
Protective Effects of PACAP in a Rat Model of Diabetic Neuropathy

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
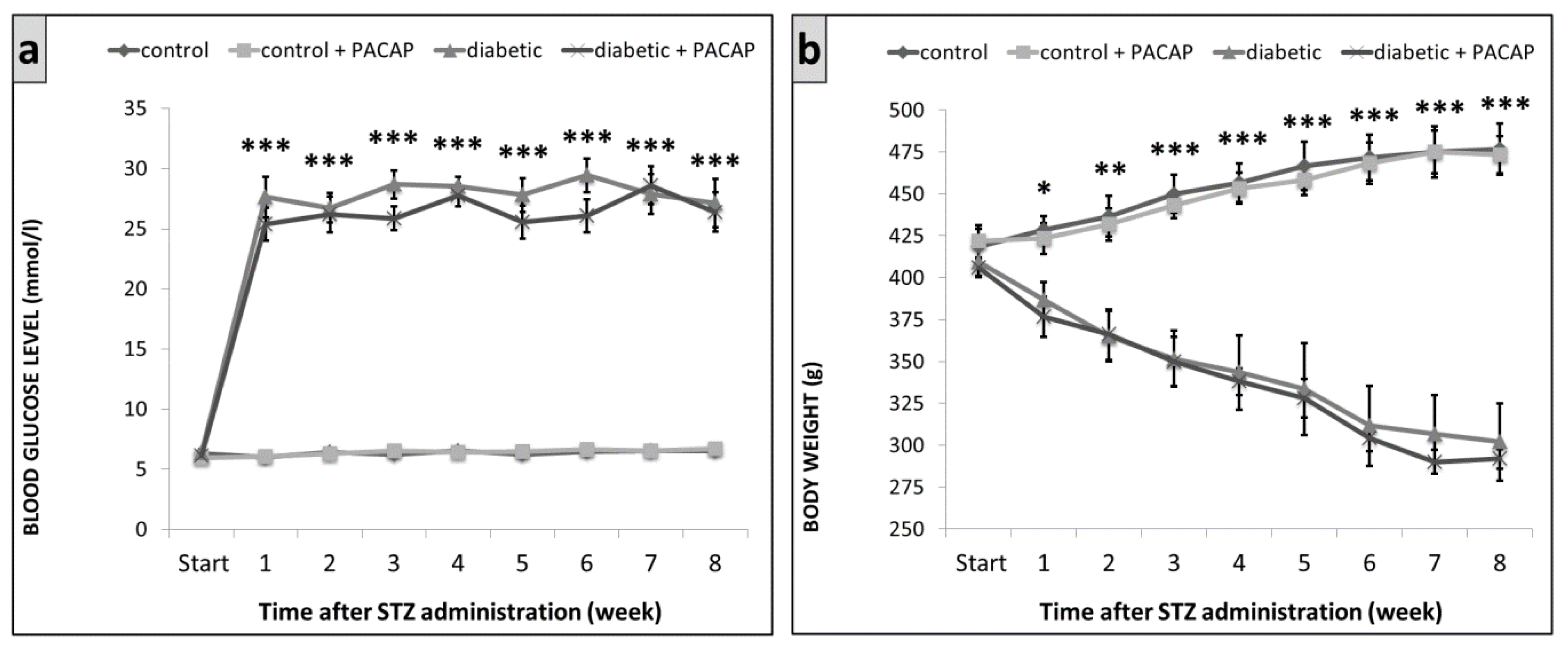
2.1. Blood Glucose Levels and Body Weight
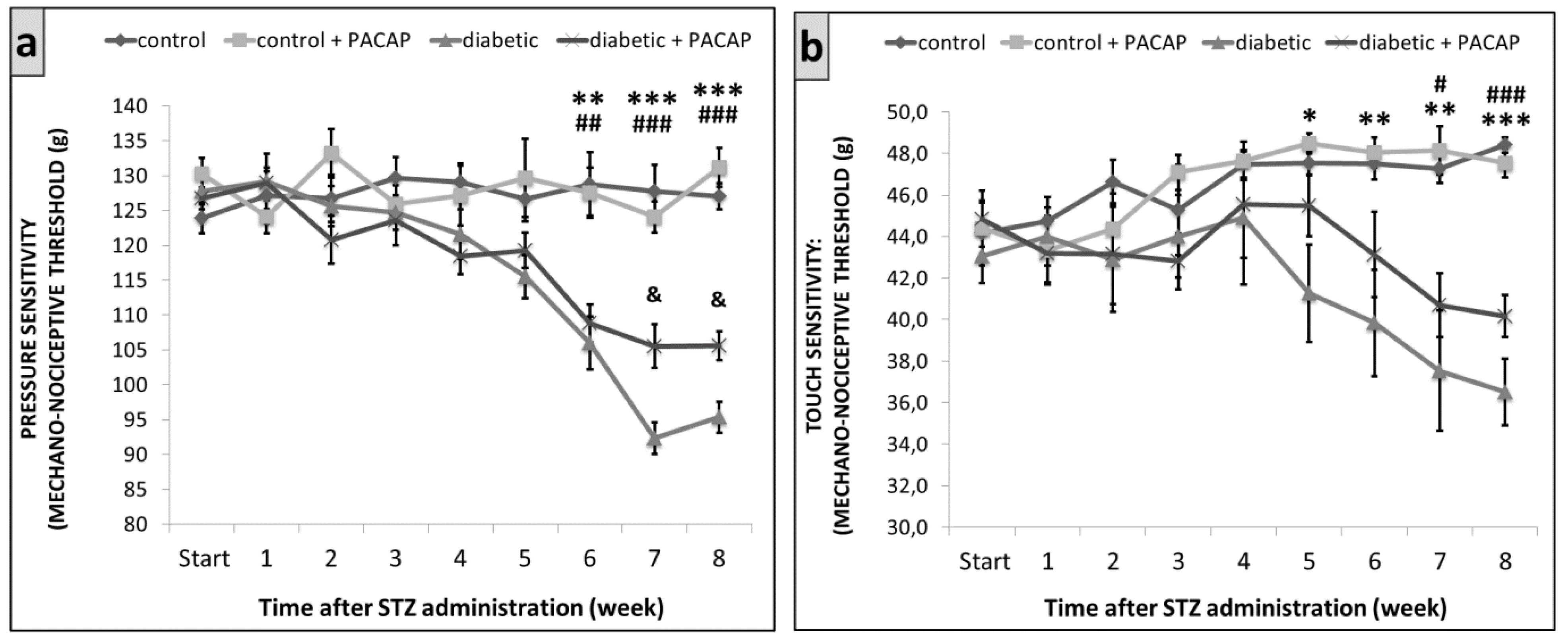
2.2. Functional Tests
2.2.1. Randall–Selitto Test
2.2.2. Dynamic Plantar Aesthesiometer (DPA)
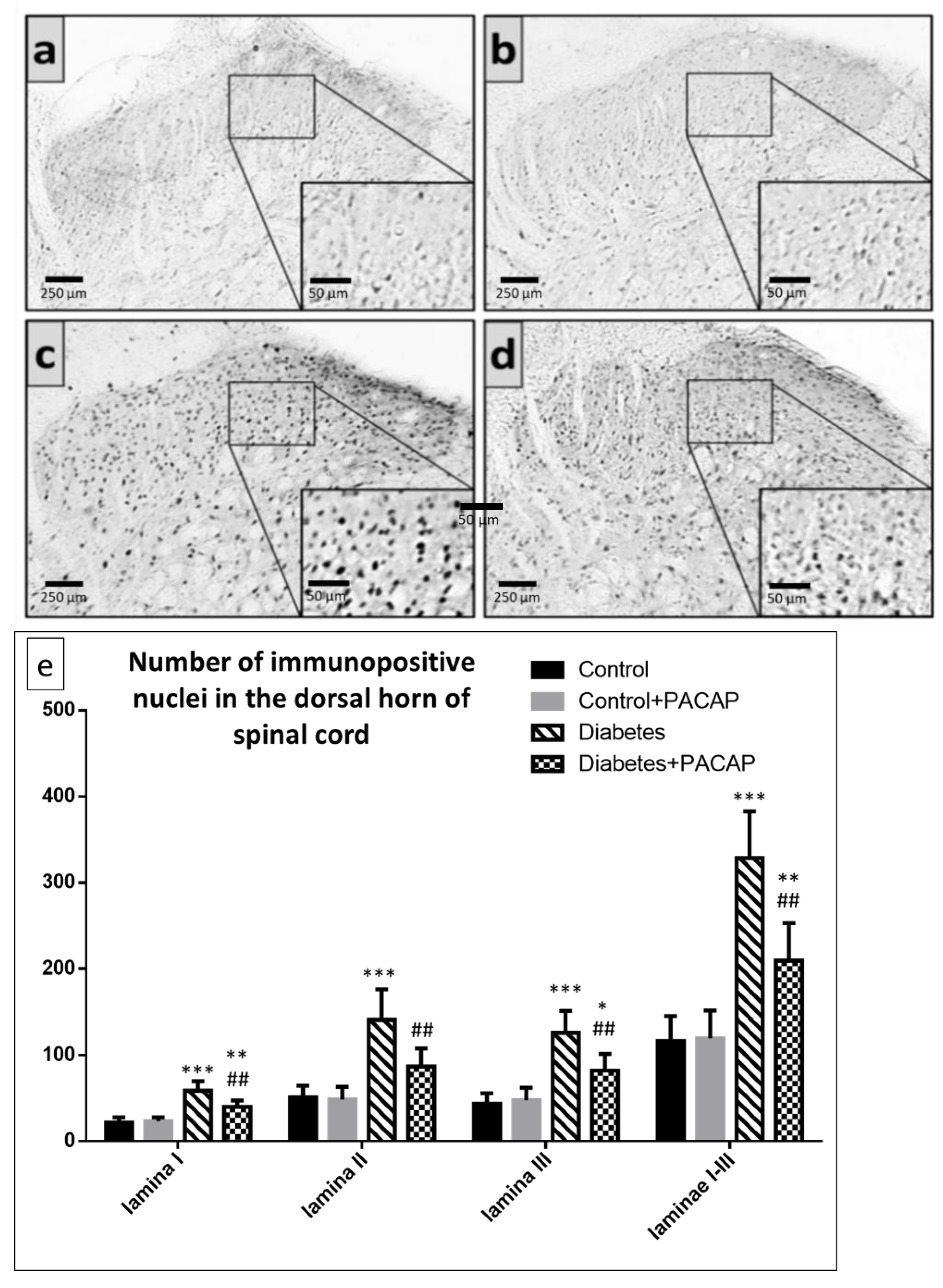
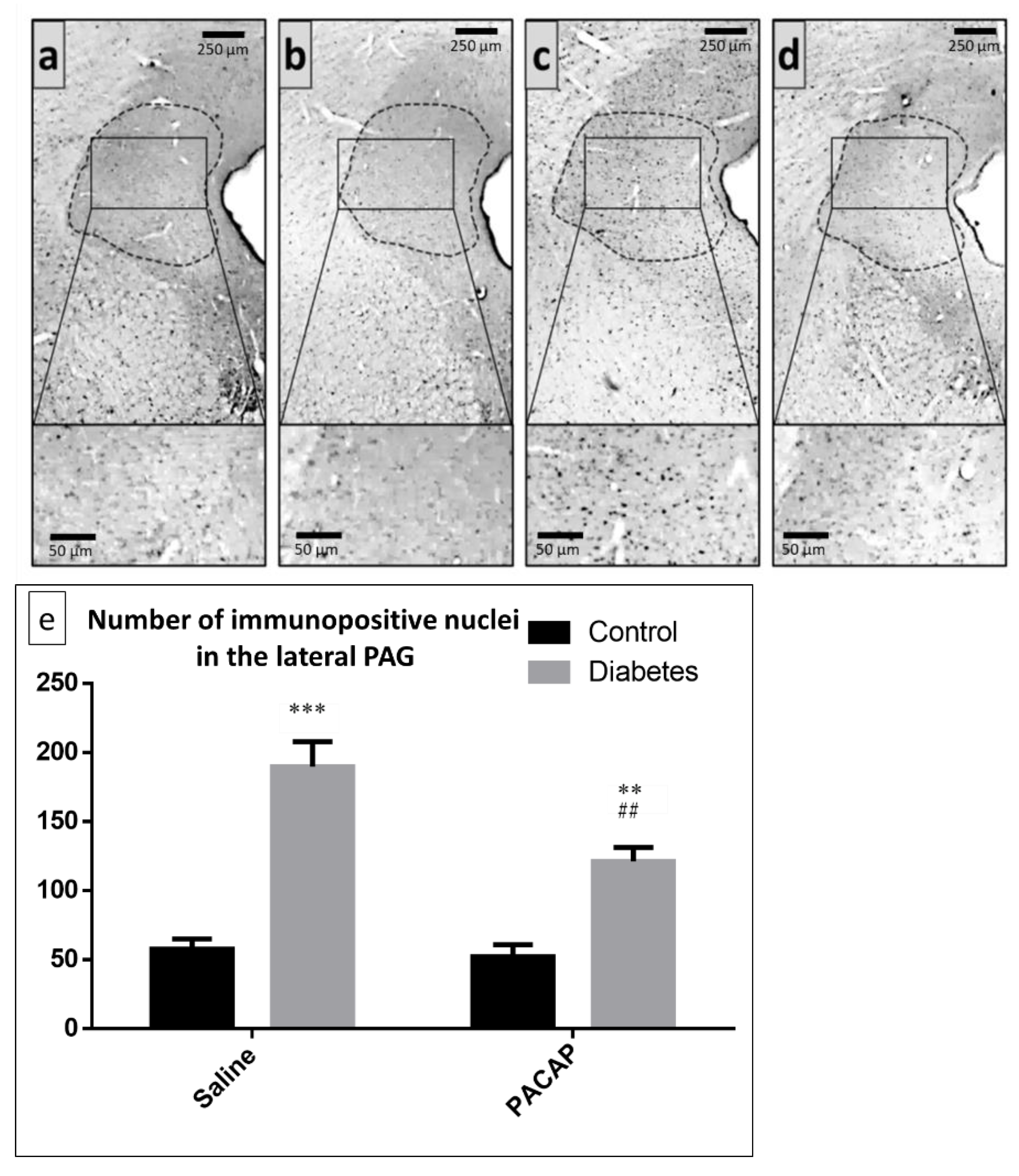
2.3. Immunohistochemistry
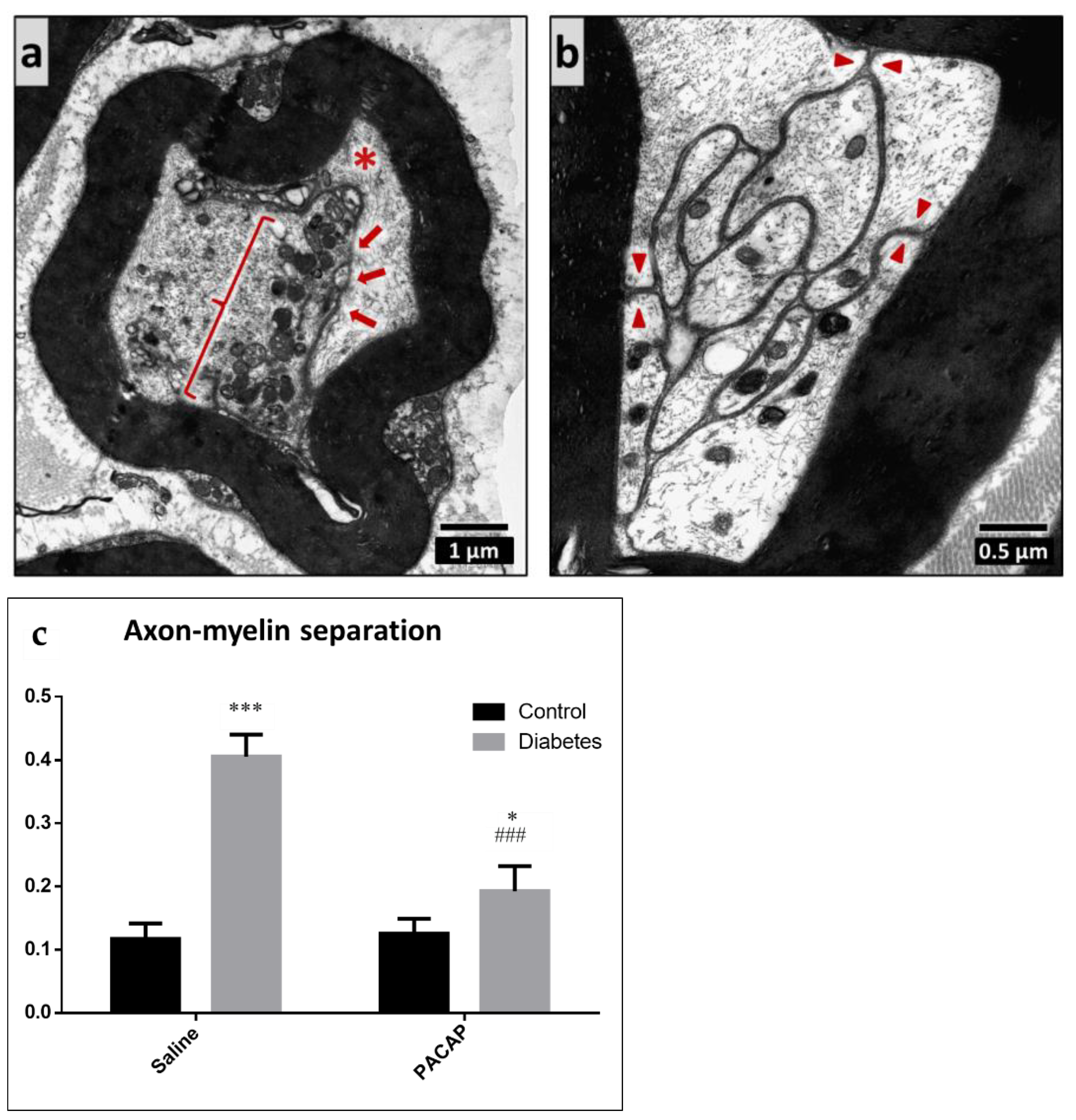
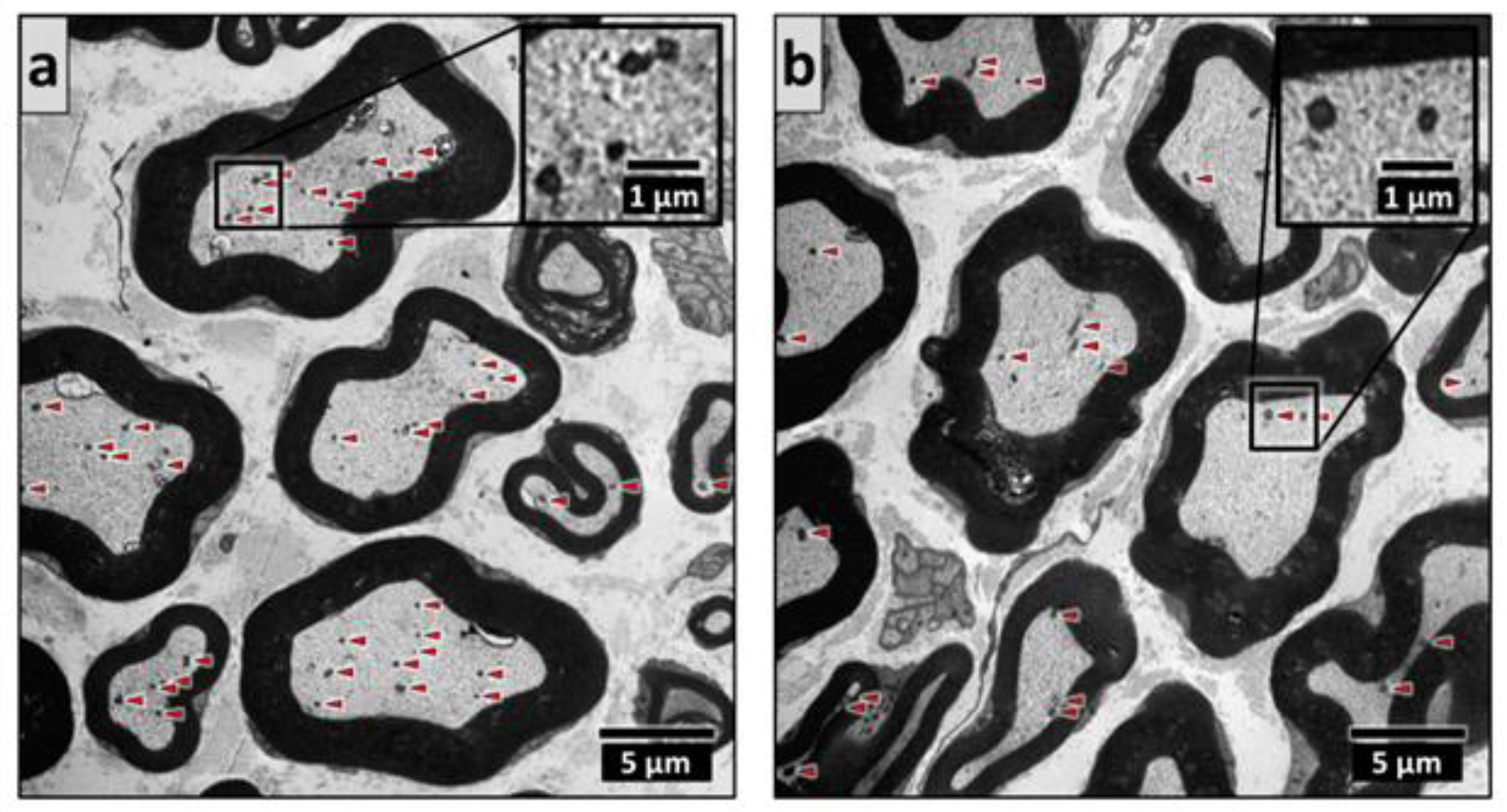
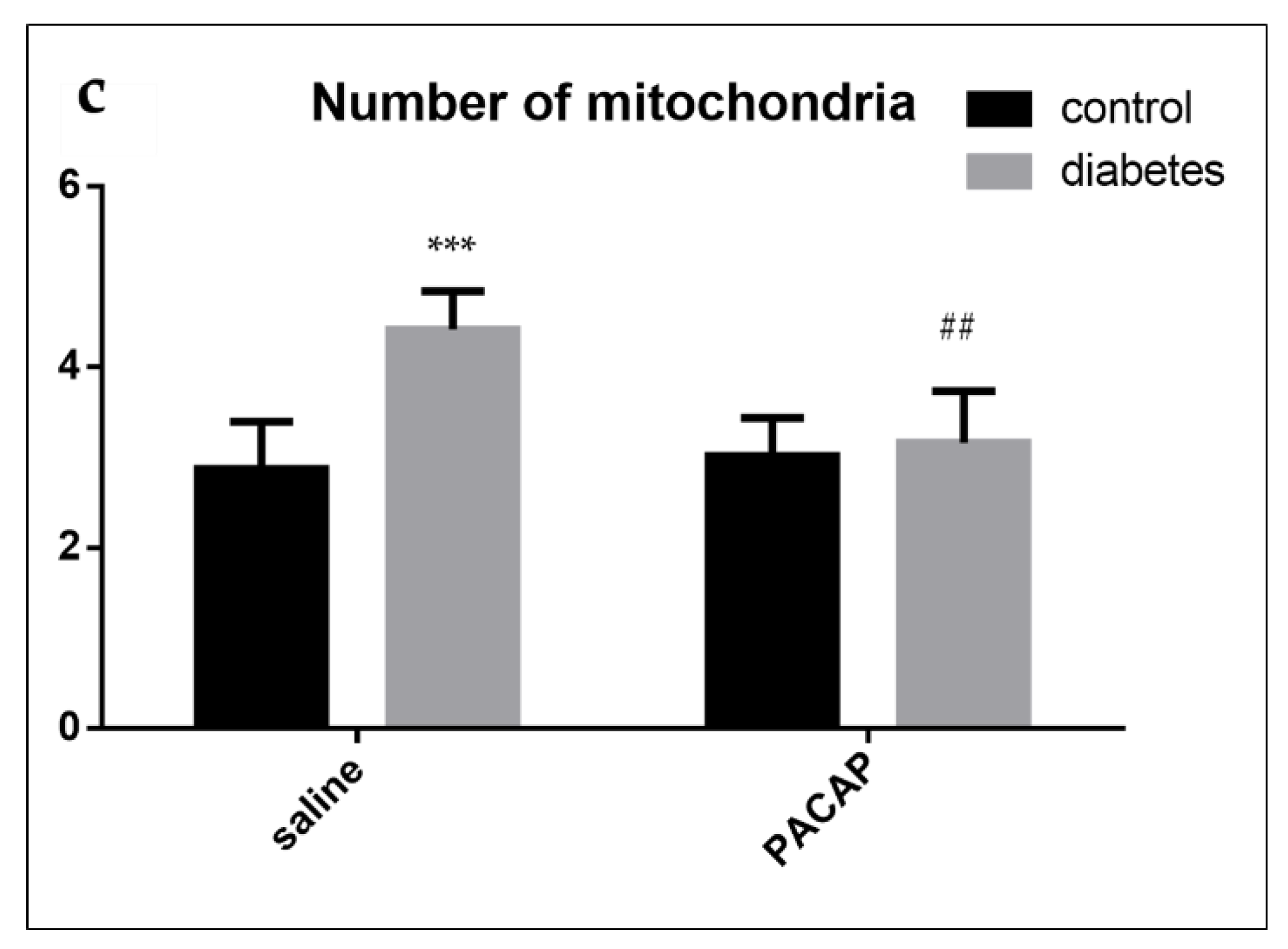
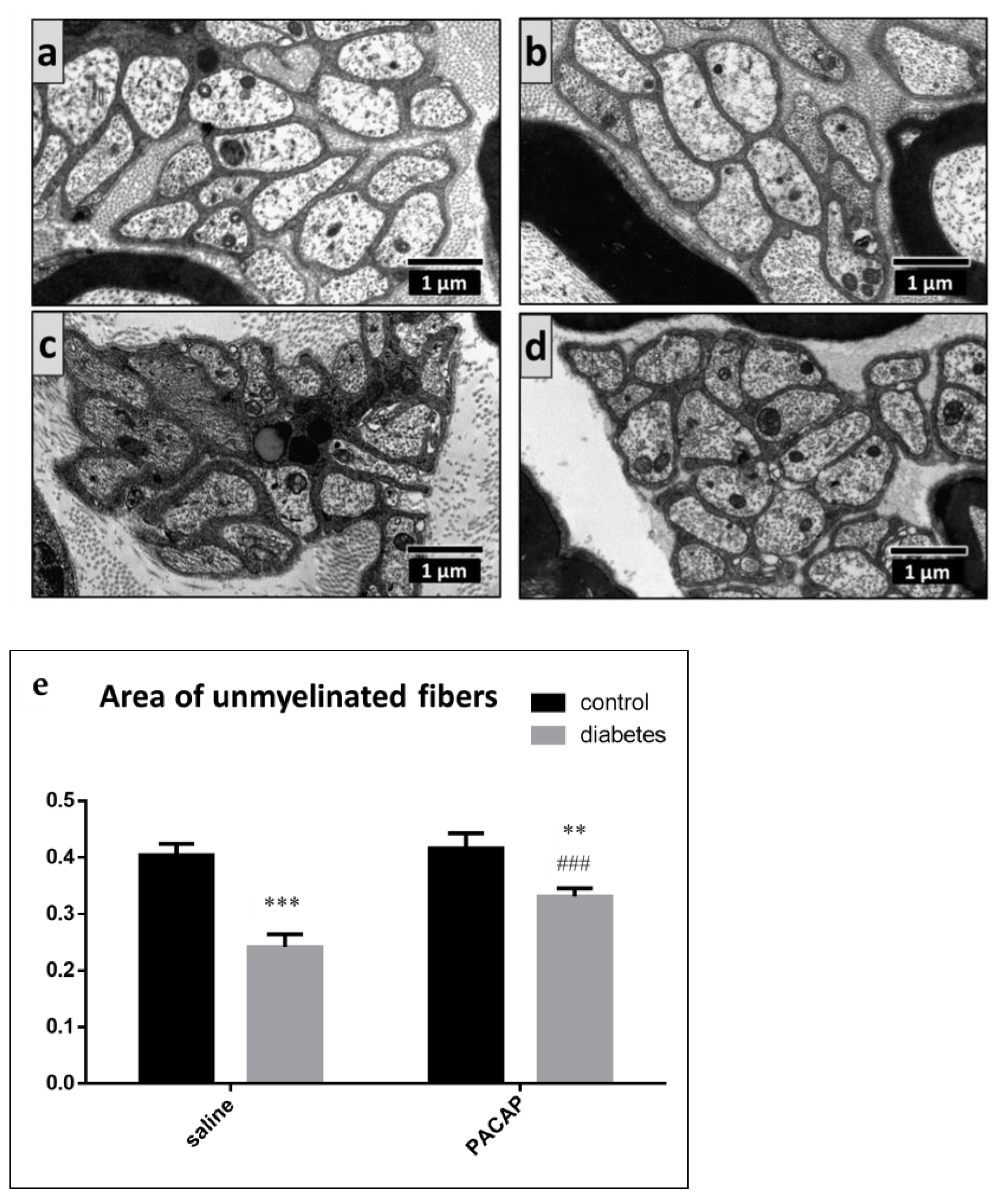
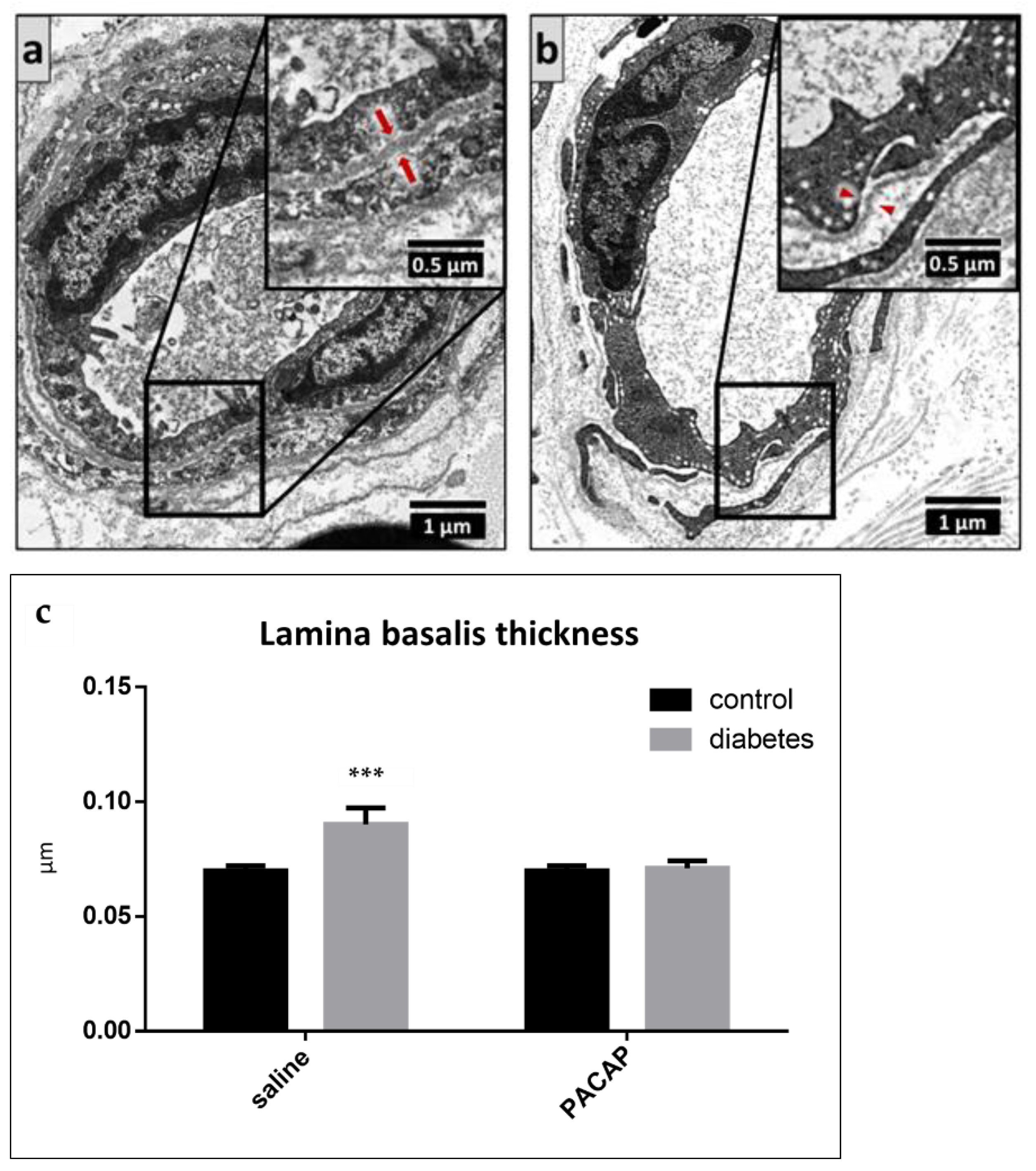
2.4. Electron Microscopy of the Sciatic Nerve
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Functional Tests
4.2.1. Mechanical Nociceptive Threshold—Randall–Selitto Test
4.2.2. Mechanical Nociceptive Threshold—Dynamic Plantar Aesthesiometer Test (DPA)
4.3. Histology
4.3.1. Tissue Collection and Preparation for Histology
4.3.2. Immunohistochemistry for FosB
4.3.3. Digital Imaging and Morphometry at Light Microscopic Level
4.3.4. Electron Microscopy
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vaudry, D.; Falluel-Morel, A.; Bourgault, S.; Basille, M.; Burel, D.; Wurtz, O.; Fournier, A.; Chow, B.K.C.; Hashimoto, H.; Galas, L.; et al. Pituitary adenylate cyclase-activating polypeptide and its receptors: 20 Years after the discovery. Pharmacol. Rev. 2009, 61, 283–357. [Google Scholar] [CrossRef]
- May, V.; Johnson, G.C.; Hammack, S.E.; Braas, K.M.; Parsons, R.L. PAC1 Receptor internalization and endosomal MEK/ERK activation is essential for PACAP-mediated neuronal excitability. J. Mol. Neurosci. 2021, 71, 1536–1542. [Google Scholar] [CrossRef]
- Messlinger, K.; Balcziak, L.K.; Russo, A.F. Cross-talk signaling in the trigeminal ganglion: Role of neuropeptides and other mediators. J. Neural Transm. 2020, 127, 431–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciranna, L.; Costa, L. Pituitary adenylate cyclase-activating polypeptide modulates hippocampal synaptic transmission and plasticity: New therapeutic suggestions for fragile X syndrome. Front. Cell. Neurosci. 2019, 13, 524. [Google Scholar] [CrossRef] [PubMed]
- Gastelum, C.; Perez, L.; Hernandez, J.; Le, N.; Vahrson, I.; Sayers, S.; Wagner, E.J. Adaptive changes in the central control of energy homeostasis occur in response to variations in energy status. Int. J. Mol. Sci. 2021, 22, 2728. [Google Scholar] [CrossRef] [PubMed]
- Vélez, E.J.; Unniappan, S. A comparative update on the neuroendocrine regulation of growth hormone in vertebrates. Front. Endocrinol. 2021, 11, 614981. [Google Scholar] [CrossRef]
- Toth, D.; Szabo, E.; Tamas, A.; Juhasz, T.; Horvath, G.; Fabian, E.; Opper, B.; Szabo, D.; Maugeri, G.; D’Amico, A.G.; et al. Protective effects of PACAP in peripheral organs. Front. Endocrinol. 2020, 11, 377. [Google Scholar] [CrossRef]
- Oride, A.; Kanasaki, H.; Mijiddorj, T.; Sukhbaatar, U.; Yamada, T.; Kyo, S. Expression and regulation of pituitary adenylate cyclase-activating polypeptide in rat placental cells. Reprod. Sci. 2016, 23, 1080–1086. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Spencer, N.J.; Brookes, S.J.H. The role of enteric inhibitory neurons in intestinal motility. Auton. Neurosci. 2021, 235, 102854. [Google Scholar] [CrossRef]
- Horvath, G.; Opper, B.; Reglodi, D. The neuropeptide pituitary adenylate cyclase-activating polypeptide (PACAP) is protective in inflammation and oxidative stress-induced damage in the kidney. Int. J. Mol. Sci. 2019, 20, 4944. [Google Scholar] [CrossRef] [Green Version]
- Szentleleky, E.; Szegeczki, V.; Karanyicz, E.; Hajdu, T.; Tamas, A.; Toth, G.; Zakany, R.; Reglodi, D.; Juhasz, T. Pituitary adenylate cyclase activating polypeptide (PACAP) reduces oxidative and mechanical stress-evoked matrix degradation in chondrifying cell cultures. Int. J. Mol. Sci. 2019, 20, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girard, B.M.; Campbell, S.E.; Beca, K.I.; Perkins, M.; Hsiang, H.; May, V.; Vizzard, M.A. Intrabladder PAC1 receptor antagonist, PACAP(6-38), reduces urinary bladder frequency and pelvic sensitivity in mice exposed to repeated variate stress (RVS). J. Mol. Neurosci. 2021, 71, 1575–1588. [Google Scholar] [CrossRef]
- Jozsa, G.; Fulop, B.D.; Kovacs, L.; Czibere, B.; Szegeczki, V.; Kiss, T.; Hajdu, T.; Tamas, A.; Helyes, Z.; Zakany, R.; et al. Lack of pituitary adenylate cyclase–activating polypeptide (PACAP) disturbs callus formation. J. Mol. Neurosci. 2021, 71, 1543–1555. [Google Scholar] [CrossRef] [Green Version]
- Rytel, L.; Wojtkiewicz, J.; Snarska, A.; Mikołajczyk, A. Changes in the neurochemical characterization of enteric neurons in the porcine duodenum after administration of low-dose salmonella enteritidis lipopolysaccharides. J. Mol. Neurosci. 2021, 71, 1556–1566. [Google Scholar] [CrossRef] [PubMed]
- Szabo, D.; Sarszegi, Z.; Polgar, B.; Saghy, E.; Nemeth, A.; Reglodi, D.; Makkos, A.; Gorbe, A.; Helyes, Z.; Ferdinandy, P.; et al. Pacap-38 in acute st-segment elevation myocardial infarction in humans and pigs: A translational study. Int. J. Mol. Sci. 2021, 22, 2883. [Google Scholar] [CrossRef]
- Chiba, Y.; Ueda, C.; Kohno, N.; Yamashita, M.; Miyakawa, Y.; Ando, Y.; Suto, W.; Hirabayashi, T.; Takenoya, F.; Takasaki, I.; et al. Attenuation of relaxing response induced by pituitary adenylate cyclase-activating polypeptide in bronchial smooth muscle of experimental asthma. Am. J. Physiol. Lung. Cell. Mol. Physiol. 2020, 319, L786–L793. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, L.; Xie, J.; Shi, L. The emerging role of neuropeptides in Parkinson’s disease. Front. Aging Neurosci. 2021, 13, 646726. [Google Scholar] [CrossRef]
- Reglodi, D.; Kiss, P.; Lubics, A.; Tamas, A. Review on the protective effects of PACAP in models of neurodegenerative diseases in vitro and in vivo. Curr. Pharm. Des. 2011, 17, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Kiss, P.; Szabadfi, K.; Atlasz, T.; Gabriel, R.; Horvath, G.; Szakaly, P.; Sandor, B.; Lubics, A.; Laszlo, E.; et al. PACAP is an endogenous protective factor-insights from PACAP-deficient mice. J. Mol. Neurosci. 2012, 48, 482–492. [Google Scholar] [CrossRef]
- Reglodi, D.; Tamas, A.; Jungling, A.; Vaczy, A.; Rivnyak, A.; Fulop, B.D.; Szabo, E.; Lubics, A.; Atlasz, T. Protective effects of pituitary adenylate cyclase activating polypeptide against neurotoxic agents. Neurotoxicology 2018, 66, 185–194. [Google Scholar] [CrossRef]
- Shioda, S.; Takenoya, F.; Wada, N.; Hirabayashi, T.; Seki, T.; Nakamachi, T. Pleiotropic and retinoprotective functions of PACAP. Anat. Sci. Int. 2016, 91, 313–324. [Google Scholar] [CrossRef]
- Lee, E.H.; Seo, S.R. Neuroprotective roles of pituitary adenylate cyclase-activating polypeptide in neurodegenerative diseases. BMB Rep. 2014, 47, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Rojas, V.A.; Jiménez-Garduño, A.M.; Michelatti, D.; Tosatto, L.; Marchioretto, M.; Arosio, D.; Basso, M.; Pennuto, M.; Musio, C. ClC-2-like chloride current alterations in a cell model of spinal and bulbar muscular atrophy, a polyglutamine disease. J. Mol. Neurosci. 2021, 71, 662–674. [Google Scholar] [CrossRef]
- D'Amico, A.G.; Maugeri, G.; Saccone, S.; Federico, C.; Cavallaro, S.; Reglodi, D.; D'Agata, V. PACAP modulates the autophagy process in an in vitro model of amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2020, 21, 2943. [Google Scholar] [CrossRef]
- Perényi, H.; Szegeczki, V.; Horvath, G.; Hinnah, B.; Tamás, A.; Radak, Z.; Abraham, D.; Zakany, R.; Reglodi, D.; Juhasz, T. Physical activity protects the pathological alterations of Alzheimer’s disease kidneys via the activation of PACAP and BMP signaling pathways. Front. Cell. Neurosci. 2020, 14, 243. [Google Scholar] [CrossRef] [PubMed]
- Schaler, A.W.; Runyan, A.M.; Clelland, C.L.; Sydney, E.J.; Fowler, S.L.; Figueroa, H.Y.; Shioda, S.; Santa-Maria, I.; Duff, K.E.; Myeku, N. PAC1 receptor-mediated clearance of tau in postsynaptic compartments attenuates tau pathology in mouse brain. Sci. Transl. Med. 2021, 13, eaba7394. [Google Scholar] [CrossRef]
- Chen, X.Y.; Du, Y.F.; Chen, L. Neuropeptides exert neuroprotective effects in Alzheimer’s disease. Front. Mol. Neurosci. 2019, 11, 493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonaka, N.; Banks, W.A.; Shioda, S. Pituitary adenylate cyclase-activating polypeptide: Protective effects in stroke and dementia. Peptides 2020, 130, 170332. [Google Scholar] [CrossRef]
- Cherait, A.; Maucotel, J.; Lefranc, B.; Leprince, J.; Vaudry, D. Intranasal administration of PACAP is an efficient delivery route to reduce infarct volume and promote functional recovery after transient and permanent middle cerebral artery occlusion. Front. Endocrinol. 2021, 11, 585082. [Google Scholar] [CrossRef] [PubMed]
- Reglodi, D.; Tamas, A.; Somogyvari-Vigh, A.; Szanto, Z.; Kertes, E.; Lenard, L.; Arimura, A.; Lengvari, I. Effects of pretreatment with PACAP on the infarct size and functional outcome in rat permanent focal cerebral ischemia. Peptides 2002, 23, 2227–2234. [Google Scholar] [CrossRef]
- Jungling, A.; Reglodi, D.; Maasz, G.; Zrinyi, Z.; Schmidt, J.; Rivnyak, A.; Horvath, G.; Pirger, Z.; Tamas, A. Alterations of nigral dopamine levels in Parkinson’s disease after environmental enrichment and PACAP treatment in aging rats. Life 2021, 11, 35. [Google Scholar] [CrossRef]
- Solés-Tarrés, I.; Cabezas-Llobet, N.; Vaudry, D.; Xifró, X. Protective effects of pituitary adenylate cyclase-activating polypeptide and vasoactive intestinal peptide against cognitive decline in neurodegenerative diseases. Front. Cell. Neurosci. 2020, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Lamine-Ajili, A.; Fahmy, A.M.; Létourneau, M.; Chatenet, D.; Labonté, P.; Vaudry, D.; Fournier, A. Effect of the pituitary adenylate cyclase-activating polypeptide on the autophagic activation observed in in vitro and in vivo models of Parkinson’s disease. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Tamas, A.; Lubics, A.; Lengvari, I.; Reglodi, D. Protective effects of PACAP in excitotoxic striatal lesion. Ann. N. Y. Acad. Sci. 2006, 1070, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Kvarik, T.; Reglodi, D.; Werling, D.; Vaczy, A.; Kovari, P.; Szabo, E.; Kovacs, K.; Hashimoto, H.; Ertl, T.; Gyarmati, J.; et al. The protective effects of endogenous PACAP in oxygen-induced retinopathy. J. Mol. Neurosci. 2021, in press. [Google Scholar] [CrossRef]
- Kovacs, A.K.; Atlasz, T.; Werling, D.; Szabo, E.; Reglodi, D.; Toth, G.K. Stability test of PACAP in eye drops. J. Mol. Neurosci. 2021, 71, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Ye, D.; Shi, Y.; Xu, Y.; Huang, J. PACAP attenuates optic nerve crush-induced retinal ganglion cell apoptosis via activation of the CREB-Bcl-2 pathway. J. Mol. Neurosci. 2019, 68, 475–484. [Google Scholar] [CrossRef]
- Banki, E.; Degrell, P.; Kiss, P.; Kovacs, K.; Kemeny, A.; Csanaky, K.; Duh, A.; Nagy, D.; Toth, G.; Tamas, A.; et al. Effect of PACAP treatment on kidney morphology and cytokine expression in rat diabetic nephropathy. Peptides 2013, 42, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Banki, E.; Kovacs, K.; Nagy, D.; Juhasz, T.; Degrell, P.; Csanaky, K.; Kiss, P.; Jancso, G.; Toth, G.; Tamas, A.; et al. Molecular mechanisms underlying the nephroprotective effects of PACAP in diabetes. J. Mol. Neurosci. 2014, 54, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Szabadfi, K.; Atlasz, T.; Kiss, P.; Danyadi, B.; Tamas, A.; Helyes, Z.; Hashimoto, H.; Shintani, N.; Baba, A.; Toth, G.; et al. Mice deficient in pituitary adenylate cyclase activating polypeptide (PACAP) are more susceptible to retinal ischemic injury in vivo. Neurotox. Res. 2012, 21, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Szabadfi, K.; Szabo, A.; Kiss, P.; Reglodi, D.; Setalo, G., Jr.; Kovacs, K.; Tamas, A.; Toth, G.; Gabriel, R. PACAP promotes neuron survival in early experimental diabetic retinopathy. Neurochem. Int. 2014, 64, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Szabadfi, K.; Reglodi, D.; Szabo, A.; Szalontai, B.; Valasek, A.; Setalo, G.; Kiss, P.; Tamas, A.; Wilhelm, M.; Gabriel, R. Pituitary adenylate cyclase activating polypeptide, a potential therapeutic agent for diabetic retinopathy in rats: Focus on the vertical information processing pathway. Neurotox. Res. 2016, 29, 432–446. [Google Scholar] [CrossRef]
- D’Amico, A.G.; Maugeri, G.; Reitano, R.; Bucolo, C.; Saccone, S.; Drago, F.; D’Agata, V. PACAP modulates expression of hypoxia-inducible factors in streptozotocin-induced diabetic rat retina. J. Mol. Neurosci. 2015, 57, 501–509. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Rasà, D.M.; Bucolo, C.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. Modulation of IL-1β and VEGF expression in rat diabetic retinopathy after PACAP administration. Peptides 2017, 97, 64–69. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, A.G.; Maugeri, G.; Musumeci, G.; Reglodi, D.; D’Agata, V. PACAP and NAP: Effect of two functionally related peptides in diabetic retinopathy. J. Mol. Neurosci. 2021, 71, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Saccone, S.; Federico, C.; Cavallaro, S.; D’Agata, V. PACAP and VIP inhibit HIF-1α-mediated VEGF expression in a model of diabetic macular edema. J. Cell. Physiol. 2017, 232, 1209–1215. [Google Scholar] [CrossRef]
- Maugeri, G.; D’Amico, A.G.; Bucolo, C.; D’Agata, V. Protective effect of PACAP-38 on retinal pigmented epithelium in an in vitro and in vivo model of diabetic retinopathy through EGFR-dependent mechanism. Peptides 2019, 119, 170108. [Google Scholar] [CrossRef]
- Scuderi, S.; D’Amico, A.G.; Castorina, A.; Imbesi, R.; Carnazza, M.L.; D’Agata, V. Ameliorative effect of PACAP and VIP against increased permeability in a model of outer blood retinal barrier dysfunction. Peptides 2013, 39, 119–124. [Google Scholar] [CrossRef]
- Fabian, E.; Horvath, G.; Opper, B.; Atlasz, T.; Toth, G.; Reglodi, D. PACAP is protective against cellular stress in retinal pigment epithelial cells. Int. J. Pept. Res. Ther. 2021, 27, 1221–1228. [Google Scholar] [CrossRef]
- Solymar, M.; Ivic, I.; Balasko, M.; Fulop, B.D.; Toth, G.; Tamas, A.; Reman, G.; Koller, A.; Reglodi, D. Pituitary adenylate cyclase-activating polypeptide ameliorates vascular dysfunction induced by hyperglycaemia. Diabetes Vasc. Dis. Res. 2018, 15, 277–285. [Google Scholar] [CrossRef]
- Feldman, E.L.; Russell, J.W.; Sullivan, K.A.; Golovoy, D. New insights into the pathogenesis of diabetic neuropathy. Curr. Opin. Neurol. 1999, 12, 553–563. [Google Scholar] [CrossRef]
- Zochodne, D.W. Is early diabetic neuropathy a disorder of the dorsal root ganglion? A hypothesis and critique of some current ideas on the etiology of diabetic neuropathy. J. Periph. Nerv. Syst. 1996, 1, 119–130. [Google Scholar]
- Sanlioglu, A.D.; Karacay, B.; Balci, M.K.; Griffith, T.S.; Sanlioglu, S. Therapeutic potential of VIP vs PACAP in diabetes. J. Mol. Endocrinol. 2012, 49, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Yang, X.; Bai, T.; Liu, Z.; Liu, T.; Wang, Y.; Cui, L.; Liu, Y.; Zhang, Y. PACAP stimulates insulin secretion by PAC1 receptor and ion channels in β-cells. Cell Signal. 2019, 61, 48–56. [Google Scholar] [CrossRef]
- Shao, S.; Yang, Y.; Yuan, G.; Zhang, M.; Yu, X. Signaling molecules involved in lipid-induced pancreatic beta-cell dysfunction. DNA Cell Biol. 2013, 32, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Nakata, M.; Shintani, N.; Hashimoto, H.; Baba, A.; Yada, T. Intra-islet PACAP protects pancreatic β-cells against glucotoxicity and lipotoxicity. J. Mol. Neurosci. 2010, 42, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, M.; D’Amico, A.G.; Musumeci, G.; Reglodi, D.; D’Agata, V. Effects of PACAP on Schwann cells: Focus on nerve injury. Int. J. Mol. Sci. 2020, 21, 8233. [Google Scholar] [CrossRef] [PubMed]
- Castorina, A.; Scuderi, S.; D’Amico, A.G.; Drago, F.; D’Agata, V. PACAP and VIP increase the expression of myelin-related proteins in rat schwannoma cells: Involvement of PAC1/VPAC2 receptor-mediated activation of PI3K/Akt signaling pathways. Exp. Cell Res. 2014, 322, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.M.; Chaverra, M.; Hapner, S.J.; Nelson, B.R.; Todd, V.; Zigmond, R.E.; Lefcort, F. PACAP promotes sensory neuron differentiation: Blockade by neurotrophic factors. Mol. Cell. Neurosci. 2004, 25, 629–641. [Google Scholar] [CrossRef]
- Fukiage, C.; Nakajima, T.; Takayama, Y.; Minagawa, Y.; Shearer, T.R.; Azuma, M. PACAP induces neurite outgrowth in cultured trigeminal ganglion cells and recovery of corneal sensitivity after flap surgery in rabbits. Am. J. Ophthalmol. 2007, 143, 255–262. [Google Scholar] [CrossRef]
- Armstrong, B.D.; Abad, C.; Chhith, S.; Cheung-Lau, G.; Hajji, O.E.; Nobuta, H.; Waschek, J.A. Impaired nerve regeneration and enhanced neuroinflammatory response in mice lacking pituitary adenylyl cyclase activating peptide. Neuroscience 2008, 151, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Baskozos, G.; Sandy-Hindmarch, O.; Clark, A.J.; Windsor, K.; Karlsson, P.; Weir, G.A.; McDermott, L.A.; Burchall, J.; Wiberg, A.; Furniss, D.; et al. Molecular and cellular correlates of human nerve regeneration: ADCYAP1/PACAP enhance nerve outgrowth. Brain 2020, 143, 2009–2026. [Google Scholar] [CrossRef]
- Wang, Z.; Shan, W.; Li, H.; Feng, J.; Lu, S.; Ou, B.; Ma, M.; Ma, Y. The PACAP-derived peptide MPAPO facilitates corneal wound healing by promoting corneal epithelial cell proliferation and trigeminal ganglion cell axon regeneration. Int. J. Biol. Sci. 2019, 15, 2676–2691. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, Y.; Guo, M.; Dong, X.; Liao, M.; Du, M.; Wang, X.; Yin, H.; Yan, H. Exosome-mediated delivery of the neuroprotective peptide PACAP38 promotes retinal ganglion cell survival and axon regeneration in rats with traumatic optic neuropathy. Front. Cell Dev. Biol. 2021, 9, 659783. [Google Scholar] [CrossRef] [PubMed]
- Harati, Y. Diabetic neuropathies: Unanswered questions. Neurol. Clin. 2007, 25, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 414, 813–820. [Google Scholar] [CrossRef]
- Duby, J.J.; Campbell, R.K.; Setter, S.M.; White, J.R.; Rasmussen, K.A. Diabetic neuropathy: An intensive review. Am. J. Heal. Pharm. 2004, 61, 160–176. [Google Scholar] [CrossRef]
- Conte, M.; Sabbatinelli, J.; Chiariello, A.; Martucci, M.; Santoro, A.; Monti, D.; Arcaro, M.; Galimberti, D.; Scarpini, E.; Bonfigli, A.R.; et al. Disease-specific plasma levels of mitokines FGF21, GDF15, and Humanin in type II diabetes and Alzheimer’s disease in comparison with healthy aging. GeroScience 2021, 43, 985–1001. [Google Scholar] [CrossRef]
- Yu, Y.; Singh, H.; Kwon, K.; Tsitrin, T.; Petrini, J.; Nelson, K.E.; Pieper, R. Protein signatures from blood plasma and urine suggest changes in vascular function and IL-12 signaling in elderly with a history of chronic diseases compared with an age-matched healthy cohort. GeroScience 2021, 43, 593–606. [Google Scholar] [CrossRef]
- Greene, D.A.; Feldman, E.L.; Stevens, M.J.; Sima, A.A.F.; Albers, J.W.; Pfeifer, M.A. Diabetic Neuropathy. In Ellenberg and Rifkin Diabetes Mellitus; Rifkin, H., Porte, D., Sherwin, R., Eds.; Appleton & Lange: Stamford, CT, USA, 1997; pp. 1009–1076. [Google Scholar]
- Morgado, C.; Terra, P.P.; Tavares, I. Neuronal hyperactivity at the spinal cord and periaqueductal grey during painful diabetic neuropathy: Effects of gabapentin. Eur. J. Pain 2010, 14, 693–699. [Google Scholar] [CrossRef]
- Johnson, M.S.; Ryals, J.M.; Wright, D.E. Diabetes-induced chemogenic hypoalgesia is paralleled by attenuated stimulus-induced Fos expression in the spinal cord of diabetic mice. J. Pain 2007, 8, 637–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgado, C.; Tavares, I. Diabetes C-fos expression at the spinal dorsal horn of streptozotocin-induced diabetic rats. Diabetes Metab. Res. Rev. 2007, 23, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Raposo, D.; Morgado, C.; Pereira-Terra, P.; Tavares, I. Nociceptive spinal cord neurons of laminae I-III exhibit oxidative stress damage during diabetic neuropathy which is prevented by early antioxidant treatment with epigallocatechin-gallate (EGCG). Brain Res. Bull. 2015, 110, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Lantéri-Minet, M.; de Pommery, J.; Herdegen, T.; Weil-Fugazza, J.; Bravo, R.; Menétrey, D. Differential time course and spatial expression of Fos, Jun, and Krox-24 proteins in spinal cord of rats undergoing subacute or chronic somatic inflammation. J. Comp. Neurol. 1993, 333, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Herdegen, T.; Tölle, T.R.; Bravo, R.; Zieglgänsberger, W.; Zimmermann, M. Sequential expression of JUN B, JUN D and FOS B proteins in rat spinal neurons: Cascade of transcriptional operations during nociception. Neurosci. Lett. 1991, 129, 221–224. [Google Scholar] [CrossRef]
- Madsen, T.M.; Bolwig, T.G.; Mikkelsen, J.D. Differential regulation of c-fos and fosB in the rat brain after amygdala kindling. Cell. Mol. Neurobiol. 2006, 26, 87–100. [Google Scholar] [CrossRef]
- Motojima, Y.; Ueta, Y.; Sakai, A. Analysis of the proportion and neuronal activity of excitatory and inhibitory neurons in the rat dorsal spinal cord after peripheral nerve injury. Neurosci. Lett. 2021, 749, 135707. [Google Scholar] [CrossRef] [PubMed]
- Aubert, N.; Vaudry, D.; Falluel-Morel, A.; Desfeux, A.; Fisch, C.; Ancian, P.; de Jouffrey, S.; Le Bigot, J.F.; Couvineau, A.; Laburthe, M.; et al. PACAP prevents toxicity induced by cisplatin in rat and primate neurons but not in proliferating ovary cells: Involvement of the mitochondrial apoptotic pathway. Neurobiol. Dis. 2008, 32, 66–80. [Google Scholar] [CrossRef]
- Lennertz, R.C.; Medler, K.A.; Bain, J.L.; Wright, D.E.; Stucky, C.L. Impaired sensory nerve function and axon morphology in mice with diabetic neuropathy. J. Neurophysiol. 2011, 106, 905–914. [Google Scholar] [CrossRef]
- Love, S.; Duchen, L.W. Morphological Abnormalities in Myelinated nerve fibres caused by Leiurus, Centruroides and Phoneutria venoms and their prevention by tetrodotoxin. Q. J. Exp. Physiol. 1986, 71, 115–122. [Google Scholar] [CrossRef]
- Low, P.A.; Dyck, P.J.; Schmelzer, J.D. Mammalian peripheral nerve sheath has unique responses to chronic elevations of endoneurial fluid pressure. Exp. Neurol. 1980, 70, 300–306. [Google Scholar] [CrossRef]
- Powell, H.C.; Myers, R.R. Axonopathy and microangiopathy in chronic alloxan diabetes. Acta Neuropathol. 1984, 65, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Vincze, A.; Reglodi, D.; Helyes, Z.; Hashimoto, H.; Shintani, N.; Abrahám, H. Role of endogenous pituitary adenylate cyclase activating polypeptide (PACAP) in myelination of the rodent brain: Lessons from PACAP-deficient mice. Int. J. Dev. Neurosci. 2011, 29, 923–935. [Google Scholar] [CrossRef]
- Castorina, A.; Tiralongo, A.; Giunta, S.; Carnazza, M.L.; Rasi, G.; D’Agata, V. PACAP and VIP prevent apoptosis in schwannoma cells. Brain Res. 2008, 1241, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Woodley, P.K.; Min, Q.; Li, Y.; Mulvey, N.F.; Parkinson, D.B.; Dun, X.P. Distinct VIP and PACAP functions in the distal nerve stump during peripheral nerve regeneration. Front. Neurosci. 2019, 13, 1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musumeci, G.; Leggio, G.M.; Marzagalli, R.; Al-Badri, G.; Drago, F.; Castorina, A. Identification of dysregulated microRNA networks in schwann cell-like cultures exposed to immune challenge: Potential crosstalk with the protective VIP/PACAP neuropeptide system. Int. J. Mol. Sci. 2018, 19, 981. [Google Scholar] [CrossRef] [Green Version]
- Edwards, J.L.; Quattrini, A.; Lentz, S.I.; Figueroa-Romero, C.; Cerri, F.; Backus, C.; Hong, Y.; Feldman, E.L. Diabetes regulates mitochondrial biogenesis and fission in mouse neurons. Diabetologia 2010, 53, 160–169. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.M.; Edwards, J.L.; McLean, L.L.; Hong, Y.; Cerri, F.; Lopez, I.; Quattrini, A.; Feldman, E.L. Mitochondrial biogenesis and fission in axons in cell culture and animal models of diabetic neuropathy. Acta Neuropathol. 2010, 120, 477–489. [Google Scholar] [CrossRef]
- Galloway, C.A.; Lee, H.; Nejjar, S.; Jhun, B.S.; Yu, T.; Hsu, W.; Yoon, Y. Transgenic control of mitochondrial fission induces mitochondrial uncoupling and relieves diabetic oxidative stress. Diabetes 2012, 61, 2093–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtaki, H.; Satoh, A.; Nakamachi, T.; Yofu, S.; Dohi, K.; Mori, H.; Ohara, K.; Miyamoto, K.; Hashimoto, H.; Shintani, N.; et al. Regulation of oxidative stress by pituitary adenylate cyclase-activating polypeptide (PACAP) mediated by PACAP receptor. J. Mol. Neurosci. 2010, 42, 397–403. [Google Scholar] [CrossRef]
- Botia, B.; Seyer, D.; Ravni, A.; Bénard, M.; Falluel-Morel, A.; Cosette, P.; Jouenne, T.; Fournier, A.; Vaudry, H.; Gonzalez, B.J.; et al. Peroxiredoxin 2 is involved in the neuroprotective effects of PACAP in cultured cerebellar granule neurons. J. Mol. Neurosci. 2008, 36, 61–72. [Google Scholar] [CrossRef]
- Kinhult, J.; Uddman, R.; Cardell, L.O. The induction of carbon monoxide-mediated airway relaxation by PACAP 38 in isolated guinea pig airways. Lung 2001, 179, 1–8. [Google Scholar] [CrossRef]
- Laszlo, E.; Juhasz, T.; Varga, A.; Czibere, B.; Kovacs, K.; Degrell, P.; Horvath, G.; Jancso, G.; Szakaly, P.; Tamas, A.; et al. Protective Effect of PACAP on Ischemia/Reperfusion-Induced Kidney Injury of Male and Female Rats: Gender Differences. J. Mol. Neurosci. 2019, 68, 408–419. [Google Scholar] [CrossRef]
- Masmoudi-Kouki, O.; Douiri, S.; Hamdi, Y.; Kaddour, H.; Bahdoudi, S.; Vaudry, D.; Basille, M.; Leprince, J.; Fournier, A.; Vaudry, H.; et al. Pituitary adenylate cyclase-activating polypeptide protects astroglial cells against oxidative stress-induced apoptosis. J. Neurochem. 2011, 117, 403–411. [Google Scholar] [CrossRef]
- Murakami, T.; Iwanaga, T.; Ogawa, Y.; Fujita, Y.; Sato, E.; Yoshitomi, H.; Sunada, Y.; Nakamura, A. Development of sensory neuropathy in streptozotocin-induced diabetic mice. Brain Behav. 2013, 3, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Tamas, A.; Reglodi, D.; Farkas, O.; Kovesdi, E.; Pal, J.; Povlishock, J.T.; Schwarcz, A.; Czeiter, E.; Szanto, Z.; Doczi, T.; et al. Effect of PACAP in central and peripheral nerve injuries. Int. J. Mol. Sci. 2012, 13, 8430–8448. [Google Scholar] [CrossRef] [Green Version]
- Toth, D.; Tamas, A.; Reglodi, D. The neuroprotective and biomarker potential of PACAP in human traumatic brain injury. Int. J. Mol. Sci. 2020, 21, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, B.J.; Basille, M.; Vaudry, D.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide promotes cell survival and neurite outgrowth in rat cerebellar neuroblasts. Neuroscience 1997, 78, 419–430. [Google Scholar] [CrossRef]
- Suarez, V.; Guntinas-Lichius, O.; Streppel, M.; Ingorokva, S.; Grosheva, M.; Neiss, W.F.; Angelov, D.N.; Klimaschewski, L. The axotomy-induced neuropeptides galanin and pituitary adenylate cyclase-activating peptide promote axonal sprouting of primary afferent and cranial motor neurones. Eur. J. Neurosci. 2006, 24, 1555–1564. [Google Scholar] [CrossRef]
- Waschek, J.A. Multiple actions of pituitary adenylyl cyclase activating peptide in nervous system development and regeneration. Dev. Neurosci. 2002, 24, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.; Kubota, H.; Ozaki, Y.; Uda, S.; Kuroda, S. Timing-dependent actions of NGF required for cell differentiation. PLoS ONE 2010, 5, e9011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gey, M.; Wanner, R.; Schilling, C.; Pedro, M.T.; Sinske, D.; Knöll, B. Atf3 mutant mice show reduced axon regeneration and impaired regeneration-associated gene induction after peripheral nerve injury. Open Biol. 2016, 6, 160091. [Google Scholar] [CrossRef] [Green Version]
- Tsuchida, M.; Nakamachi, T.; Sugiyama, K.; Tsuchikawa, D.; Watanabe, J.; Hori, M.; Yoshikawa, A.; Imai, N.; Kagami, N.; Matkovits, A.; et al. PACAP Stimulates Functional Recovery after Spinal Cord Injury through Axonal Regeneration. J. Mol. Neurosci. 2014, 54, 380–387. [Google Scholar] [CrossRef]
- Armstrong, B.D.; Abad, C.; Chhith, S.; Cheung-Lau, G.; Hajji, O.E.; Coute, A.C.; Ngo, D.H.; Waschek, J.A. Impairment of axotomy-induced pituitary adenylyl cyclase-activating peptide gene expression in T helper 2 lymphocyte-deficient mice. Neuroreport 2006, 17, 309–312. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H.; Kawatani, M.; Ito, E.; Ishikawa, K. Effects of pituitary adenylate cyclase-activating polypeptide on facial nerve recovery in the guinea pig. Laryngoscope 2003, 113, 1000–1006. [Google Scholar] [CrossRef]
- Kimura, H.; Kawatani, M.; Ito, E.; Ishikawa, K. PACAP facilitate the nerve regeneration factors in the facial nerve injury. Regul. Pept. 2004, 123, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Giardino, I.; Edelstein, D.; Brownlee, M. Changes in diabetic retinal matrix protein mRNA levels in a common transgenic mouse strain. Curr. Eye Res. 2000, 21, 581–587. [Google Scholar] [CrossRef]
- Roy, S.; Maiello, M.; Lorenzi, M. Increased expression of basement membrane collagen in human diabetic retinopathy. J. Clin. Invest. 1994, 93, 438–442. [Google Scholar] [CrossRef]
- Shimizu, F.; Sano, Y.; Haruki, H.; Kanda, T. Advanced glycation end-products induce basement membrane hypertrophy in endoneurial microvessels and disrupt the blood-nerve barrier by stimulating the release of TGF-β and vascular endothelial growth factor (VEGF) by pericytes. Diabetologia 2011, 54, 1517–1526. [Google Scholar] [CrossRef] [Green Version]
- Castorina, A.; Giunta, S.; Mazzone, V.; Cardile, V.; D’Agata, V. Effects of PACAP and VIP on hyperglycemia-induced proliferation in murine microvascular endothelial cells. Peptides 2010, 31, 2276–2283. [Google Scholar] [CrossRef] [PubMed]
- Rácz, B.; Gasz, B.; Borsiczky, B.; Gallyas, F.; Tamás, A.; Józsa, R.; Lubics, A.; Kiss, P.; Roth, E.; Ferencz, A.; et al. Protective effects of pituitary adenylate cyclase activating polypeptide in endothelial cells against oxidative stress-induced apoptosis. Gen. Comp. Endocrinol. 2007, 153, 115–123. [Google Scholar] [CrossRef]
- Malcangio, M.; Tomlinson, D.R. A pharmacologic analysis of mechanical hyperalgesia in streptozotocin/diabetic rats. Pain 1998, 76, 151–157. [Google Scholar] [CrossRef]
- Németh, J.; Reglofdi, D.; Pozsgai, G.; Szabó, Á.; Elekes, K.; Pintér, E.; Szolcsányi, J.; Helyes, Z. Effect of pituitary adenylate cyclase activating polypeptide-38 on sensory neuropeptide release and neurogenic inflammation in rats and mice. Neuroscience 2006, 143, 223–230. [Google Scholar] [CrossRef]
- Perrotti, L.I.; Hadeishi, Y.; Ulery, P.G.; Barrot, M.; Monteggia, L.; Duman, R.S.; Nestler, E.J. Induction of deltaFosB in reward-related brain structures after chronic stress. J. Neurosci. 2004, 24, 10594–10602. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Tao, X.; Huang, S.T.; Wu, L.; Tang, H.L.; Song, Y.; Zhang, G.; Zhang, Y.M. Chronic stress is associated with pain precipitation and elevation in DeltaFosB expression. Front. Pharmacol. 2016, 7, 138. [Google Scholar] [CrossRef] [Green Version]
- Luis-Delgado, O.E.; Barrot, M.; Rodeau, J.L.; Ulery, P.G.; Freund-Mercier, M.J.; Lasbennes, F. The transcription factor deltaFosB is recruited by inflammatory pain. J. Neurochem. 2006, 98, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Heinricher, M.M.; Tavares, I.; Leith, J.L.; Lumb, B.M. Descending control of nociception: Specificity, recruitment and plasticity. Brain Res. Rev. 2009, 60, 214–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaszner, B.; Kormos, V.; Kozicz, T.; Hashimoto, H.; Reglodi, D.; Helyes, Z. The behavioral phenotype of pituitary adenylate-cyclase activating polypeptide-deficient mice in anxiety and depression tests is accompanied by blunted c-Fos expression in the bed nucleus of the stria terminalis, central projecting Edinger-Westphal nucleus, ventral lateral septum, and dorsal raphe nucleus. Neuroscience 2012, 202, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, F.; Fujiwara, K.; Tsukahara, S.; Yada, T. Pituitary adenylate cyclase-activating polypeptide neurons of the ventromedial hypothalamus project to the midbrain central gray. Neuroreport 2006, 17, 221–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castorina, A.; Vogiatzis, M.; Kang, J.W.M.; Keay, K.A. PACAP and VIP expression in the periaqueductal grey of the rat following sciatic nerve constriction injury. Neuropeptides 2019, 74, 60–69. [Google Scholar] [CrossRef]
- Yamaoka, S.; Oshima, Y.; Horiuchi, H.; Morino, T.; Hino, M.; Miura, H.; Ogata, T. Altered gene expression of RNF34 and PACAP possibly involved in mechanism of exercise-induced analgesia for neuropathic pain in rats. Int. J. Mol. Sci. 2017, 18, 1962. [Google Scholar] [CrossRef] [Green Version]
- Yokai, M.; Kurihara, T.; Miyata, A. Spinal astrocytic activation contributes to both induction and maintenance of pituitary adenylate cyclase-activating polypeptide type 1 receptor-induced long-lasting mechanical allodynia in mice. Mol. Pain 2016, 12, 1744806916646383. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, D.; Birklein, F.; Reeh, P.W.; Sauer, S.K. Sensitized peripheral nociception in experimental diabetes of the rat. Pain 2010, 151, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Randall, L.O.; Selitto, J.J. A method for measurement of analgesic activity on inflamed tissue. Arch. Int. Pharmacodyn. Ther. 1957, 111, 409–419. [Google Scholar]
- Bölcskei, K.; Helyes, Z.; Szabó, Á.; Sándor, K.; Elekes, K.; Németh, J.; Almási, R.; Pintér, E.; Petho, G.; Szolcsányi, J. Investigation of the role of TRPV1 receptors in acute and chronic nociceptive processes using gene-deficient mice. Pain 2005, 117, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Kormos, V.; Gáspár, L.; Kovács, L.; Farkas, J.; Gaszner, T.; Csernus, V.; Balogh, A.; Hashimoto, H.; Reglodi, D.; Helyes, Z.; et al. Reduced response to chronic mild stress in PACAP mutant mice is associated with blunted FosB expression in limbic forebrain and brainstem centers. Neuroscience 2016, 330, 335–358. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.L.; Vialou, V.; Rios, L.; Carle-Florence, T.L.; Chakravarty, S.; Kumar, A.; Graham, D.L.; Green, T.A.; Kirk, A.; Iñiguez, S.D.; et al. The influence of ΔfosB in the nucleus accumbens on natural reward-related behavior. J. Neurosci. 2008, 28, 10272–10277. [Google Scholar] [CrossRef] [PubMed]
- Sterrenburg, L.; Gaszner, B.; Boerrigter, J.; Santbergen, L.; Bramini, M.; Elliott, E.; Chen, A.; Peeters, B.W.M.M.; Roubos, E.W.; Kozicz, T. Chronic stress induces sex-specific alterations in methylation and expression of corticotropin-releasing factor gene in the rat. PLoS ONE 2011, 6, e28128. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiss, P.; Banki, E.; Gaszner, B.; Nagy, D.; Helyes, Z.; Pal, E.; Reman, G.; Toth, G.; Tamas, A.; Reglodi, D. Protective Effects of PACAP in a Rat Model of Diabetic Neuropathy. Int. J. Mol. Sci. 2021, 22, 10691. https://doi.org/10.3390/ijms221910691
Kiss P, Banki E, Gaszner B, Nagy D, Helyes Z, Pal E, Reman G, Toth G, Tamas A, Reglodi D. Protective Effects of PACAP in a Rat Model of Diabetic Neuropathy. International Journal of Molecular Sciences. 2021; 22(19):10691. https://doi.org/10.3390/ijms221910691
Chicago/Turabian StyleKiss, Peter, Eszter Banki, Balazs Gaszner, Daniel Nagy, Zsuzsanna Helyes, Endre Pal, Gyongyver Reman, Gabor Toth, Andrea Tamas, and Dora Reglodi. 2021. "Protective Effects of PACAP in a Rat Model of Diabetic Neuropathy" International Journal of Molecular Sciences 22, no. 19: 10691. https://doi.org/10.3390/ijms221910691