
Methotrexate Alters the Expression of microRNA in Fibroblast-like Synovial Cells in Rheumatoid Arthritis

 , ,
, ,
Abstract
:1. Introduction
2. Results
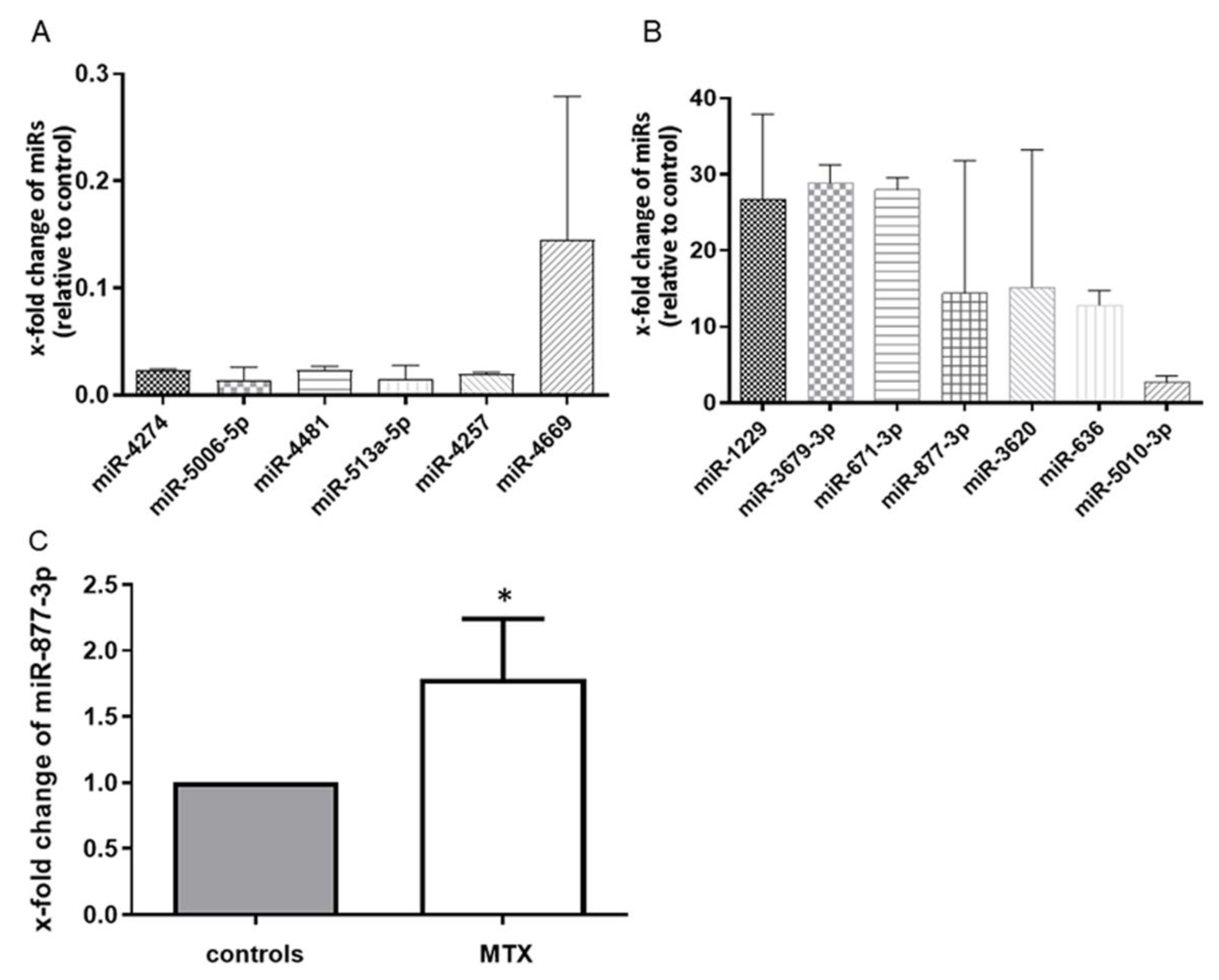
2.1. miR-877-3p is an MTX-Inducible miRNA in RA-FLS
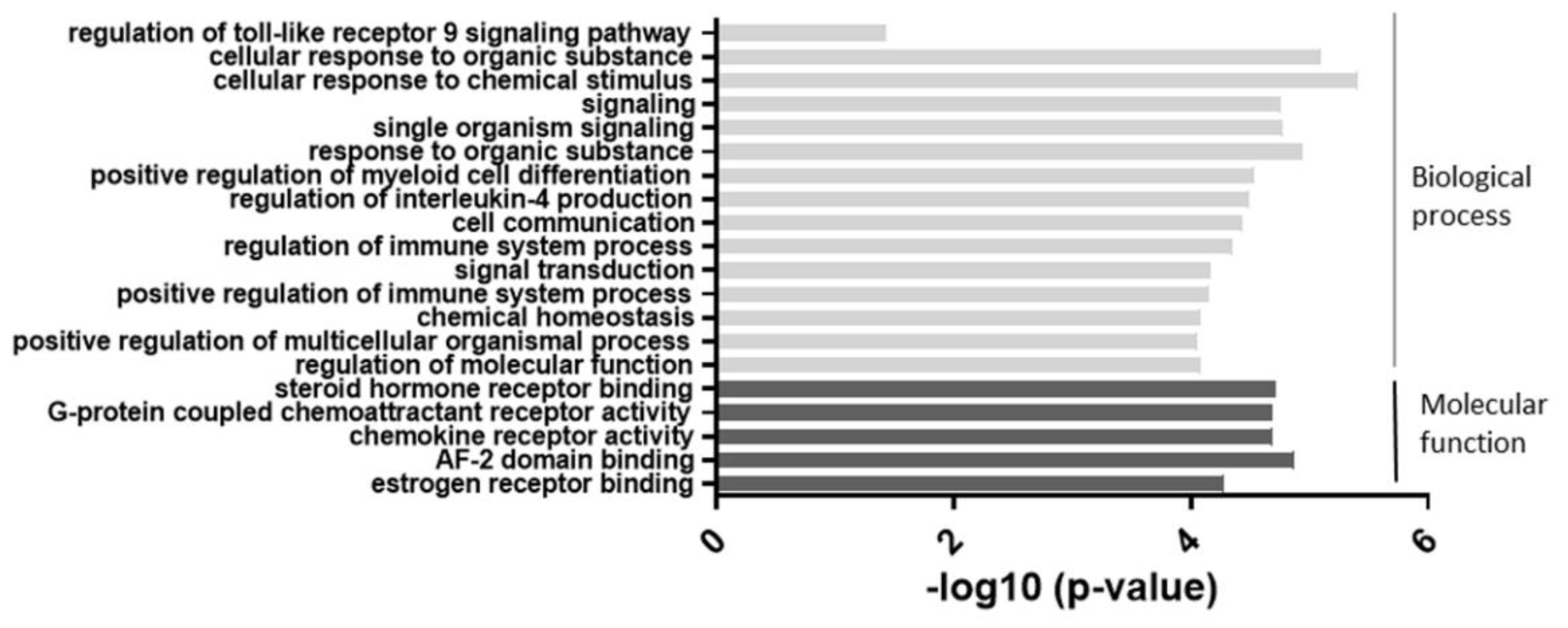
2.2. miR-877-3p Transfection Microarrays
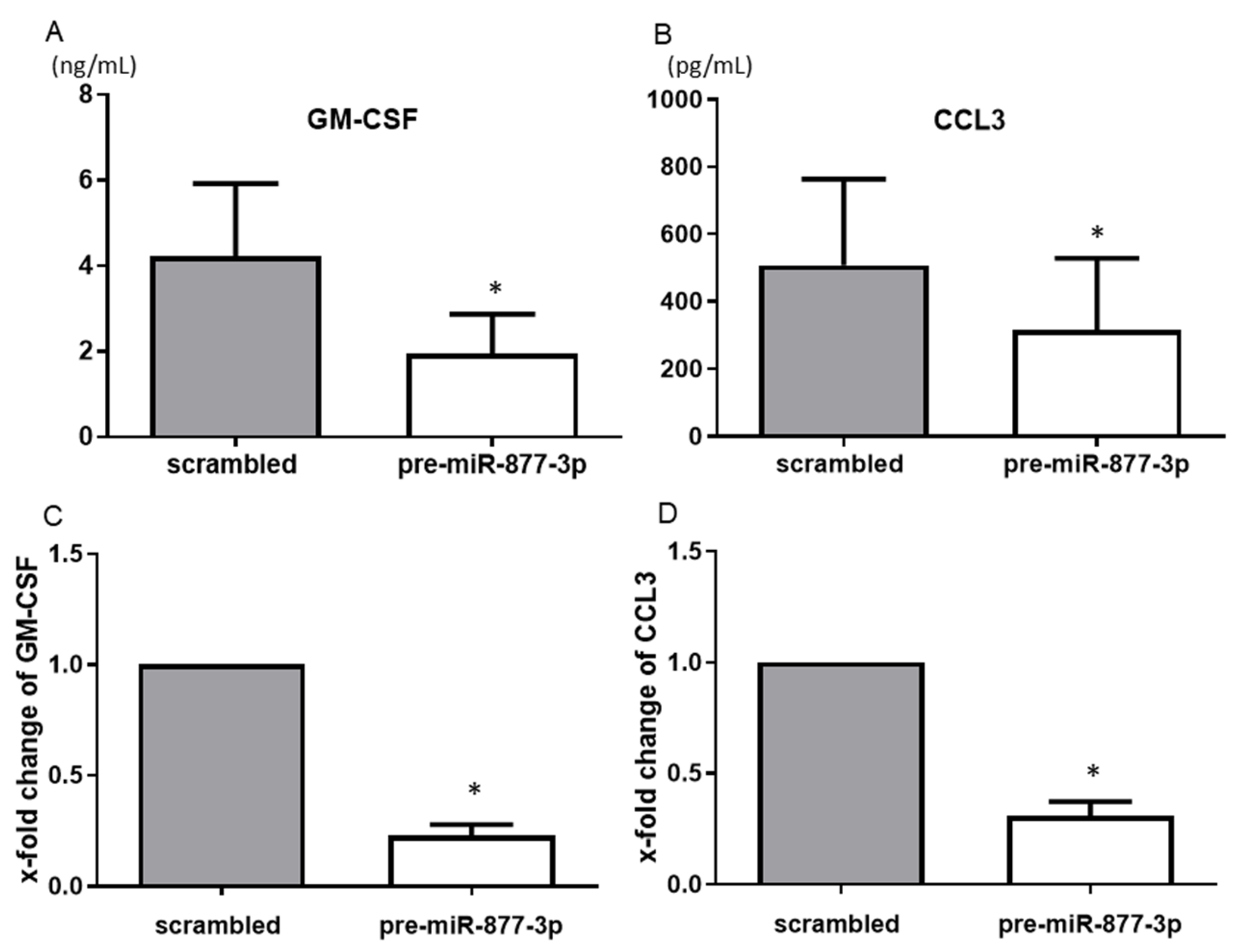
2.3. miR-877-3p Reduced the Productions of GM-CSF and CCL3
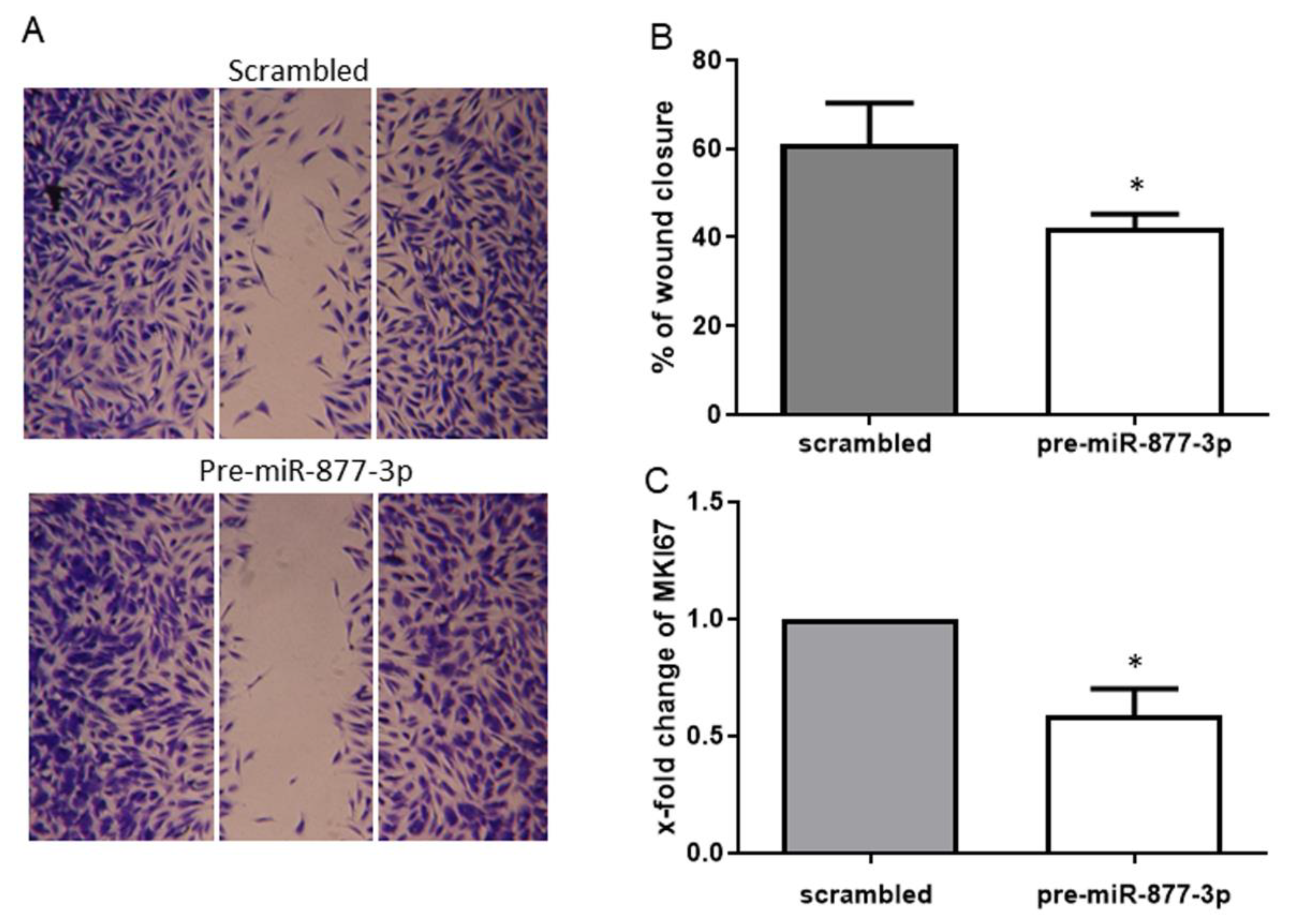
2.4. miR-877-3p Inhibited the Migratory Ability of RA-FLS
3. Discussion
4. Materials and Methods
4.1. Isolation of FLS and Treatment by MTX
4.2. RNA isolation and Quantitative Real-Time PCR Analysis
4.3. MicroRNA and DNA Microarray Assay Analyses
4.4. Transfection Experiments
4.5. Multiplex Cytokines/Chemokines Bead Assay and ELISA
4.6. Wound Healing Assay
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACR | American College of Rheumatology |
| CCL | CC chemokine ligand |
| DMARD | disease-modifying antirheumatic drugs |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ELISA | enzyme-linked immunosorbent assay |
| EULAR | European League against Rheumatism |
| FBS | fetal bovine serum |
| FLS | fibroblast-like synovial cells |
| GM-CSF | granulocyte macrophage colony-stimulating factor |
| GO | gene ontology |
| IL-1β | interleukin-1β |
| JAK/STAT | Janus kinase/signal transducer and activator of transcription |
| LPS | lipopolysaccharide |
| MKI67 | marker of proliferation Ki-67 |
| miRNA | microRNA |
| MTX | methotrexate |
| PI3K/AKT | phosphoinositide 3-kinase/v-akt murine thymoma |
| pre-miR | precursor miRNA |
| RA | rheumatoid arthritis |
| RNU6B | U6B small nuclear RNA |
| ROS | reactive oxygen species |
| RT-PCR | real-time polymerase chain reaction |
| SD | standard deviation |
| TNF-α | tumor necrosis factor-alpha |
References
- Bustamante, M.F.; Garcia-Carbonell, R.; Whisenant, K.D.; Guma, M. Fibroblast-like synoviocyte metabolism in the pathogenesis of rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Landewé, R.B.; Bijlsma, J.W.; Burmester, G.R.; Dougados, M.; Kerschbaumer, A.; McInnes, I.B.; Sepriano, A.; Van Vollehoven, R.F.; De Wit, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs: Update. Ann. Rheum. Dis. 2019, 79, 685–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutolo, M.; Sulli, A.; Pizzorni, C.; Seriolo, B.; Straub, R.H. Anti-inflammatory mechanisms of methotrexate in rheumatoid arthritis. Ann. Rheum. Dis. 2001, 60, 729–735. [Google Scholar] [CrossRef] [Green Version]
- Hirata, S.; Matsubara, T.; Saura, R.; Tateishi, H.; Hirohata, K. Inhibition of in vitro vascular endothelial cell proliferation and in vivo neovascularization by low-dose methotrexate. Arthritis Rheum. 1989, 32, 1065–1073. [Google Scholar] [CrossRef]
- Nakajima, A.; Hakoda, M.; Yamanaka, H.; Kamatani, N.; Kashiwazaki, S. Divergent effects of methotrexate on the clonal growth of T and B lymphocytes and synovial adherent cells from patients with rheumatoid arthritis. Ann. Rheum. Dis. 1996, 55, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, C.R.; Pickett, W.C.; Bell, D.H.; McClintock, D.K.; Oronsky, A.L.; Kerwar, S.S. Effect of low dose methotrexate on neutrophil chemotaxis induced by leukotriene B4 and complement C5a. J. Rheumatol. 1987, 14, 9–11. [Google Scholar]
- Lee, S.Y.; Park, S.H.; Lee, S.W.; Lee, S.H.; Son, M.K.; Choi, Y.H.; Chung, W.T.; Yoo, Y.H. Synoviocyte apoptosis may differentiate responder and non-responder patients to methotrexate treatment in rheumatoid arthritis. Arch. Pharm. Res. 2014, 37, 1286–1294. [Google Scholar] [CrossRef] [PubMed]
- Sung, J.Y.; Hong, J.H.; Kang, H.S.; Choi, I.; Lim, S.D.; Lee, J.K.; Seok, J.H.; Lee, J.H.; Hur, G.M. Methotrexate suppresses the interleukin-6 induced generation of reactive oxygen species in the synoviocytes of rheumatoid arthritis. Immunopharmacology 2000, 47, 35–44. [Google Scholar] [CrossRef]
- Spurlock, C.F., 3rd; Gass, H.M., 4th; Bryant, C.J.; Wells, B.C.; Olsen, N.J.; Aune, T.M. Methotrexate-mediated inhibition of nuclear factor kappaB activation by distinct pathways in T cells and fibroblast-like synoviocytes. Rheumatology 2015, 54, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Zou, Y.; Gao, G.; Zheng, S.; Wu, S.; Xie, X.; Tang, H. The biogenesis, function and clinical significance of circular RNAs in breast cancer. Cancer Biol Med. 2021, 18, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, N.; Kawakami, A. Recent findings regarding the effects of microRNAs on fibroblast-like synovial cells in rheumatoid arthritis. Immunol. Med. 2019, 42, 156–161. [Google Scholar] [CrossRef]
- Hong, S.M.; Liu, C.; Yin, Z.; Wu, L.; Qu, B.; Shen, N. MicroRNAs in Systemic Lupus Erythematosus: A Perspective on the Path from Biological Discoveries to Clinical Practice. Curr. Rheumatol. Rep. 2020, 22, 17. [Google Scholar] [CrossRef]
- Potenza, N.; Mosca, N.; Zappavigna, S.; Castiello, F.; Panella, M.; Ferri, C.; Vanacore, D.; Giordano, A.; Stiuso, P.; Caraglia, M.; et al. MicroRNA-125a-5p Is a Downstream Effector of Sorafenib in Its Antiproliferative Activity Toward Human Hepatocellular Carcinoma Cells. J. Cell Physiol. 2017, 232, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Takwi, A.A.; Li, Y.; Becker Buscaglia, L.E.; Zhang, J.; Choudhury, S.; Park, A.K.; Liu, M.; Young, K.H.; Park, W.Y.; Martin, R.C.; et al. A statin-regulated microRNA represses human c-Myc expression and function. EMBO Mol. Med. 2012, 4, 896–909. [Google Scholar] [CrossRef]
- Shah, M.Y.; Pan, X.; Fix, L.N.; Farwell, M.A.; Zhang, B. 5-Fluorouracil drug alters the microRNA expression profiles in MCF-7 breast cancer cells. J. Cell Physiol. 2011, 226, 1868–1878. [Google Scholar] [CrossRef]
- Duan, C.; Liu, Y.; Li, Y.; Chen, H.; Liu, X.; Chen, X.; Yue, J.; Zhou, X.; Yang, J. Sulfasalazine alters microglia phenotype by competing endogenous RNA effect of miR-136-5p and long non-coding RNA HOTAIR in cuprizone-induced demyelination. Biochem. Pharmacol. 2018, 155, 110–123. [Google Scholar] [CrossRef]
- Cunningham, C.C.; Wade, S.; Floudas, A.; Orr, C.; McGarry, T.; Cregan, S.; Fearon, U.; Veale, D.J. Serum miRNA Signature in Rheumatoid Arthritis and At-Risk Individuals. Front. Immunol. 2021, 12, 633201. [Google Scholar] [CrossRef]
- Daraghmeh, D.N.; King, C.; Wiese, M.D. A review of liquid biopsy as a tool to assess epigenetic, cfDNA and miRNA variability as methotrexate response predictors in patients with rheumatoid arthritis. Pharmacol. Res. 2021, 173, 105887. [Google Scholar] [CrossRef] [PubMed]
- Sode, J.; Krintel, S.B.; Carlsen, A.L.; Hetland, M.L.; Johansen, J.S.; Hørslev-Petersen, K.; Stengaard-Pedersen, K.; Ellingsen, T.; Burton, M.; Junker, P.; et al. Plasma MicroRNA Profiles in Patients with Early Rheumatoid Arthritis Responding to Adalimumab plus Methotrexate vs Methotrexate Alone: A Placebo-controlled Clinical Trial. J. Rheumatol. 2018, 45, 53–61. [Google Scholar] [CrossRef]
- Hong, H.; Yang, H.; Xia, Y. Circulating miR-10a as Predictor of Therapy Response in Rheumatoid Arthritis Patients Treated with Methotrexate. Curr. Pharm. Biotechnol. 2018, 19, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Patro, P.S.; Aggarwal, A. MicroRNA-132, miR-146a, and miR-155 as potential biomarkers of methotrexate response in patients with rheumatoid arthritis. Clin. Rheumatol. 2019, 38, 877–884. [Google Scholar] [CrossRef]
- Fiehn, C.; Wermann, M.; Pezzutto, A.; Hufner, M.; Heilig, B. Plasma GM-CSF concentrations in rheumatoid arthritis, systemic lupus erythematosus and spondyloarthropathy. Z. Rheumatol. 1992, 51, 121–126. [Google Scholar] [PubMed]
- Hazenberg, B.P.; Van Leeuwen, M.A.; Van Rijswijk, M.H.; Stern, A.C.; Vellenga, E. Correction of granulocytopenia in Felty’s syndrome by granulocyte-macrophage colony-stimulating factor. Simultaneous induction of interleukin-6 release and flare-up of the arthritis. Blood 1989, 74, 2769–2770. [Google Scholar] [CrossRef] [Green Version]
- Burgess, A.W.; Metcalf, D. The nature and action of granulocyte-macrophage colony stimulating factors. Blood 1980, 56, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.D.; Hamilton, J.A. Investigational therapies targeting the granulocyte macrophage colony-stimulating factor receptor-alpha in rheumatoid arthritis: Focus on mavrilimumab. Ther. Adv. Musculoskelet. Dis. 2018, 10, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuipers, H.F.; Rappert, A.A.; Mommaas, A.M.; van Haastert, E.S.; van der Valk, P.; Boddeke, H.W.; Biber, K.P.; van den Elsen, P.J. Simvastatin affects cell motility and actin cytoskeleton distribution of microglia. Glia 2006, 53, 115–123. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, H.B.; Zhou, L.; Cui, X.Q.; Fan, X.H. CCL3 participates in the development of rheumatoid arthritis by activating AKT. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6625–6632. [Google Scholar] [CrossRef]
- Rolle, K.; Piwecka, M.; Belter, A.; Wawrzyniak, D.; Jeleniewicz, J.; Barciszewska, M.Z.; Barciszewski, J. The Sequence and Structure Determine the Function of Mature Human miRNAs. PLoS ONE 2016, 11, e0151246. [Google Scholar] [CrossRef]
- Phillips, D.C.; Woollard, K.J.; Griffiths, H.R. The anti-inflammatory actions of methotrexate are critically dependent upon the production of reactive oxygen species. Br. J. Pharmacol. 2003, 138, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Hsu, P.; Hung, Y.; Liao, Y.; Liu, C.; Hour, C.; Kao, M.; Tsay, G.J.; Hung, H.; Liu, G.Y. Ornithine decarboxylase prevents methotrexate-induced apoptosis by reducing intracellular reactive oxygen species production. Apoptosis 2005, 10, 895–907. [Google Scholar] [CrossRef]
- Hsu, P.C.; Hour, T.C.; Liao, Y.F.; Hung, Y.C.; Liu, C.C.; Chang, W.H.; Kao, M.C.; Tsay, G.J.; Hung, H.C.; Liu, G.Y. Increasing ornithine decarboxylase activity is another way of prolactin preventing methotrexate-induced apoptosis: Crosstalk between ODC and BCL. Apoptosis 2006, 11, 389–399. [Google Scholar] [CrossRef]
- Quemeneur, L.; Gerland, L.M.; Flacher, M.; Ffrench, M.; Revillard, J.P.; Genestier, L. Differential control of cell cycle, proliferation, and survival of primary T lymphocytes by purine and pyrimidine nucleotides. J. Immunol. 2003, 170, 4986–4995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brzustewicz, E.; Bryl, E. The role of cytokines in the pathogenesis of rheumatoid arthritis--Practical and potential application of cytokines as biomarkers and targets of personalized therapy. Cytokine 2015, 76, 527–536. [Google Scholar] [CrossRef]
- Li, X.; Xu, H.; Huang, J.; Luo, D.; Lv, S.; Lu, X.; Xiao, C. Dysfunctions, Molecular Mechanisms, and Therapeutic Strategies of Regulatory T Cells in Rheumatoid Arthritis. Front. Pharmacol. 2021, 12, 716081. [Google Scholar] [CrossRef]
- Li, S.; Zhu, Y.; Liang, Z.; Wang, X.; Meng, S.; Xu, X.; Wu, J.; Ji, A.; Hu, Z.; Lin, Y.; et al. Up-regulation of p16 by miR-877-3p inhibits proliferation of bladder cancer. Oncotarget 2016, 7, 51773–51783. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Li, K. HOXD-AS1 facilitates cell migration and invasion as an oncogenic lncRNA by competitively binding to miR-877-3p and upregulating FGF2 in human cervical cancer. BMC Cancer 2020, 20, 924. [Google Scholar] [CrossRef]
- He, G.; Chen, J.; Huang, D. miR-877-3p promotes TGF-beta1-induced osteoblast differentiation of MC3T3-E1 cells by targeting Smad. Exp. Ther. Med. 2019, 18, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosik, K.S. MicroRNAs and cellular phenotypy. Cell 2010, 143, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Sood, P.; Krek, A.; Zavolan, M.; Macino, G.; Rajewsky, N. Cell-type-specific signatures of microRNAs on target mRNA expression. Proc. Natl. Acad. Sci. USA 2006, 103, 2746–2751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 rheumatoid arthritis classification criteria: An American College of Rheumatology/European League Against Rheumatism collaborative initiative. Ann. Rheum. Dis. 2010, 69, 1580–1588. [Google Scholar] [CrossRef]
- Arnett, F.C.; Edworthy, S.M.; Bloch, D.A.; McShane, D.J.; Fries, J.F.; Cooper, N.S.; Healey, L.A.; Kaplan, S.R.; Liang, M.H.; Luthra, H.S.; et al. The American Rheumatism Association 1987 revised criteria for the classification of rheumatoid arthritis. Arthritis Rheum. 1988, 31, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, T.; Kawakami, A.; Nakashima, T.; Yamasaki, S.; Tamai, M.; Tanaka, F.; Kamachi, M.; Ida, H.; Migita, K.; Origuchi, T.; et al. Osteoprotegerin (OPG) acts as an endogenous decoy receptor in tumour necrosis factor-related apoptosis-inducing ligand (TRAIL)-mediated apoptosis of fibroblast-like synovial cells. Clin. Exp. Immunol. 2004, 137, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Iwamoto, N.; Yamasaki, S.; Nishino, A.; Nakashima, Y.; Horai, Y.; Kawashiri, S.Y.; Ichinose, K.; Arima, K.; Tamai, M.; et al. Upregulation of Thrombospondin 1 Expression in Synovial Tissues and Plasma of Rheumatoid Arthritis: Role of Transforming Growth Factor-beta1 toward Fibroblast-like Synovial Cells. J. Rheumatol. 2015, 42, 943–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GMCSF forward | 5′-CATGATGGCCAGCCACTACAA-3′ |
| GMCSF reverse | 5′-ACTGGCTCCCAGCAGTCAAAG-3′ |
| CCL3 forward | 5′-CGGCAGATTCCACAGAATTTCATA-3′ |
| CCL3 reverse | 5′-AGATGACACCGGGCTTGGAG-3′ |
| MKI67 forward | 5′-CCATATGCCTGTGGAGTGGAA-3′ |
| MKI67 reverse | 5′-CCACCCTTAGCGTGCTCTTGA-3′ |
| GAPDH forward | 5′-GCACCGTCAAGGCTGAGAAC-3′ |
| GAPDH reverse | 5′-TGGTGAAGACGCCAGTGGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwamoto, N.; Furukawa, K.; Endo, Y.; Shimizu, T.; Sumiyoshi, R.; Umeda, M.; Koga, T.; Kawashiri, S.-y.; Igawa, T.; Ichinose, K.; et al. Methotrexate Alters the Expression of microRNA in Fibroblast-like Synovial Cells in Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 11561. https://doi.org/10.3390/ijms222111561
Iwamoto N, Furukawa K, Endo Y, Shimizu T, Sumiyoshi R, Umeda M, Koga T, Kawashiri S-y, Igawa T, Ichinose K, et al. Methotrexate Alters the Expression of microRNA in Fibroblast-like Synovial Cells in Rheumatoid Arthritis. International Journal of Molecular Sciences. 2021; 22(21):11561. https://doi.org/10.3390/ijms222111561
Chicago/Turabian StyleIwamoto, Naoki, Kaori Furukawa, Yushiro Endo, Toshimasa Shimizu, Remi Sumiyoshi, Masataka Umeda, Tomohiro Koga, Shin-ya Kawashiri, Takashi Igawa, Kunihiro Ichinose, and et al. 2021. "Methotrexate Alters the Expression of microRNA in Fibroblast-like Synovial Cells in Rheumatoid Arthritis" International Journal of Molecular Sciences 22, no. 21: 11561. https://doi.org/10.3390/ijms222111561