer Acrosomal Membrane of the Human Spermatozoon. J. Reprod. Fertil. 1987, 81, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Grishin, A.V.; Sverdlov, V.E.; Kostina, M.B.; Modyanov, N.N. Cloning and Characterization of the Entire CDNA Encoded by ATP1AL1--a Member of the Human Na,K/H,K-ATPase Gene Family. FEBS Lett. 1994, 349, 144–150. [Google Scholar] [CrossRef] [Green Version]
- Bublitz, M.; Morth, J.P.; Nissen, P. P-Type ATPases at a Glance. J. Cell Sci. 2011, 124, 2515–2519. [Google Scholar] [CrossRef] [Green Version]
- Blanco, G.; Sánchez, G.; Melton, R.J.; Tourtellotte, W.G.; Mercer, R.W. The Alpha4 Isoform of the Na,K-ATPase Is Expressed in the Germ Cells of the Testes. J. Histochem. Cytochem. 2000, 48, 1023–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, T.; Sánchez, G.; Wertheimer, E.; Blanco, G. Activity of the Na,K-ATPase Alpha4 Isoform Is Important for Membrane Potential, Intracellular Ca2+, and PH to Maintain Motility in Rat Spermatozoa. Reproduction 2010, 139, 835–845. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, T.; McDermott, J.P.; Sánchez, G.; Blanco, G. Na,K-ATPase Alpha4 Isoform Is Essential for Sperm Fertility. Proc. Natl. Acad. Sci. USA 2011, 108, 644–649. [Google Scholar] [CrossRef] [Green Version]
- McDermott, J.; Sánchez, G.; Nangia, A.K.; Blanco, G. Role of Human Na,K-ATPase Alpha 4 in Sperm Function, Derived from Studies in Transgenic Mice. Mol. Reprod. Dev. 2015, 82, 167–181. [Google Scholar] [CrossRef] [Green Version]
- Clausen, M.V.; Hilbers, F.; Poulsen, H. The Structure and Function of the Na,K-ATPase Isoforms in Health and Disease. Front. Physiol. 2017, 8, 371. [Google Scholar] [CrossRef]
- Jimenez, T.; Sanchez, G.; McDermott, J.P.; Nguyen, A.-N.; Kumar, T.R.; Blanco, G. Increased Expression of the Na,K-ATPase Alpha4 Isoform Enhances Sperm Motility in Transgenic Mice. Biol. Reprod. 2011, 84, 153–161. [Google Scholar] [CrossRef] [Green Version]
- Peralta-Arias, R.D.; Vívenes, C.Y.; Camejo, M.I.; Piñero, S.; Proverbio, T.; Martínez, E.; Marín, R.; Proverbio, F. ATPases, Ion Exchangers and Human Sperm Motility. Reproduction 2015, 149, 475–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Shafeeque, C.M.; Sharma, S.K.; Singh, R.; Mohan, J.; Sastry, K.V.H.; Saxena, V.K.; Azeez, P.A. Chicken Sperm Transcriptome Profiling by Microarray Analysis. Genome 2016, 59, 185–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, A.; Tang, C.; Xie, Y.; Ortogero, N.; Yuan, S.; Yan, W. SpermBase: A Database for Sperm-Borne RNA Contents. Biol. Reprod. 2016, 95, 99. [Google Scholar] [CrossRef] [Green Version]
- Escoffier, J.; Arnaud, B.; Kaba, M.; Hograindleur, J.P.; Le Blévec, E.; Martinez, G.; Stévant, I.; Ray, P.F.; Arnoult, C.; Nef, S. Pantoprazole, a Proton-Pump Inhibitor, Impairs Human Sperm Motility and Capacitation In Vitro. Andrology 2020, 8, 1795–1804. [Google Scholar] [CrossRef]
- Li, J.; Codina, J.; Petroske, E.; Werle, M.J.; Willingham, M.C.; DuBose, T.D.J. The Effect of Beta-Subunit Assembly on Function and Localization of the Colonic H+,K+-ATPase Alpha-Subunit. Kidney Int. 2004, 66, 1068–1075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef]
- Aldana, A.; Carneiro, J.; Martínez-Mekler, G.; Darszon, A. Discrete Dynamic Model of the Mammalian Sperm Acrosome Reaction: The Influence of Acrosomal PH and Physiological Heterogeneity. Front. Physiol. 2021, 12, 682790. [Google Scholar] [CrossRef]
- Nakamura, N.; Tanaka, S.; Teko, Y.; Mitsui, K.; Kanazawa, H. Four Na+/H+ Exchanger Isoforms Are Distributed to Golgi and Post-Golgi Compartments and Are Involved in Organelle PH Regulation. J. Biol. Chem. 2005, 280, 1561–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oberheide, K.; Puchkov, D.; Jentsch, T.J. Loss of the Na(+)/H(+) Exchanger NHE8 Causes Male Infertility in Mice by Disrupting Acrosome Formation. J. Biol. Chem. 2017, 292, 10845–10854. [Google Scholar] [CrossRef] [Green Version]
- Chávez, J.C.; De la Vega-Beltrán, J.L.; José, O.; Torres, P.; Nishigaki, T.; Treviño, C.L.; Darszon, A. Acrosomal Alkalization Triggers Ca(2+) Release and Acrosome Reaction in Mammalian Spermatozoa. J. Cell Physiol. 2018, 233, 4735–4747. [Google Scholar] [CrossRef]
- Nakanishi, T.; Ikawa, M.; Yamada, S.; Toshimori, K.; Okabe, M. Alkalinization of Acrosome Measured by GFP as a PH Indicator and Its Relation to Sperm Capacitation. Dev. Biol. 2001, 237, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Koh, J.-Y. Roles for H(+) /K(+) -ATPase and Zinc Transporter 3 in CAMP-Mediated Lysosomal Acidification in Bafilomycin A1-Treated Astrocytes. Glia 2021, 69, 1110–1125. [Google Scholar] [CrossRef] [PubMed]
- Tragni, V.; Cotugno, P.; De Grassi, A.; Cavalluzzi, M.M.; Mincuzzi, A.; Lentini, G.; Sanzani, S.M.; Ippolito, A.; Pierri, C.L. Targeting Penicillium Expansum GMC Oxidoreductase with High Affinity Small Molecules for Reducing Patulin Production. Biology 2020, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Todisco, S.; Di Noia, M.A.; Onofrio, A.; Parisi, G.; Punzi, G.; Redavid, G.; De Grassi, A.; Pierri, C.L. Identification of New Highly Selective Inhibitors of the Human ADP/ATP Carriers by Molecular Docking and in Vitro Transport Assays. Biochem. Pharmacol. 2016, 100, 112–132. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.F.; Alfaro, V.; Montenegro, V.; Rodriguez-Martinez, H. Computer-Assisted Analysis of Sperm Motion in Goats and Its Relationship with Sperm Migration in Cervical Mucus. Theriogenology 2006, 66, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Muzzachi, S.; Guerra, L.; Martino, N.A.; Favia, M.; Punzi, G.; Silvestre, F.; Guaricci, A.C.; Roscino, M.T.; Pierri, C.L.; Dell’Aquila, M.E.; et al. Effect of Cariporide on Ram Sperm PH Regulation and Motility: Possible Role of NHE1. Reproduction 2018, 155, 433–445. [Google Scholar] [CrossRef]
- Hasan, M.M.; Reshi, Q.U.A.; Lättekivi, F.; Viil, J.; Godakumara, K.; Dissanayake, K.; Andronowska, A.; Jaakma, Ü.; Fazeli, A. Bovine Follicular Fluid Derived Extracellular Vesicles Modulate the Viability, Capacitation and Acrosome Reaction of Bull Spermatozoa. Biology 2021, 10, 1154. [Google Scholar] [CrossRef] [PubMed]
- Meyer, D.; Voigt, A.; Widmayer, P.; Borth, H.; Huebner, S.; Breit, A.; Marschall, S.; de Angelis, M.H.; Boehm, U.; Meyerhof, W.; et al. Expression of Tas1 Taste Receptors in Mammalian Spermatozoa: Functional Role of Tas1r1 in Regulating Basal Ca2+ and CAMP Concentrations in Spermatozoa. PLoS ONE 2012, 7, e32354. [Google Scholar] [CrossRef] [Green Version]
- Bossis, F.; De Grassi, A.; Palese, L.L.; Pierri, C.L. Prediction of High- and Low-Affinity Quinol-Analogue-Binding Sites in the Aa3 and Bo3 Terminal Oxidases from Bacillus Subtilis and Escherichia Coli1. Biochem. J. 2014, 461, 305–314. [Google Scholar] [CrossRef]
- Trisolini, L.; Gambacorta, N.; Gorgoglione, R.; Montaruli, M.; Laera, L.; Colella, F.; Volpicella, M.; De Grassi, A.; Pierri, C.L. FAD/NADH Dependent Oxidoreductases: From Different Amino Acid Sequences to Similar Protein Shapes for Playing an Ancient Function. J. Clin. Med. 2019, 8, 2117. [Google Scholar] [CrossRef] [Green Version]
- Pietropaolo, A.; Pierri, C.L.; Palmieri, F.; Klingenberg, M. The Switching Mechanism of the Mitochondrial ADP/ATP Carrier Explored by Free-Energy Landscapes. Biochim. Biophys. Acta 2016, 1857, 772–781. [Google Scholar] [CrossRef]
- Pierri, C.L.; Parisi, G.; Porcelli, V. Computational Approaches for Protein Function Prediction: A Combined Strategy from Multiple Sequence Alignment to Molecular Docking-Based Virtual Screening. Biochim. Biophys. Acta 2010, 1804, 1695–1712. [Google Scholar] [CrossRef] [PubMed]
- Pierri, C.L.; Bossis, F.; Punzi, G.; De Grassi, A.; Cetrone, M.; Parisi, G.; Tricarico, D. Molecular Modeling of Antibodies for the Treatment of TNFα-Related Immunological Diseases. Pharmacol. Res. Perspect. 2016, 4, e00197. [Google Scholar] [CrossRef] [PubMed]
- Axelsen, K.B.; Palmgren, M.G. Evolution of Substrate Specificities in the P-Type ATPase Superfamily. J. Mol. Evol. 1998, 46, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Procter, J.B.; Carstairs, G.M.; Soares, B.; Mourão, K.; Ofoegbu, T.C.; Barton, D.; Lui, L.; Menard, A.; Sherstnev, N.; Roldan-Martinez, D.; et al. Alignment of Biological Sequences with Jalview. Methods Mol. Biol. 2021, 2231, 203–224. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Gregersen, J.L.; Mattle, D.; Fedosova, N.U.; Nissen, P.; Reinhard, L. Isolation, Crystallization and Crystal Structure Determination of Bovine Kidney Na(+),K(+)-ATPase. Acta Crystallogr. F Struct. Biol. Commun. 2016, 72, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Irie, K.; Nakanishi, H.; Suzuki, H.; Fujiyoshi, Y. Crystal Structures of the Gastric Proton Pump. Nature 2018, 556, 214–218. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Joo, K.; Lee, J.; Lee, J.; Raman, S.; Thompson, J.; Tyka, M.; Baker, D.; Karplus, K. Improving Physical Realism, Stereochemistry, and Side-Chain Accuracy in Homology Modeling: Four Approaches That Performed Well in CASP8. Proteins 2009, 77 (Suppl. 9), 114–122. [Google Scholar] [CrossRef] [Green Version]
- Van Durme, J.; Delgado, J.; Stricher, F.; Serrano, L.; Schymkowitz, J.; Rousseau, F. A Graphical Interface for the FoldX Forcefield. Bioinformatics 2011, 27, 1711–1712. [Google Scholar] [CrossRef] [Green Version]
- Schymkowitz, J.; Borg, J.; Stricher, F.; Nys, R.; Rousseau, F.; Serrano, L. The FoldX Web Server: An Online Force Field. Nucleic Acids Res. 2005, 33, W382–W388. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Complex | Subunit α | Subunit β |
|---|---|---|
| Na+/K+ ATPase | α1 (ATP1A1) | β1 (ATP1B1) |
| α2 (ATP1A2) | β2 (ATP1B2) | |
| α3 (ATP1A3) | β3 (ATP1B3) | |
| α4 (ATP1A4) | β4 (ATP1B4) | |
| “gastric” H+/K+ ATPase | α (ATP4A) | β (ATP4B) |
| “non gastric” H+/K+ ATPase | α (ATP12A) | β non-specific |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Favia, M.; Gerbino, A.; Notario, E.; Tragni, V.; Sgobba, M.N.; Dell’Aquila, M.E.; Pierri, C.L.; Guerra, L.; Ciani, E. The Non-Gastric H+/K+ ATPase (ATP12A) Is Expressed in Mammalian Spermatozoa. Int. J. Mol. Sci. 2022, 23, 1048. https://doi.org/10.3390/ijms23031048
Favia M, Gerbino A, Notario E, Tragni V, Sgobba MN, Dell’Aquila ME, Pierri CL, Guerra L, Ciani E. The Non-Gastric H+/K+ ATPase (ATP12A) Is Expressed in Mammalian Spermatozoa. International Journal of Molecular Sciences. 2022; 23(3):1048. https://doi.org/10.3390/ijms23031048
Chicago/Turabian StyleFavia, Maria, Andrea Gerbino, Elisabetta Notario, Vincenzo Tragni, Maria Noemi Sgobba, Maria Elena Dell’Aquila, Ciro Leonardo Pierri, Lorenzo Guerra, and Elena Ciani. 2022. "The Non-Gastric H+/K+ ATPase (ATP12A) Is Expressed in Mammalian Spermatozoa" International Journal of Molecular Sciences 23, no. 3: 1048. https://doi.org/10.3390/ijms23031048