
Nanoplastic Impact on the Gut-Brain Axis: Current Knowledge and Future Directions

 , , and
, , and
Abstract
:1. Introduction
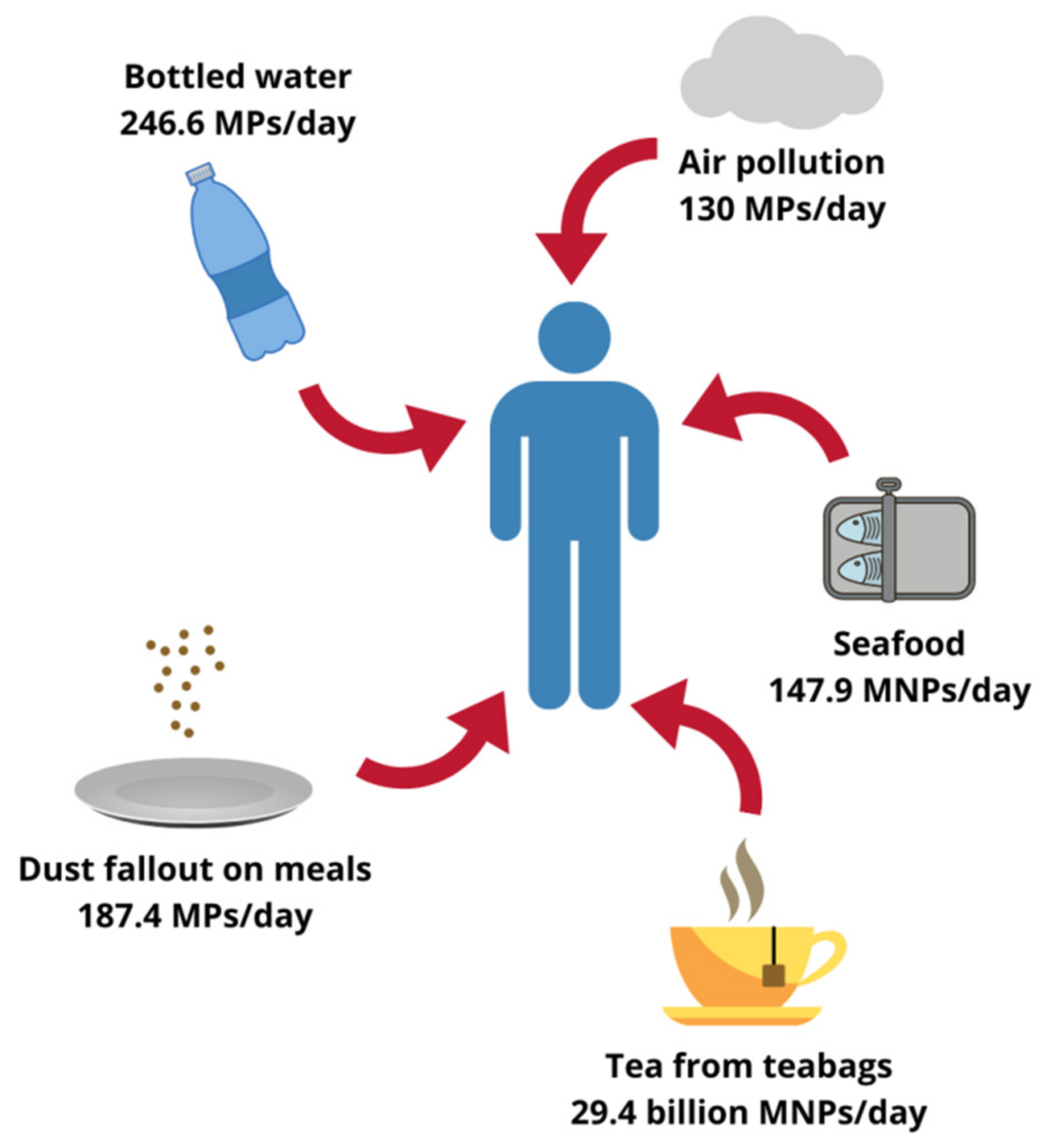
2. Plastics in Human Environment
3. Toxic Potential of Plastic Particles
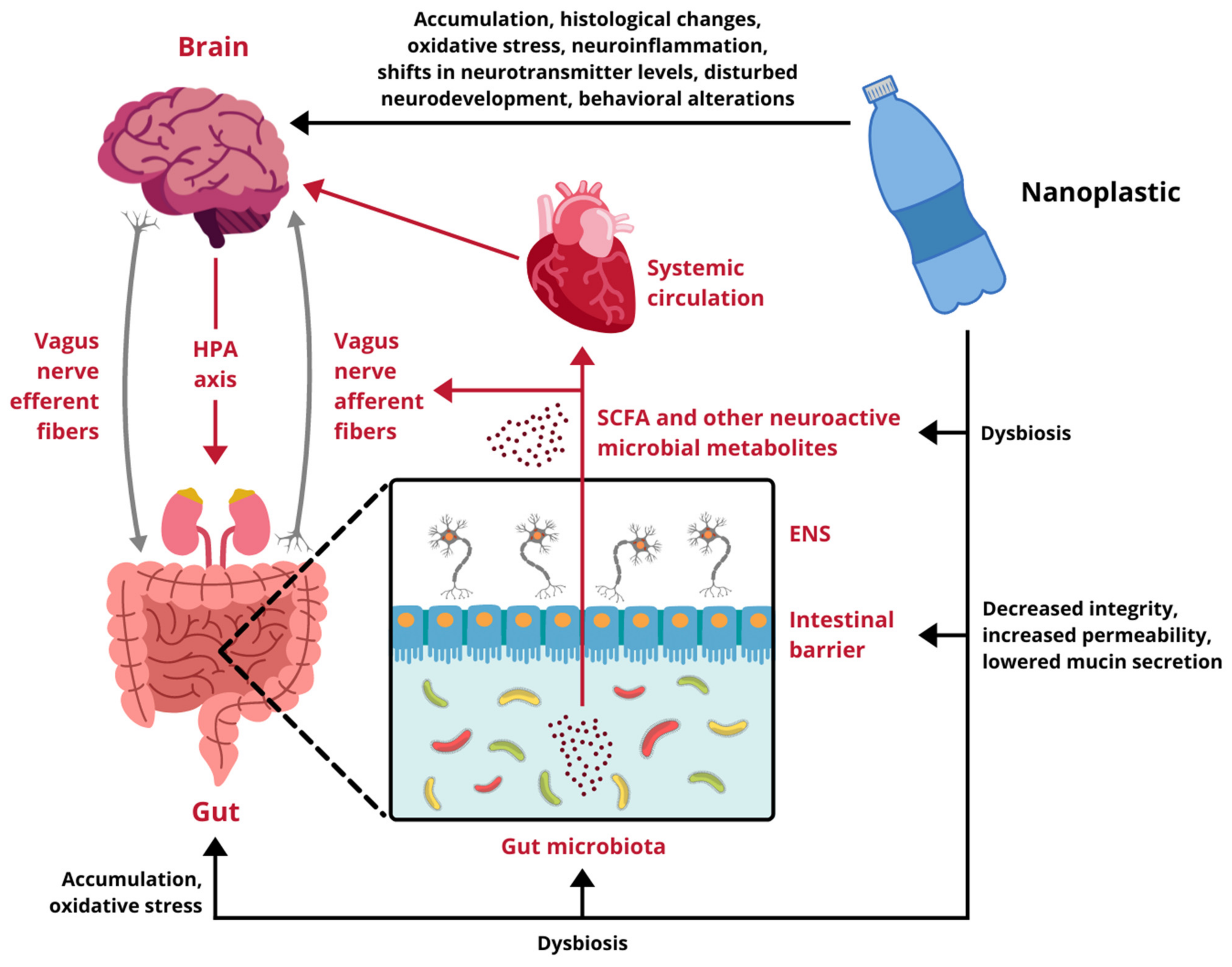
4. The Gut-Brain Axis
5. Impact of Nanoplastic Exposure on the Gut-Brain Axis
5.1. In Vitro Studies on Cellular Cultures
5.2. In Vivo Studies on Fish
5.3. In Vivo Studies on Rodents
6. Main Findings
7. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Haram, L.E.; Carlton, J.T.; Ruiz, G.M.; Maximenko, N.A. A Plasticene Lexicon. Mar. Pollut. Bull. 2020, 150, 110714. [Google Scholar] [CrossRef]
- Alimba, C.G.; Faggio, C. Microplastics in the marine environment: Current trends in environmental pollution and mechanisms of toxicological profile. Environ. Toxicol. Pharmacol. 2019, 68, 61–74. [Google Scholar] [CrossRef]
- Plastic Europe—Association of Plastics Manufactures. Plastics—The Facts 2020; Plastic Europe: Brussels, Belgium, 2020; Volume 1, pp. 1–64. [Google Scholar]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, 25–29. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, M.; Lebreton, L.C.M.; Carson, H.S.; Thiel, M.; Moore, C.J.; Borerro, J.C.; Galgani, F.; Ryan, P.G.; Reisser, J. Plastic Pollution in the World’s Oceans: More than 5 Trillion Plastic Pieces Weighing over 250,000 Tons Afloat at Sea. PLoS ONE 2014, 9, e111913. [Google Scholar] [CrossRef] [Green Version]
- Gigault, J.; ter Halle, A.; Baudrimont, M.; Pascal, P.-Y.; Gauffre, F.; Phi, T.-L.; El Hadri, H.; Grassl, B.; Reynaud, S. Current opinion: What is a nanoplastic? Environ. Pollut. 2018, 235, 1030–1034. [Google Scholar] [CrossRef]
- Prüst, M.; Meijer, J.; Westerink, R.H.S. The plastic brain: Neurotoxicity of micro- and nanoplastics. Part. Fibre Toxicol. 2020, 17, 24. [Google Scholar] [CrossRef]
- Yong, C.Q.Y.; Valiyaveetill, S.; Tang, B.L. Toxicity of microplastics and nanoplastics in Mammalian systems. Int. J. Environ. Res. Public Health 2020, 17, 1509. [Google Scholar] [CrossRef] [Green Version]
- Waring, R.H.; Harris, R.M.; Mitchell, S.C. Plastic contamination of the food chain: A threat to human health? Maturitas 2018, 115, 64–68. [Google Scholar] [CrossRef]
- Blair, R.M.; Waldron, S.; Phoenix, V.; Gauchotte-Lindsay, C. Micro- and Nanoplastic Pollution of Freshwater and Wastewater Treatment Systems. Springer Sci. Rev. 2017, 5, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, H.; Hollman, P.C.H.; Peters, R.J.B. Potential Health Impact of Environmentally Released Micro- and Nanoplastics in the Human Food Production Chain: Experiences from Nanotoxicology. Environ. Sci. Technol. 2015, 49, 8932–8947. [Google Scholar] [CrossRef]
- Chen, C.-L. Regulation and Management of Marine Litter. Mar. Anthropog. Litter 2015, 395–428. [Google Scholar] [CrossRef] [Green Version]
- Lehner, R.; Weder, C.; Petri-Fink, A.; Rothen-Rutishauser, B. Emergence of Nanoplastic in the Environment and Possible Impact on Human Health. Environ. Sci. Technol. 2019, 53, 1748–1765. [Google Scholar] [CrossRef]
- Alvarez-Román, R.; Naik, A.; Kalia, Y.N.; Guy, R.H.; Fessi, H. Skin penetration and distribution of polymeric nanoparticles. J. Control. Release 2004, 99, 53–62. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef]
- Dris, R.; Gasperi, J.; Saad, M.; Mirande, C.; Tassin, B. Synthetic fibers in atmospheric fallout: A source of microplastics in the environment? Mar. Pollut. Bull. 2016, 104, 290–293. [Google Scholar] [CrossRef]
- Dris, R.; Gasperi, J.; Mirande, C.; Mandin, C.; Guerrouache, M.; Langlois, V.; Tassin, B. A first overview of textile fibers, including microplastics, in indoor and outdoor environments. Environ. Pollut. 2017, 221, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Prata, J.C. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef]
- Hernandez, L.M.; Xu, E.G.; Larsson, H.C.E.; Tahara, R.; Maisuria, V.B.; Tufenkji, N. Plastic Teabags Release Billions of Microparticles and Nanoparticles into Tea. Environ. Sci. Technol. 2019, 53, 12300–12310. [Google Scholar] [CrossRef]
- Danopoulos, E.; Jenner, L.C.; Twiddy, M.; Rotchell, J.M. Microplastic contamination of seafood intended for human consumption: A systematic review and meta-analysis. Environ. Health Perspect. 2020, 128, 126002. [Google Scholar] [CrossRef]
- Rubio, L.; Marcos, R.; Hernández, A. Potential adverse health effects of ingested micro- and nanoplastics on humans. Lessons learned from in vivo and in vitro mammalian models. J. Toxicol. Environ. Health Part B Crit. Rev. 2020, 23, 51–68. [Google Scholar] [CrossRef]
- Toussaint, B.; Raffael, B.; Angers-Loustau, A.; Gilliland, D.; Kestens, V.; Petrillo, M.; Rio-Echevarria, I.M.; Van den Eede, G. Review of micro- and nanoplastic contamination in the food chain. Food Addit. Contam. Part A 2019, 36, 639–673. [Google Scholar] [CrossRef]
- Catarino, A.I.; Macchia, V.; Sanderson, W.G.; Thompson, R.C.; Henry, T.B. Low levels of microplastics (MP) in wild mussels indicate that MP ingestion by humans is minimal compared to exposure via household fibres fallout during a meal. Environ. Pollut. 2018, 237, 675–684. [Google Scholar] [CrossRef]
- Cox, K.D.; Covernton, G.A.; Davies, H.L.; Dower, J.F.; Juanes, F.; Dudas, S.E. Human Consumption of Microplastics. Environ. Sci. Technol. 2019, 53, 7068–7074. [Google Scholar] [CrossRef] [Green Version]
- Senathirajah, K.; Attwood, S.; Bhagwat, G.; Carbery, M.; Wilson, S.; Palanisami, T. Estimation of the mass of microplastics ingested—A pivotal first step towards human health risk assessment. J. Hazard. Mater. 2021, 404, 124004. [Google Scholar] [CrossRef]
- Schwabl, P.; Koppel, S.; Konigshofer, P.; Bucsics, T.; Trauner, M.; Reiberger, T.; Liebmann, B. Detection of various microplastics in human stool: A prospective case series. Ann. Intern. Med. 2019, 171, 453–457. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, M.C.A.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Shen, L. Functional morphology of the gastrointestinal tract. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 337, pp. 1–35. [Google Scholar] [CrossRef]
- Deisseroth, K. A Look inside the Brain. Sci. Am. 2016, 315, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Cryan, J.F.; O’riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The microbiota-gut-brain axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Ma, Q.; Xing, C.; Long, W.; Wang, H.Y.; Liu, Q.; Wang, R.F. Impact of microbiota on central nervous system and neurological diseases: The gut-brain axis. J. Neuroinflamm. 2019, 16, 53. [Google Scholar] [CrossRef] [Green Version]
- Morais, L.H.; Schreiber, H.L.; Mazmanian, S.K. The gut microbiota–brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef]
- Liang, S.; Wu, X.; Jin, F. Gut-brain psychology: Rethinking psychology from the microbiota–gut–brain axis. Front. Integr. Neurosci. 2018, 12, 33. [Google Scholar] [CrossRef]
- Erny, D.; De Angelis, A.L.H.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Carbery, M.; O’Connor, W.; Palanisami, T. Trophic transfer of microplastics and mixed contaminants in the marine food web and implications for human health. Environ. Int. 2018, 115, 400–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strużyńska, L.; Skalska, J. Mechanisms underlying neurotoxicity of silver nanoparticles. Adv. Exp. Med. Biol. 2018, 1048, 227–250. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, D.M.; Chircov, C.; Grumezescu, A.M.; Teleanu, R.I. Neurotoxicity of nanomaterials: An up-to-date overview. Nanomaterials 2019, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Calderón-Garcidueñas, L.; Reynoso-Robles, R.; Pérez-Guillé, B.; Mukherjee, P.S.; Gónzalez-Maciel, A. Combustion-derived nanoparticles, the neuroenteric system, cervical vagus, hyperphosphorylated alpha synuclein and tau in young Mexico City residents. Environ. Res. 2017, 159, 186–201. [Google Scholar] [CrossRef] [Green Version]
- Busch, M.; Bredeck, G.; Kämpfer, A.A.M.; Schins, R.P.F. Investigations of acute effects of polystyrene and polyvinyl chloride micro- and nanoplastics in an advanced in vitro triple culture model of the healthy and inflamed intestine. Environ. Res. 2021, 193, 110536. [Google Scholar] [CrossRef]
- Domenech, J.; Hernández, A.; Rubio, L.; Marcos, R.; Cortés, C. Interactions of polystyrene nanoplastics with in vitro models of the human intestinal barrier. Arch. Toxicol. 2020, 94, 2997–3012. [Google Scholar] [CrossRef]
- Hesler, M.; Aengenheister, L.; Ellinger, B.; Drexel, R.; Straskraba, S.; Jost, C.; Wagner, S.; Meier, F.; von Briesen, H.; Büchel, C.; et al. Multi-endpoint toxicological assessment of polystyrene nano- and microparticles in different biological models in vitro. Toxicol. In Vitro 2019, 61, 104610. [Google Scholar] [CrossRef]
- Walczak, A.P.; Kramer, E.; Hendriksen, P.J.M.; Helsdingen, R.; Van Der Zande, M.; Rietjens, I.M.C.M.; Bouwmeester, H. In vitro gastrointestinal digestion increases the translocation of polystyrene nanoparticles in an in vitro intestinal co-culture model. Nanotoxicology 2015, 9, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Jung, B.K.; Han, S.W.; Park, S.H.; Bae, J.S.; Choi, J.; Ryu, K.Y. Neurotoxic potential of polystyrene nanoplastics in primary cells originating from mouse brain. Neurotoxicology 2020, 81, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Schirinzi, G.F.; Pérez-Pomeda, I.; Sanchís, J.; Rossini, C.; Farré, M.; Barceló, D. Cytotoxic effects of commonly used nanomaterials and microplastics on cerebral and epithelial human cells. Environ. Res. 2017, 159, 579–587. [Google Scholar] [CrossRef]
- Murali, K.; Kenesei, K.; Li, Y.; Demeter, K.; Környei, Z.; Madarász, E. Uptake and bio-reactivity of polystyrene nanoparticles is affected by surface modifications, ageing and LPS adsorption: In vitro studies on neural tissue cells. Nanoscale 2015, 7, 4199–4210. [Google Scholar] [CrossRef]
- Hoelting, L.; Scheinhardt, B.; Bondarenko, O.; Schildknecht, S.; Kapitza, M.; Tanavde, V.; Tan, B.; Lee, Q.Y.; Mecking, S.; Leist, M.; et al. A 3-dimensional human embryonic stem cell (hESC)-derived model to detect developmental neurotoxicity of nanoparticles. Arch. Toxicol. 2013, 87, 721–733. [Google Scholar] [CrossRef] [Green Version]
- Weston, J.N.J.; Carrillo-Barragan, P.; Linley, T.D.; Reid, W.D.K.; Jamieson, A.J. New species of Eurythenes from hadal depths of the Mariana Trench, Pacific Ocean (Crustacea: Amphipoda). Zootaxa 2020, 4748, 163–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veneman, W.J.; Spaink, H.P.; Brun, N.R.; Bosker, T.; Vijver, M.G. Pathway analysis of systemic transcriptome responses to injected polystyrene particles in zebrafish larvae. Aquat. Toxicol. 2017, 190, 112–120. [Google Scholar] [CrossRef]
- Kang, H.M.; Byeon, E.; Jeong, H.; Kim, M.S.; Chen, Q.; Lee, J.S. Different effects of nano- and microplastics on oxidative status and gut microbiota in the marine medaka Oryzias melastigma. J. Hazard. Mater. 2021, 405, 124207. [Google Scholar] [CrossRef]
- Kashiwada, S. Distribution of nanoparticles in the see-through medaka (Oryzias latipes). Environ. Health Perspect. 2006, 114, 1697–1702. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.A.; Kozal, J.S.; Jayasundara, N.; Massarsky, A.; Trevisan, R.; Geitner, N.; Wiesner, M.; Levin, E.D.; Di Giulio, R.T. Uptake, tissue distribution, and toxicity of polystyrene nanoparticles in developing zebrafish (Danio rerio). Aquat. Toxicol. 2018, 194, 185–194. [Google Scholar] [CrossRef]
- Chae, Y.; Kim, D.; Kim, S.W.; An, Y.J. Trophic transfer and individual impact of nano-sized polystyrene in a four-species freshwater food chain. Sci. Rep. 2018, 8, 284. [Google Scholar] [CrossRef]
- Mattsson, K.; Ekvall, M.T.; Hansson, L.A.; Linse, S.; Malmendal, A.; Cedervall, T. Altered behavior, physiology, and metabolism in fish exposed to polystyrene nanoparticles. Environ. Sci. Technol. 2015, 49, 553–561. [Google Scholar] [CrossRef]
- Mattsson, K.; Johnson, E.V.; Malmendal, A.; Linse, S.; Hansson, L.A.; Cedervall, T. Brain damage and behavioural disorders in fish induced by plastic nanoparticles delivered through the food chain. Sci. Rep. 2017, 7, 11452. [Google Scholar] [CrossRef]
- Chen, Q.; Gundlach, M.; Yang, S.; Jiang, J.; Velki, M.; Yin, D.; Hollert, H. Quantitative investigation of the mechanisms of microplastics and nanoplastics toward zebrafish larvae locomotor activity. Sci. Total Environ. 2017, 584–585, 1022–1031. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Siregar, P.; Malhotra, N.; Lai, Y.H.; Liang, S.T.; Chen, J.R.; Chen, K.H.C.; Hsiao, C. Der Nanoplastics cause neurobehavioral impairments, reproductive and oxidative damages, and biomarker responses in zebrafish: Throwing up alarms of wide spread health risk of exposure. Int. J. Mol. Sci. 2020, 21, 1410. [Google Scholar] [CrossRef] [Green Version]
- Walczak, A.P.; Hendriksen, P.J.M.; Woutersen, R.A.; van der Zande, M.; Undas, A.K.; Helsdingen, R.; van den Berg, H.H.J.; Rietjens, I.M.C.M.; Bouwmeester, H. Bioavailability and biodistribution of differently charged polystyrene nanoparticles upon oral exposure in rats. J. Nanopart. Res. 2015, 17, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reineke, J.J.; Cho, D.Y.; Dingle, Y.T.; Morello, A.P.; Jacob, J.; Thanos, C.G.; Mathiowitz, E. Unique insights into the intestinal absorption, transit, and subsequent biodistribution of polymer-derived microspheres. Proc. Natl. Acad. Sci. USA 2013, 110, 13803–13808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fournier, S.B.; D’Errico, J.N.; Adler, D.S.; Kollontzi, S.; Goedken, M.J.; Fabris, L.; Yurkow, E.J.; Stapleton, P.A. Nanopolystyrene translocation and fetal deposition after acute lung exposure during late-stage pregnancy. Part. Fibre Toxicol. 2020, 17, 55. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Wan, Z.; Luo, T.; Fu, Z.; Jin, Y. Polystyrene microplastics induce gut microbiota dysbiosis and hepatic lipid metabolism disorder in mice. Sci. Total Environ. 2018, 631–632, 449–458. [Google Scholar] [CrossRef]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Rafiee, M.; Dargahi, L.; Eslami, A.; Beirami, E.; Jahangiri-rad, M.; Sabour, S.; Amereh, F. Neurobehavioral assessment of rats exposed to pristine polystyrene nanoplastics upon oral exposure. Chemosphere 2018, 193, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Jiang, X.; Zhou, Y.; Sumayyah, G.; Zhou, L.; Tu, B.; Qin, Q.; Qiu, J.; Qin, X.; Zou, Z.; et al. Results of a 30-day safety assessment in young mice orally exposed to polystyrene nanoparticles. Environ. Pollut. 2021, 292, 118184. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Ma, Y.; Han, X.; Chen, Y. Systematic toxicity evaluation of polystyrene nanoplastics on mice and molecular mechanism investigation about their internalization into Caco-2 cells. J. Hazard. Mater. 2021, 417, 126092. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Cell Models | NPs Type and Size | Exposure | Effects Related to the GB Axis | Reference |
|---|---|---|---|---|
| Human intestinal Caco-2, HT29-MTX-E12 and THP-1 monocul tures; triple culture human intestinal Caco-2/HT29-MTX-E12/THP-1 model(healthy or inflamed) | Pristine/amino-modified PS NPs (59 nm) PVC NPs (279 nm) | 24 h (1–50 µg/mL) | Monocultures/amino-modified PS: metabolic disruption, inflammation, DNA damage; healthy triple culture model/amino-modified PS: increased cytotoxicity, decrease of tight junction protein 1; inflamed triple culture model/PVC: loss of nuclei | [40] |
| Human intestinal Caco-2/HT29 and Caco-2/HT29 + Raji-B cells | PS NPs (5–100 nm) | 24 h (1–100 µg/mL) | No significant toxic effects | [41] |
| Human intestinal Caco-2/HT29-MTX-E12co-culture model | Carboxylated PS NPs (50 and 500 nm) | 24 h (0.1–100 µg/mL) | Uptake of NPs | [42] |
| Human intestinal Caco-2/HT29-MTX co-culture model | Pristine/positively/negatively charged PS NPs (50 nm), non-digestedor digested in vitro | 24 h (250 µg/mL) | Digested NPs: enhanced translocation across cells; positively charged NPs: increased intestinal barrier permeability | [43] |
| Murine mixed neuronal cells; primary astrocytes | PS NPs (100 nm) | 48 h (50–200 µg/mL) | Uptake of NPs; mixed neuronal cells: reduced cell viability, altered expression of Tubb3 and Gfap; primary astrocytes: increased expression of Tnfa and Il1b | [44] |
| Human neuronal T98G cells | PE NPs (100–600 nm) PS NPs (40–250 nm) | 24 h (0.05–10 µg/mL) | Increased ROS generation | [45] |
| Murine NE-4C Neuroectodermal stem cells; neuron-enriched primary brain cell cultures; primary astrocytes; microglial cultures; brain vascular endothelial cell cultures | Carboxylated/PEGylated PS NPs (45–70 nm), “fresh” or “aged”(6 months < of storage) | 1 h (50 µg/mL) 24 h (7.8–250 µg/mL) | “Fresh” carboxylated NPs: uptake by microglia; “aged” NPs: uptake and cytotoxicity in NE-4C neuronal stem cells and microglia; enhanced cellular uptake of NPs caused by LPS adsorption | [46] |
| Embryonic stem cell (hESC)-derived 3-dimensional model of human neural development | PE NPs (33 nm) | 48 h (5.6–1440 µg/mL) 18 days (5.6–360 µg/mL) | Uptake of NPs; reduced cell viability; oxidative stress; down-regulation of HES5, NOTCH1, FOXG1, NEUROD1 and ASCL1 | [47] |
| Fish | NPs Type and Size | Exposure | Effects Related to the GB Axis | Reference |
|---|---|---|---|---|
| Zebrafish (D. rerio) | PS NPs (700 nm) | Single-dose injection (5 mg/mL) | Altered expression of 26 genes 1 day and 51 genes 3 days post-injection; activation of the complement system; activation of oxidative stress-related pathways | [49] |
| Marine medaka (O. melastigma) | PS NPs (50 nm) | In water for 24 h (10 µg/mL) or 14 days (2.5 µg/mL) | NPs accumulation in the digestive system; induction of apoptosis in the gut; increased activity of SOD and CAT in the gut | [50] |
| Japanese medaka (O. latipes) | PS NPs (39.4 nm) | In water for 7 days (10 µg/mL) | NPs accumulation in the gut and brain | [51] |
| Zebrafish (D. rerio) | PS NPs (51 nm) | In water for 114 h (0.1–10 µg/mL) | NPs accumulation in the gut and head; behavioral alterations | [52] |
| Chinese medaka (O. sinensis) Dark chub (Z. temminckii) | PS NPs (51 nm) | In water for 7 days (5 µg/mL, individual toxicity) For 48 h (O. sinensis) or 24 h (Z. temminckii) via trophic transfer (C. reinhardtii → D. magna → O. sinensis → Z. temminckii) | Individual toxicity: behavioral alterations; O. sinensis/trophic transfer: NPs accumulation in the gut; Z. temminckii/trophic transfer: NPs accumulation in the gut and stomach | [53] |
| Crucian carp (C. carassius) | Sulfonated PS NPs (24 and 27 nm) | For 61 days via trophic transfer(Scenedesmus sp. → D. magna → C. carassius) | Histological changes in the brain; behavioral alterations | [54] |
| Crucian carp (C. carassius) | Amino-modified PS NPs (53 and 180 nm) | For 67 days via trophic transfer(Scenedesmus sp. → D. magna → C. carassius) | NPs accumulation in the brain; behavioral alterations | [55] |
| Zebrafish (D. rerio) | PS NPs (50 nm) | In water for 117 h (1 µg/mL) | Up-regulation of Gfap and α1-tubulin; decreased AChE activity; decreased levels of GSH; decreased body length; behavioral alterations | [56] |
| Zebrafish (D. rerio) | PS NPs (70 nm) | 7 days (0.5 and 1.5 µg/mL) 30 days (1.5 µg/mL) 7 weeks (5 µg/mL) | NPs accumulation in the gut and brain; lowered levels of AChE, dopamine, melatonin, vasopressin, 5-hydroxytryptophan, kisspeptin, γ-aminobutyric acid and oxytocin; behavioral alterations | [57] |
| Rodent | NPs Type and Size | Exposure | Effects Related to the GB Axis | Reference |
|---|---|---|---|---|
| Fischer rat | Pristine/positively/negatively charged PS NPs (50 nm) | Single-dose orally (125 mg/kg bw) | NPs accumulation in the gut | [58] |
| Sprague-Dawley rat | PS NPs (500 nm) | Orally for 5 h (100–125 mg/kg bw) | Accumulation in the GI tract and brain | [59] |
| Sprague-Dawley rat (pregnant) | PS NPs (20 nm) | Single-dose inhalation (2.64 × 1014 particles) | NPs accumulation in fetal brain | [60] |
| ICR mouse | PS NPs (500 nm) | Orally in drinking water for 5 weeks (0.1 or 1 µg/mL) | Higher load: decreased body weight; decrease in gut mucin secretion; lowered expression of Muc1 and Klf4; dysbiosis | [61] |
| Wistar rat | PS NPs (38.9 nm) | Orally for 35 days (1–10 mg/kg bw) | No changes in behavior | [63] |
| C57BL/6J mice | PS NPs (around 50 nm) | Orally for 30 days (0.2–10 mg/kg bw) | No changes in behavior; no inflammation/oxidative stress in the gut and brain; highest dose: damage to the intestinal wall; changes in microbiota composition | [64] |
| BALB/c mice | Pristine/carboxyl-/amino-modified PS NPs (100 nm) | Orally for 28 days (1 mg/day) | NPs accumulation in the gut and brain; histological damage to the gut and brain; inflammation in the brain; intestinal cells penetration confirmed in vitro | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodzicki, W.; Dziendzikowska, K.; Gromadzka-Ostrowska, J.; Kruszewski, M. Nanoplastic Impact on the Gut-Brain Axis: Current Knowledge and Future Directions. Int. J. Mol. Sci. 2021, 22, 12795. https://doi.org/10.3390/ijms222312795
Grodzicki W, Dziendzikowska K, Gromadzka-Ostrowska J, Kruszewski M. Nanoplastic Impact on the Gut-Brain Axis: Current Knowledge and Future Directions. International Journal of Molecular Sciences. 2021; 22(23):12795. https://doi.org/10.3390/ijms222312795
Chicago/Turabian StyleGrodzicki, Wojciech, Katarzyna Dziendzikowska, Joanna Gromadzka-Ostrowska, and Marcin Kruszewski. 2021. "Nanoplastic Impact on the Gut-Brain Axis: Current Knowledge and Future Directions" International Journal of Molecular Sciences 22, no. 23: 12795. https://doi.org/10.3390/ijms222312795