
Regulation and Function of Laminin A5 during Mouse and Human Decidualization

 ,
,
Abstract
:1. Introduction
2. Results
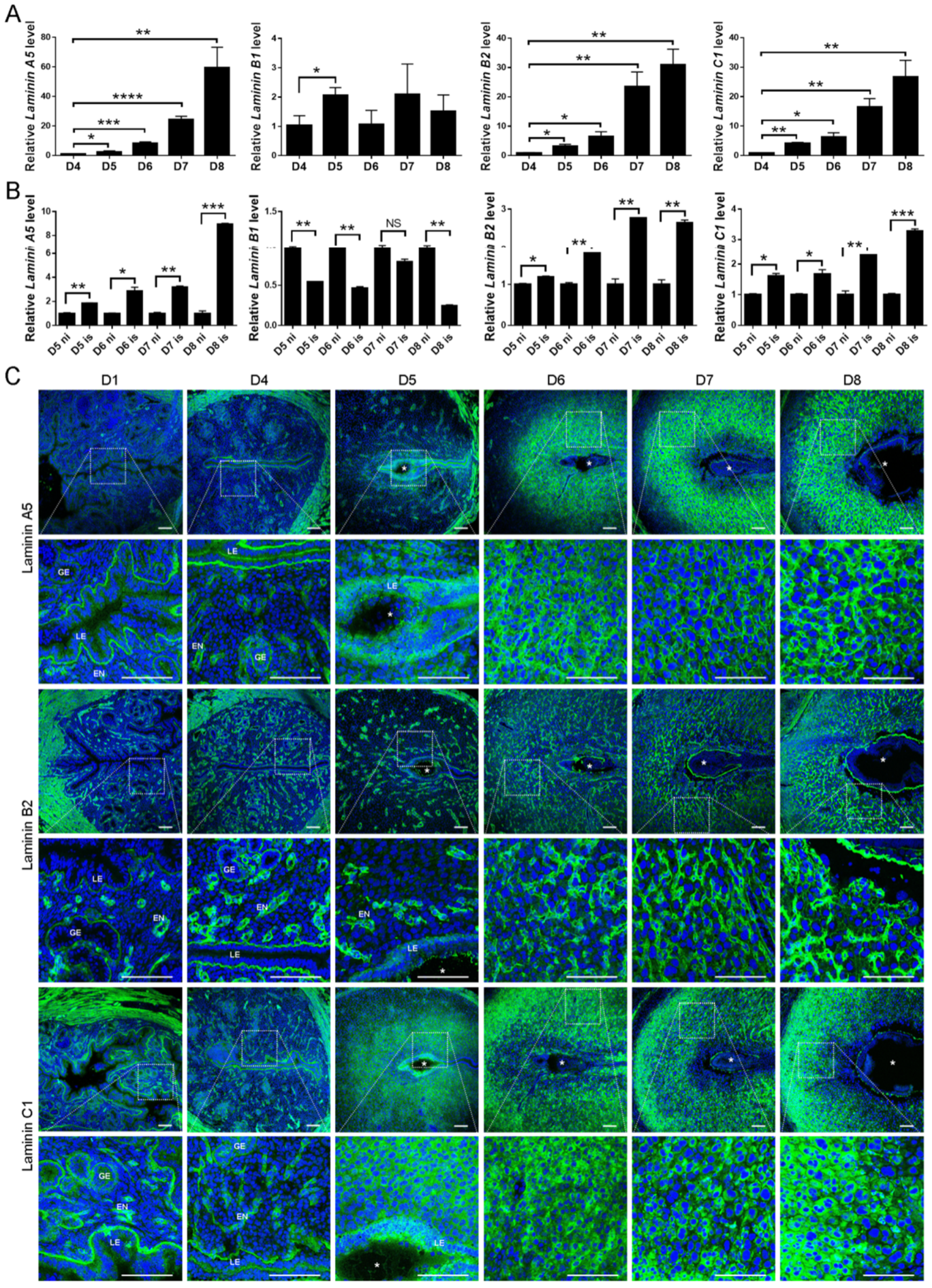
2.1. Spatiotemporal Expression of Laminin A5, B1, B2, and C1 in the Mouse Uteri during Early Pregnancy
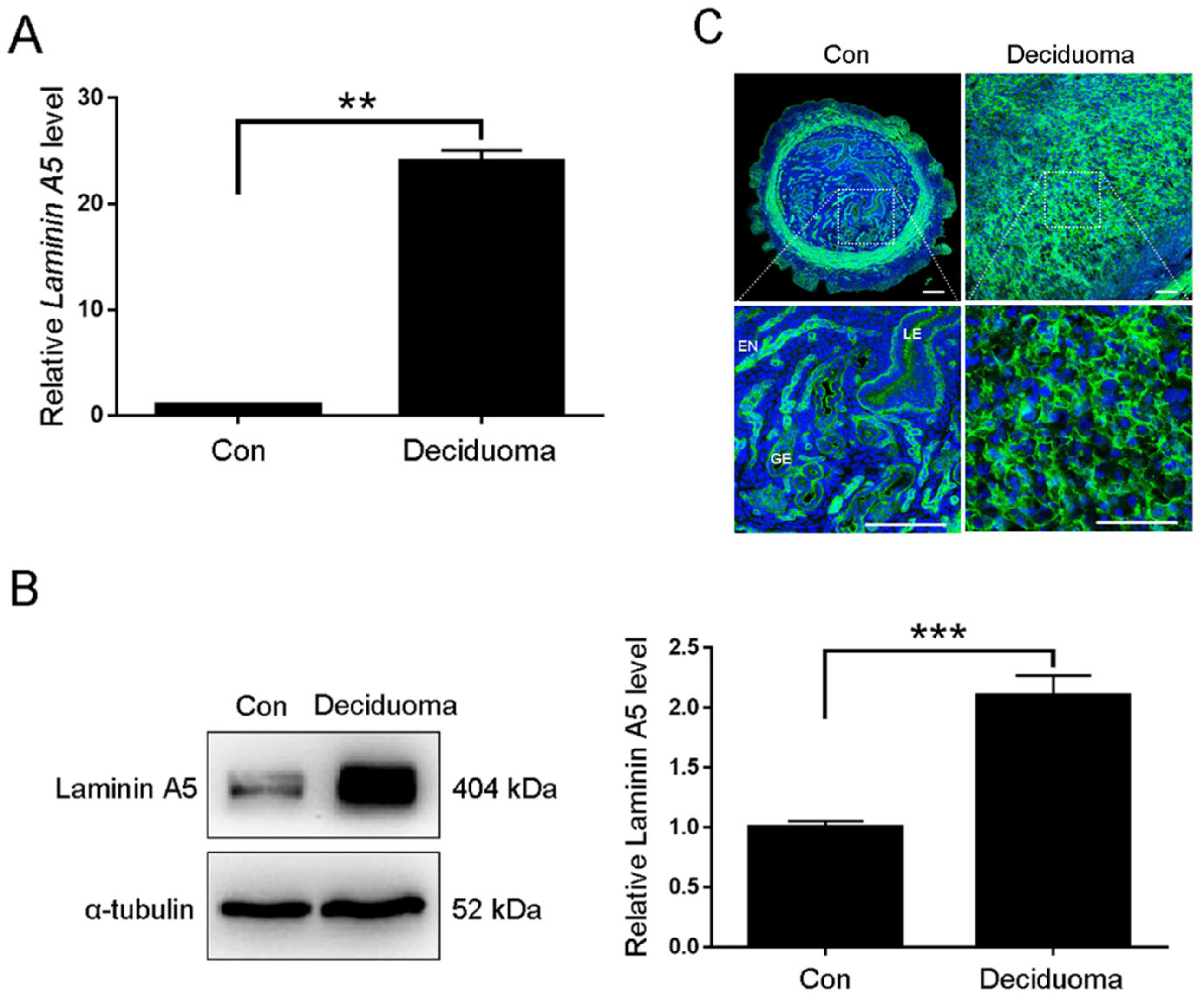
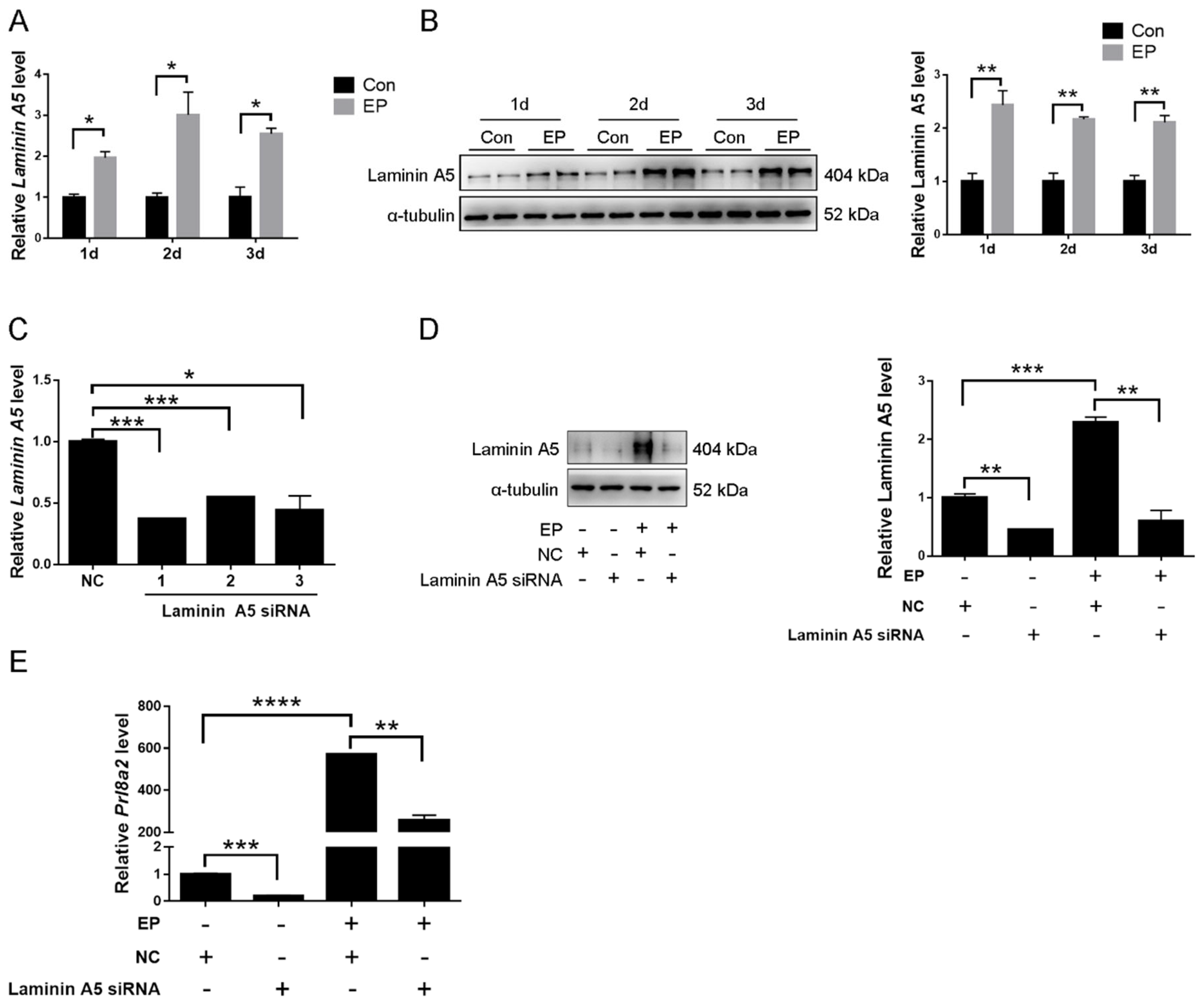
2.2. Effects of Laminin A5 on the Mouse In Vitro Decidualization
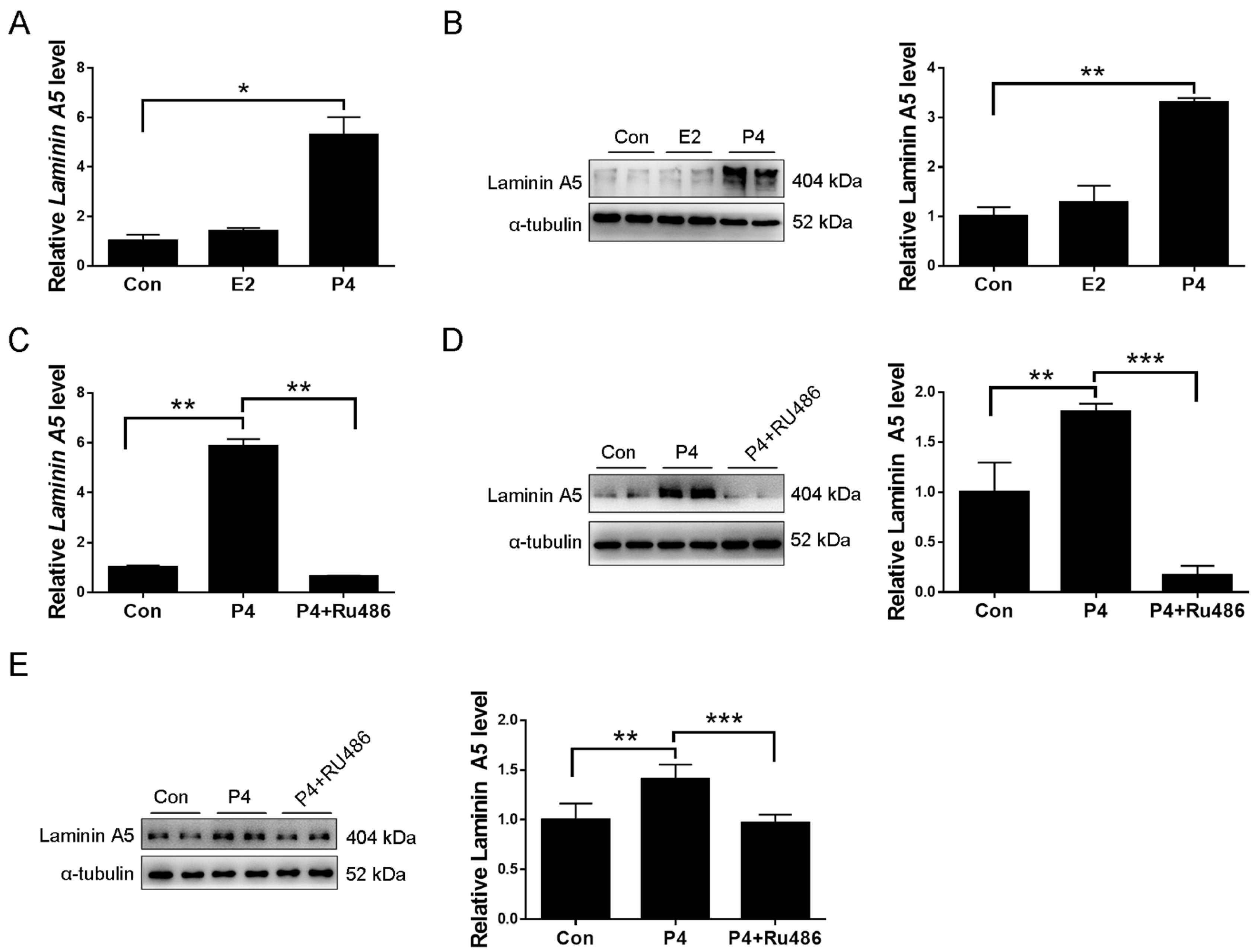
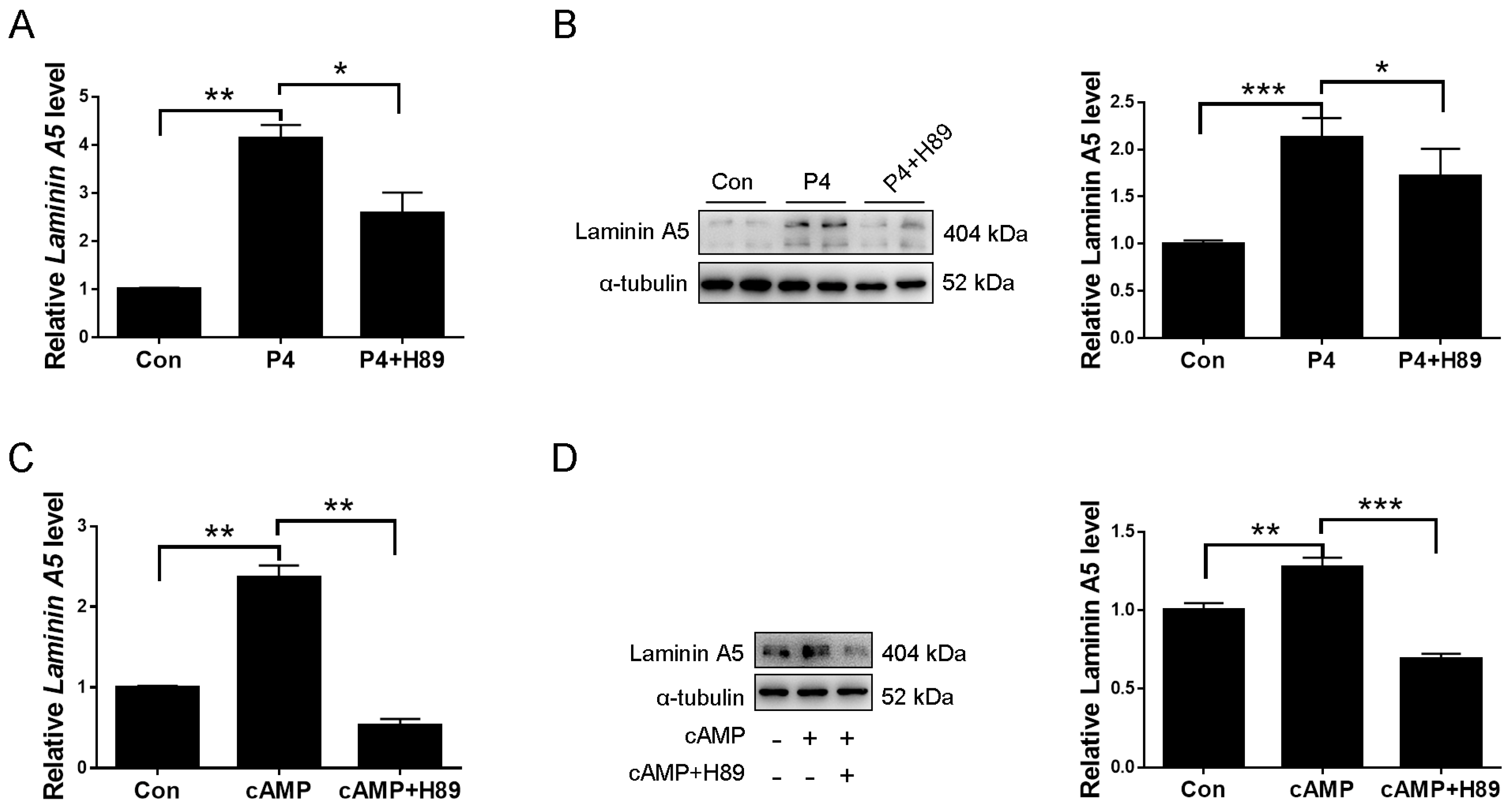
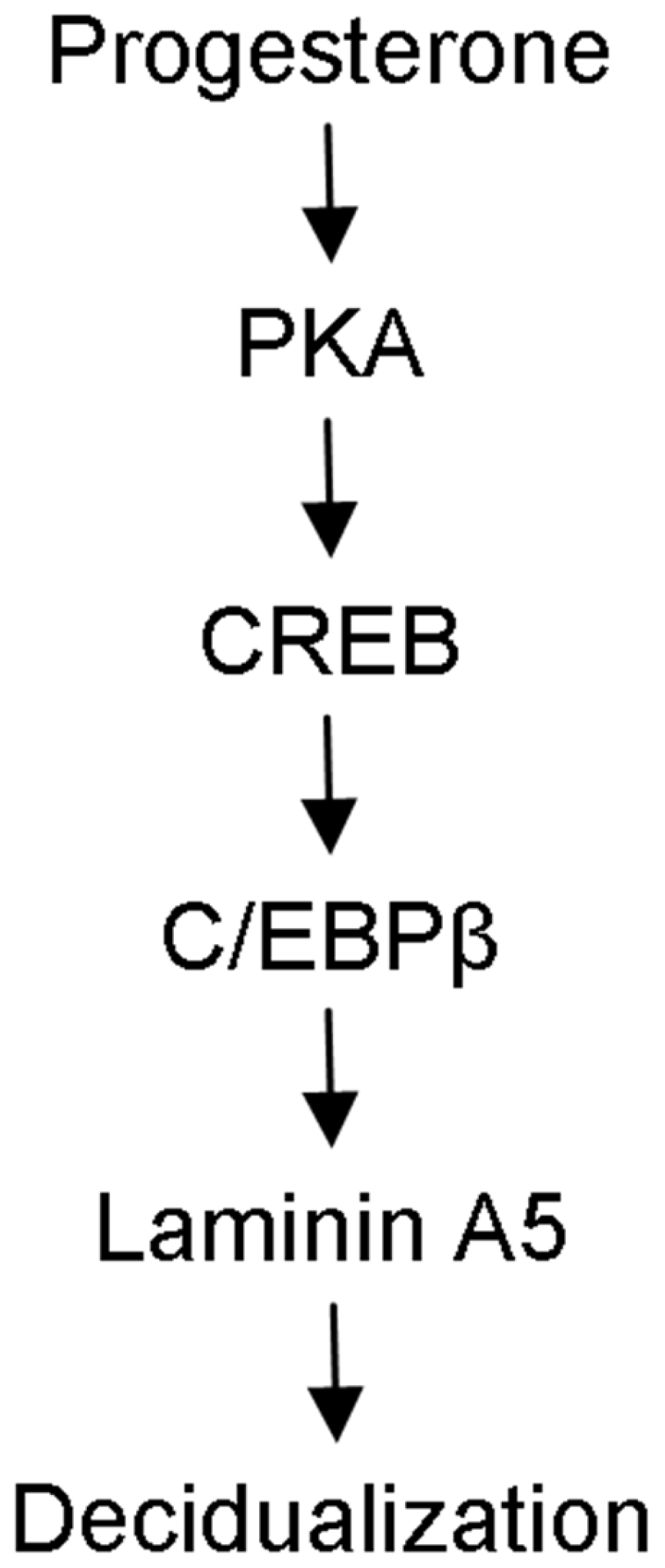
2.3. Laminin A5 Regulation by Progesterone
2.4. Progesterone Regulates Laminin A5 Expression through the cAMP-PKA Pathway
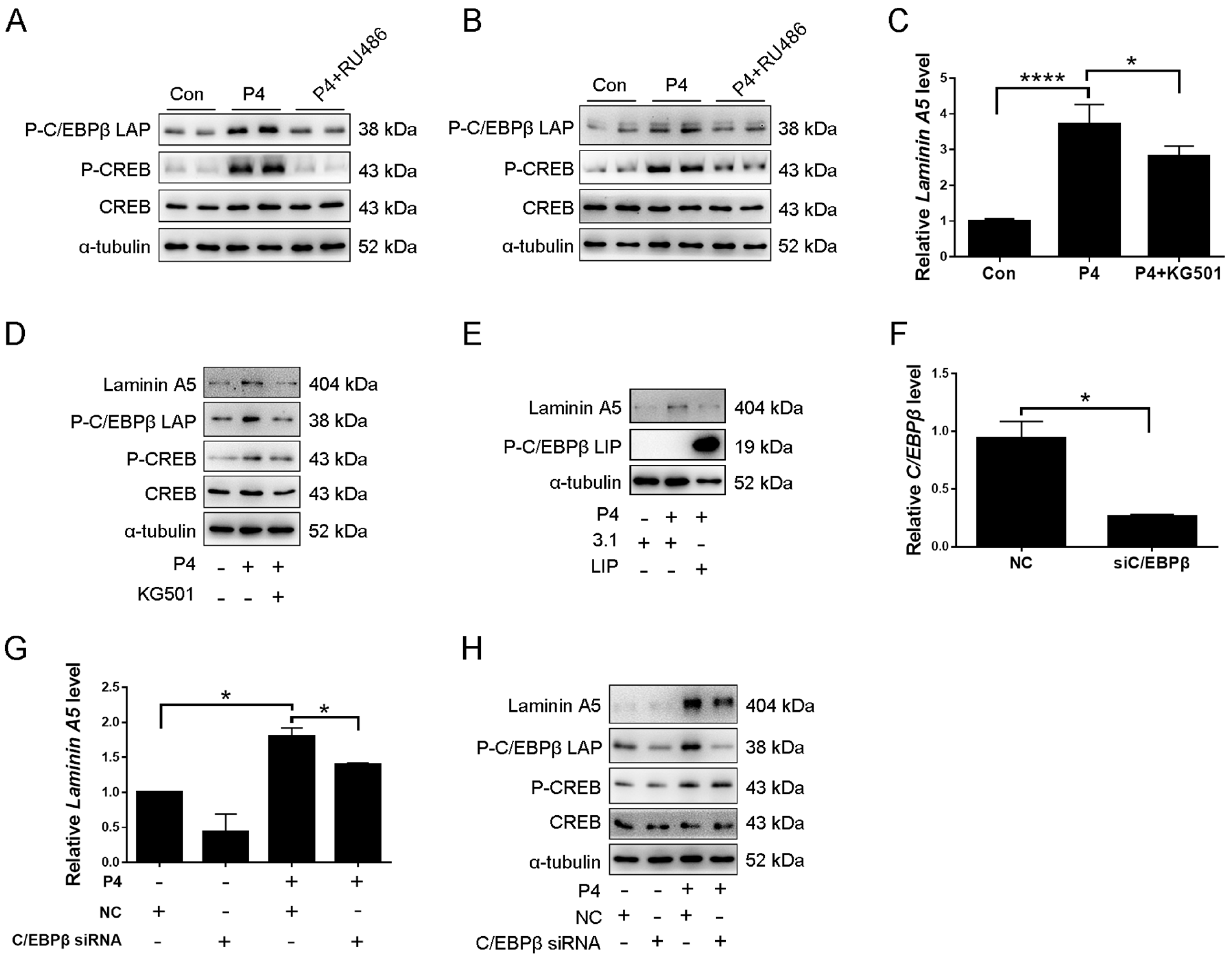
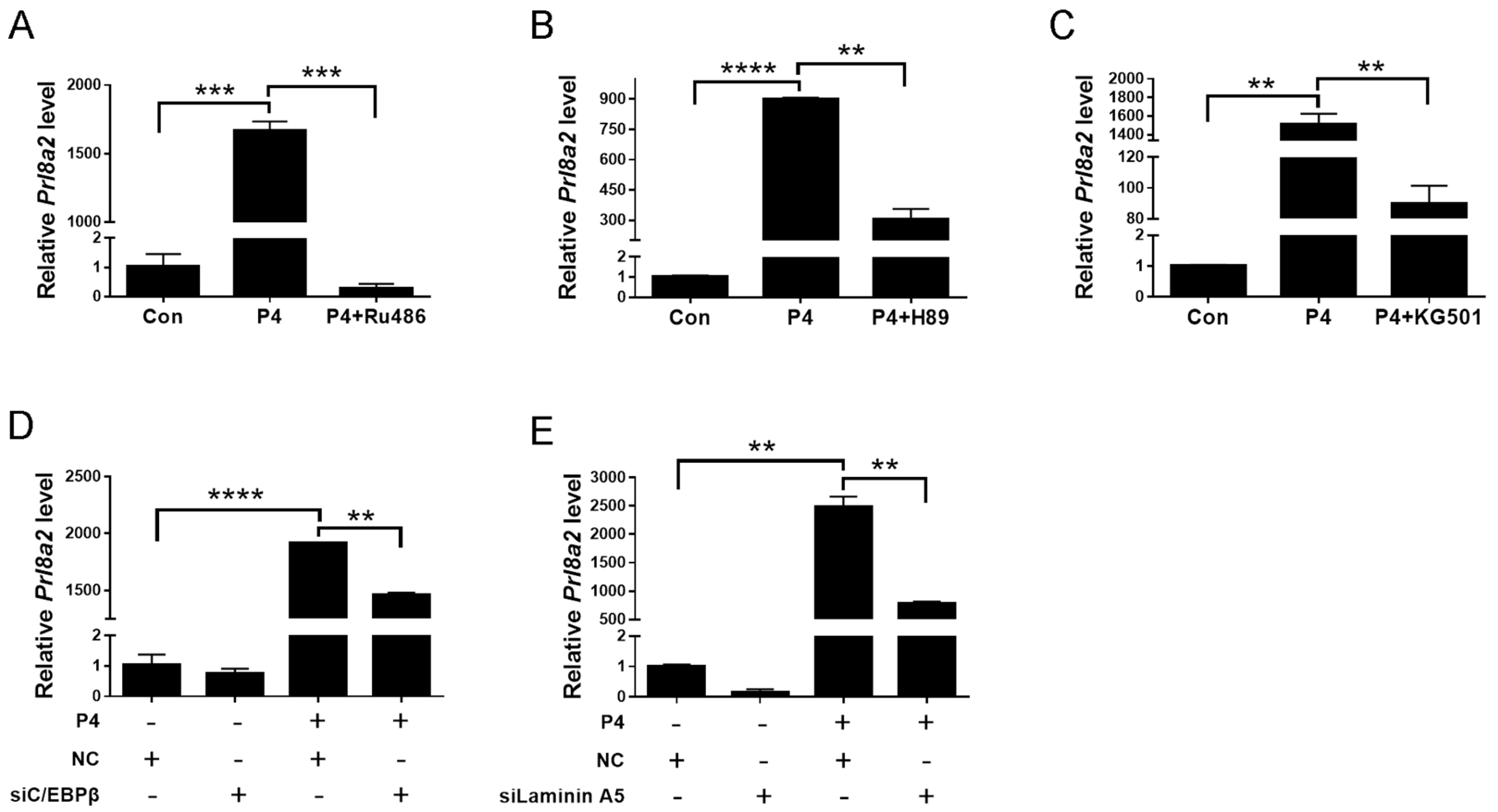
2.5. CREB-C/EBPβ Mediates the Stimulation of Progesterone on Laminin A5 Expression
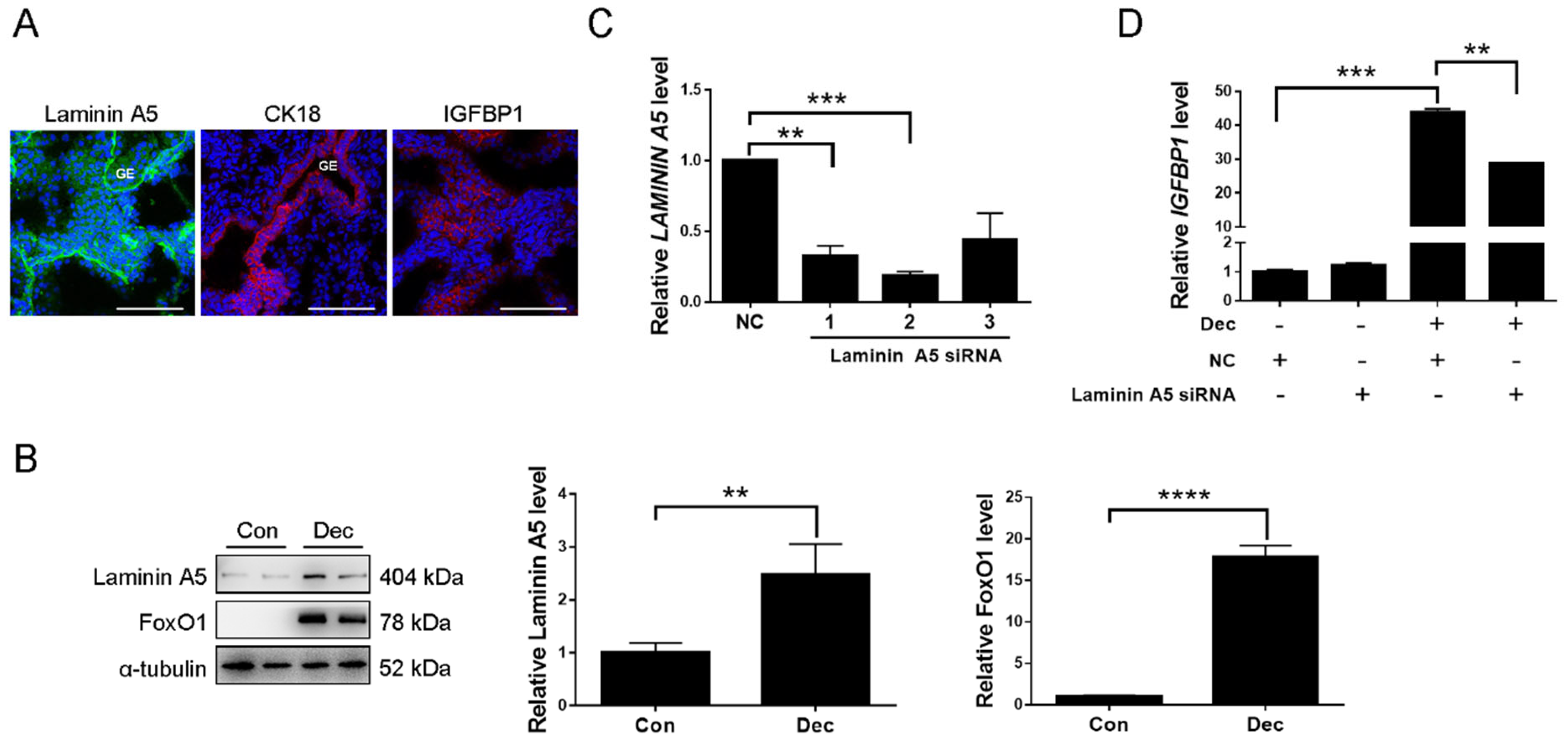
2.6. Expression and Function of Laminin A5 during Human Decidualization
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Collection of Human Decidual Samples
4.3. Isolation and Treatment of Mouse Endometrial Stromal Cells
4.4. Culture and Treatment of Human Endometrial Stromal Cells
4.5. siRNA Transfection
4.6. C/EBPβ LIP Overexpression
4.7. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.8. Immunofluorescence
4.9. Western Blot
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| C/EBPβ | CCAAT enhancer-binding protein β |
| CREB | cAMP-response element-binding protein |
| H89 | PKA inhibitor |
| IGFBP1 | Insulin growth factor-binding protein 1 |
| KG-501 | CREB inhibitor |
| PKA | Protein kinase A |
| Prl8a2 | Prolactin family 8, subfamily A, member 2 |
| RU486 | progesterone receptor antagonist |
References
- Wang, H.; Dey, S.K. Roadmap to embryo implantation: Clues from mouse models. Nat. Rev. Genet. 2006, 7, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Akyaw, A.; Krishnamoorthy, K.; Goldsmith, L.T.; Morelli, S.S. The role of mesenchymal-epithelial transition in endometrial function. Hum. Reprod. Update 2019, 25, 114–133. [Google Scholar] [CrossRef]
- Pawar, S.; Hantak, A.M.; Bagchi, I.C.; Bagchi, M.K. Minireview: Steroid-regulated paracrine mechanisms controlling implantation. Mol. Endocrinol. 2014, 28, 1408–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, H.; Tsuzuki, T.; Murata, H. Decidualization of the human endometrium. Reprod. Med. Biol. 2018, 17, 220–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sekiguchi, R.; Yamada, K.M. Basement membranes in development and disease. Curr. Top Dev. Biol. 2018, 130, 143–191. [Google Scholar] [CrossRef]
- Jayadev, R.; Sherwood, D.R. Basement membranes. Curr. Biol. 2017, 27, R207–R211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theocharis, A.D.; Skandalis, S.S.; Gialeli, C.; Karamanos, N.K. Extracellular matrix structure. Adv. Drug Deliv. Rev. 2016, 97, 4–27. [Google Scholar] [CrossRef]
- Schittny, J.C.; Yurchenco, P.D. Basement membranes: Molecular organization and function in development and disease. Curr. Opin. Cell Biol. 1989, 1, 983–988. [Google Scholar] [CrossRef]
- Paulsson, M. Basement membrane proteins: Structure, assembly, and cellular interactions. Crit. Rev. Biochem. Mol. Biol. 1992, 27, 93–127. [Google Scholar] [CrossRef]
- Timpl, R.; Rohde, H.; Robey, P.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin—A glycoprotein from basement membranes. J. Biol. Chem. 1979, 254, 9933–9937. [Google Scholar] [CrossRef]
- Chung, A.E.; Jaffe, R.; Freeman, I.L.; Vergnes, J.P.; Braginski, J.E.; Carlin, B. Properties of a basement membrane-related glycoprotein synthesized in culture by a mouse embryonal carcinoma-derived cell line. Cell 1979, 16, 277–287. [Google Scholar] [CrossRef]
- Timpl, R.; Dziadek, M.; Fujiwara, S.; Nowack, H.; Wick, G. Nidogen: A new, self-aggregating basement membrane protein. Eur. J. Biochem. 1983, 137, 455–465. [Google Scholar] [CrossRef] [PubMed]
- Hassell, J.R.; Robey, P.G.; Barrach, H.J.; Wilczek, J.; Rennard, S.I.; Martin, G.R. Isolation of a heparan sulfate-containing proteoglycan from basement membrane. Proc. Natl. Acad. Sci. USA 1980, 77, 4494–4498. [Google Scholar] [CrossRef] [Green Version]
- Mason, I.J.; Taylor, A.; Williams, J.G.; Sage, H.; Hogan, B.L. Evidence from molecular cloning that SPARC, a major product of mouse embryo parietal endoderm, is related to an endothelial cell ‘culture shock’ glycoprotein of Mr 43,000. EMBO J. 1986, 5, 1465–1472. [Google Scholar] [CrossRef] [PubMed]
- Tsen, G.; Halfter, W.; Kröger, S.; Cole, G.J. Agrin is a heparan sulfate proteoglycan. J. Biol. Chem. 1995, 270, 3392–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halfter, W.; Dong, S.; Schurer, B.; Cole, G.J. Collagen XVIII is a basement membrane heparan sulfate proteoglycan. J. Biol. Chem. 1998, 273, 25404–25412. [Google Scholar] [CrossRef] [Green Version]
- Timpl, R.; Sasaki, T.; Kostka, G.; Chu, M.L. Fibulins: A versatile family of extracellular matrix proteins. Nat. Rev. Mol. Cell Biol. 2003, 4, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Jones-Paris, C.R.; Paria, S.; Berg, T.; Saus, J.; Bhave, G.; Paria, B.C.; Hudson, B.G. Embryo implantation triggers dynamic spatiotemporal expression of the basement membrane toolkit during uterine reprogramming. Matrix Biol. 2017, 57–58, 347–365. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, A.; Feng, L.; Wang, Y.; Zhang, H.; Zhang, I.; Bany, B.M.; Ma, L. Heparan sulfate proteoglycan sulfation regulates uterine differentiation and signaling during embryo implantation. Endocrinology 2018, 159, 2459–2472. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, T.; Okada, H.; Nakajima, T.; Ikuta, A.; Yasuda, K.; Kanzaki, H. Progesterone induces the fibulin-1 expression in human endometrial stromal cells. Hum. Reprod. 2005, 20, 1447–1455. [Google Scholar] [CrossRef] [Green Version]
- Ramathal, C.; Wang, W.; Hunt, E.; Bagchi, I.C.; Bagchi, M.K. Transcription factor CCAAT enhancer-binding protein beta (C/EBPbeta) regulates the formation of a unique extracellular matrix that controls uterine stromal differentiation and embryo implantation. J. Biol. Chem. 2011, 286, 19860–19871. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.W.; Lai, Z.Z.; Yang, H.L.; Yang, S.L.; Wang, C.J.; Ao, D.; Ruan, L.Y.; Shen, H.H.; Zhou, W.J.; Mei, J.; et al. Collagen at the maternal-fetal interface in human pregnancy. Int. J. Biol. Sci. 2020, 16, 2220–2234. [Google Scholar] [CrossRef]
- Burgeson, R.E.; Chiquet, M.; Deutzmann, R.; Ekblom, P.; Engel, J.; Kleinman, H.; Martin, G.R.; Meneguzzi, G.; Paulsson, M.; Sanes, J.; et al. A new nomenclature for the laminins. Matrix Biol. 1994, 14, 209–211. [Google Scholar] [CrossRef]
- Yao, Y. Laminin: Loss-of-function studies. Cell Mol. Life Sci. 2017, 74, 1095–1115. [Google Scholar] [CrossRef]
- Hohenester, E. Structural biology of laminins. Essays Biochem. 2019, 63, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Yap, L.; Tay, H.G.; Nguyen, M.; Tjin, M.S.; Tryggvason, K. Laminins in cellular differentiation. Trends Cell. Biol. 2019, 29, 987–1000. [Google Scholar] [CrossRef] [Green Version]
- Aumailley, M. The laminin family. Cell. Adh. Migr. 2013, 7, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miner, J.H.; Patton, B.L.; Lentz, S.I.; Gilbert, D.J.; Snider, W.D.; Jenkins, N.A.; Copeland, N.G.; Sanes, J.R. The laminin alpha chains: Expression, developmental transitions, and chromosomal locations of alpha1-5, identification of heterotrimeric laminins 8–11, and cloning of a novel alpha3 isoform. J. Cell. Biol. 1997, 137, 685–701. [Google Scholar] [CrossRef]
- Miner, J.H.; Cunningham, J.; Sanes, J.R. Roles for laminin in embryogenesis: Exencephaly, syndactyly, and placentopathy in mice lacking the laminin alpha5 chain. J. Cell. Biol. 1998, 143, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Noakes, P.G.; Gautam, M.; Mudd, J.; Sanes, J.R.; Merlie, J.P. Aberrant differentiation of neuromuscular junctions in mice lacking s-laminin/laminin beta 2. Nature 1995, 374, 258–262. [Google Scholar] [CrossRef]
- Smyth, N.; Vatansever, H.S.; Murray, P.; Meyer, M.; Frie, C.; Paulsson, M.; Edgar, D. Absence of basement membranes after targeting the LAMC1 gene results in embryonic lethality due to failure of endoderm differentiation. J. Cell. Biol. 1999, 144, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.; Edgar, D. Regulation of programmed cell death by basement membranes in embryonic development. J. Cell. Biol. 2000, 150, 1215–1221. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Soares, M.J.; Srivastava, R.K.; Gibori, G. Expression of decidual prolactin-related protein in the rat decidua. Endocrinology 1994, 135, 1422–1427. [Google Scholar] [CrossRef]
- Dey, S.K.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.C.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev. 2004, 25, 341–373. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, H.; Kong, S.; Wang, S.; Wang, H.; Wang, H.; Armant, D.R. Physiological and molecular determinants of embryo implantation. Mol. Aspects Med. 2013, 34, 939–980. [Google Scholar] [CrossRef] [Green Version]
- Gellersen, B.; Brosens, J. Cyclic AMP and progesterone receptor cross-talk in human endometrium: A decidualizing affair. J. Endocrinol. 2003, 178, 357–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.F.; Tao, R.; Yang, Z.Q.; Wang, K.; Yue, Z.P.; Guo, B. Ptn functions downstream of C/EBPβ to mediate the effects of cAMP on uterine stromal cell differentiation through targeting Hand2 in response to progesterone. J. Cell. Physiol. 2018, 233, 1612–1626. [Google Scholar] [CrossRef]
- Deng, W.B.; Liang, X.H.; Liu, J.L.; Yang, Z.M. Regulation and function of deiodinases during decidualization in female mice. Endocrinology 2014, 155, 2704–2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Wang, T.S.; Zhao, Y.C.; Zuo, R.J.; Deng, W.B.; Chi, Y.J.; Yang, Z.M. Cyclic adenosine monophosphate-induced argininosuccinate synthase 1 expression is essential during mouse decidualization. Mol. Cell. Endocrinol. 2014, 388, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Ruffell, D.; Mourkioti, F.; Gambardella, A.; Kirstetter, P.; Lopez, R.G.; Rosenthal, N.; Nerlov, C. A CREB-C/EBPbeta cascade induces M2 macrophage-specific gene expression and promotes muscle injury repair. Proc. Natl. Acad. Sci. USA 2009, 106, 17475–17480. [Google Scholar] [CrossRef] [Green Version]
- Al-Sabbagh, M.; Fusi, L.; Higham, J.; Lee, Y.; Lei, K.; Hanyaloglu, A.C.; Lam, E.W.; Christian, M.; Brosens, J.J. NADPH oxidase-derived reactive oxygen species mediate decidualization of human endometrial stromal cells in response to cyclic AMP signaling. Endocrinology 2011, 152, 730–740. [Google Scholar] [CrossRef] [Green Version]
- Gellersen, B.; Brosens, J.J. Cyclic decidualization of the human endometrium in reproductive health and failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.W.; Chen, Z.C.; Yang, Z.S.; Yang, Y.; Yan, Y.P.; Liu, Y.F.; Pan, J.M.; Su, R.W.; Yang, Z.M. Blastocyst-induced ATP release from luminal epithelial cells initiates decidualization through the P2Y2 receptor in mice. Sci. Signal 2020, 13, 3396. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Shembrey, C.; Smith, J.; Paquet-Fifield, S.; Behrenbruch, C.; Beyit, L.M.; Thomson, B.; Heriot, A.G.; Cao, Y.; Hollande, F. Laminin 521 enhances self-renewal via STAT3 activation and promotes tumor progression in colorectal cancer. Cancer Lett. 2020, 476, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Liang, X.; Liang, X.H.; Wang, T.S.; Qi, Q.R.; Deng, W.B.; Sha, A.G.; Yang, Z.M. The mesenchymal-epithelial transition during in vitro decidualization. Reprod. Sci. 2013, 20, 354–360. [Google Scholar] [CrossRef] [Green Version]
- Kisalus, L.L.; Herr, J.C.; Little, C.D. Immunolocalization of extracellular matrix proteins and collagen synthesis in first-trimester human decidua. Anat. Rec. 1987, 218, 402–415. [Google Scholar] [CrossRef]
- Church, H.J.; Vićovac, L.M.; Williams, J.D.; Hey, N.A.; Aplin, J.D. Laminins 2 and 4 are expressed by human decidual cells. Lab. Investig. 1996, 74, 21–32. [Google Scholar]
- Blankenship, T.N.; Given, R.L. Loss of laminin and type IV collagen in uterine luminal epithelial basement membranes during blastocyst implantation in the mouse. Anat. Rec. 1995, 243, 27–36. [Google Scholar] [CrossRef]
- Scott, L.E.; Weinberg, S.H.; Lemmon, C.A. Mechanochemical signaling of the extracellular matrix in epithelial-mesenchymal transition. Front. Cell. Dev. Biol. 2019, 7, 135. [Google Scholar] [CrossRef]
- Pitsidianaki, I.; Morgan, J.; Adams, J.; Campbell, K. Mesenchymal-to-epithelial transitions require tissue-specific interactions with distinct laminins. J. Cell. Biol. 2021, 220, e202010154. [Google Scholar] [CrossRef]
- Large, M.J.; DeMayo, F.J. The regulation of embryo implantation and endometrial decidualization by progesterone receptor signaling. Mol. Cell. Endocrinol. 2012, 358, 155–165. [Google Scholar] [CrossRef] [Green Version]
- Paria, B.C.; Tan, J.; Lubahn, D.B.; Dey, S.K.; Das, S.K. Uterine decidual response occurs in estrogen receptor-alpha-deficient mice. Endocrinology 1999, 2704–2710. [Google Scholar] [CrossRef] [PubMed]
- Vilar-Rojas, C.; Castro-Osuna, G.; Hicks, J.J. Cyclic AMP and cyclic GMP in the implantation site of the rat. Int. J. Fertil. 1982, 27, 56–59. [Google Scholar]
- Rankin, J.C.; Ledford, B.E.; Baggett, B. Early involvement of cyclic nucleotides in the artificially stimulated decidual cell reaction in the mouse uterus. Biol. Reprod. 1977, 17, 549–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, P.V.; Bergström, S. Induction of blastocyst implantation in mice by cyclic AMP. J. Reprod. Fertil. 1975, 43, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Telgmann, R.; Maronde, E.; Taskén, K.; Gellersen, B. Activated protein kinase A is required for differentiation-dependent transcription of the decidual prolactin gene in human endometrial stromal cells. Endocrinology 1997, 138, 929–937. [Google Scholar] [CrossRef]
- Ruan, Y.C.; Guo, J.H.; Liu, X.; Zhang, R.; Tsang, L.L.; Dong, J.D.; Chen, H.; Yu, M.K.; Jiang, X.; Zhang, X.H.; et al. Activation of the epithelial Na+ channel triggers prostaglandin E2 release and production required for embryo implantation. Nat. Med. 2012, 18, 1112–1117. [Google Scholar] [CrossRef]
- Mantena, S.R.; Kannan, A.; Cheon, Y.P.; Li, Q.; Johnson, P.F.; Bagchi, I.C.; Bagchi, M.K. C/EBPbeta is a critical mediator of steroid hormone-regulated cell proliferation and differentiation in the uterine epithelium and stroma. Proc. Natl. Acad. Sci. USA 2006, 103, 1870–1875. [Google Scholar] [CrossRef] [Green Version]
- Li, D.D.; Yang, Z.Q.; Guo, C.H.; Yue, L.; Duan, C.C.; Cao, H.; Guo, B.; Yue, Z.P. Hmgn1 acts downstream of C/EBPβ to regulate the decidualization of uterine stromal cells in mice. Cell. Cycle 2015, 14, 3461–3474. [Google Scholar] [CrossRef] [Green Version]
- Niehof, M.; Manns, M.P.; Trautwein, C. CREB controls LAP/C/EBP beta transcription. Mol. Cell. Biol. 1997, 17, 3600–3613. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.R.; Zhao, X.Y.; Zuo, R.J.; Wang, T.S.; Gu, X.W.; Liu, J.L.; Yang, Z.M. Involvement of atypical transcription factor E2F8 in the polyploidization during mouse and human decidualization. Cell. Cycle 2015, 14, 1842–1858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.T.; Zhang, H.Y.; Chen, S.T.; Li, M.Y.; Fu, T.; Yang, Z.M. The detrimental effects of stress-induced glucocorticoid exposure on mouse uterine receptivity and decidualization. FASEB J. 2020, 34, 14200–14216. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.X.; Hu, W.; Jin, Z.Y.; Diao, H.L.; Liu, L.; Yang, Y.; Fu, T.; Yang, Z.M. Nucleolar stress regulates stromal-epithelial transition via NPM1 during decidualization. Reproduction 2020, 160, 491–500. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Species | Sequence (5’-3’) | Application | Accession Number | Product Size |
|---|---|---|---|---|---|
| Rpl7 | Mouse | GCAGATGTACCGCACTGAGATTC ACCTTTGGGCTTACTCCATTGATA | RT-qPCR | NM_011291.5 | 129 bp |
| Prl8a2 | Mouse | AGCCAGAAATCACTGCCACT TGATCCATGCACCCATAAAA | RT-qPCR | NM_010088 | 119 bp |
| Laminin A5 | Mouse | AGCAAGGCGATCCAAGTGTT TTGAGGTCCACGTACTTGCC | RT-qPCR | NM_001081171.2 | 242 bp |
| Laminin B1 | Mouse | GCGCTGAACAATAGCTGCTC AATAAGCCCCTTCAGGCACC | RT-qPCR | NM_008482.3 | 241 bp |
| Laminin B2 | Mouse | CTGGTGACCAACCGAGAGAC CAGCGCAGTAGCAGGTCATA | RT-qPCR | NM_008483.3 | 128 bp |
| Laminin C1 | Mouse | CGGAGTTTGTTAATGCCGCC TCGGCCTGGTTGTTGTAGTC | RT-qPCR | NM_010683.2 | 187 bp |
| C/EBPβ | Mouse | GACAAGCTGAGCGACGAGTA TGCTTGAACAAGTTCCGCAG | RT-qPCR | NM_001287738.1 | 197 bp |
| IGFBP1 | Human | CCAAACTGCAACAAGAATG GTAGACGCACCAGCAGAG | RT-qPCR | NM_001013029 | 87 bp |
| RPL7 | Human | CTGCTGTGCCAGAAACCCTT TCTTGCCATCCTCGCCAT | RT-qPCR | NM_000971 | 194 bp |
| LAMININ A5 | Human | AGTGCCAGTCCTGTAACTGC GATAGGTGCCATCCAGGCTC | RT-qPCR | NM_005560.6 | 99 bp |
| NC | - | CTCCGAACGTGTCACGT | siRNA | ||
| Laminin A5 | Mouse | GCGACTGTGAGTCAGACTT | siRNA | ||
| C/EBPβ | Mouse | GAGCGACGAGUACAAGAUG | siRNA | ||
| LAMININ A5 | Human | GCCTCGTGCTGTTGTATGA | siRNA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.-S.; Pan, H.-Y.; Shi, W.-W.; Chen, S.-T.; Wang, Y.; Li, M.-Y.; Zhang, H.-Y.; Yang, C.; Liu, A.-X.; Yang, Z.-M. Regulation and Function of Laminin A5 during Mouse and Human Decidualization. Int. J. Mol. Sci. 2022, 23, 199. https://doi.org/10.3390/ijms23010199
Yang Z-S, Pan H-Y, Shi W-W, Chen S-T, Wang Y, Li M-Y, Zhang H-Y, Yang C, Liu A-X, Yang Z-M. Regulation and Function of Laminin A5 during Mouse and Human Decidualization. International Journal of Molecular Sciences. 2022; 23(1):199. https://doi.org/10.3390/ijms23010199
Chicago/Turabian StyleYang, Zhen-Shan, Hai-Yang Pan, Wen-Wen Shi, Si-Ting Chen, Ying Wang, Meng-Yuan Li, Hai-Yi Zhang, Chen Yang, Ai-Xia Liu, and Zeng-Ming Yang. 2022. "Regulation and Function of Laminin A5 during Mouse and Human Decidualization" International Journal of Molecular Sciences 23, no. 1: 199. https://doi.org/10.3390/ijms23010199