
Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
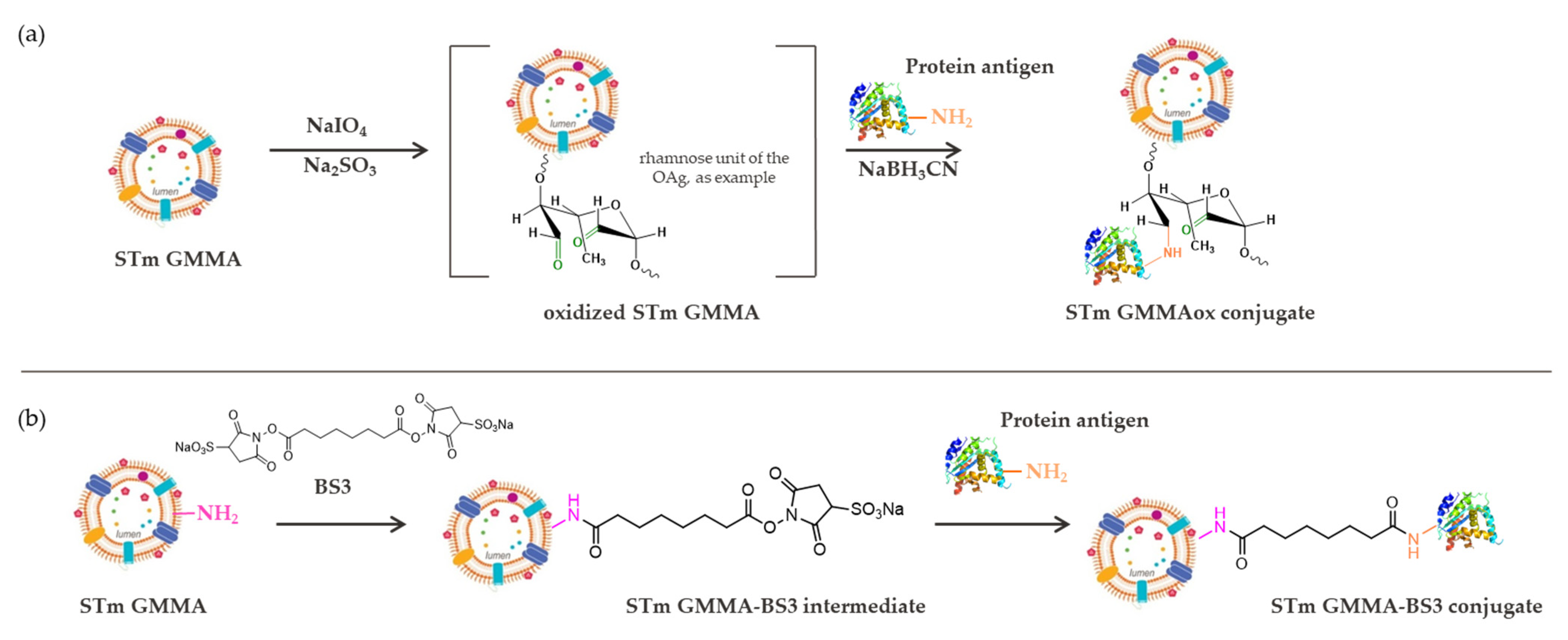
2.1. Conjugation Chemistry Targeting LPS/LOS on GMMA
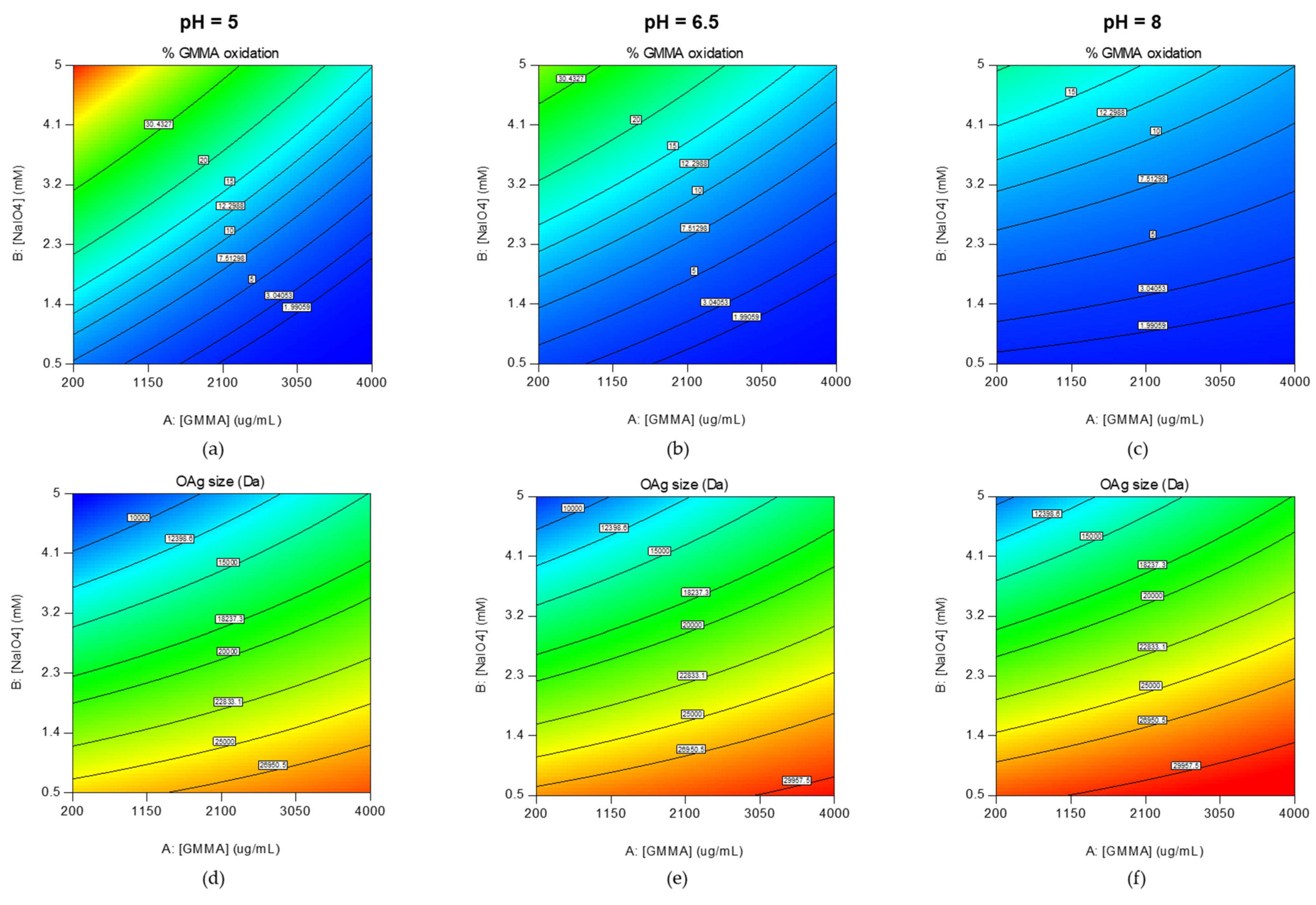
2.1.1. Design of Experiment to Identify the Main Parameters Affecting GMMA Oxidation
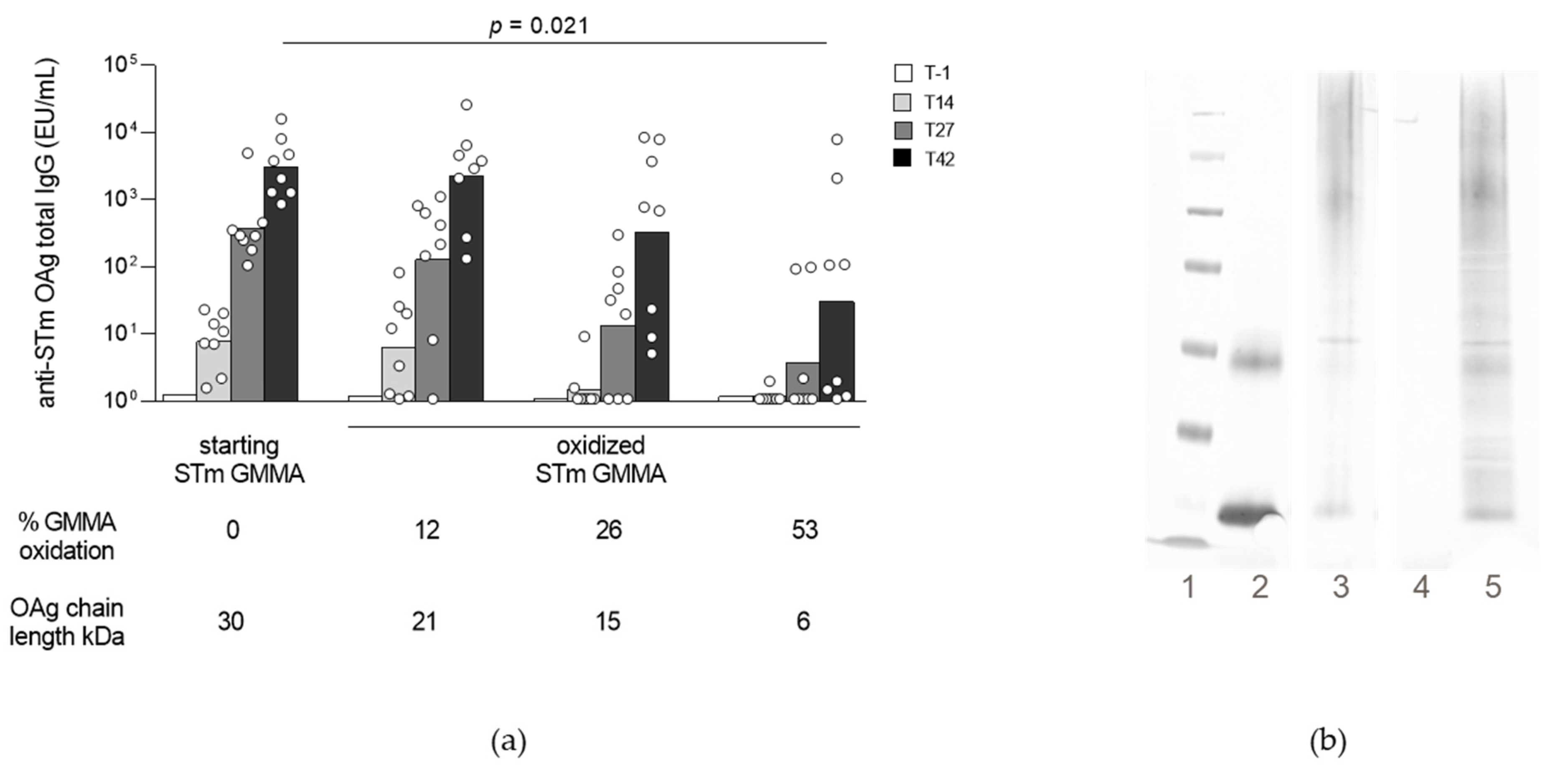
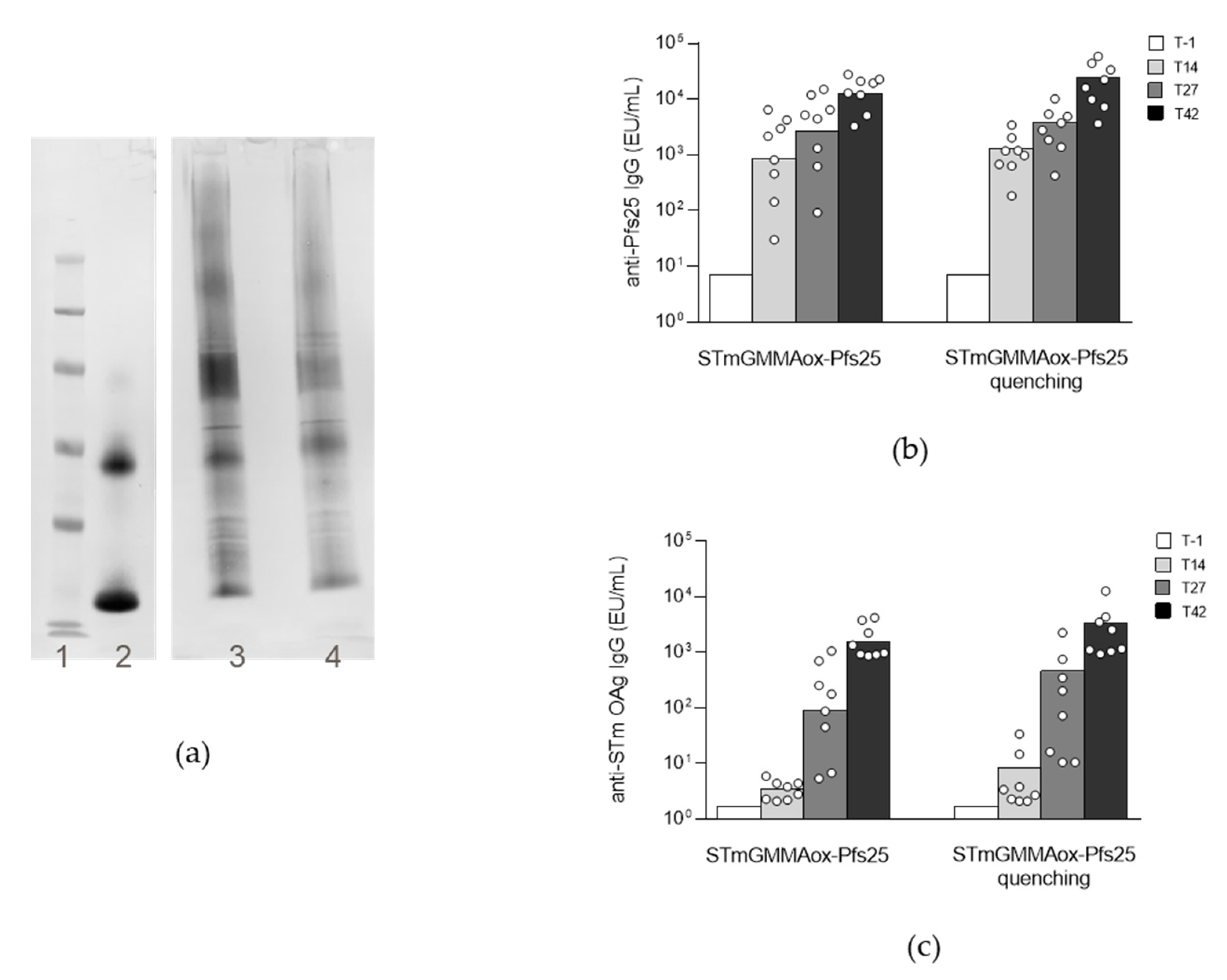
2.1.2. Impact of Oxidation Degree on GMMA Immunogenicity and Conjugation Efficiency
2.2. Conjugation Chemistry by Targeting Proteins on GMMA
2.2.1. DoE to Identify Optimal Conditions for GMMA Functionalization with BS3
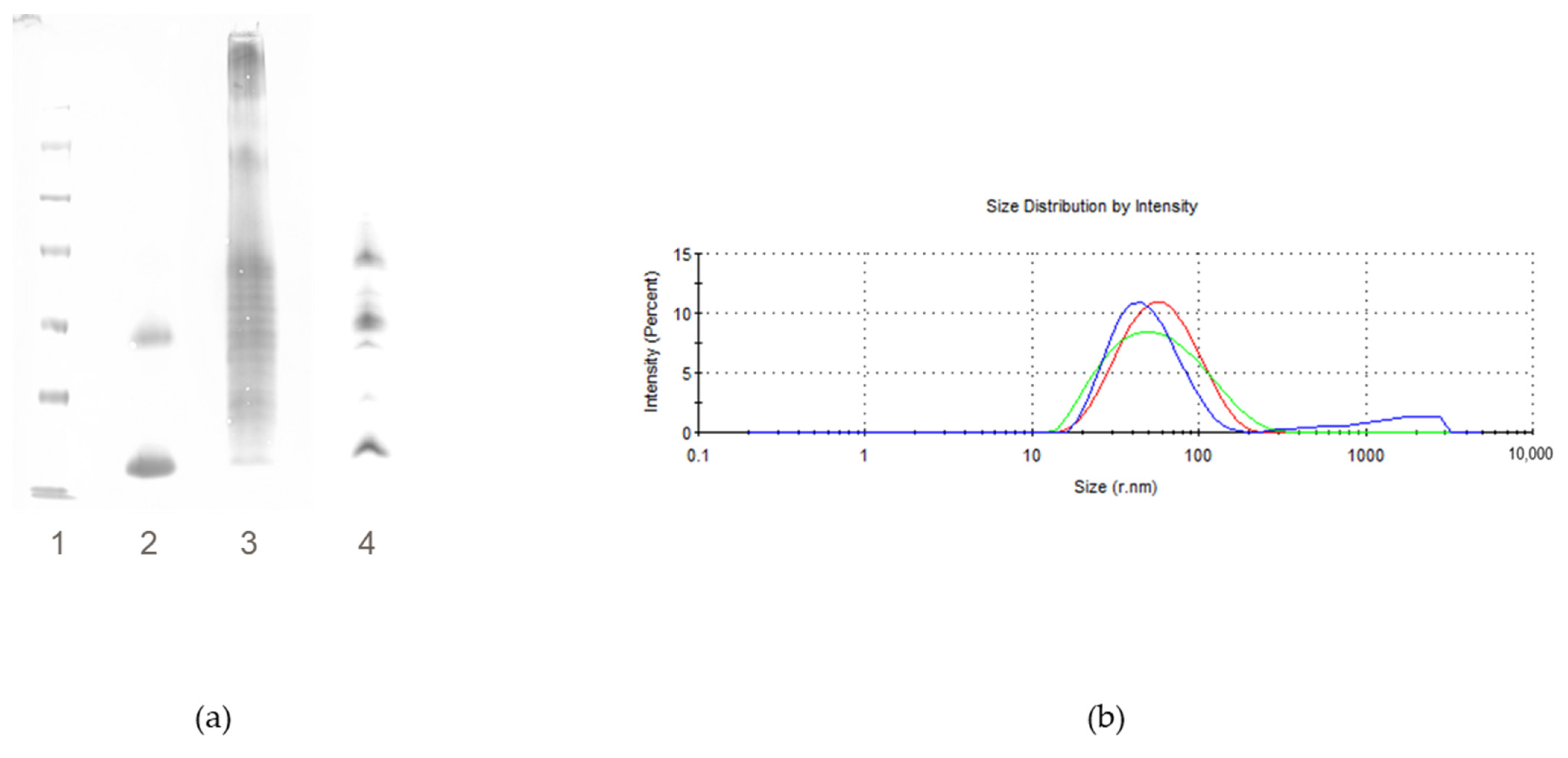
2.2.2. Impact of % Active Ester Groups on Conjugation Efficiency
2.3. Comparison of the Two Conjugation Approaches in Mice
2.4. Further Improvements of the Conjugation Processes
2.4.1. Conjugation by Reductive Amination: Quenching the Excess of Periodate with Sodium Sulfite
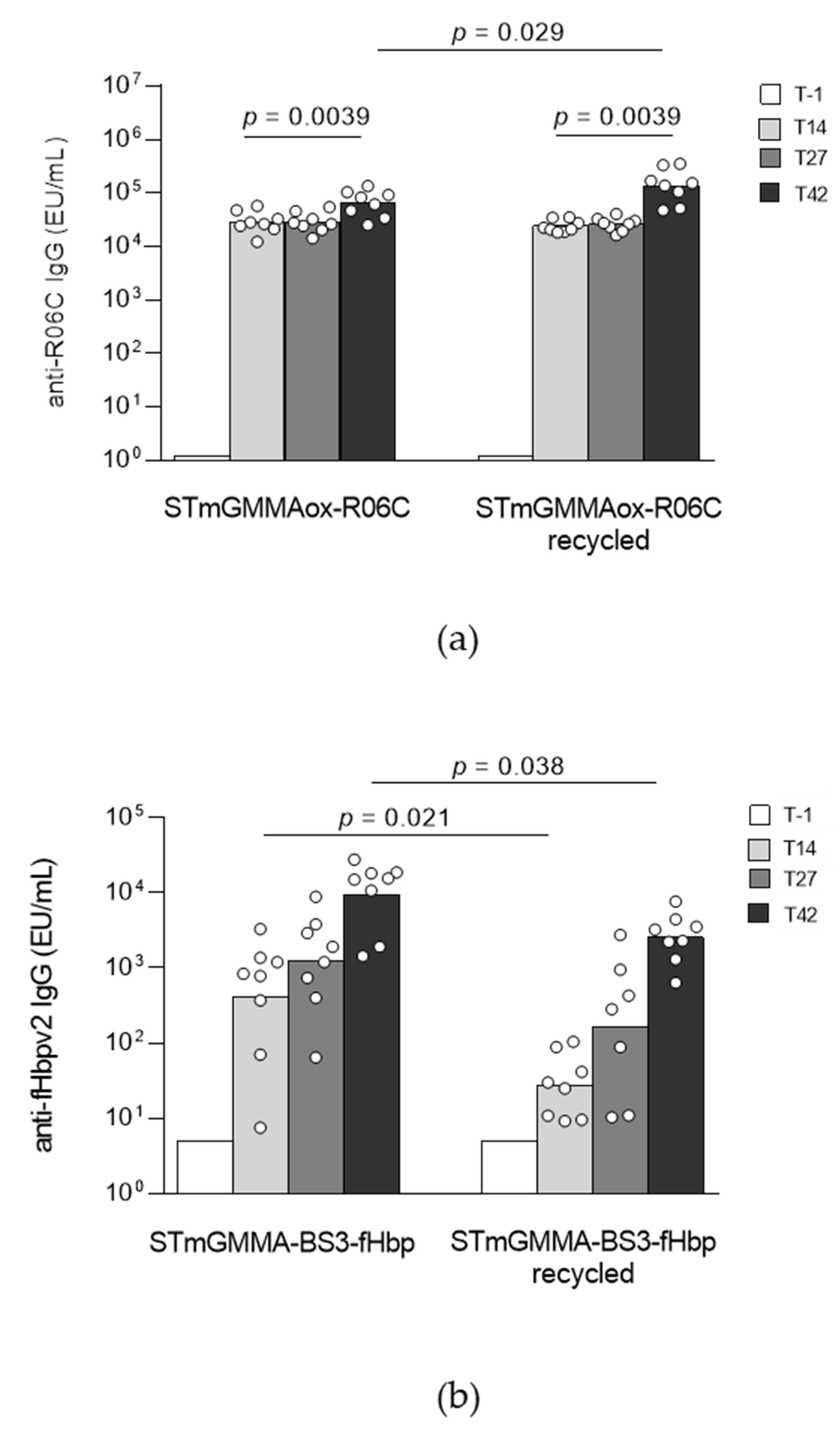
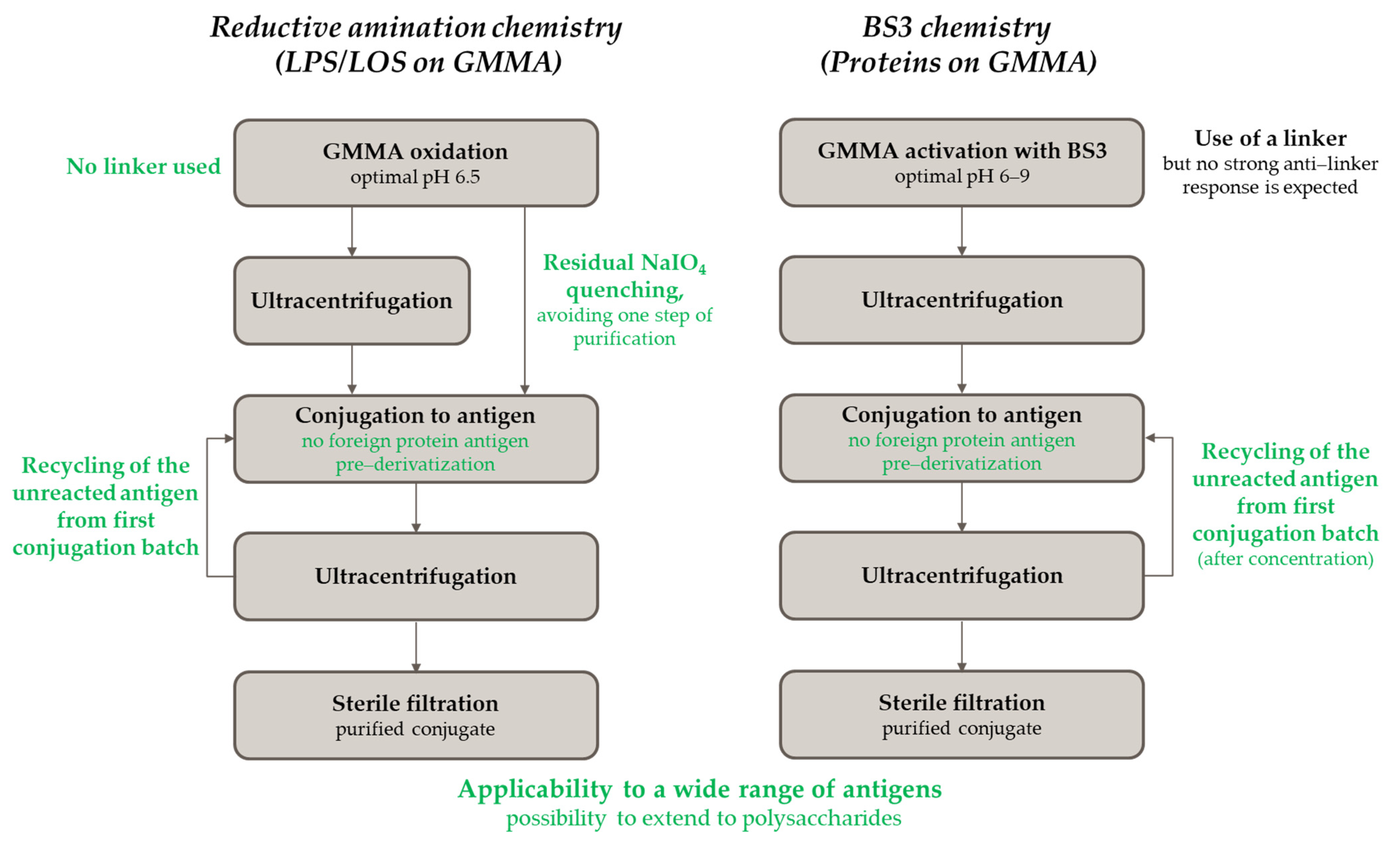
2.4.2. Application of Both Chemistries to Additional Antigens and Introduction of a Recycling Step
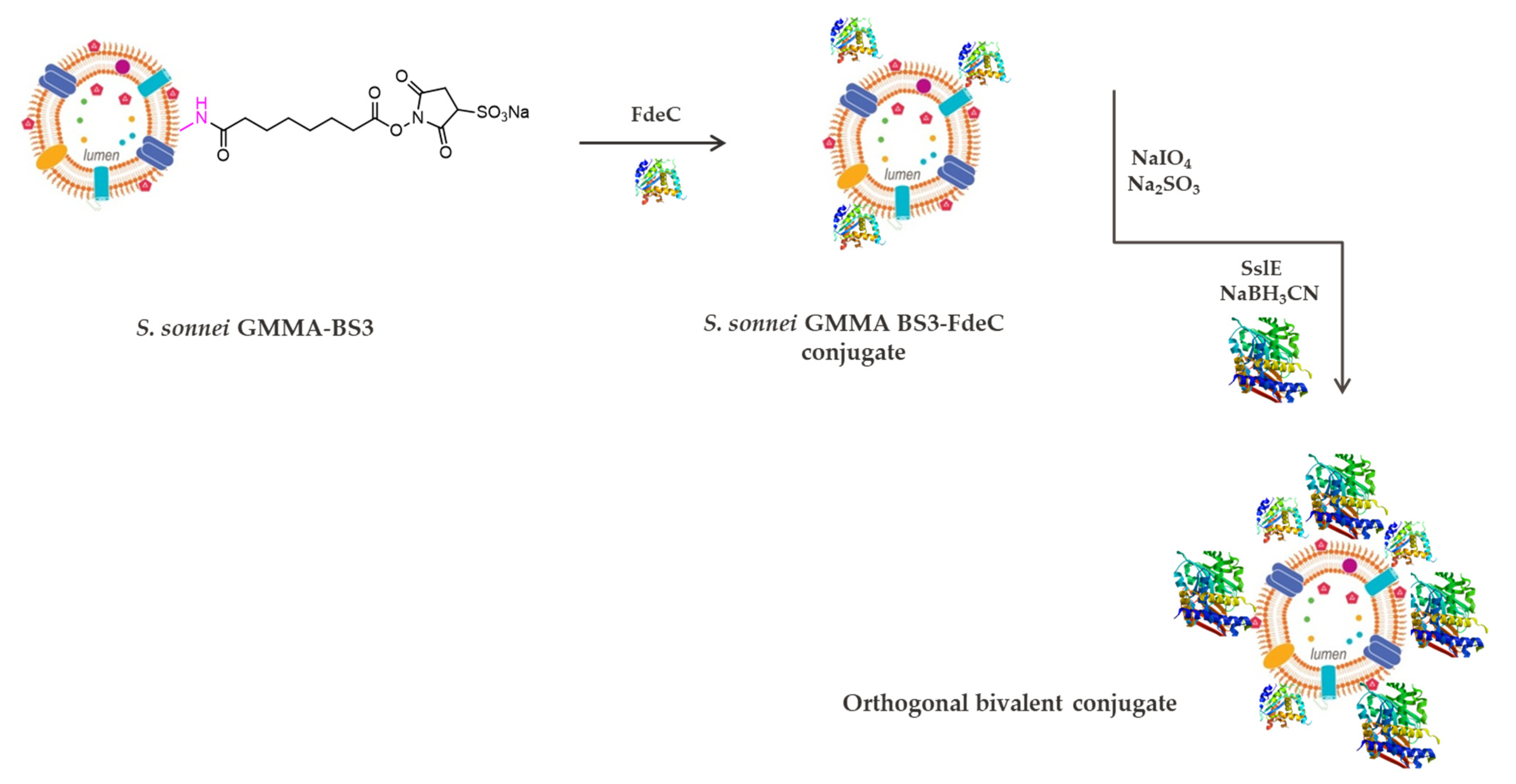
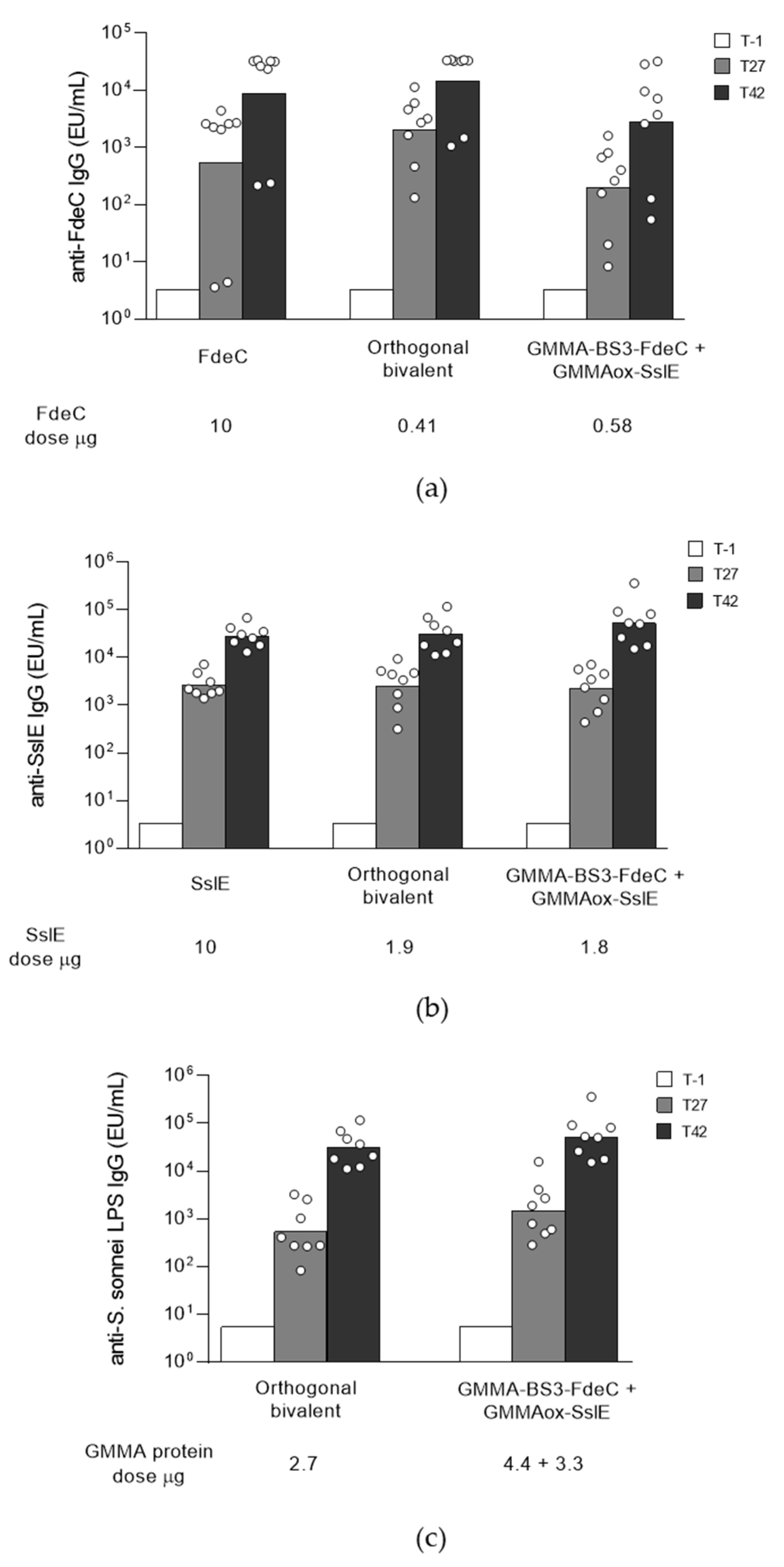
2.5. Reductive Amination and BS3 Chemistries for the Introduction of Two Different Proteins on the Same GMMA
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Conjugation Chemistry Targeting LPS/LOS on GMMA
4.2.1. GMMA Oxidation
4.2.2. Conjugation via Reductive Amination
4.3. Conjugation Chemistry by Targeting Proteins on GMMA
4.3.1. GMMA Activation with BS3
4.3.2. Conjugation via BS3 Chemistry
4.4. Synthesis of the Bivalent Conjugate
4.5. Design of Experiment (DoE)
4.6. Analytical Methods
4.6.1. High-Performance Liquid Chromatography–Size Exclusion Chromatography (HPLC-SEC) to Determine % Active Ester Groups Introduced
4.6.2. Competitive ELISA (cELISA)
4.7. Immunogenicity Studies in Mice
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; MacLennan, C.A. Outer membrane vesicle vaccines. Semin. Immunol. 2020, 50, 101433. [Google Scholar] [CrossRef] [PubMed]
- Gerritzen, M.J.H.; Martens, D.E.; Wijffels, R.H.; van der Pol, L.; Stork, M. Bioengineering bacterial outer membrane vesicles as vaccine platform. Biotechnol. Adv. 2017, 35, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Valguarnera, E.; Feldman, M.F. Glycoengineered Outer Membrane Vesicles as a Platform for Vaccine Development. Methods Enzymol. 2017, 597, 285–310. [Google Scholar] [PubMed]
- Kesty, N.C.; Kuehn, M.J. Incorporation of heterologous outer membrane and periplasmic proteins into Escherichia coli outer membrane vesicles. J. Biol. Chem. 2004, 279, 2069–2076. [Google Scholar] [CrossRef] [Green Version]
- Bartolini, E.; Ianni, E.; Frigimelica, E.; Petracca, R.; Galli, G.; Berlanda Scorza, F.; Norais, N.; Laera, D.; Giusti, F.; Pierleoni, A.; et al. Recombinant outer membrane vesicles carrying Chlamydia muridarum HtrA induce antibodies that neutralize chlamydial infection in vitro. J. Extracell. Vesicles 2013, 2, 20181. [Google Scholar] [CrossRef]
- Chen, D.J.; Osterrieder, N.; Metzger, S.M.; Buckles, E.; Doody, A.M.; DeLisa, M.P.; Putnam, D. Delivery of foreign antigens by engineered outer membrane vesicle vaccines. Proc. Natl. Acad. Sci. USA 2010, 107, 3099–3104. [Google Scholar] [CrossRef] [Green Version]
- Fantappiè, L.; de Santis, M.; Chiarot, E.; Carboni, F.; Bensi, G.; Jousson, O.; Margarit, I.; Grandi, G. Antibody-mediated immunity induced by engineered Escherichia coli OMVs carrying heterologous antigens in their lumen. J. Extracell. Vesicles 2014, 3, 24015. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Wang, S.; Yao, Y.; Xia, Y.; Yang, X.; Li, K.; Sun, P.; Liu, C.; Sun, W.; Bai, H.; et al. Employing Escherichia coli-derived outer membrane vesicles as an antigen delivery platform elicits protective immunity against Acinetobacter baumannii infection. Sci. Rep. 2016, 6, 37242. [Google Scholar] [CrossRef]
- Koeberling, O.; Ispasanie, E.; Hauser, J.; Rossi, O.; Pluschke, G.; Caugant, D.A.; Saul, A.; MacLennan, C.A. A broadly-protective vaccine against meningococcal disease in sub-Saharan Africa based on generalized modules for membrane antigens (GMMA). Vaccine 2014, 32, 2688–2695. [Google Scholar] [CrossRef] [Green Version]
- Muralinath, M.; Kuehn, M.J.; Roland, K.L.; Curtiss, R., 3rd. Immunization with Salmonella enterica serovar Typhimurium-derived outer membrane vesicles delivering the pneumococcal protein PspA confers protection against challenge with Streptococcus pneumoniae. Infect. Immun. 2011, 79, 887–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Valentine, J.L.; Huang, C.J.; Endicott, C.E.; Moeller, T.D.; Rasmussen, J.A.; Fletcher, J.R.; Boll, J.M.; Rosenthal, J.A.; Dobruchowska, J.; et al. Outer membrane vesicles displaying engineered glycotopes elicit protective antibodies. Proc. Natl. Acad. Sci. USA 2016, 113, E3609–E3618. [Google Scholar] [CrossRef] [Green Version]
- Price, N.L.; Goyette-Desjardins, G.; Nothaft, H.; Valguarnera, E.; Szymanski, C.M.; Segura, M.; Feldman, M.F. Glycoengineered Outer Membrane Vesicles: A Novel Platform for Bacterial Vaccines. Sci. Rep. 2016, 6, 24931. [Google Scholar] [CrossRef]
- Valentine, J.L.; Chen, L.; Perregaux, E.C.; Weyant, K.B.; Rosenthal, J.A.; Heiss, C.; Azadi, P.; Fisher, A.C.; Putnam, D.; Moe, G.R.; et al. Immunization with Outer Membrane Vesicles Displaying Designer Glycotopes Yields Class-Switched, Glycan-Specific Antibodies. Cell Chem. Biol. 2016, 23, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Mancini, F.; Rossi, O.; Necchi, F.; Micoli, F. OMV Vaccines and the Role of TLR Agonists in Immune Response. Int. J. Mol. Sci. 2020, 21, 4416. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Del Bino, L.; Alfini, R.; Carboni, F.; Romano, M.R.; Adamo, R. Glycoconjugate vaccines: Current approaches towards faster vaccine design. Expert Rev. Vaccines 2019, 18, 881–895. [Google Scholar] [CrossRef] [PubMed]
- Alves, N.J.; Turner, K.B.; Daniele, M.A.; Oh, E.; Medintz, I.L.; Walper, S.A. Bacterial Nanobioreactors--Directing Enzyme Packaging into Bacterial Outer Membrane Vesicles. ACS Appl. Mater. Interfaces 2015, 7, 24963–24972. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Palmieri, E.; Balocchi, C.; Gasperini, G.; et al. GMMA Is a Versatile Platform to Design Effective Multivalent Combination Vaccines. Vaccines 2020, 8, 540. [Google Scholar] [CrossRef] [PubMed]
- van den Berg van Saparoea, H.B.; Houben, D.; Kuijl, C.; Luirink, J.; Jong, W.S.P. Combining Protein Ligation Systems to Expand the Functionality of Semi-Synthetic Outer Membrane Vesicle Nanoparticles. Front. Microbiol. 2020, 11, 890. [Google Scholar] [CrossRef]
- Fukasawa, L.O.; Schenkman, R.P.; Perciani, C.T.; Carneiro, S.M.; Dias, W.O.; Tanizaki, M.M. Optimization of the conjugation method for a serogroup B/C meningococcal vaccine. Biotechnol. Appl. Biochem. 2006, 45 Pt 3, 141–146. [Google Scholar]
- Siadat, S.D.; Vaziri, F.; Eftekhary, M.; Karbasian, M.; Moshiri, A.; Aghasadeghi, M.R.; Ardestani, M.S.; Alitappeh, M.A.; Arsang, A.; Fateh, A.; et al. Preparation and Evaluation of a New Lipopolysaccharide-based Conjugate as a Vaccine Candidate for Brucellosis. Osong Public Health Res. Perspect. 2015, 6, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Gerritzen, M.J.H.; Salverda, M.L.M.; Martens, D.E.; Wijffels, R.H.; Stork, M. Spontaneously released Neisseria meningitidis outer membrane vesicles as vaccine platform: Production and purification. Vaccine 2019, 37, 6978–6986. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Alfini, R.; Di Benedetto, R.; Necchi, F.; Schiavo, F.; Mancini, F.; Carducci, M.; Oldrini, D.; Pitirollo, O.; Gasperini, G.; et al. Generalized Modules for Membrane Antigens as Carrier for Polysaccharides: Impact of Sugar Length, Density, and Attachment Site on the Immune Response Elicited in Animal Models. Front. Immunol. 2021, 12, 3544. [Google Scholar] [CrossRef]
- De Benedetto, G.; Alfini, R.; Cescutti, P.; Caboni, M.; Lanzilao, L.; Necchi, F.; Saul, A.; MacLennan, C.A.; Rondini, S.; Micoli, F. Characterization of O-antigen delivered by Generalized Modules for Membrane Antigens (GMMA) vaccine candidates against nontyphoidal Salmonella. Vaccine 2017, 35, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Gasperini, G.; Alfini, R.; Arato, V.; Mancini, F.; Aruta, M.G.; Kanvatirth, P.; Pickard, D.; Necchi, F.; Saul, A.; Rossi, O.; et al. Salmonella Paratyphi A Outer Membrane Vesicles Displaying Vi Polysaccharide as a Multivalent Vaccine against Enteric Fever. Infect. Immun. 2021, 89, e00699-20. [Google Scholar] [CrossRef]
- Gerke, C.; Colucci, A.M.; Giannelli, C.; Sanzone, S.; Vitali, C.G.; Sollai, L.; Rossi, O.; Martin, L.B.; Auerbach, J.; Di Cioccio, V.; et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE 2015, 10, e0134478. [Google Scholar] [CrossRef]
- Raso, M.M.; Gasperini, G.; Alfini, R.; Schiavo, F.; Aruta, M.G.; Carducci, M.; Forgione, M.C.; Martini, S.; Cescutti, P.; Necchi, F.; et al. GMMA and Glycoconjugate Approaches Compared in Mice for the Development of a Vaccine against Shigella flexneri Serotype 6. Vaccines 2020, 8, 160. [Google Scholar] [CrossRef] [Green Version]
- Schager, A.E.; Dominguez-Medina, C.C.; Necchi, F.; Micoli, F.; Goh, Y.S.; Goodall, M.; Flores-Langarica, A.; Bobat, S.; Cook, C.N.L.; Arcuri, M.; et al. IgG Responses to Porins and Lipopolysaccharide within an Outer Membrane-Based Vaccine against Nontyphoidal Salmonella Develop at Discordant Rates. mBio 2018, 9, e02379-17. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.S.; Laskowich, E.R.; Michon, F.; Kaiser, R.E.; Arumugham, R.G. Monitoring activation sites on polysaccharides by GC-MS. Anal. Biochem. 2006, 358, 136–142. [Google Scholar] [CrossRef]
- Lanzilao, L.; Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F.; Rondini, S. Strain Selection for Generation of O-Antigen-Based Glycoconjugate Vaccines against Invasive Nontyphoidal Salmonella Disease. PLoS ONE 2015, 10, e0139847. [Google Scholar]
- Rondini, S.; Micoli, F.; Lanzilao, L.; Gavini, M.; Alfini, R.; Brandt, C.; Clare, S.; Mastroeni, P.; Saul, A.; MacLennan, C.A. Design of glycoconjugate vaccines against invasive African Salmonella enterica serovar Typhimurium. Infect. Immun. 2015, 83, 996–1007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, D.C.; Robbins, J.B.; Szu, S.C. Protection of mice against Salmonella typhimurium with an O-specific polysaccharide-protein conjugate vaccine. Infect. Immun. 1992, 60, 4679–4686. [Google Scholar] [CrossRef] [Green Version]
- Stefanetti, G.; Rondini, S.; Lanzilao, L.; Saul, A.; MacLennan, C.A.; Micoli, F. Impact of conjugation chemistry on the immunogenicity of S. Typhimurium conjugate vaccines. Vaccine 2014, 32, 6122–6129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miura, K.; Keister, D.B.; Muratova, O.V.; Sattabongkot, J.; Long, C.A.; Saul, A. Transmission-blocking activity induced by malaria vaccine candidates Pfs25/Pvs25 is a direct and predictable function of antibody titer. Malar. J. 2007, 6, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimp, R.L., Jr.; Rowe, C.; Reiter, K.; Chen, B.; Nguyen, V.; Aebig, J.; Rausch, K.M.; Kumar, K.; Wu, Y.; Jin, A.J.; et al. Development of a Pfs25-EPA malaria transmission blocking vaccine as a chemically conjugated nanoparticle. Vaccine 2013, 31, 2954–2962. [Google Scholar] [CrossRef] [Green Version]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef]
- Mabey, D.C.; Brown, A.; Greenwood, B.M. Plasmodium falciparum malaria and Salmonella infections in Gambian children. J. Infect. Dis. 1987, 155, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, C.A.; Levine, M.M. Invasive nontyphoidal Salmonella disease in Africa: Current status. Expert Rev. Anti-Infect. Ther. 2013, 11, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Slauch, J.M.; Mahan, M.J.; Michetti, P.; Neutra, M.R.; Mekalanos, J.J. Acetylation (O-factor 5) affects the structural and immunological properties of Salmonella typhimurium lipopolysaccharide O antigen. Infect. Immun. 1995, 63, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.K.; Roeffen, W.; Andersen, G.; Bousema, T.; Christiansen, M.; Sauerwein, R.; Theisen, M. A Plasmodium falciparum 48/45 single epitope R0.6C subunit protein elicits high levels of transmission blocking antibodies. Vaccine 2015, 33, 1981–1986. [Google Scholar] [CrossRef]
- Singh, S.K.; Roeffen, W.; Mistarz, U.H.; Chourasia, B.K.; Yang, F.; Rand, K.D.; Sauerwein, R.W.; Theisen, M. Construct design, production, and characterization of Plasmodium falciparum 48/45 R0.6C subunit protein produced in Lactococcus lactis as candidate vaccine. Microb. Cell Factories 2017, 16, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biagini, M.; Spinsanti, M.; De Angelis, G.; Tomei, S.; Ferlenghi, I.; Scarselli, M.; Rigat, F.; Messuti, N.; Biolchi, A.; Muzzi, A.; et al. Expression of factor H binding protein in meningococcal strains can vary at least 15-fold and is genetically determined. Proc. Natl. Acad. Sci. USA 2016, 113, 2714–2719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacLennan, C.A.; Martin, L.B.; Micoli, F. Vaccines against invasive Salmonella disease: Current status and future directions. Hum. Vaccines Immunother. 2014, 10, 1478–1493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uche, I.V.; MacLennan, C.A.; Saul, A. A Systematic Review of the Incidence, Risk Factors and Case Fatality Rates of Invasive Nontyphoidal Salmonella (iNTS) Disease in Africa (1966 to 2014). PLoS Negl. Trop. Dis. 2017, 11, e0005118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nesta, B.; Valeri, M.; Spagnuolo, A.; Rosini, R.; Mora, M.; Donato, P.; Alteri, C.J.; Del Vecchio, M.; Buccato, S.; Pezzicoli, A.; et al. SslE elicits functional antibodies that impair in vitro mucinase activity and in vivo colonization by both intestinal and extraintestinal Escherichia coli strains. PLoS Pathog. 2014, 10, e1004124. [Google Scholar] [CrossRef] [Green Version]
- Valeri, M.; Rossi Paccani, S.; Kasendra, M.; Nesta, B.; Serino, L.; Pizza, M.; Soriani, M. Pathogenic, E. coli exploits SslE mucinase activity to translocate through the mucosal barrier and get access to host cells. PLoS ONE 2015, 10, e0117486. [Google Scholar] [CrossRef] [Green Version]
- Nesta, B.; Spraggon, G.; Alteri, C.; Moriel, D.G.; Rosini, R.; Veggi, D.; Smith, S.; Bertoldi, I.; Pastorello, I.; Ferlenghi, I.; et al. FdeC, a novel broadly conserved Escherichia coli adhesin eliciting protection against urinary tract infections. mBio 2012, 3, e00010-12. [Google Scholar] [CrossRef] [Green Version]
- Walker, R.I. An assessment of enterotoxigenic Escherichia coli and Shigella vaccine candidates for infants and children. Vaccine 2015, 33, 954–965. [Google Scholar] [CrossRef] [Green Version]
- Kolb, H.C.; Finn, M.G.; Sharpless, K.B. Click Chemistry: Diverse Chemical Function from a Few Good Reactions. Angew. Chem. Int. Ed. Engl. 2001, 40, 2004–2021. [Google Scholar] [CrossRef]
- Patel, K.G.; Swartz, J.R. Surface functionalization of virus-like particles by direct conjugation using azide-alkyne click chemistry. Bioconjug. Chem. 2011, 22, 376–387. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Przysiecki, C.; Flanagan, E.; Bello-Irizarry, S.N.; Ionescu, R.; Muratova, O.; Dobrescu, G.; Lambert, L.; Keister, D.; Rippeon, Y.; et al. Sustained high-titer antibody responses induced by conjugating a malarial vaccine candidate to outer-membrane protein complex. Proc. Natl. Acad. Sci. USA 2006, 103, 18243–18248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamo, R.; Hu, Q.-Y.; Torosantucci, A.; Crotti, S.; Brogioni, G.; Allan, M.; Chiani, P.; Bromuro, C.; Quinn, D.; Tontini, M.; et al. Deciphering the structure–immunogenicity relationship of anti-Candida glycoconjugate vaccines. Chem. Sci. 2014, 5, 4302–4311. [Google Scholar] [CrossRef] [Green Version]
- Adamo, R.; Nilo, A.; Castagner, B.; Boutureira, O.; Berti, F.; Bernardes, G.J. Synthetically defined glycoprotein vaccines: Current status and future directions. Chem. Sci. 2013, 4, 2995–3008. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.-Y.; Allan, M.; Adamo, R.; Quinn, D.; Zhai, H.; Wu, G.; Clark, K.; Zhou, J.; Ortiz, S.; Wang, B.; et al. Synthesis of a well-defined glycoconjugate vaccine by a tyrosine-selective conjugation strategy. Chem. Sci. 2013, 4, 3827–3832. [Google Scholar] [CrossRef] [Green Version]
- Nilo, A.; Allan, M.; Brogioni, B.; Proietti, D.; Cattaneo, V.; Crotti, S.; Sokup, S.; Zhai, H.; Margarit, I.; Berti, F.; et al. Tyrosine-directed conjugation of large glycans to proteins via copper-free click chemistry. Bioconjug. Chem. 2014, 25, 2105–2111. [Google Scholar] [CrossRef]
- Stefanetti, G.; Hu, Q.Y.; Usera, A.; Robinson, Z.; Allan, M.; Singh, A.; Imase, H.; Cobb, J.; Zhai, H.; Quinn, D.; et al. Sugar-Protein Connectivity Impacts on the Immunogenicity of Site-Selective Salmonella O-Antigen Glycoconjugate Vaccines. Angew. Chem. Int. Ed. Engl. 2015, 54, 13198–13203. [Google Scholar] [CrossRef] [Green Version]
- Stefanetti, G.; Saul, A.; MacLennan, C.A.; Micoli, F. Click Chemistry Applied to the Synthesis of Salmonella Typhimurium O-Antigen Glycoconjugate Vaccine on Solid Phase with Sugar Recycling. Bioconjug. Chem. 2015, 26, 2507–2513. [Google Scholar] [CrossRef] [PubMed]
- Nilo, A.; Passalacqua, I.; Fabbrini, M.; Allan, M.; Usera, A.; Carboni, F.; Brogioni, B.; Pezzicoli, A.; Cobb, J.; Romano, M.R.; et al. Exploring the Effect of Conjugation Site and Chemistry on the Immunogenicity of an anti-Group B Streptococcus Glycoconjugate Vaccine Based on GBS67 Pilus Protein and Type V Polysaccharide. Bioconjug. Chem. 2015, 26, 1839–1849. [Google Scholar] [CrossRef]
- Stefanetti, G.; Allan, M.; Usera, A.; Micoli, F. Click chemistry compared to thiol chemistry for the synthesis of site-selective glycoconjugate vaccines using CRM(197) as carrier protein. Glycoconj. J. 2020, 37, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Zakeri, B.; Fierer, J.O.; Celik, E.; Chittock, E.C.; Schwarz-Linek, U.; Moy, V.T.; Howarth, M. Peptide tag forming a rapid covalent bond to a protein, through engineering a bacterial adhesin. Proc. Natl. Acad. Sci. USA 2012, 109, E690–E697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kis, Z.; Shattock, R.; Shah, N.; Kontoravdi, C. Emerging Technologies for Low-Cost, Rapid Vaccine Manufacture. Biotechnol. J. 2019, 14, e1800376. [Google Scholar] [CrossRef] [Green Version]
- Micoli, F.; Adamo, R.; Costantino, P. Protein Carriers for Glycoconjugate Vaccines: History, Selection Criteria, Characterization and New Trends. Molecules 2018, 23, 1451. [Google Scholar] [CrossRef] [Green Version]
- Beg, S.; Swain, S.; Rahman, M.; Hasnain, M.S.; Imam, S.S. Chapter 3—Application of Design of Experiments (DoE) in Pharmaceutical Product and Process Optimization. In Pharmaceutical Quality by Design; Beg, S., Hasnain, M.S., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 43–64. [Google Scholar]
- Montgomery, D.C. Response surface methods and other approaches to process optimization. In Design and Analysis of Experiments; John Wiley & Sons: Hoboken, NJ, USA, 1997; pp. 427–510. [Google Scholar]
- Goh, Y.S.; Clare, S.; Micoli, F.; Saul, A.; Mastroeni, P.; MacLennan, C.A. Monoclonal Antibodies of a Diverse Isotype Induced by an O-Antigen Glycoconjugate Vaccine Mediate In Vitro and In Vivo Killing of African Invasive Nontyphoidal Salmonella. Infect. Immun. 2015, 83, 3722–3731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rondini, S.; Lanzilao, L.; Necchi, F.; O’Shaughnessy, C.M.; Micoli, F.; Saul, A.; MacLennan, C.A. Invasive African Salmonella Typhimurium induces bactericidal antibodies against O-antigens. Microb. Pathog. 2013, 63, 19–23. [Google Scholar] [CrossRef] [PubMed]
- De Benedetto, G.; Cescutti, P.; Giannelli, C.; Rizzo, R.; Micoli, F. Multiple Techniques for Size Determination of Generalized Modules for Membrane Antigens from Salmonella typhimurium and Salmonella enteritidis. ACS Omega 2017, 2, 8282–8289. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Ravenscroft, N.; Cescutti, P.; Stefanetti, G.; Londero, S.; Rondini, S.; Maclennan, C.A. Structural analysis of O-polysaccharide chains extracted from different Salmonella Typhimurium strains. Carbohydr. Res. 2014, 385, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Satake, K.; Okuyama, T.; Ohashi, M.; Shinoda, T. The spectrophotometric determination of amine, amino acid and peptide with 2, 4, 6-trinitrobenzene 1-sulfonic acid. J. Biochem. 1960, 47, 654–660. [Google Scholar] [CrossRef]
- Necchi, F.; Stefanetti, G.; Alfini, R.; Palmieri, E.; Carducci, M.; Di Benedetto, R.; Schiavo, F.; Aruta, M.G.; Giusti, F.; Ferlenghi, I.; et al. Neisseria meningitidis Factor H Binding Protein Surface Exposure on Salmonella Typhimurium GMMA Is Critical to Induce an Effective Immune Response against Both Diseases. Pathogens 2021, 10, 726. [Google Scholar] [CrossRef]
- Rossi, O.; Aruta, M.G.; Acquaviva, A.; Mancini, F.; Micoli, F.; Necchi, F. Characterization of Competitive ELISA and Formulated Alhydrogel Competitive ELISA (FAcE) for Direct Quantification of Active Ingredients in GMMA-Based Vaccines. Methods Protoc. 2020, 3, 62. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Conditions for Oxidation | Expected GMMA ox % (95% TI) | Expected OAg Size Da (95% TI) | I Set Experiments (Used for Study in Mice) | II Set Experiments (Used for Conjugation Tests) | ||||
|---|---|---|---|---|---|---|---|---|---|
| [GMMA] mg/mL | [NaIO4] mM | pH | GMMA ox % | OAg Size Da | GMMA ox % | OAg Size Da | |||
| 5 | 0.2 | 5 | 5 | 57.1 (42.2–74.1) | 6200 (2211–14,610) | 53 | 6039 | 60 | 7038 |
| 9 | 2.1 | 5 | 6.5 | 23 (15.1–32.2) | 12,266 (4794–19,738) | 26 | 15,327 | 26 1 | 15,327 1 |
| 14 | 2.1 | 2.75 | 6.5 | 9.1 (4.7–14.8) | 20,729 (13,733–27,725) | 12 | 21,163 | 14 | 23,464 |
| Run | Conditions for GMMA-BS3 Activation | % Active Ester Groups Introduced | |||
|---|---|---|---|---|---|
| [GMMA] mg/mL | [BS3] mg/mL (eq Respect to NH2 on GMMA) | pH | Expected (95% TI) | Obtained | |
| Optimized condition | 4 | 50 (10.4) | 9 | 31.3 (11.3–51.3) | 32 |
| 1 | 0.2 | 0.2 (5) | 6 | 10.6 (0–30.2) | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Benedetto, R.; Alfini, R.; Carducci, M.; Aruta, M.G.; Lanzilao, L.; Acquaviva, A.; Palmieri, E.; Giannelli, C.; Necchi, F.; Saul, A.; et al. Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens. Int. J. Mol. Sci. 2021, 22, 10180. https://doi.org/10.3390/ijms221910180
Di Benedetto R, Alfini R, Carducci M, Aruta MG, Lanzilao L, Acquaviva A, Palmieri E, Giannelli C, Necchi F, Saul A, et al. Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens. International Journal of Molecular Sciences. 2021; 22(19):10180. https://doi.org/10.3390/ijms221910180
Chicago/Turabian StyleDi Benedetto, Roberta, Renzo Alfini, Martina Carducci, Maria Grazia Aruta, Luisa Lanzilao, Alessandra Acquaviva, Elena Palmieri, Carlo Giannelli, Francesca Necchi, Allan Saul, and et al. 2021. "Novel Simple Conjugation Chemistries for Decoration of GMMA with Heterologous Antigens" International Journal of Molecular Sciences 22, no. 19: 10180. https://doi.org/10.3390/ijms221910180