
Intranasal Delivery of RGD-Containing Osteopontin Heptamer Peptide Confers Neuroprotection in the Ischemic Brain and Augments Microglia M2 Polarization

Abstract
:1. Introduction
2. Results
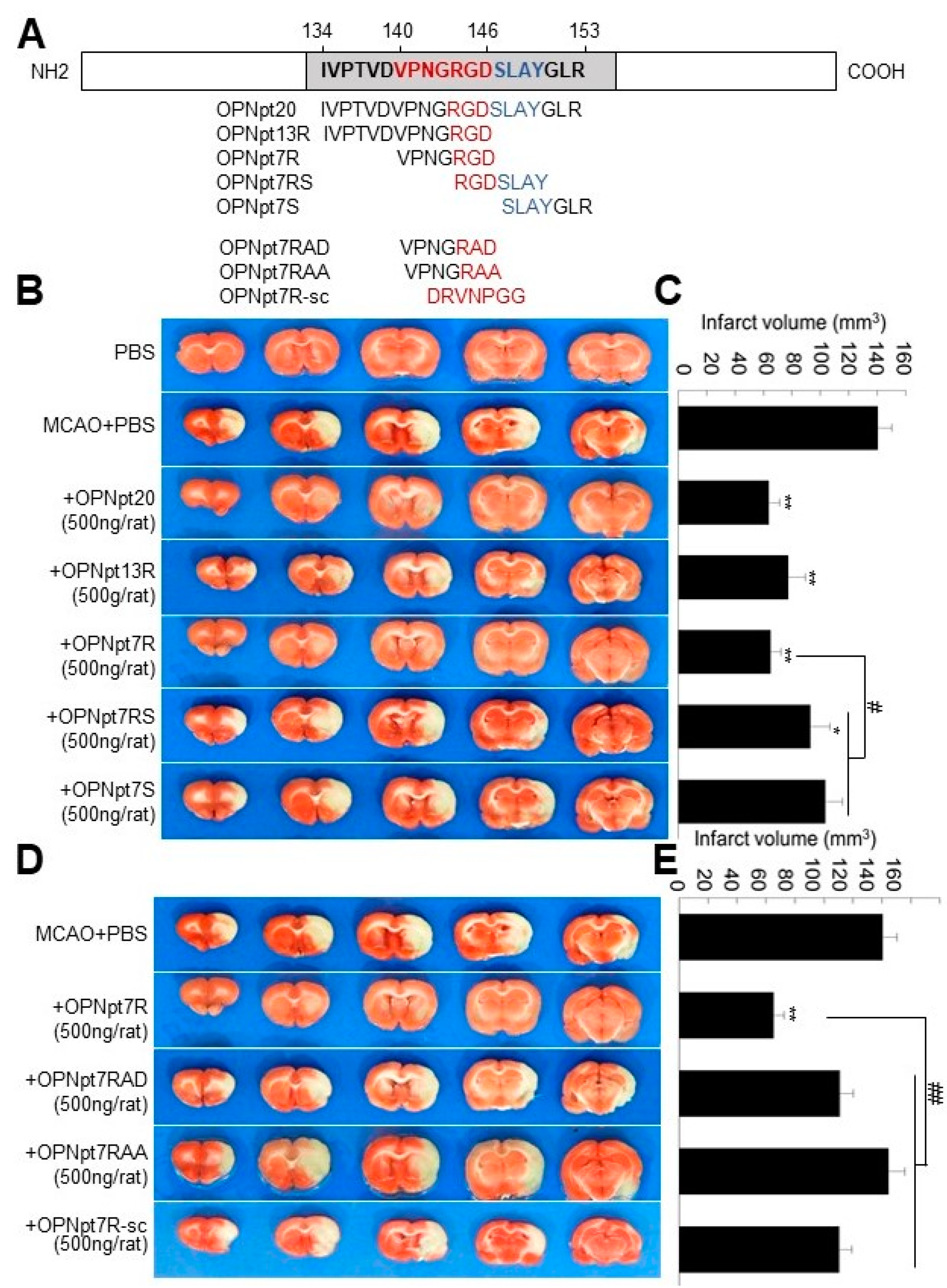
2.1. OPN Heptamer Peptides (OPNpt7) Containing an RGD Motif Confers a Neuroprotective Effect in the Post-Ischemic Rat Brain
2.2. RGD Motif Plays a Critical Role in OPNpt7R-Mediated Infarct Suppression in the Post-Ischemic Brain
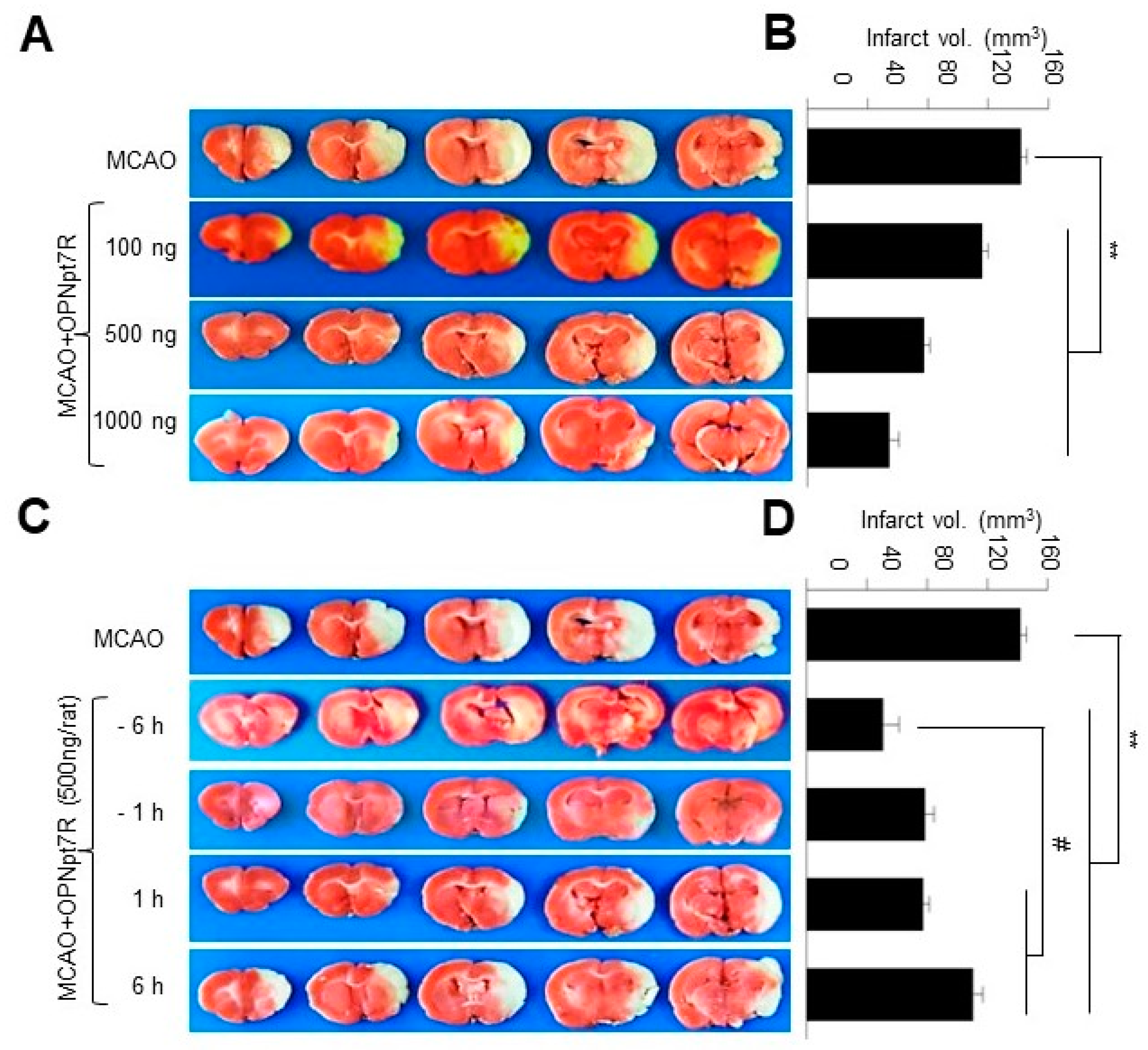
2.3. OPNpt7 Exhibits a Wide Therapeutic Time Window in the Post-Ischemic Brain
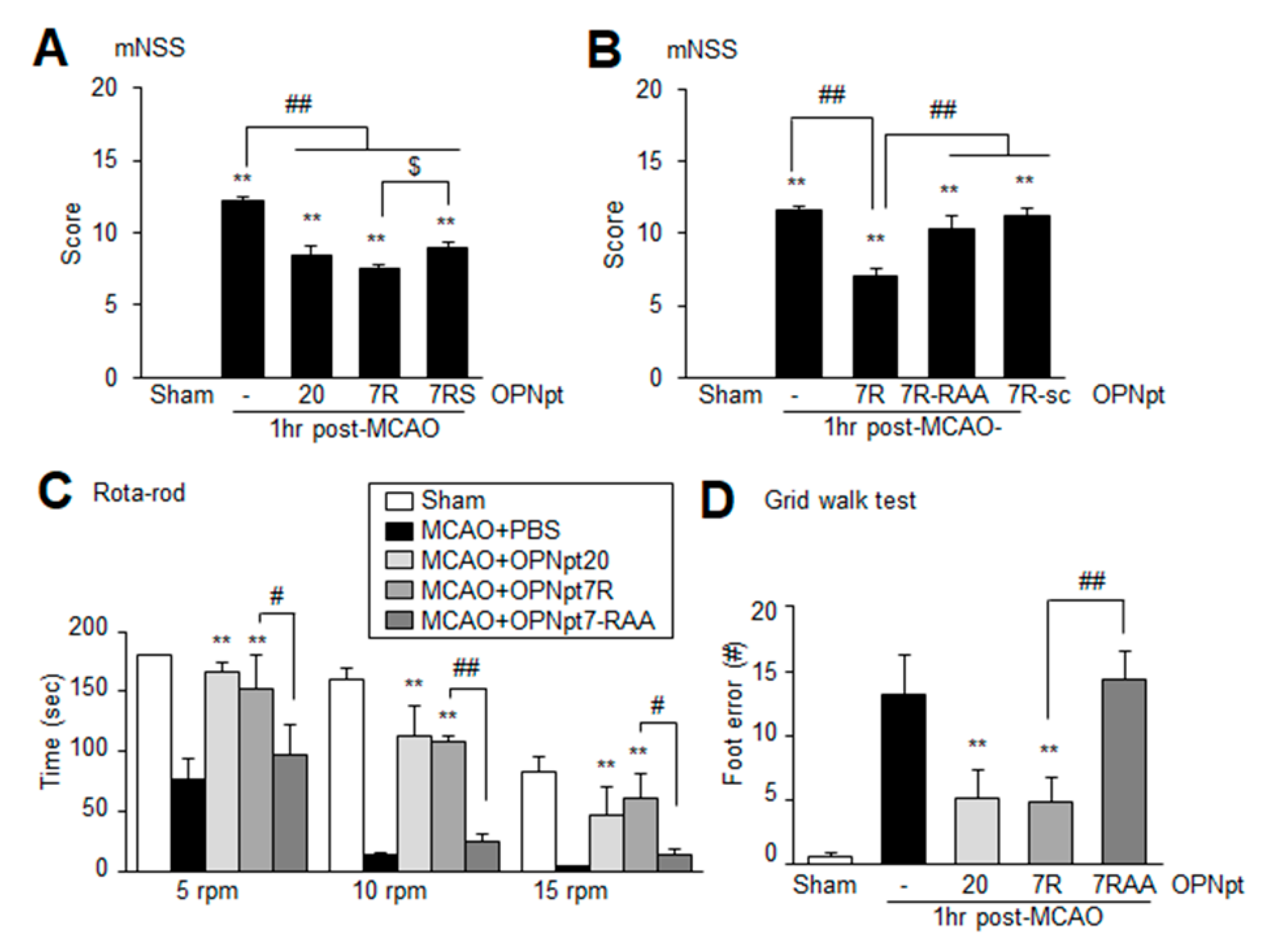
2.4. OPNpt7 Ameliorates Neurological Deficits and Motor Impairments
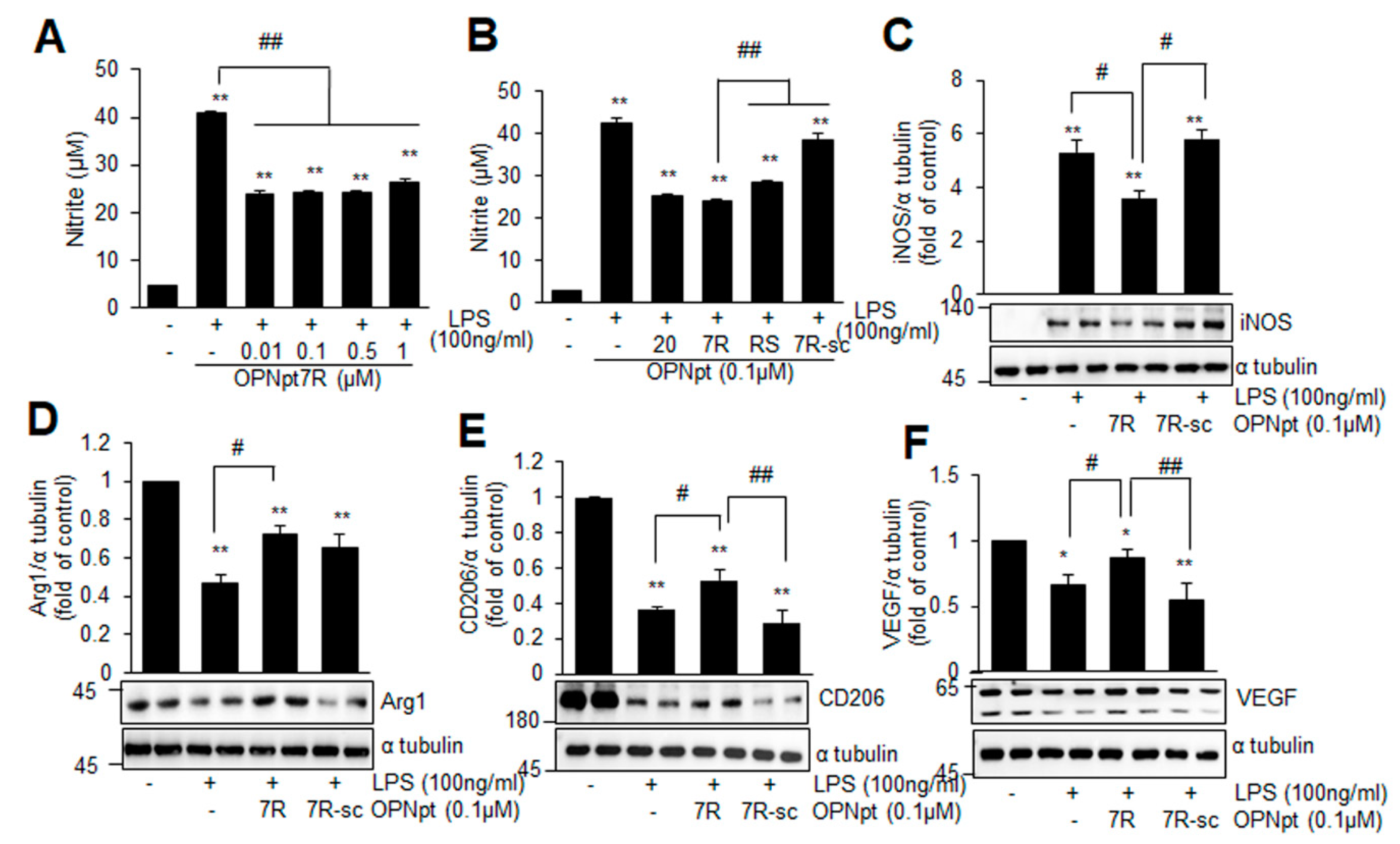
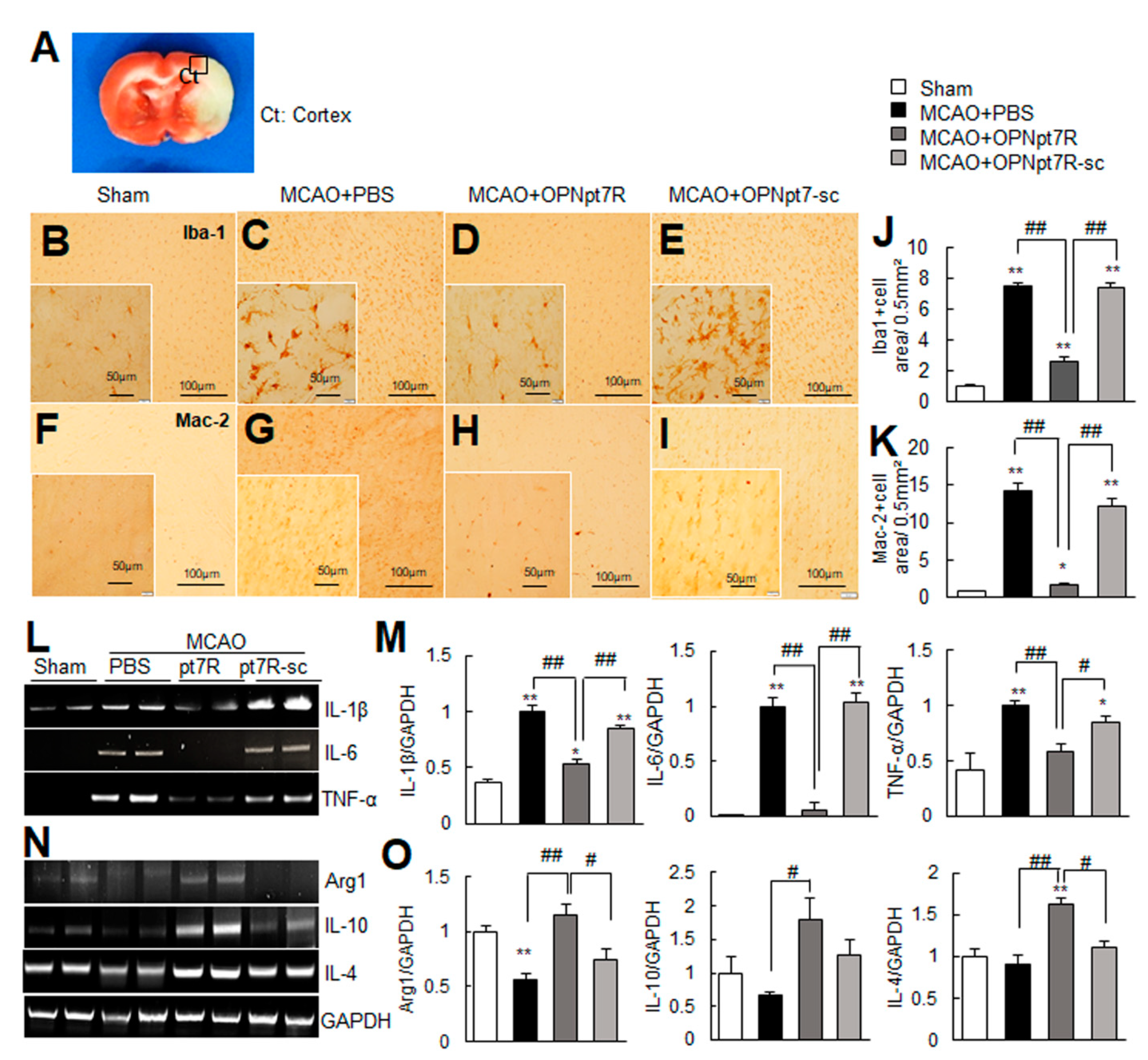
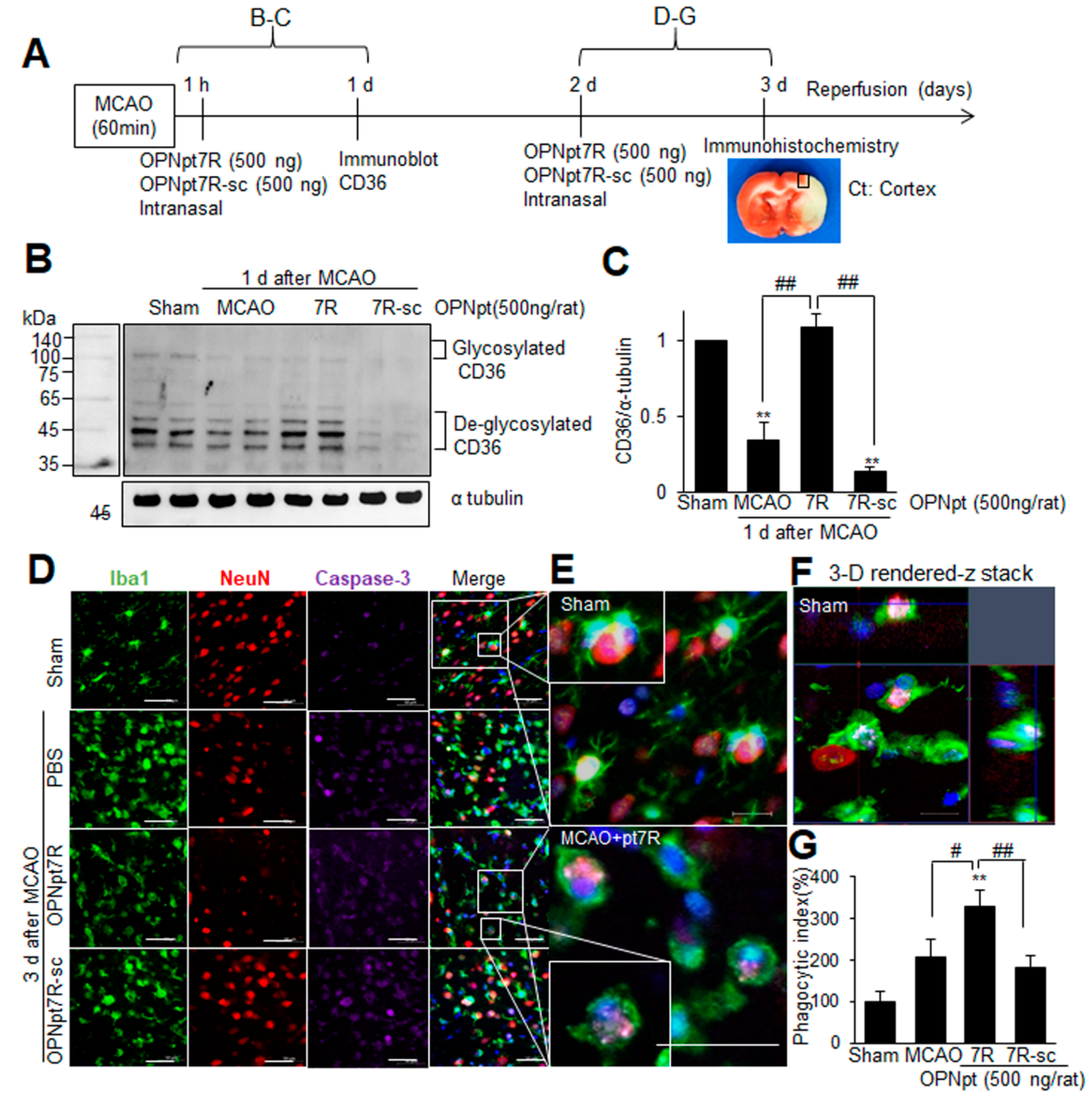
2.5. OPNpt7R Suppresses Activation of Microglial Cells and Augments M2 Polarization
2.6. Suppression of Inflammation and Induction of M2 Markers by OPNpt7R in the Post-Ischemic Brain
2.7. Enhancement of Phagocytic Activity of Microglia after Administration of OPNpt7R in the Post-Ischemic Brain
3. Discussion
4. Materials and Methods
4.1. Peptides
4.2. Surgical Procedure Used for Middle Cerebral Artery Occlusion
4.3. Intranasal Administration
4.4. Infarct Volume Assessment
4.5. Modified Neurological Severity Scores (mNSS)
4.6. Rotarod Test
4.7. Grid Walking Test
4.8. Primary Microglia Culture
4.9. Nitrite Measurements
4.10. Immunoblot Analysis
4.11. Immunohistochemistry
4.12. Immunofluorescence Staining
4.13. RNA Preparation and Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Denhardt, D.T.; Giachelli, C.M.; Rittling, S.R. Role of osteopontin in cellular signaling and toxicant injury. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 723–749. [Google Scholar] [CrossRef] [PubMed]
- Iczkiewicz, J.; Broom, L.; Cooper, J.D.; Wong, A.M.S.; Rose, S.; Jenner, P. The RGD-containing peptide fragment of osteopontin protects tyrosine hydroxylase positive cells against toxic insult in primary ventral mesencephalic cultures and in the rat substantia nigra. J. Neurochem. 2010, 114, 1792–1804. [Google Scholar] [CrossRef] [PubMed]
- Ailane, S.; Long, P.; Jenner, P.; Rose, S. Expression of integrin and CD44 receptors recognising osteopontin in the normal and LPS-lesioned rat substantia nigra. Eur. J. Neurosci. 2013, 38, 2468–2476. [Google Scholar] [CrossRef] [PubMed]
- Steinman, L.; Martin, R.; Bernard, C.; Conlon, P.; Oksenberg, J.R. Multiple Sclerosis: Deeper Understanding of Its Pathogenesis Reveals New Targets for Therapy. Annu. Rev. Neurosci. 2002, 25, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, Y.; Ota, K.; Ikeguchi, R.; Kubo, S.; Kabasawa, C.; Uchiyama, S. Plasma osteopontin levels are associated with disease activity in the patients with multiple sclerosis and neuromyelitis optica. J. Neuroimmunol. 2013, 263, 148–151. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, H.; Hasegawa, Y.; Kanamaru, K.; Zhang, J.H. Mechanisms of Osteopontin-Induced Stabilization of Blood-Brain Barrier Disruption After Subarachnoid Hemorrhage in Rats. Stroke 2010, 41, 1783–1790. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Ma, Q.; Suzuki, H.; Chen, C.; Liu, W.; Tang, J.; Zhang, J.H. Recombinant Osteopontin Attenuates Brain Injury after Intracerebral Hemorrhage in Mice. Neurocritical Care 2010, 14, 109–117. [Google Scholar] [CrossRef]
- Meller, R.; Stevens, S.L.; Minami, M.; Cameron, J.A.; King, S.; Rosenzweig, H.L.; Doyle, K.; Lessov, N.S.; Simon, R.P.; Stenzel-Poore, M.P. Neuroprotection by Osteopontin in Stroke. Br. J. Pharmacol. 2005, 25, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, M.; Zickler, P.; Denhardt, D.T.; Hartung, H.-P.; Jander, S. Increased thalamic neurodegeneration following ischaemic cortical stroke in osteopontin-deficient mice. Brain 2006, 129, 1426–1437. [Google Scholar] [CrossRef]
- Chen, W.; Ma, Q.; Suzuki, H.; Hartman, R.; Tang, J.; Zhang, J.H. Osteopontin Reduced Hypoxia–Ischemia Neonatal Brain Injury by Suppression of Apoptosis in a Rat Pup Model. Stroke 2011, 42, 764–769. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Kim, I.-Y.; Kim, I.-D.; Lee, H.-K.; Park, J.-Y.; Han, P.-L.; Kim, K.K.; Choi, H.; Lee, J.-K. Biodegradable gelatin microspheres enhance the neuroprotective potency of osteopontin via quick and sustained release in the post-ischemic brain. Acta Biomater. 2014, 10, 3126–3135. [Google Scholar] [CrossRef]
- Doyle, K.; Yang, T.; Lessov, N.S.; Ciesielski, T.M.; Stevens, S.L.; Simon, R.P.; King, J.S.; Stenzel-Poore, M.P. Nasal Administration of Osteopontin Peptide Mimetics Confers Neuroprotection in Stroke. Br. J. Pharmacol. 2008, 28, 1235–1248. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.C.; Lee, H.; Kim, S.W.; Kim, I.D.; Lee, H.K.; Lee, Y.; Han, P.L.; Lee, J.K. Intranasal Delivery of RGD Motif-Containing Osteopontin Icosamer Confers Neuroprotection in the Postischemic Brain via alphavbeta3 Integrin Binding. Mol. Neurobiol. 2016, 53, 5652–5663. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-D.; Lee, H.; Jin, Y.-C.; Lee, J.-K. Osteopontin Peptide Icosamer Containing RGD and SLAYGLR Motifs Enhances the Motility and Phagocytic Activity of Microglia. Exp. Neurobiol. 2017, 26, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Jin, Y.-C.; Kim, S.-W.; Kim, I.-D.; Lee, H.-K.; Lee, J.-K. Proangiogenic functions of an RGD-SLAY-containing osteopontin icosamer peptide in HUVECs and in the postischemic brain. Exp. Mol. Med. 2018, 50, e430. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, J.; Wang, Y.; Yang, G.-Y. The biphasic function of microglia in ischemic stroke. Prog. Neurobiol. 2017, 157, 247–272. [Google Scholar] [CrossRef]
- Jiang, C.; Wu, W.; Deng, Y.; Ge, J. Modulators of microglia activation and polarization in ischemic stroke (Review). Mol. Med. Rep. 2020, 21, 2006–2018. [Google Scholar] [CrossRef] [Green Version]
- Ladwig, A.; Walter, H.L.; Hucklenbroich, J.; Willuweit, A.; Langen, K.-J.; Fink, G.R.; Rueger, M.A.; Schroeter, M. Osteopontin Augments M2 Microglia Response and Separates M1- and M2-Polarized Microglial Activation in Permanent Focal Cerebral Ischemia. Mediat. Inflamm. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Shin, Y.-J.; Kim, H.L.; Choi, J.-S.; Choi, J.-Y.; Cha, J.-H.; Lee, M.-Y. Osteopontin: Correlation with phagocytosis by brain macrophages in a rat model of stroke. Glia 2011, 59, 413–423. [Google Scholar] [CrossRef]
- Rentsendorj, A.; Sheyn, J.; Fuchs, D.T.; Daley, D.; Salumbides, B.C.; Schubloom, H.E.; Hart, N.J.; Li, S.; Hayden, E.Y.; Teplow, D.B.; et al. A novel role for osteopontin in macrophage-mediated amyloid-beta clearance in Alzheimer’s models. Brain Behav. Immun. 2018, 67, 163–180. [Google Scholar] [CrossRef] [Green Version]
- Dashdulam, D.; Kim, I.-D.; Lee, H.; Lee, H.-K.; Kim, S.-W.; Lee, J.-K. Osteopontin heptamer peptide containing the RGD motif enhances the phagocytic function of microglia. Biochem. Biophys. Res. Commun. 2020, 524, 371–377. [Google Scholar] [CrossRef]
- Takahashi, K.; Takahashi, F.; Tanabe, K.K.; Takahashi, H.; Fukuchi, Y. The carboxyl-terminal fragment of osteopontin suppresses arginine-glycine-asparatic acid-dependent cell adhesion. Biochem. Mol. Boil. Int. 1998, 46, 1081–1092. [Google Scholar] [CrossRef]
- Hamada, Y.; Nokihara, K.; Okazaki, M.; Fujitani, W.; Matsumoto, T.; Matsuo, M.; Umakoshi, Y.; Takahashi, J.; Matsuura, N. Angiogenic activity of osteopontin-derived peptide SVVYGLR. Biochem. Biophys. Res. Commun. 2003, 310, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Yuki, K.; Okazaki, M.; Fujitani, W.; Matsumoto, T.; Hashida, M.K.; Harutsugu, K.; Nokihara, K.; Daito, M.; Matsuura, N.; et al. Osteopontin-derived Peptide SVVYGLR Induces Angiogenesis In Vivo. Dent. Mater. J. 2004, 23, 650–655. [Google Scholar] [CrossRef] [Green Version]
- Uchinaka, A.; Kawaguchi, N.; Hamada, Y.; Mori, S.; Miyagawa, S.; Saito, A.; Sawa, Y.; Matsuura, N. Transplantation of myoblast sheets that secrete the novel peptide SVVYGLR improves cardiac function in failing hearts. Cardiovasc. Res. 2013, 99, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchinaka, A.; Hamada, Y.; Mori, S.; Miyagawa, S.; Saito, A.; Sawa, Y.; Matsuura, N.; Yamamoto, H.; Kawaguchi, N. SVVYGLR motif of the thrombin-cleaved N-terminal osteopontin fragment enhances the synthesis of collagen type III in myocardial fibrosis. Mol. Cell. Biochem. 2015, 408, 191–203. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-S.; Kim, H.-Y.; Cha, J.-H.; Choi, J.-Y.; Lee, M.-Y. Transient microglial and prolonged astroglial upregulation of osteopontin following transient forebrain ischemia in rats. Brain Res. 2007, 1151, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Louden, C.; Yue, T.-L.; Ellison, J.A.; Barone, F.C.; Solleveld, H.A.; Feuerstein, G.Z. Delayed Expression of Osteopontin after Focal Stroke in the Rat. J. Neurosci. 1998, 18, 2075–2083. [Google Scholar] [CrossRef]
- Borges, K.; Gearing, M.; Rittling, S.; Sørensen, E.S.; Kotloski, R.; Denhardt, D.T.; Dingledine, R. Characterization of osteopontin expression and function after status epilepticus. Epilepsia 2008, 49, 1675–1685. [Google Scholar] [CrossRef] [Green Version]
- Maetzler, W.; Berg, D.; Schalamberidze, N.; Melms, A.; Schott, K.; Mueller, J.C.; Liaw, L.; Gasser, T.; Nitsch, C. Osteopontin is elevated in Parkinson’s disease and its absence leads to reduced neurodegeneration in the MPTP model. Neurobiol. Dis. 2007, 25, 473–482. [Google Scholar] [CrossRef]
- Wu, X.-L.; Lin, K.-J.; Bai, A.-P.; Wang, W.-X.; Meng, X.-K.; Su, X.-L.; Hou, M.-X.; Dong, P.-D.; Zhang, J.-J.; Wang, Z.-Y.; et al. Osteopontin knockdown suppresses the growth and angiogenesis of colon cancer cells. World J. Gastroenterol. 2014, 20, 10440–10448. [Google Scholar] [CrossRef]
- Ortiz-Martínez, F.; Sanmartín, E.; Pomares-Navarro, E.; Perez-Balaguer, A.; Andrés, L.; Sánchez-Payá, J.; Aranda, F.I.; Lerma, E.; Peiró, G. Osteopontin Regulates VEGFA and ICAM-1 mRNA Expression in Breast Carcinoma. Am. J. Clin. Pathol. 2015, 143, 812–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, S.A.; Boddeke, H.W.G.M.; Kettenmann, H. Microglia in Physiology and Disease. Annu. Rev. Physiol. 2017, 79, 619–643. [Google Scholar] [CrossRef]
- Rabenstein, M.; Vay, S.U.; Flitsch, L.J.; Fink, G.R.; Schroeter, M.; Rueger, M.A. Osteopontin directly modulates cytokine expression of primary microglia and increases their survival. J. Neuroimmunol. 2016, 299, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, A.; Rogall, R.; Hucklenbroich, J.; Willuweit, A.; Schoeneck, M.; Langen, K.-J.; Fink, G.R.; Rueger, M.A.; Schroeter, M. Osteopontin Attenuates Secondary Neurodegeneration in the Thalamus after Experimental Stroke. J. Neuroimmune Pharmacol. 2018, 14, 295–311. [Google Scholar] [CrossRef] [PubMed]
- Esposito, E.; Hayakawa, K.; Ahn, B.J.; Chan, S.J.; Xing, C.; Liang, A.C.; Kim, K.-W.; Arai, K.; Lo, E.H. Effects of ischemic post-conditioning on neuronal VEGF regulation and microglial polarization in a rat model of focal cerebral ischemia. J. Neurochem. 2018, 146, 160–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, R.; Shen, Q.; Xu, P.; Luo, J.; Tang, Y. Phagocytosis of Microglia in the Central Nervous System Diseases. Mol. Neurobiol. 2014, 49, 1422–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuch, K.; Wanko, B.; Ambroz, K.; Castelo-Rosa, A.; Moreno-Viedma, V.; Grün, N.G.; Leitner, L.; Staffler, G.; Zeyda, M.; Stulnig, T.M. Osteopontin affects macrophage polarization promoting endocytic but not inflammatory properties. Obesity 2016, 24, 1489–1498. [Google Scholar] [CrossRef]
- Kim, J.-B.; Choi, J.S.; Min-Hyung, L.; Nam, K.; Piao, C.-S.; Kim, S.-W.; Lee, M.-H.; Han, P.-L.; Park, J.-S.; Lee, J.-K. HMGB1, a Novel Cytokine-Like Mediator Linking Acute Neuronal Death and Delayed Neuroinflammation in the Postischemic Brain. J. Neurosci. 2006, 26, 6413–6421. [Google Scholar] [CrossRef] [Green Version]
- Kim, I.-D.; Shin, J.-H.; Kim, S.-W.; Choi, S.; Ahn, J.; Han, P.-L.; Park, J.-S.; Lee, J.-K. Intranasal Delivery of HMGB1 siRNA Confers Target Gene Knockdown and Robust Neuroprotection in the Postischemic Brain. Mol. Ther. 2012, 20, 829–839. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Sanberg, P.R.; Li, Y.; Wang, L.; Lu, M.; Willing, A.E.; Sanchez-Ramos, J.; Chopp, M. Intravenous Administration of Human Umbilical Cord Blood Reduces Behavioral Deficits After Stroke in Rats. Stroke 2001, 32, 2682–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.-B.; Lim, C.-M.; Yu, Y.-M.; Lee, J.-K. Induction and subcellular localization of high-mobility group box–1 (HMGB1) in the postischemic rat brain. J. Neurosci. Res. 2008, 86, 1125–1131. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PBS-Treated Group | OPNpt7R-Treated Group | |||
|---|---|---|---|---|
| Base | During Ischemia | Base | During Ischemia | |
| pH | 7.6 ± 0.1 | 7.6 ± 0.2 | 7.5 ± 0.2 | 7.5 ± 0.7 |
| pO2, mmHg | 146.8 ± 2.1 | 151 ± 4.0 | 144.6 ± 2.1 | 146.2 ± 6.4 |
| pCO2, mmHg | 35.9 ± 0.5 | 33.2 ± 2.4 | 35.2 ± 2.2 | 33.9 ± 2.6 |
| Glucose, mg/dL | 109.4 ± 3.5 | 100.6 ± 5.1 | 114 ± 5.7 | 97.6 ± 7.8 |
| Temperature | 36.6 ± 0.7 | 37.8 ± 1.7 | 36.5 ± 2.5 | 37.5 ± 0.6 |
| Gene (GenBank Accession No.) | Oligonucleotide Primer Sequences | PCR Product Size (bp) | Tm | Cycle |
|---|---|---|---|---|
| IL-1β (M98820) | 5′-AGC ATC CAG CTT CAA ATC TCA-3′ 5′-CGA GGC ATT TTT GTT GTT CAT-3′ | 268 | 54 | 25 |
| IL-6 (M26744) | 5′-CAA GAG ACT TCC AGC GAG TTG-3′ 5′-GAA ACG GAA CTC CAG AAG ACC-3′ | 350 | 54 | 25 |
| TNF-α (NM012675) | 5′-CTC CGT GAT GTC TAA GTA CT-3′ 5′-CTC AAA ACT CGA GTG ACA AG-3′ | 422 | 54 | 25 |
| IL-10 (XM006249712) | 5′-CTT TCA CTT GCC CTC ATC-3′ 5′-ACA AAC AAT ACG CCA TTC-3′ | 265 | 47 | 30 |
| Arg1 (AH002138) | 5′-CAG AAG AAT GGA AGA GTC AG-3′ 5′-CAG ATA TGC AGG GAG TCA CC-3′ | 524 | 54 | 30 |
| IL-4 (AY496861) | 5′-ACC TTG GTG TCA CCC TGT TCT GC-3′ 5′-GTT GTG AGC GTG GAC TCA TTC ACG-3′ | 292 | 55 | 30 |
| GAPDH (DQ403053) | 5′-TCATTGACCTCAACTACATGGT-3′ 5′-CTAAGCAGTTGGTGGTGCAG -3′ | 363 | 55 | 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davaanyam, D.; Kim, I.-D.; Lee, J.-K. Intranasal Delivery of RGD-Containing Osteopontin Heptamer Peptide Confers Neuroprotection in the Ischemic Brain and Augments Microglia M2 Polarization. Int. J. Mol. Sci. 2021, 22, 9999. https://doi.org/10.3390/ijms22189999
Davaanyam D, Kim I-D, Lee J-K. Intranasal Delivery of RGD-Containing Osteopontin Heptamer Peptide Confers Neuroprotection in the Ischemic Brain and Augments Microglia M2 Polarization. International Journal of Molecular Sciences. 2021; 22(18):9999. https://doi.org/10.3390/ijms22189999
Chicago/Turabian StyleDavaanyam, Dashdulam, Il-Doo Kim, and Ja-Kyeong Lee. 2021. "Intranasal Delivery of RGD-Containing Osteopontin Heptamer Peptide Confers Neuroprotection in the Ischemic Brain and Augments Microglia M2 Polarization" International Journal of Molecular Sciences 22, no. 18: 9999. https://doi.org/10.3390/ijms22189999