
The Membrane Proteome of Spores and Vegetative Cells of the Food-Borne Pathogen Bacillus cereus

 and
and
Abstract
:1. Introduction
2. Results
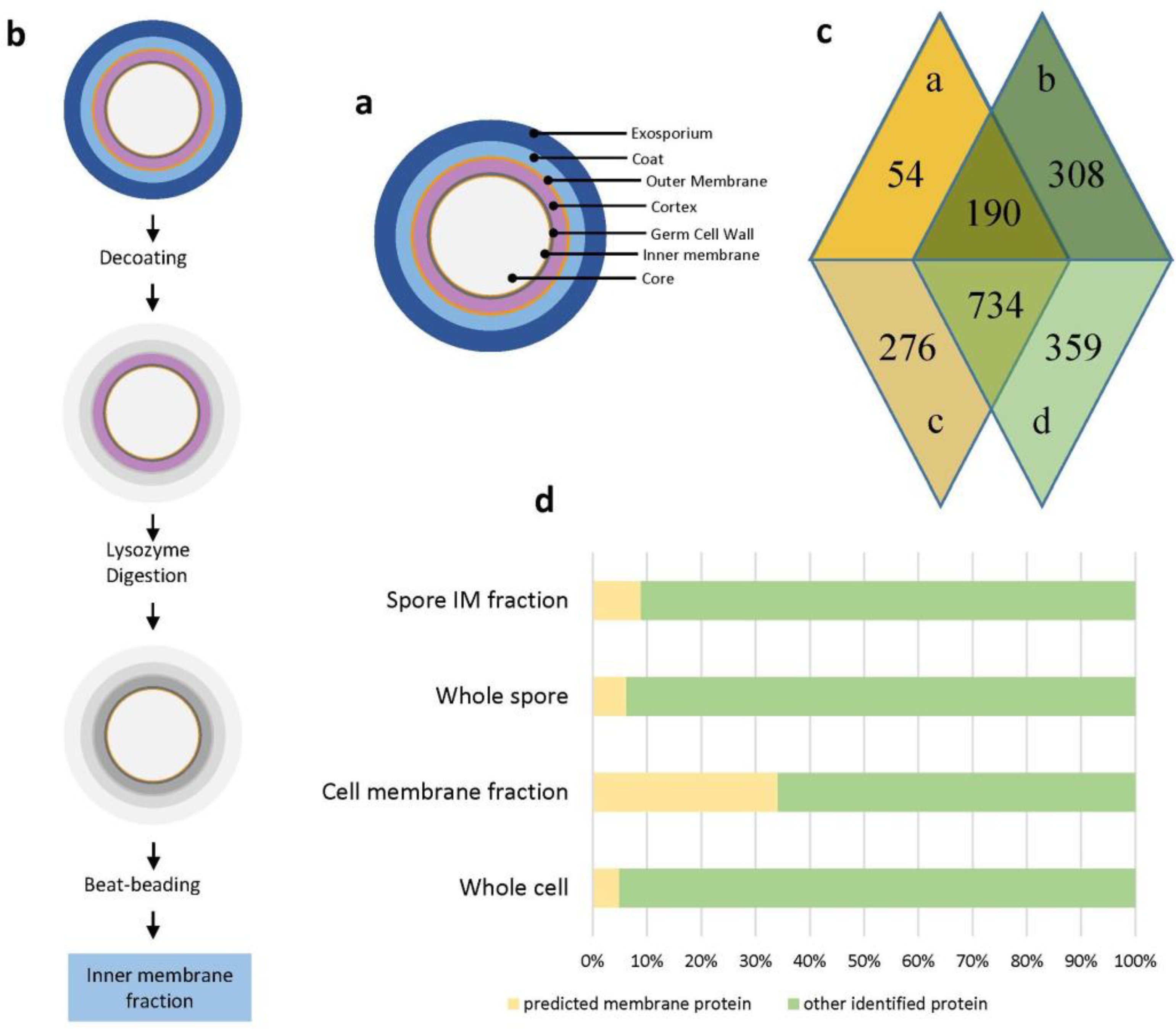
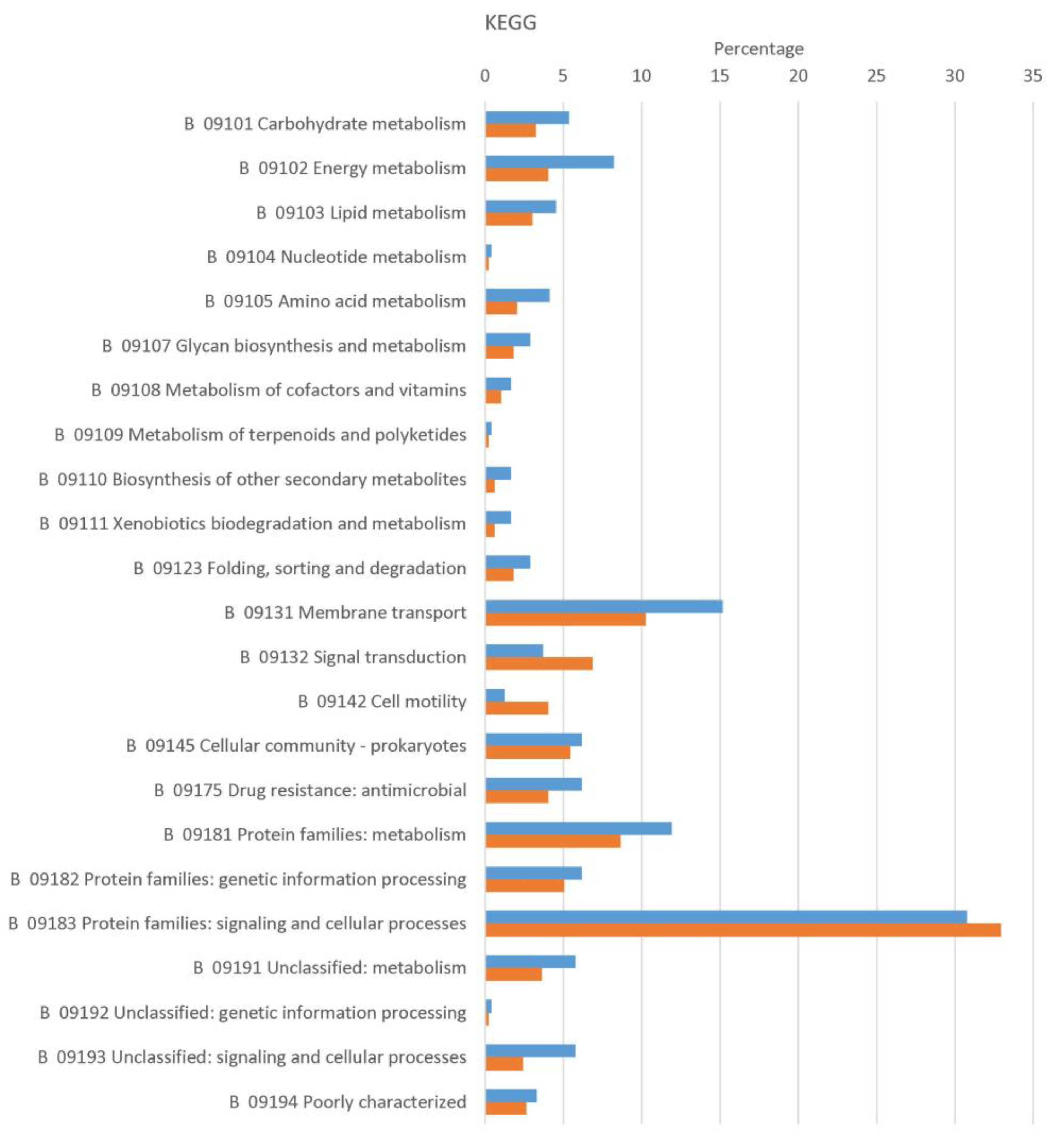
2.1. Identification, Prediction and Functional Classification of Spore Inner Membrane and Vegetative Cell Membrane Proteins
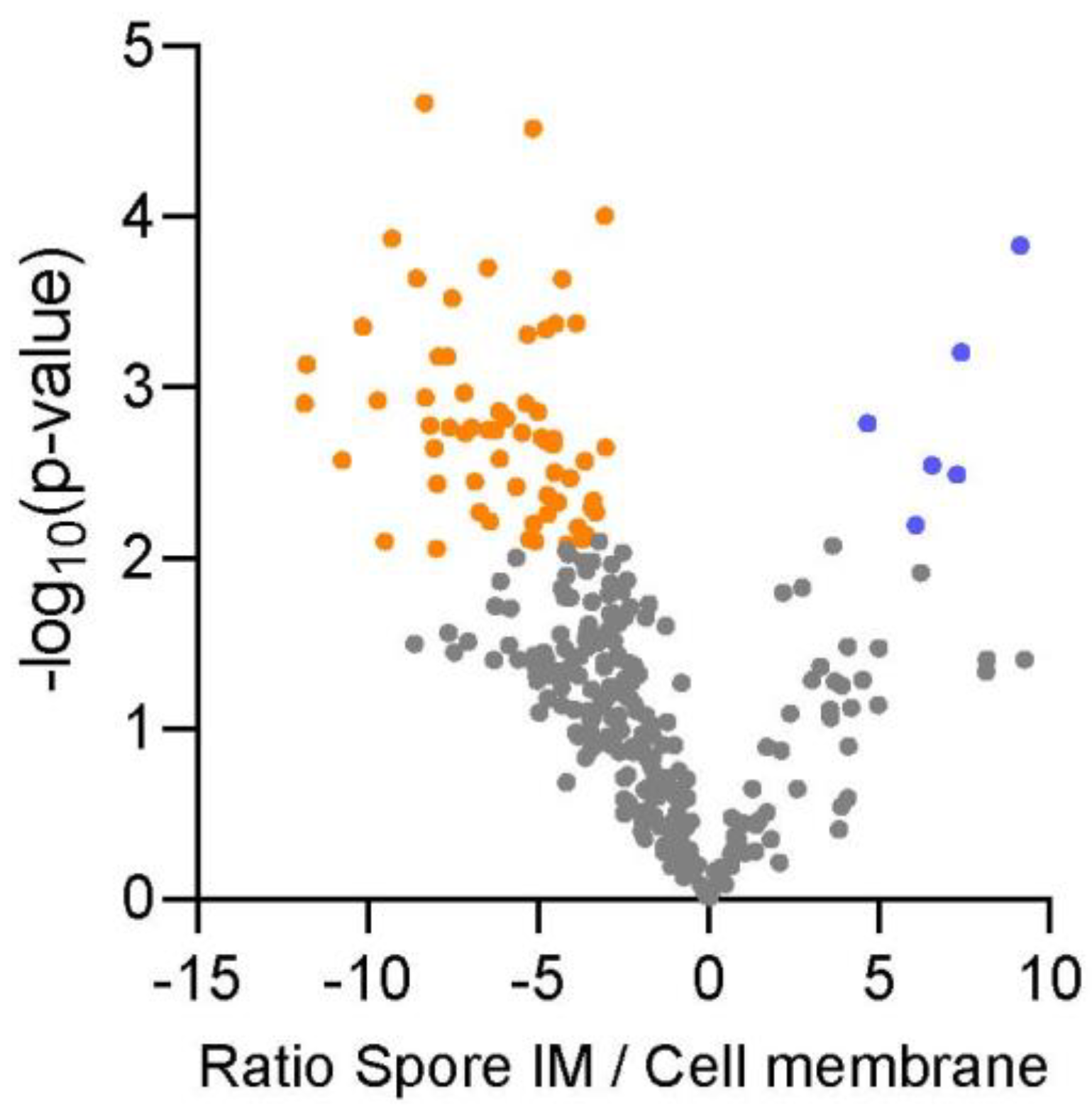
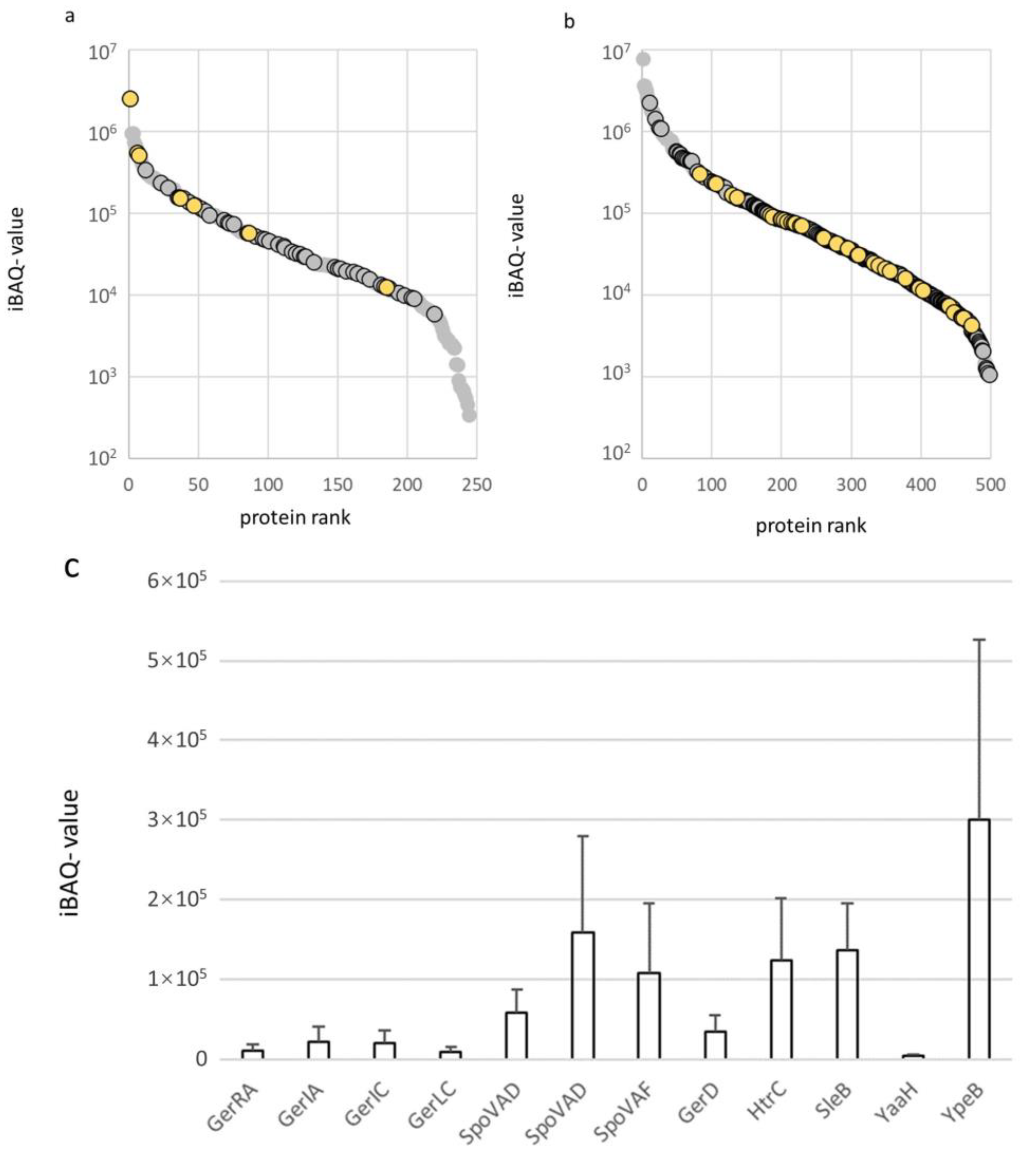
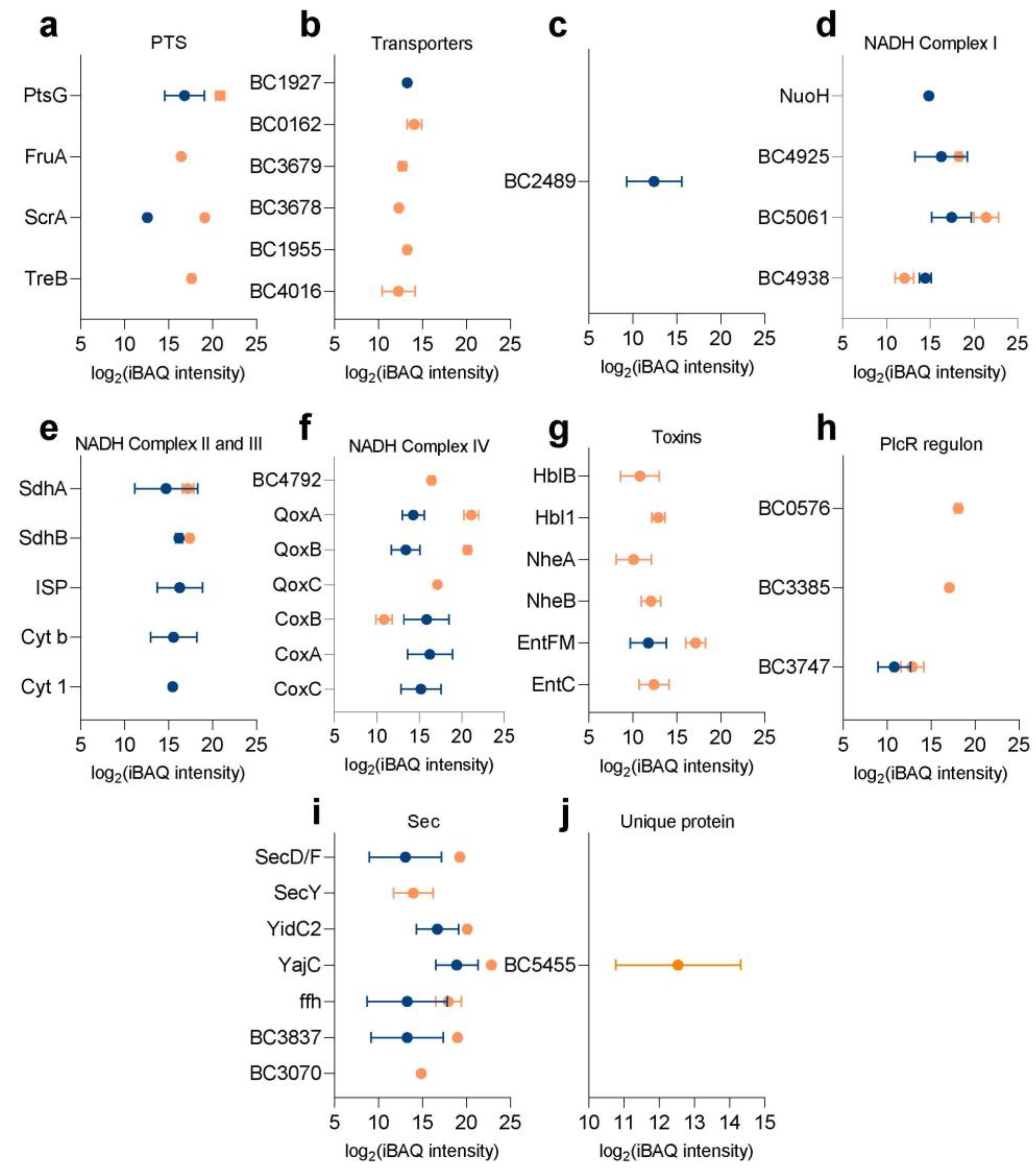
2.2. Protein Expression Levels of Membrane Proteins in the Spore Inner Membrane and Vegetative Cell Membrane of B. cereus
2.3. Specific Membrane Bound Proteins of Interest
2.3.1. Membrane Transport
2.3.2. Metabolism
2.3.3. Toxins and Virulence
2.4. Unique Proteins Detected in B. cereus
3. Discussion
4. Materials and Methods
4.1. Strain and Culture Conditions
4.2. Isolation of Spore Inner Membrane and Vegetative Cell Membrane
4.3. Sample Preparation for Proteomics Analysis
4.4. LC-MS/MS Analysis
4.5. Data Analysis
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abee, T.; Groot, M.N.; Tempelaars, M.; Zwietering, M.; Moezelaar, R.; van der Voort, M. Germination and outgrowth of spores of Bacillus cereus group members: Diversity and role of germinant receptors. Food Microbiol. 2011, 28, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Setlow, P. Germination of spores of Bacillus species: What we know and do not know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Cowan, A.E.; Olivastro, E.M.; Koppel, D.E.; Loshon, C.A.; Setlow, B.; Setlow, P. Lipids in the inner membrane of dormant spores of Bacillus species are largely immobile. Proc. Natl. Acad. Sci. USA 2004, 101, 7733–7738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laue, M.; Han, H.M.; Dittmann, C.; Setlow, P. Intracellular membranes of bacterial endospores are reservoirs for spore core membrane expansion during spore germination. Sci. Rep. 2018, 8, 11388. [Google Scholar] [CrossRef]
- Kang, M.K.; Tullman-Ercek, D. Engineering expression and function of membrane proteins. Methods 2018, 147, 66–72. [Google Scholar] [CrossRef]
- Frick, M.; Schmidt, C. Mass spectrometry—A versatile tool for characterising the lipid environment of membrane protein assemblies. Chem. Phys. Lipids 2019, 221, 145–157. [Google Scholar] [CrossRef]
- Otto, A.; Bernhardt, J.; Meyer, H.; Schaffer, M.; Herbst, F.A.; Siebourg, J.; Mäder, U.; Lalk, M.; Hecker, M.; Becher, D. Systems-wide temporal proteomic profiling in glucose-starved Bacillus subtilis. Nat. Commun. 2010, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Boekhorst, J.; Francke, C.; Siezen, R.J. LocateP: Genome-scale subcellular-location predictor for bacterial proteins. BMC Bioinform. 2008, 9, 173. [Google Scholar] [CrossRef] [Green Version]
- Yu, N.Y.; Wagner, J.R.; Laird, M.R.; Melli, G.; Rey, S.; Lo, R.; Dao, P.; Sahinalp, S.C.; Ester, M.; Foster, L.J.; et al. PSORTb 3.0: Improved protein subcellular localization prediction with refined localization subcategories and predictive capabilities for all prokaryotes. Bioinformatics 2010, 26, 1608–1615. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Abhyankar, W.; Ouwerling, N.; Dekker, H.L.; van Veen, H.; van der Wel, N.N.; Roseboom, W.; de Koning, L.J.; Brul, S.; de Koster, C.G. Bacillus subtilis spore inner membrane proteome. J. Proteome Res. 2016, 15, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Warda, A.K.; Xiao, Y.; Boekhorst, J.; Wells-Bennik, M.H.; Nierop Groot, M.N.; Abee, T. Analysis of germination capacity and germinant receptor (sub) clusters of genome-sequenced Bacillus cereus environmental isolates and model strains. Appl. Environ. Microbiol. 2017, 83, e02490–16. [Google Scholar] [CrossRef] [Green Version]
- Bernhards, C.B.; Chen, Y.; Toutkoushian, H.; Popham, D.L. HtrC is involved in proteolysis of YpeB during germination of Bacillus anthracis and Bacillus subtilis spores. J. Bacteriol. 2015, 197, 326–336. [Google Scholar] [CrossRef] [Green Version]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global quantification of mammalian gene expression control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerveny, L.; Straskova, A.; Dankova, V.; Hartlova, A.; Ceckova, M.; Staud, F.; Stulik, J. Tetratricopeptide repeat motifs in the world of bacterial pathogens: Role in virulence mechanisms. Infect. Immun. 2013, 81, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Taylor, C.W.; Da Fonseca, P.C.; Morris, E.P. IP3 receptors: The search for structure. Trends Biochem. Sci. 2004, 29, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Stelder, S.K.; Benito de Moya, C.; Hoefsloot, H.C.; De Koning, L.J.; Brul, S.; De Koster, C.G. Stoichiometry, absolute abundance, and localization of proteins in the Bacillus cereus spore coat insoluble fraction determined using a QconCAT approach. J. Proteome Res. 2017, 17, 903–917. [Google Scholar] [CrossRef] [Green Version]
- Stewart, K.A.V.; Setlow, P. Numbers of individual nutrient germinant receptors and other germination proteins in spores of Bacillus subtilis. J. Bacteriol. 2013, 195, 3575–3582. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; de Boer, R.; Vischer, N.; van Haastrecht, P.; Setlow, P.; Brul, S. Visualization of germination proteins in putative Bacillus cereus germinosomes. Int. J. Mol. Sci. 2020, 21, 5198. [Google Scholar] [CrossRef]
- Galinier, A.; Deutscher, J. Sophisticated regulation of transcriptional factors by the bacterial phosphoenolpyruvate: Sugar phosphotransferase system. J. Mol. Biol. 2017, 429, 773–789. [Google Scholar] [CrossRef]
- VanDrisse, C.M.; Escalante-Semerena, J.C. Protein acetylation in bacteria. Annu. Rev. Microbiol. 2019, 73, 111–132. [Google Scholar] [CrossRef]
- Ingledew, W.; Poole, R. The respiratory chains of Escherichia coli. Microbiol. Rev. 1984, 48, 222–271. [Google Scholar] [CrossRef] [PubMed]
- Hatefi, Y. The mitochondrial electron transport and oxidative phosphorylation system. Annu. Rev. Biochem. 1985, 54, 1015–1069. [Google Scholar] [CrossRef] [PubMed]
- Chagolla-López, A.; de la Vara, L.G.; Cabellos-Avelar, T.; Gómez-Lojero, C.; Cirlos, E.B.G. The composition of the Bacillus subtilis aerobic respiratory chain supercomplexes. J. Bioenerg. Biomembr. 2012, 44, 473–486. [Google Scholar]
- Sorokin, A.; Candelon, B.; Guilloux, K.; Galleron, N.; Wackerow-Kouzova, N.; Ehrlich, S.D.; Bourguet, D.; Sanchis, V. Multiple-locus sequence typing analysis of Bacillus cereus and Bacillus thuringiensis reveals separate clustering and a distinct population structure of psychrotrophic strains. Appl. Environ. Microbiol. 2006, 72, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [Green Version]
- Ehling-Schulz, M.; Vukov, N.; Schulz, A.; Shaheen, R.; Andersson, M.; Märtlbauer, E.; Scherer, S. Identification and partial characterization of the nonribosomal peptide synthetase gene responsible for cereulide production in emetic Bacillus cereus. Appl. Environ. Microbiol. 2005, 71, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Gohar, M.; Faegri, K.; Perchat, S.; Ravnum, S.; Økstad, O.A.; Gominet, M.; Kolstø, A.B.; Lereclus, D. The PlcR virulence regulon of Bacillus cereus. PLoS ONE 2008, 3, e2793. [Google Scholar] [CrossRef]
- Tsirigotaki, A.; De Geyter, J.; Šoštaric, N.; Economou, A.; Karamanou, S. Protein export through the bacterial Sec pathway. Nat. Rev. Microbiol. 2017, 15, 21–36. [Google Scholar] [CrossRef]
- Palmer, T.; Berks, B.C. The twin-arginine translocation (Tat) protein export pathway. Nat. Rev. Microbiol. 2012, 10, 483–496. [Google Scholar] [CrossRef]
- Auclair, S.M.; Bhanu, M.K.; Kendall, D.A. Signal peptidase I: Cleaving the way to mature proteins. Protein Sci. 2012, 21, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Barat, B.; Ray, W.K.; Helm, R.F.; Melville, S.B.; Popham, D.L. Membrane proteomes and ion transporters in Bacillus anthracis and Bacillus subtilis dormant and germinating spores. J. Bacteriol. 2019, 201, e00662–18. [Google Scholar] [CrossRef] [Green Version]
- Anderson, I.; Sorokin, A.; Kapatral, V.; Reznik, G.; Bhattacharya, A.; Mikhailova, N.; Burd, H.; Joukov, V.; Kaznadzey, D.; Walunas, T.; et al. Comparative genome analysis of Bacillus cereus group genomes with Bacillus subtilis. FEMS Microbiol. Lett. 2005, 250, 175–184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groot, P.W.; de Boer, A.D.; Cunningham, J.; Dekker, H.L.; de Jong, L.; Hellingwerf, K.J.; de Koster, C.; Klis, F.M. Proteomic analysis of Candida albicans cell walls reveals covalently bound carbohydrate-active enzymes and adhesins. Eukaryot. Cell 2004, 3, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Abhyankar, W.; Beek, A.T.; Dekker, H.; Kort, R.; Brul, S.; de Koster, C.G. Gel-free proteomic identification of the Bacillus subtilis insoluble spore coat protein fraction. Proteomics 2011, 11, 4541–4550. [Google Scholar] [CrossRef]
- Papanastasiou, M.; Orfanoudaki, G.; Koukaki, M.; Kountourakis, N.; Sardis, M.F.; Aivaliotis, M.; Karamanou, S.; Economou, A. The Escherichia coli peripheral inner membrane proteome. Mol. Cell. Proteom. 2013, 12, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.M. Strategies for the purification of membrane proteins. In Protein Chromatography; Springer: Berlin/Heidelberg, Germany, 2011; pp. 485–496. [Google Scholar]
- Madsen, C.S.; TerAvest, M.A. NADH dehydrogenases Nuo and Nqr1 contribute to extracellular electron transfer by Shewanella oneidensis MR-1 in bioelectrochemical systems. Sci. Rep. 2019, 9, 14959. [Google Scholar] [CrossRef]
- Gyan, S.; Shiohira, Y.; Sato, I.; Takeuchi, M.; Sato, T. Regulatory loop between redox sensing of the NADH/NAD+ ratio by Rex (YdiH) and oxidation of NADH by NADH dehydrogenase Ndh in Bacillus subtilis. J. Bacteriol. 2006, 188, 7062–7071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, S.L.; Guillemet, E.; Gohar, M.; Lereclus, D.; Ramarao, N. CwpFM (EntFM) is a Bacillus cereus potential cell wall peptidase implicated in adhesion, biofilm formation, and virulence. J. Bacteriol. 2010, 192, 2638–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanova, N.; Sorokin, A.; Anderson, I.; Galleron, N.; Candelon, B.; Kapatral, V.; Bhattacharyya, A.; Reznik, G.; Mikhailova, N.; Lapidus, A.; et al. Genome sequence of Bacillus cereus and comparative analysis with Bacillus anthracis. Nature 2003, 423, 87–91. [Google Scholar] [CrossRef]
- Abhyankar, W.; Hossain, A.H.; Djajasaputra, A.; Permpoonpattana, P.; Ter Beek, A.; Dekker, H.L.; Cutting, S.M.; Brul, S.; de Koning, L.J.; de Koster, C.G. In pursuit of protein targets: Proteomic characterization of bacterial spore outer layers. J. Proteome Res. 2013, 12, 4507–4521. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized ppb-range mass accuracies and proteome- wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Gene | Accession No. | N † | Description |
|---|---|---|---|---|
| GerKA | BC_0635 | Q81HZ1 | 1 | spore germination protein KA |
| GerKC | BC_0633 | Q81HZ3 | 1 | spore germination protein KC |
| GerSA | BC_3574 | Q81AJ2 | 1 | spore germination protein SA |
| GerRA | BC_0784 | Q81HM0 | 2 | GerA family spore germination protein |
| GerRC | BC_0783 | Q81HM1 | 2 | GerC family spore germination protein |
| GerQC | BC_3097 | Q81BQ4 | 2 | spore germination protein QC |
| GerIA | BC_4731 | Q816T6 | 2 | spore germination protein IA |
| GerLC | BC_0706 | Q81HS6 | 2 | spore germination protein LC |
| SpoVAA | BC_4070 | Q819B6 | 2 | stage V sporulation protein AA |
| SpoVAC | BC_5147 | Q815K0 | 2 | stage V sporulation protein AC |
| SpoVAD | BC_4067 | Q819B9 | 2 | stage V sporulation protein AD |
| SpoVAD | BC_5148 | Q815J9 | 3 | stage V sporulation protein AD |
| SpoVAE | BC_4065 | Q819C1 | 1 | stage V sporulation protein AE |
| SpoVAF | BC_4064 | Q819C2 | 3 | stage V sporulation protein AF |
| GerD | BC_0169 | Q81J09 | 3 | spore germination protein GerD |
| HtrC | BC_5458 | Q814H6 | 3 | serine protease YyxA |
| SleB | BC_2753 | P0A3V0 | 3 | Spore cortex-lytic enzyme |
| YaaH | BC_3607 | Q81AG3 | 3 | spore peptidoglycan hydrolase |
| YpeB | BC_2752 | Q813I5 | 3 | hypothetical protein (sporulation protein) |
| † Protein identified in N number of replicates out of 3. | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, X.; Swarge, B.N.; Dekker, H.L.; Roseboom, W.; Brul, S.; Kramer, G. The Membrane Proteome of Spores and Vegetative Cells of the Food-Borne Pathogen Bacillus cereus. Int. J. Mol. Sci. 2021, 22, 12475. https://doi.org/10.3390/ijms222212475
Gao X, Swarge BN, Dekker HL, Roseboom W, Brul S, Kramer G. The Membrane Proteome of Spores and Vegetative Cells of the Food-Borne Pathogen Bacillus cereus. International Journal of Molecular Sciences. 2021; 22(22):12475. https://doi.org/10.3390/ijms222212475
Chicago/Turabian StyleGao, Xiaowei, Bhagyashree N. Swarge, Henk L. Dekker, Winfried Roseboom, Stanley Brul, and Gertjan Kramer. 2021. "The Membrane Proteome of Spores and Vegetative Cells of the Food-Borne Pathogen Bacillus cereus" International Journal of Molecular Sciences 22, no. 22: 12475. https://doi.org/10.3390/ijms222212475