
Cystine/Glutamate Antiporter in Schizophrenia: From Molecular Mechanism to Novel Biomarker and Treatment


1
Institute of Clinical Medical Science, China Medical University, Taichung 40402, Taiwan
2
Department of Psychiatry, China Medical University Hospital, Taichung 40402, Taiwan
3
Graduate Institute of Biomedical Sciences, China Medical University, Taichung 40402, Taiwan
4
Department of Psychiatry, Kaohsiung Chang Gung Memorial Hospital, Chang Gung University College of Medicine, Kaohsiung 83301, Taiwan
5
School of Medicine, Chang Gung University, Taoyuan 33302, Taiwan
6
Department of Psychology, College of Medical and Health Sciences, Asia University, Taichung 41354, Taiwan
*
Authors to whom correspondence should be addressed.
†
Chieh-Hsin Lin and Hsien-Yuan Lane contributed equally in this manuscript as corresponding authors.
Int. J. Mol. Sci. 2021, 22(18), 9718; https://doi.org/10.3390/ijms22189718
Submission received: 26 July 2021
/
Revised: 6 September 2021
/
Accepted: 6 September 2021
/
Published: 8 September 2021
(This article belongs to the Collection Feature Papers in Molecular Pharmacology)
{kind=link}
Abstract
:Glutamate, a crucial excitatory neurotransmitter, plays a major role in the modulation of schizophrenia’s pathogenesis. New drug developments for schizophrenia have been prompted by the hypoglutamatergic hypothesis of schizophrenia. The cystine/glutamate antiporter system xc− is related to glutamate-release regulation. Patients with schizophrenia were recently discovered to exhibit downregulation of xc− subunits—the solute carrier (SLC) family 3 member 2 and the SLC family 7 member 11. We searched for relevant studies from 1980, when Bannai and Kitamura first identified the protein subunit system xc− in lung fibroblasts, with the aim of compiling the biological, functional, and pharmacological characteristics of antiporter xc−, which consists of several subunits. Some of them can significantly stimulate the human brain through the glutamate pathway. Initially, extracellular cysteine activates neuronal xc−, causing glutamate efflux. Next, excitatory amino acid transporters enhance the unidirectional transportation of glutamate and sodium. These two biochemical pathways are also crucial to the production of glutathione, a protective agent for neural and glial cells and astrocytes. Investigation of the expression of system xc− genes in the peripheral white blood cells of patients with schizophrenia can facilitate better understanding of the mental disorder and future development of novel biomarkers and treatments for schizophrenia. In addition, the findings further support the hypoglutamatergic hypothesis of schizophrenia.
1. Introduction
Schizophrenia is a chronic brain disease affecting approximately 1% of the world population that causes a severe health burden [1,2]. Many theories and studies have argued for the role of an imbalance in serial neurotransmitters, such as dopamine and glutamate, in the pathophysiology of schizophrenia. In all these studies, increased dopaminergic and decreased glutamatergic neurotransmissions have been hypothesized to play crucial roles in the neuropsychiatric etiology of schizophrenia [3,4]. The symptoms of schizophrenia include psychotic episodes, such as delusions and hallucinations, and cognitive impairment, leading to social withdrawal and a lack of motivation [5]. People with schizophrenia experience mental illness and disability during the course of the disease, leading to dysfunction in their daily routine and lower life expectancy. Investigation of the underlying etiology of the disease can assist in the future development of diagnostic tools and treatment options.
Evidence concerning neurodevelopmental origin as well as genetic evidence suggest concordance with regard to interference in polygenic inheritance by N-methyl-D-aspartate (NMDA) neurotransmission in schizophrenia [5]. The human brain develops during adolescence, the period in which many schizophrenia symptoms appear or worsen, rendering the patients subject to suffering from the deterioration of executive performance, cognitive function and social interaction throughout their adulthood [6]. In addition, the genetic predisposition and environmental disturbances that lead to changes or imbalances in these developmental processes may also increase the schizophrenia risk [7].
Not only hyperdopaminergic but also hypoglutamatergic hypotheses of schizophrenia pathophysiology have been studied and tested [4]. Moreover, the literature shows that patients with schizophrenia may display an abnormal mechanism in antioxidant protection in peripheral blood [8,9,10], cerebrospinal fluid (CSF) [11], and postmortem brain tissues [12,13]. Evidence also suggests that patients with schizophrenia tend to have genetic bases that result in decreased appropriate antioxidant defense systems [14]. Overall, the biomolecular pathophysiology of schizophrenia is involved in many fields, from neurotransmission to oxidative stresses.
The protein subunit system xc−, related to glutamate, is composed of a heavy-chain subunit 4F2hc, the solute carrier (SLC) family 3 member 2 (SLC3A2), the light-chain subunit xCT, and the SLC family 7 member 11 (SLC7A11) [15]. Cystine is reduced to cysteine intracellularly after its incorporation into system xc−. Cysteine is the rate-limiting substrate in the biosynthesis of the antioxidant glutathione (GSH), which is one of the major antioxidants in the brain [16]. The aforementioned pathways are involved in the cortical function through glutamatergic stimulation in the human brain. Two types of glutamate receptors are present, namely metabotropic and ionotropic receptors. The metabotropic glutamatergic receptors (mGluRs) are crucial in the initiation and modulation of glutamate neurotransmission and are composed of guanine nucleotide-binding G protein. The mGluRs are activated by glutamate to release guanosine diphosphate (GDP). Activated mGluRs can therefore influence and modulate the enzyme functions, ion channels, and vesical transports. The mGluRs are divided into eight subtypes and classified into three groups on the basis of their signaling pathways, pharmacological properties, and DNA sequences. Group 1 mGluRs increase the presynaptic glutamate release and the signaling cascade involved in phospholipase C, which cleaves phosphatidylinositol-4-5 bisphosphate into diacylglycerol and inositol 1,4,5-triphosphate, resulting in calcium release. In contrast, group 2 and group 3 mGluRs both induce the interactions with G1/o species, and the signaling proteins include adenyl cyclase, creating cyclic adenosine monophosphate (cAMP). The above mGluRs have been found at presynaptic glutamate terminals and GABA interneurons. Activation of mGluRs facilitates the potentiation of NMDA receptor (NMDAR) currents. Therefore, NMDARs are depolarized, resulting in the activation of a serine/threonine protein phosphatase that dephosphorylates mGluR and depolarizes it again [5]. Glutamate outside the cell membrane should be controlled and regulated to enhance appropriate neurotransmission. The mechanism ensures fluctuation of the altered activity of metabotropic and ionotropic glutamate receptors, resulting in cognitive processing and behavioral manifestation.
The relationship between xc− and schizophrenia pathogenesis was investigated in a recent study [17], in which the mRNA expression of SLC7A11 and SLC3A2 in peripheral white blood cells was discovered to be lower in patients with schizophrenia than in healthy people. The laboratory findings were consistent with the hypothesis of hypoglutamatergic neurotransmission in schizophrenia pathophysiology. We suggest that physiological markers could play diagnostic and therapeutic roles in patients with schizophrenia.
2. Biomolecular Mechanisms Involved in the Hypoglutamatergic Hypothesis of Schizophrenia
2.1. System xc− Protein and Its Genetic Modulation in Schizophrenia Pathophysiology
In 1980, the subunit system xc− protein was first identified in human fetal lung fibroblasts by Bannai and Kitamura [18]. The system xc− is a sodium-independent and chloride-dependent antiporter of the anionic forms of cystine and glutamate. It is composed of a heavy-chain subunit (4F2hc and SLC3A2) and a light-chain subunit (xCT and SLC7A11) [15,19]. Inside its protein structure, cystine is transformed into cysteine in a 1:1 ratio, and cysteine serves as the rate-limiting substrate for the biosynthesis of GSH, which is one of the major antioxidants in the brain [16]. Since decreased GSH was observed in older humans, GSH was postulated to play as a key role in the cognitive deficiency of aged people. [20]. The influence of the aforementioned system xc− on the concentration inside human brain cells was hypothesized to play an important role in the modulation of many neurotransmitter pathways.
The concentration of system xc− has been found to be higher in human brain astrocyte cells than in other brain cells [21,22,23]. The modulation of system xc− expression has been related to many neurological and psychiatric disorders [24,25,26], including schizophrenia. Different expressions of system xc− subunits are documented in cancer, immune responses, and neurodegenerative diseases [26]. In addition, the postmortem brain of a patient with schizophrenia showed a high system xc− protein level in the dorsolateral prefrontal cortex compared with control brains without considerable changes in the anterior cingulate cortex and hippocampus [27]. Since system xc− plays a critical role in glutamate release and the hypoglutamatergic hypothesis concerns the contribution to schizophrenia, it has been hypothesize that system xc− may be involved in the pathogenesis of schizophrenia.
Recently, we investigated the mRNA expression of system xc− obtained from peripheral blood of patients with schizophrenia. To develop convenient and efficient diagnostic tools for schizophrenia, such as peripheral gene expression as a useful surrogate for gene expression in the central nervous system (CNS), we planned to examine whether the expression of the two system xc− subunits is altered in patients with schizophrenia. Furthermore, we wanted to know whether this expression can serve as a surrogate diagnostic biomarker. We thus measured the mRNA expression levels of SLC3A2 and SLC7A11 in peripheral white blood cells in well-characterized, unrelated patients with schizophrenia and healthy controls. We eliminated confounding factors caused by antipsychotic agents used in schizophrenia treatment. Altered SLC3A2 and SLC7A11 gene expression was found to be associated with system xc− impairment [17]. The results showed that the two subunits in system xc− were less abundant in patients with schizophrenia. In addition, lower activity of system xc− was discovered to be associated with the reduction of extracellular glutamate.
The mRNA levels of the two system xc− subunits, SLC3A2 and SLC7A11, were not significantly different between drug-free patients and medicated patients. Such a limitation implies that psychotropics potentially affect the expression of these subunits, and further evaluation is required. Laboratory analysis revealed that patients with schizophrenia tend to have significantly decreased mRNA expression for the aforementioned genetic particles.
2.2. Relationship between GSH and System xc−
GSH is a free radical reactive oxygen species (ROS) scavenger that modulates cell metabolism. Oxidative stress causes an imbalance between ROS and antioxidant defense systems in human tissues and body fluids. Therefore, excitotoxicity resulting from oxidative stressors has been postulated to be involved in the pathogenesis of most neurodegenerative diseases. GSH is a tripeptide consisting of the amino acids glutamate, glycine, and cysteine. Inside a somatic cell, GSH plays a crucial role in antioxidation, particularly through the mechanism of cystine/glutamate antiporter system xc− [26]. Cystine is reduced to cysteine either by intracellular GSH through the formation of a mixed disulfide intermediate or by thioredoxin reductase 1 [28]. Overall, excitatory amino acid transporters are considered to play a role in cysteine importation [29].
GSH is oxidized into GSH disulfide and then either reduced by GSH reductase in a reaction requiring the reduced nicotinamide adenine dinucleotide phosphate (NADPH) or exported from the cell by multiple-drug-resistant proteins [15]. NADPH is generated through the hexose monophosphate shunt, an alternative pathway of glucose metabolism. GSH is formed by GSH S-transferases, which are formed as secondary metabolites during cell metabolism and electrophilic xenobiotics [15,30].
2.3. Role of Glutamate in GSH and System xc−
Glutamate receptors are divided into two types, namely ionotropic and metabotropic receptors. The concentration of extracellular glutamate is regulated for effective neurotransmission in the brain network. Fluctuation in the extracellular glutamate concentration can lead to altered activity of metabotropic and ionotropic glutamate receptors and subsequent changes in neuronal activity, leading to the change of cognitive function and subsequent behavioral manifestations [26].
Ionotropic receptors are composed of synthetic glutamate derivatives, including NMDA and 2-amino-3-(5-methyl-3-oxo-1, 2-oxazol-4-yl) propanoic acid (AMPA). Both NMDA and AMPA receptors possess the excise channel that is stimulated for glutamate. In addition to the original glutamate binding site, NMDARs have a binding site for endogenous modulatory amino acids such as glycine and D-serine. This binding site is polyamine and redox sensitive, assisting in glutamate stimulation and neurotransmission for the integration of brain cognitive function. The neurotransmission glutamate pathway in the brain modulates its activity at multiple ionotropic and metabotropic receptors, assisting its function in human brain plasticity of ensuring learning and memory. The mechanism also influences cognitive processes and human behavior.
Extracellular glutamate concentrations are postulated to be tightly regulated to ensure adequate neurotransmission. Hence, glutamate neurotransmission represents a modulation system for controlling human neuronal plasticity, learning, and memory [31]. A literature review reported deficits in the working memory of a rodent model of schizophrenia. The above memory deficits were reversed by using the cystine prodrug N-acetylcysteine to promote the stimulation of system xc− [27]. As serial cognitive impairments, such as memory deficits and judgmental deterioration, are one of the main features of the disease course and psychopathology of schizophrenia, the effects of N-acetylcysteine are relevant to the hypoglutamatergic hypothesis of schizophrenia.
On the other hand, in a rodent phencyclidine (PCP) model of schizophrenia, deficits in working memory were reversed by using cystine prodrug N-acetylcysteine pretreatment to enhance and facilitate the activity of system xc− [27]. These lines of evidence implicate system xc− in schizophrenia pathogenesis through the regulation of extracellular glutamate and GSH, which can prevent the brain from oxidative damage [31]. Hence, the function of system xc− could be beneficial in reversing worsened cognition, deteriorated memory, and behavioral problems in patients with schizophrenia. In brief summation, system xc− may be implicated in schizophrenia pathogenesis through the regulation of extracellular glutamate and GSH. System xc− also plays a crucial role in the determination of the cognition and behavioral manifestations of schizophrenia. The investigation of xc− might lead us to a better understanding of the disease nature of schizophrenia.
2.4. Proposed and Tentative Biomarkers for Schizophrenia Diagnosis
According to the review of the literature, decreased mRNA expression of system xc− subunits SLC7A11 and SLC3A2 in patients with schizophrenia supports the hypoglutamatergic neurotransmission hypothesis regarding the pathogenesis of schizophrenia. A hypothesis of NMDA neurotransmission in schizophrenia has been proposed for some time, with a basis in evidence from and experience with the NMDAR antagonist PCP and the dissociative anesthetic ketamine, which induces psychiatric symptomatology resembling the clinical presentation of schizophrenia. Abnormal concentrations of glutamate and gamma-aminobutyric acid have been proposed to play important roles in schizophrenia. Hence, the current review indicates an effective association and provides clear evidence that system xc− subunits SLC7A11 and SLC3A2 may serve as important biomarkers for schizophrenia [32,33,34].
In addition, the upstream mRNA expression of SLC7A11 and SLC3A2 was related to β-amyloid expression in transgenic mice and an in vitro model of human Alzheimer’s disease [35]. A rat model of Parkinson’s disease revealed a relationship with system xc− subunits [36]. Most studies have indicated that system xc− subunits play a crucial role in the pathophysiology of neurological and psychiatric diseases [24,37,38,39,40,41,42,43]. Psychiatric diseases, including schizophrenia, are diagnosed on the basis of the clinician’s experience and face-to-face interviews. Hence, a biomarker would be essential as an objective tool for comprehensive validation of the diagnosis of psychiatric disorders. Appropriate and effective methods could support the accuracy and efficacy of disease diagnosis and treatment. From the perspective of the hypoglutamatergic hypothesis and experimental evidence, decreased mRNA expression levels of SLC7A11 and SLC3A2 in peripheral blood could be developed as accurate tools for diagnosing schizophrenia.
Four variants of SLC3A2 exist (namely transcript variants 2, 3, 5, and 6), and our previous study [17] specifically evaluated transcript variant 3 (NM_002394.5). Comprehensive and thorough studies can clarify whether all variants of SLC3A2 are related to schizophrenia’s pathophysiology and add to the current findings. The association between the extracellular glutamate concentration and mRNA expression levels of the two system xc− subunits was not clarified in the study. The confounding factors included the effects of additive food preservatives from daily food consumption in the Han-Chinese population. The peripheral mRNA expression levels of two system xc− subunits, SLC7A11 and SLC3A2 (particularly SLC3A2), are decreased in patients with schizophrenia. The conclusions of this study support the findings of other studies demonstrating the hypofunction of glutamate neurotransmission in schizophrenia [5,17].
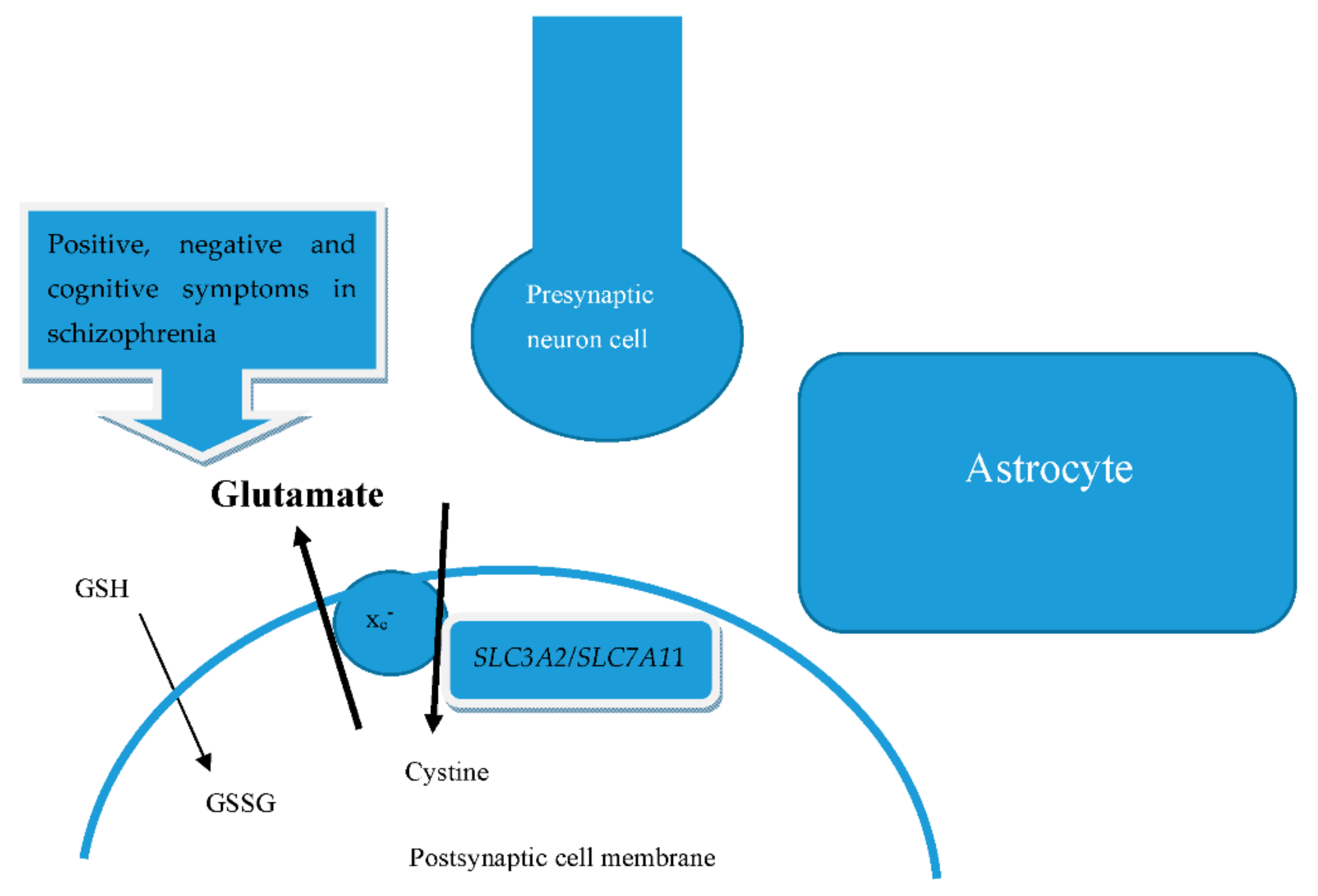
We show a summary of the mentioned biomolecular in Figure 1. Glutamate and cystine are transported bidirectionally via the integral membrane protein of the antiporter system by xc− subunits, regulated by mRNA SLC7A11/SLC3A2. Glutamate is hence abundant in the space formed by the presynaptic neuron cell, postsynaptic cell membrane, and astrocytes. The concentration of glutamate can therefore influence the positive, negative, and cognitive symptoms of schizophrenia. GSH also interferes with the cognitive function of schizophrenia.
3. Interplay between the Hypoglutamatergic Transmission and Oxidative Stress in the Etiology of Schizophrenia
NMDARs are regulated by agonists, coagonists, antagonists such as PCP and ketamine, and other molecules, including polyamines, proton, zinc, and magnesium [32]. While NMDARs can be affected by both endogenous and exogenous signals, they maintain synaptic plasticity and neuronal development in brain areas involving cognition, memory, and judgment, as well as psychosis [44,45]. NMDAR overactivation causes neurotoxicity, whereas its hypofunction results in neurodegeneration. As NMDAR hypofunction as shown to influence positive, negative, and cognitive symptoms in patients with schizophrenia, it was concluded that NMDA is an important neurotransmitter in schizophrenia [46]. Hence, NMDAR activity must be maintained within an appropriate range for human brain protection and mental health preservation [47].
Furthermore, the mechanism of increased oxidative stress contributes to the aging processes [46] and to neurodegenerative diseases through apoptotic decline in cells and tissues [12], whereas free radicals damage cells and tissues [48]. Antioxidants can assist in the prevention and reversal of cognitive deficits induced by free radicals [49]. A study has indicated cross-links among age-related NMDAR dysfunction, oxidative stress, senescence, and related forms of cognitive decline [50]. Among NMDA-related molecules, sodium benzoate works through the reduction of the activity of D-amino acid oxidase (DAO), a flavoenzyme of peroxisomes responsible for degrading D-serine and D-alanine [51,52]. Sodium benzoate can thereby inhibit ROS [53] and increase the activity of catalase, an antioxidant, in patients with schizophrenia [54]. From another perspective, pharmacological therapy with DAO results in depletion of D-serine, which attenuates NMDAR activity in cerebellar and hippocampal slices, hippocampal cell cultures, and retina preparations [55,56]. D-serine levels are decreased in the cerebrospinal fluid of drug-naive patients with schizophrenia [57]. Furthermore, both reduced brain serine racemase (SRR) and increased DAO protein levels may contribute to a decrease in CSF D-serine levels in schizophrenia [58].
In addition, a decrease in D-serine along with an increase in L-serine suggests a dysfunction in SRR activity [59] and reveals that DAO inhibitors can facilitate the effects of D-serine on prepulse inhibition (PPI) [60]. The PPI is one of the principle forms of information processing measured in schizophrenia patients and rodents treated with DAO inhibitors or NMDA antagonists. These results indicate that NMDA is associated with cognitive function in humans. Moreover, fluctuations and changes in the aforementioned SRR protein expression have been found in the postmortem brains of patients with schizophrenia [58,61,62]. Similarly, glycine levels have been found to be decreased in drug-free patients with schizophrenia and inversely correlated with the severity of negative symptoms [63]. Not only is the concentration of glycine increased but homocysteine has also been found to contribute to the pathophysiological base of schizophrenia [64]. Using high-dose glycine can damage the prepulse inhibition measure of the sensorimotor gating in humans, leading to the argument that glycine seemingly does not support cognition recovery or reservation [65].
Other regulators involved in D-serine metabolism—such as D-amino oxidase (DAO)/D-amino oxidase activator (DAOA) [58,62], the protein interacting with kinase C [66], and alanine–serine–cysteine transporter 1 [67]—are related to D-serine levels. Therefore, D-serine depletion was investigated and determined to be associated with NMDAR by its mediation of neurological functions and NMDAR-induced neurotoxicity, as well as NMDAR-dependent long-term potentiation (LTP), in many brain regions, particularly the hippocampus [57]. Brain glial cells, presumably astrocytes, pass through, with the availability of D-serine being involved in line with the deficits in synaptic learning and memory mechanisms that occur during the course of aging [68]. Overall, NMDA neurotransmission is crucial in normal human cognitive function as well as in many CNS and psychiatric disease models, forming a critical and novel hypothesis of the pathogenesis of schizophrenia.
In summary, medications acting on NMDARs can stimulate the glutamatergic reaction through various approaches and pathways in the human brain. A D-serine supplement can entirely reverse the effects of decreased NMDAR-mediated neurotransmission [55,56,68]. In human genetic studies, significant associations have been observed between the gene DAO [69] and the G72 (DAOA)/G30 [70] gene polymorphisms and schizophrenia in case–control association analyses. Additionally, the functions of these genes can be applied as therapeutic agents of schizophrenia. The therapeutic effects include not only the positive symptoms of schizophrenia but also negative symptoms or cognitive deficiencies in the course of the mental illness [71,72]. Moreover, these compounds may possess other mechanisms, such as antioxidant and mechanistic target of rapamycin (mTOR) effects, which are involved in many aspects of memory and cognition in the healthy human brain, as well as in the brains of patients with schizophrenia [71,73]. Therefore, modulators of glutamate receptors may be novel candidate targets for the treatment of refractory schizophrenia [71].
4. From the Invention of Biomarkers as Precision Medicine for Schizophrenia to Possible Treatment Options
The findings of the current review could lead to not only a tentative improvement in diagnostic skills but also the achievement of novel neuropharmacological therapy for schizophrenia. Neuropsychiatric drug development has opened new avenues that can help scholars understand the neurological basis of schizophrenia [7]. Although medical treatments have advanced for chronic psychiatric diseases, the diagnostic tools available remain limited. In addition, early detection and correct diagnosis of schizophrenia can decrease the disease’s severity and lead to better outcomes in patients. Advances in current neuropsychiatric pharmacology could benefit the clinical process of mental disease diagnosis and treatment outcomes. Methods for early diagnosis and improved treatment would also decrease the disease burden on societies and nations worldwide.
On the basis of the reviewed studies, we can briefly conclude that some of the candidate genes involved in the neurodevelopment and glutamate-associated signaling that are relevant to schizophrenia, such as DAO and G72 (DAOA), are directly involved in NMDA neurotransmission; the rest are indirectly related to NMDA synapses. If NMDA function can be modulated through augmentation or stimulation, poor NMDA function based on various vulnerabilities in the NMDA synapse can be reversed, regardless of its origin. Further evidence regarding the involvement of the glutamatergic system in schizophrenia would increase the understanding of the disease’s nature and open a window to a new generation of antipsychotic treatments. Considering each individual’s NMDA pathology or an accumulation of evidence on variant candidate genes, rather than any single gene, may serve as a general model for the pathogenesis of schizophrenia.
Nevertheless, on the basis of the studies reviewed herein, peripheral blood could be examined for mRNA expression of SLC7A11 and SLC3A2 because such expression is significantly lower in patients with schizophrenia than in healthy individuals. Furthermore, studies have suggested D-serine levels in peripheral blood as a diagnostic and therapeutic marker for schizophrenia [59]. Decreased mRNA expression of SLC7A11 and SLC3A2 is in accordance with the hypoglutamatergic hypothesis of schizophrenia. Hence, people with mental diseases and healthy individuals could be differentiated using well-designed biomarkers.
Our findings and review of previous studies provide new insights and perspectives for the validation of schizophrenia diagnostic tools. Newer drugs designed on the basis of the hypothesis of hypoglutamatergic and disturbed glutamate signaling in schizophrenia could contribute to crucial treatment choices in the future.
5. Conclusions
The genetic approaches to schizophrenia treatment are both competitive and challenging because the pathophysiology of schizophrenia is complex. Moreover, many environmental and epigenetic factors influence the nature of this disease and thus contribute to the heterogeneous manifestations of schizophrenia [44]. Regarding the hypoglutamatergic hypothesis, clinical trials have indicated that sarcosine and sodium benzoate may be more potent than glycine, D-serine, and D-cycloserine in relieving the psychopathological symptoms and signs of schizophrenia without adverse effects or safety concerns [29,54]. As the etiology of schizophrenia is multifactorial and heterogeneous [74], precise diagnosis is difficult. With the diagnostic tools for objective screening enabled by the use of the two system xc− subunits, we might be able to improve the disease outcome of schizophrenia in the future. In addition to D-serine, D-aspartate (another endogenous NMDAR agonist enriched in the developing brain of humans) has been found to be involved in schizophrenia’s pathogenesis [75]. Reduced activity of system xc− has been found to be linked to a reduction in extracellular glutamate levels [76]. The literature review suggests that mRNA expression of system xc− subunits is prominent in the brains of both mice [19] and humans [77]. As oxidative stress and excitotoxicity inside cells are among the key factors associated with the neurodegenerative process, system xc−, which modulates and regulates GSH and glutamate, has been postulated to play a major role in the pathogenesis of many CNS disorders in addition to schizophrenia [78]. In transgenic mice and in vitro models of Alzheimer’s disease, the gene expression of the two system xc− subunits SLC7A11 and SLC3A2 was found to be increased in the presence of β-amyloid [35]. Increased SLC7A11 protein expression was also discovered in the striatum in a Parkinson’s disease rat model [36]. System xc− activity was downregulated after nicotine administration in a rat model of addiction, and treatment with N-acetylcysteine to promote system xc− activity reduced the cigarette smoking frequency in humans [79].
To briefly conclude, we propose that system xc− subunits can serve as a diagnostic tool for schizophrenia based on the study of mRNA SLC7A11, and SLC3A2. Current studies focus on distinguishing potential biomarkers between medicated schizophrenia, drug-free schizophrenia, and healthy subjects. However, the course of schizophrenia is complicated and heterogeneous. Whether these biomarkers are significantly different in first-stage schizophrenia with refractory treatment or in other psychopathologies remains unclear. Additional studies with larger sample sizes for patients with first-episode schizophrenia and refractory treatment might be needed for clarification of the theory. Therefore, study of the mechanism through which system xc− could serve as a diagnostic tool, as well as a therapeutic target, for schizophrenia and other CNS disorders is crucial. Research on the mRNA expression of SLC7A11 and SLC3A2 has revealed that diagnostic tools for schizophrenia in a clinical setting must be validated. Moreover, future development of a novel therapeutic treatment for schizophrenia, based on this mechanism, is warranted.
Author Contributions
Writing, review of the article and draft preparation: C.-C.H., C.-H.L., H.-Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the China Medical University Hospital (DMR-109-246), the National Health Research Institutes (NHRI-EX109-10731NI), and the Ministry of Science and Technology (MOST 109-2314-B-039 -001; MOST 109-2314-B-039-039-MY3; MOST 110-2622-B039-001).
Conflicts of Interest
The authors declare no conflict of interest.
Nonstandard Abbreviations
| Abbreviation | Full name |
| AMPA | 2-amino-3-(5-methyl-3-oxo-1,2-oxazol-4-yl) propanoic acid |
| CAT | catalase |
| DAO | D-amino acid oxidase |
| DAOA | D-amino acid oxidase activator |
| GDP | guanosine diphosphate |
| GSH | glutathione |
| GSSG | glutathione disulfide |
| LTP | long-term potentiation |
| mTOR | mechanistic target of rapamycin |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| NMDA | N-methyl-D-aspartate |
| PCP | phencyclidine |
| PPI | prepulse inhibition |
| ROS | reactive oxygen species |
| SLC | solute carrier |
| SRR | serine racemase |
References
- Jablensky, A.; Sartorius, N.; Ernberg, G.; Anker, M.; Korten, A.; Cooper, J.E.; Day, R.; Bertelsen, A. Schizophrenia: Manifestations, incidence and course in different cultures A World Health Organization Ten-Country Study. Psychol. Med. Monogr. Suppl. 1992, 20, 1–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablensky, A. Epidemiology of schizophrenia: The global burden of disease and disability. Eur. Arch. Psychiatry Clin. Neurosci. 2000, 250, 274–285. [Google Scholar] [CrossRef]
- Howes, O.D.; McCutcheon, R.; Owen, M.J.; Murray, R. The Role of Genes, Stress, and Dopamine in the Development of Schizophrenia. Biol. Psychiatry 2016, 81, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-H.; Lane, H.-Y.; Tsai, G.E. Glutamate signaling in the pathophysiology and therapy of schizophrenia. Pharmacol. Biochem. Behav. 2012, 100, 665–677. [Google Scholar] [CrossRef]
- Lewis, A.D.; Lieberman, A.J. Catching Up on Schizophrenia: Natural History and Neurobiology. Neuron 2000, 28, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Keshavan, M.S.; Giedd, J.; Lau, J.Y.; Lewis, D.A.; Paus, T. Changes in the adolescent brain and the pathophysiology of psychotic disorders. Lancet Psychiatry 2014, 1, 549–558. [Google Scholar] [CrossRef]
- Michelson, A.M. Biological role of the superoxide anion radical and of superoxyde-dismutase in cellular metabolism. Comptes Rendus Seances Soc. Biol. Fil. 1976, 170, 1137–1146. [Google Scholar]
- Okusaga, O. Accelerated Aging in Schizophrenia Patients: The Potential Role of Oxidative Stress. Aging Dis. 2014, 5, 256–262. [Google Scholar] [CrossRef]
- Sirota, P.; Gavrieli, R.; Wolach, B. Overproduction of neutrophil radical oxygen species corre-lates with negative symptoms in schizophrenic patients: Parallel studies on neutrophil chemotaxis, superoxide production and bactericidal activity. Psychiatry Res. 2003, 121, 123–132. [Google Scholar] [CrossRef]
- Do, K.Q.; Trabesinger, A.; Kirsten-Krüger, M.; Lauer, C.J.; Dydak, U.; Hell, D.; Holsboer, F.; Boesiger, P.; Cuénod, M. Schizophrenia: Glutathione deficit in cerebrospinal fluid and prefrontal cortex in vivo. Eur. J. Neurosci. 2000, 12, 3721–3728. [Google Scholar] [CrossRef]
- Do, K.Q.; Cabungcal, J.H.; Frank, A.; Steullet, P.; Cuenod, M. Redox dysregulation, neurodevelopment, and schizophrenia. Curr. Opin. Neurobiol. 2009, 19, 220–230. [Google Scholar] [CrossRef]
- Yao, J.K.; Leonard, S.; Reddy, R.D. Increased nitric oxide radicals in postmortem brain from patients with schizophrenia. Schizophr. Bull. 2004, 30, 923–934. [Google Scholar] [CrossRef] [Green Version]
- Chowdari, K.V.; Bamne, M.N.; Nimgaonkar, V.L. Genetic Association Studies of Antioxidant Pathway Genes and Schizophrenia. Antioxid. Redox Signal. 2011, 15, 2037–2045. [Google Scholar] [CrossRef] [Green Version]
- Bridges, R.J.; Natale, N.R.; A Patel, S. System xc− cystine/glutamate antiporter: An update on molecular pharmacology and roles within the CNS. Br. J. Pharmacol. 2011, 165, 20–34. [Google Scholar] [CrossRef] [Green Version]
- Dringen, R.; Hirrlinger, J. Glutathione Pathways in the Brain. Biol. Chem. 2003, 384, 505–516. [Google Scholar] [CrossRef]
- Lin, C.H.; Lin, P.P.; Lin, C.Y.; Lin, C.H.; Huang, C.H.; Huang, Y.J.; Lane, H.Y. Decreased mRNA expression for the two subunits of system xc−, SLC3A2 and SLC7A11, in WBC in patients with schizophrenia: Evidence in support of the hypo-glutamatergic hypothesis of schizophrenia. J. Psychiatr. Res. 2016, 72, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Bannai, S.; Kitamura, E. Transport interaction of L-cystine and L-glutamate in human diploid fibroblasts in culture. J. Biol. Chem. 1980, 255, 2372–2376. [Google Scholar] [CrossRef]
- Sato, H.; Tamba, M.; Ishii, T.; Bannai, S. Cloning and expression of a plasma membrane cystine/glutamate exchange transporter composed of two distinct proteins. J. Biol. Chem. 1999, 274, 11455–11458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, H.; Shenvi, S.; Hagen, T.M.; Liu, R.-M. Glutathione Metabolism during Aging and in Alzheimer Disease. Ann. N. Y. Acad. Sci. 2004, 1019, 346–349. [Google Scholar] [CrossRef]
- Pow, D.V. Visualising the activity of the cystine-glutamate antiporter in glial cells using antibodies to aminoadipic acid, a selectively transported substrate. Glia 2001, 34, 27–38. [Google Scholar] [CrossRef]
- Sato, H.; Tamba, M.; Okuno, S.; Sato, K.; Keino-Masu, K.; Masu, M.; Bannai, S. Distribution of Cystine/Glutamate Exchange Transporter, System xc−, in the Mouse Brain. J. Neurosci. 2002, 22, 8028–8033. [Google Scholar] [CrossRef] [Green Version]
- Burdo, J.; Dargusch, R.; Schubert, D. Distribution of the cystine/glutamate antiporter system x− c in the brain, kidney, and duodenum. J. Histochem. Cytochem. 2006, 54, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massie, A.; Boillee, S.; Hewett, S.; Knackstedt, L.; Lewerenz, J. Main path and byways: Non-vesicular glutamate release by system xc− as an important modifier of glutamatergic neurotransmission. J. Neurochem. 2015, 135, 1062–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, R.; Lutgen, V.; Lobner, D.; Baker, D.A. Thinking Outside the Cleft to Understand Synaptic Activity: Contribution of the Cystine-Glutamate Antiporter (System xc−) to Normal and Pathological Glutamatergic Signaling. Pharmacol. Rev. 2012, 64, 780–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewerenz, J.; Hewett, S.J.; Huang, Y.; Lambros, M.; Gout, P.W.; Kalivas, P.W.; Massie, A.; Smolders, I.; Methner, A.; Pergande, M.; et al. The cystine/glutamate antiporter system xc− in health and disease: From molecular mechanisms to novel therapeutic opportunities. Antioxid. Redox Signal. 2013, 18, 522–555. [Google Scholar] [CrossRef] [Green Version]
- A Baker, D.; Madayag, A.; Kristiansen, L.; Meador-Woodruff, J.H.; Haroutunian, V.; Raju, I. Contribution of Cystine–Glutamate Antiporters to the Psychotomimetic Effects of Phencyclidine. Neuropsychopharmacology 2007, 33, 1760–1772. [Google Scholar] [CrossRef] [Green Version]
- Mandal, P.K.; Seiler, A.; Perisic, T.; Kölle, P.; Canak, A.B.; Förster, H.; Weiss, N.; Kremmer, E.; Lieberman, M.W.; Bannai, S.; et al. System xc− and thioredoxin reductase 1 cooperatively rescue glutathione deficiency. J. Biol. Chem. 2010, 285, 22244–22253. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Swanson, R. The glutamate transporters EAAT2 and EAAT3 mediate cysteine uptake in cortical neuron cultures. J. Neurochem. 2003, 84, 1332–1339. [Google Scholar] [CrossRef]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef]
- Dun, Y.; Mysona, B.; Van Ells, T.; Amarnath, L.; Ola, M.S.; Ganapathy, V.; Smith, S.B. Expression of the cystine-glutamate exchanger (xc−) in retinal ganglion cells and regulation by nitric oxide and oxidative stress. Cell Tissue Res. 2006, 324, 189–202. [Google Scholar] [CrossRef] [Green Version]
- Javitt, D.C.; Schoepp, D.D.; Kalivas, P.W.; Volkow, N.D.; A Zarate, C.; Merchant, K.; Bear, M.F.; Umbricht, D.; Hajos, M.; Potter, W.Z.; et al. Translating Glutamate: From Pathophysiology to Treatment. Sci. Transl. Med. 2011, 3, 102mr2. [Google Scholar] [CrossRef] [Green Version]
- Zink, M.; Correll, C.U. Glutamatergic agents for schizophrenia: Current evidence and perspectives. Expert Rev. Clin. Pharmacol. 2015, 8, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.-C.; Liu, C.-M.; Liu, Y.-L.; Chen, C.-W.; Chang, C.C.; Fann, C.S.J.; Chiou, J.-J.; Yang, U.-C.; Chen, C.-H.; Faraone, S.V.; et al. The DAO Gene Is Associated with Schizophrenia and Interacts with Other Genes in the Taiwan Han Chinese Population. PLoS ONE 2013, 8, e60099. [Google Scholar] [CrossRef] [Green Version]
- Qin, S.; Colin, C.; Hinners, I.; Gervais, A.; Cheret, C.; Mallat, M. System Xc- and Apolipoprotein E Expressed by Microglia Have Opposite Effects on the Neurotoxicity of Amyloid-beta Peptide 1–40. J. Neurosci. 2006, 26, 3345–3356. [Google Scholar] [CrossRef] [PubMed]
- Massie, A.; Schallier, A.; Mertens, B.; Vermoesen, K.; Bannai, S.; Sato, H.; Smolders, I.; Michotte, Y. Time-dependent changes in striatal xCT protein expression in hemi-Parkinson rats. NeuroReport 2008, 19, 1589–1592. [Google Scholar] [CrossRef]
- Fournier, M.; Monin, A.; Ferrari, C.; Baumann, P.S.; Conus, P.; Do, K. Implication of the glutamate–cystine antiporter xCT in schizophrenia cases linked to impaired GSH synthesis. NPJ Schizophr. 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, Y.; Nakaso, K.; Horikoshi, Y.; Morimoto, M.; Omotani, T.; Otsuki, A.; Inagaki, Y.; Sato, H.; Matsura, T. System xc− in microglia is a novel therapeutic target for post-septic neurological and psychiatric illness. Sci. Rep. 2019, 9, 7562. [Google Scholar] [CrossRef] [Green Version]
- Torres, V.I.; Vallejo, D.; Inestrosa, N.C. Emerging synaptic molecules as candidates in the etiology of neurological disorders. Neural Plast. 2017, 2017, 8081758. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Kharkar, P.S.; Nandave, M. Emerging roles of system [Formula: See text] antiporter and its inhibition in CNS disorders. Mol. Membr. Biol. 2015, 32, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Murrough, J.W.; Abdallah, C.; Mathew, S.J. Targeting glutamate signalling in depression: Progress and prospects. Nat. Rev. Drug Discov. 2017, 16, 472–486. [Google Scholar] [CrossRef] [PubMed]
- Ayers-Ringler, J.R.; Jia, Y.-F.; Qiu, Y.-Y.; Choi, D.-S. Role of astrocytic glutamate transporter in alcohol use disorder. World J. Psychiatry 2016, 6, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.J.; Gray, L.J.; Finkelstein, D.I.; Crouch, P.J.; Pow, D.; Pang, T.Y.; Li, S.; Smith, Z.M.; Francis, P.S.; Renoir, T.; et al. N-acetylcysteine modulates glutamatergic dysfunction and depressive behavior in Huntington’s disease. Hum. Mol. Genet. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawa, A.; Snyder, S.H. Schizophrenia: Neural Mechanisms for Novel Therapies. Mol. Med. 2003, 9, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bliss, T.V.; Collingridge, G.L. A synaptic model of memory: Long-term potentiation in the hippocampus. Nature 1993, 361, 31–39. [Google Scholar] [CrossRef]
- Lin, C.H.; Huang, Y.J.; Lin, C.J.; Lane, H.Y.; E Tsai, G. NMDA neurotransmission dysfunction in mild cognitive impairment and Alzheimer’s disease. Curr. Pharm. Des. 2014, 20, 5169–5179. [Google Scholar] [CrossRef]
- Huang, Y.J.; Lin, C.H.; Lane, H.Y.; E Tsai, G. NMDA neurotransmission dysfunction in behavioral and psychological symptoms of Alzheimer’s disease. Curr. Neuropharmacol. 2012, 10, 272–285. [Google Scholar] [CrossRef] [Green Version]
- Serrano, F.; Klann, E. Reactive oxygen species and synaptic plasticity in the aging hippocampus. Ageing Res. Rev. 2004, 3, 431–443. [Google Scholar] [CrossRef]
- Bickford, P.C.; Gould, T.; Briederick, L.; Chadman, K.; Pollock, A.; Young, D.; Shukitt-Hale, B.; Joseph, J. Antioxidant-rich diets improve cerebellar physiology and motor learning in aged rats. Brain Res. 2000, 866, 211–217. [Google Scholar] [CrossRef]
- Guidi, M.; Kumar, A.; Foster, T.C. Impaired attention and synaptic senescence of the prefrontal cortex involves redox regulation of NMDA receptors. J. Neurosci. 2015, 35, 3966–3977. [Google Scholar] [CrossRef] [Green Version]
- Fukui, K.; Miyake, Y. Molecular cloning and chromosomal localization of a human gene encoding D-amino-acid oxidase. J. Biol. Chem. 1992, 267, 18631–18638. [Google Scholar] [CrossRef]
- Sasabe, J.; Miyoshi, Y.; Suzuki, M.; Mita, M.; Konno, R.; Matsuoka, M.; Hamase, K.; Aiso, S. D-Amino acid oxidase controls motoneuron degeneration through D-serine. Proc. Natl. Acad. Sci. USA 2011, 109, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Modi, K.K.; Roy, A.; Brahmachari, S.; Rangasamy, S.B.; Pahan, K. Cinnamon and Its Metabolite Sodium Benzoate Attenuate the Activation of p21rac and Protect Memory and Learning in an Animal Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0130398. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Lin, C.H.; Chang, Y.C.; Huang, Y.J.; Chen, P.W.; Yang, H.T.; Lane, H.Y. Sodium benzoate, a D-amino acid oxidase inhibitor, added to clozapine for the treatment of schizophrenia: A randomized, double-blind, placebo-controlled trial. Biol. Psychiatry 2018, 84, 422–432. [Google Scholar] [CrossRef]
- Mothet, J.P.; Parent, A.T.; Wolosker, H.; Brady, R.O.; Linden, D.J.; Ferris, C.D.; Rogawski, M.A.; Snyder, S.H. D-serine is an endogenous ligand for the glycine site of the N-methyl-D-aspartate receptor. Proc. Natl. Acad. Sci. USA 2000, 97, 4926–4931. [Google Scholar] [CrossRef] [Green Version]
- Gustafson, E.C.; Stevens, E.R.; Wolosker, H.; Miller, R.F. Endogenous d-Serine Contributes to NMDA-Receptor–Mediated Light-Evoked Responses in the Vertebrate Retina. J. Neurophysiol. 2007, 98, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Engberg, G.; Shimizu, E.; Nordin, C.; Lindström, L.H.; Iyo, M. Reduced D-serine to total serine ratio in the cerebrospinal fluid of drug naive schizophrenic patients. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2005, 29, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Bendikov, I.; Nadri, C.; Amar, S.; Panizzutti, R.; De Miranda, J.; Wolosker, H.; Agam, G. A CSF and postmortem brain study of D-serine metabolic parameters in schizophrenia. Schizophr. Res. 2007, 90, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Fukushima, T.; Shimizu, E.; Komatsu, N.; Watanabe, H.; Shinoda, N.; Nakazato, M.; Kumakiri, C.; Okada, S.; Hisanori, H. Decreased serum levels of D-serine in patients with schizophrenia: Evidence in support of the N-methyl-D-aspartate receptor hypofunction hypothesis of schizophrenia. Arch. Gen. Psychiatry 2003, 60, 572–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, K.; Fujita, Y.; Horio, M.; Kunitachi, S.; Iyo, M.; Ferraris, D.; Tsukamoto, T. Co-administration of a D-amino acid oxidase inhibitor potentiates the efficacy of D-serine in attenuating prepulse inhibition deficits after administration of dizocilpine. Biol. Psychiatry 2009, 65, 1103–1106. [Google Scholar] [CrossRef] [PubMed]
- Steffek, A.E.; Haroutunian, V.; Meador-Woodruff, J.H. Serine racemase protein expression in cortex and hippocampus in schizophrenia. Neuroreport 2006, 17, 1181–1185. [Google Scholar] [CrossRef] [PubMed]
- Verrall, L.; Walker, M.; Rawlings, N.; Benzel, I.; Kew, J.N.C.; Harrison, P.J.; Burnet, P.W.J. d-Amino acid oxidase and serine racemase in human brain: Normal distribution and altered expression in schizophrenia. Eur. J. Neurosci. 2007, 26, 1657–1669. [Google Scholar] [CrossRef] [Green Version]
- Sumiyoshi, T.; Anil, A.E.; Jin, D.; Jayathilake, K.; Lee, M.; Meltzer, H.Y. Plasma glycine and serine levels in schizophrenia compared to normal controls and major depression: Relation to negative symptoms. Int. J. Neuropsychopharmacol. 1999, 7, 1–8. [Google Scholar] [CrossRef]
- Neeman, G.; Blanaru, M.; Bloch, B.; Kremer, I.; Ermilov, M.; Javitt, D.C.; Heresco-Levy, U. Relation of Plasma Glycine, Serine, and Homocysteine Levels to Schizophrenia Symptoms and Medication Type. Am. J. Psychiatry 2005, 162, 1738–1740. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, B.V.; Croft, R.J.; Mann, C.; Dang, O.; Leung, S.; Galloway, M.P.; Phan, K.L.; Nathan, P.J. High-dose glycine impairs the prepulse inhibition measure of sensorimotor gating in humans. J. Psychopharmacol. 2011, 25, 1632–1638. [Google Scholar] [CrossRef]
- Beneyto, M.; Meador-Woodruff, J.H. Lamina-specific abnormalities of AMPA receptor trafficking and signaling molecule transcripts in the prefrontal cortex in schizophrenia. Synapse 2006, 60, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Burnet, P.W.J.; Hutchinson, L.; Von Hesling, M.; Gilbert, E.J.; Brandon, N.J.; Rutter, A.R.; Hutson, P.H.; Harrison, P.J. Expression of D-serine and glycine transporters in the prefrontal cortex and cerebellum in schizophrenia. Schizophr. Res. 2008, 102, 283–294. [Google Scholar] [CrossRef]
- Mothet, J.P.; Rouaud, E.; Sinet, P.M.; Potier, B.; Jouvenceau, A.; Dutar, P.; Dutar, C.V.; Billard, J.M. A critical role for the glial-derived neuromodulator D-serine in the age-related deficits of cellular mechanisms of learning and memory. Aging Cell 2006, 5, 267–274. [Google Scholar] [CrossRef]
- Ohnuma, T.; Shibata, N.; Maeshima, H.; Baba, H.; Hatano, T.; Hanzawa, R.; Arai, H. Association analysis of glycine- and serine-related genes in a Japanese population of patients with schizophrenia. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 511–518. [Google Scholar] [CrossRef]
- Shinkai, T.; De Luca, V.; Hwang, R.; Muller, D.J.; Lanktree, M.; Zai, G.; Shaikh, S.; Wong, G.; Sicard, T.; Potapova, N.; et al. Association analyses of the DAOA/G30 and D-amino-acid oxidase genes in schizophrenia: Further evidence for a role in schizophrenia. Neuromol. Med. 2007, 9, 169–177. [Google Scholar] [CrossRef]
- Kotzadimitriou, D.; Nissen, W.; Paizs, M.; Newton, K.; Harrison, P.J.; Paulsen, O.; Lamsa, K. Neuregulin 1 type I overexpression is associated with reduced NMDA receptor-mediated synaptic signaling in hippocampal interneurons expressing PV or CCK. Eneuro 2018, 5, ENEURO.0418-17.2018. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.-J.; Lane, H.-Y.; Tsai, G. NMDA Pathology and Treatment of Schizophrenia. Curr. Pharm. Des. 2014, 20, 5118–5126. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Wei, P.; Jin, L.; Zheng, T.; Chen, W.Y.; Liu, X.Y.; Shi, X.; Hao, J.R.; Sun, N.; Gao, C. Overexpression of EphB2 in hippocampus rescues impaired NMDA receptors trafficking and cognitive dysfunction in Alzheimer model. Cell Death Dis. 2017, 8, e2717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, I.W.; Glausier, J.R. Different Paths to Core Pathology: The Equifinal Model of the Schizophrenia Syndrome. Schizophr. Bull. 2015, 42, 542–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Errico, F.; Mothet, J.-P.; Usiello, A. d-Aspartate: An endogenous NMDA receptor agonist enriched in the developing brain with potential involvement in schizophrenia. J. Pharm. Biomed. Anal. 2015, 116, 7–17. [Google Scholar] [CrossRef]
- Baker, D.; McFarland, K.; Lake, R.W.; Shen, H.; Tang, X.-C.; Toda, S.; Kalivas, P.W. Neuroadaptations in cystine-glutamate exchange underlie cocaine relapse. Nat. Neurosci. 2003, 6, 743–749. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kanai, Y.; Chairoungdua, A.; Cha, S.H.; Matsuo, H.; Kim, D.K.; Inatomi, J.; Sawa, H.; Ida, Y.; Endou, H. Human cystine/glutamate transporter: cDNA cloning and upregulation by oxidative stress in glioma cells. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1512, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, A.L.; Robinson, M.B. The role of glutamate transporters in neurodegenerative diseases and potential opportunities for intervention. Neurochem. Int. 2007, 51, 333–355. [Google Scholar] [CrossRef] [Green Version]
- Knackstedt, L.A.; LaRowe, S.; Mardikian, P.; Malcolm, R.; Upadhyaya, H.; Hedden, S.; Markou, A.; Kalivas, P.W. The Role of Cystine-Glutamate Exchange in Nicotine Dependence in Rats and Humans. Biol. Psychiatry 2009, 65, 841–845. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Biomolecular mechanisms between glutamate and system xc−.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hung, C.-C.; Lin, C.-H.; Lane, H.-Y. Cystine/Glutamate Antiporter in Schizophrenia: From Molecular Mechanism to Novel Biomarker and Treatment. Int. J. Mol. Sci. 2021, 22, 9718. https://doi.org/10.3390/ijms22189718
AMA Style
Hung C-C, Lin C-H, Lane H-Y. Cystine/Glutamate Antiporter in Schizophrenia: From Molecular Mechanism to Novel Biomarker and Treatment. International Journal of Molecular Sciences. 2021; 22(18):9718. https://doi.org/10.3390/ijms22189718
Chicago/Turabian StyleHung, Chung-Chieh, Chieh-Hsin Lin, and Hsien-Yuan Lane. 2021. "Cystine/Glutamate Antiporter in Schizophrenia: From Molecular Mechanism to Novel Biomarker and Treatment" International Journal of Molecular Sciences 22, no. 18: 9718. https://doi.org/10.3390/ijms22189718
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.