
Sample Preparation as a Critical Aspect of Blood Platelet Mitochondrial Respiration Measurements—The Impact of Platelet Activation on Mitochondrial Respiration

 ,
,
Abstract
:1. Introduction
2. Results
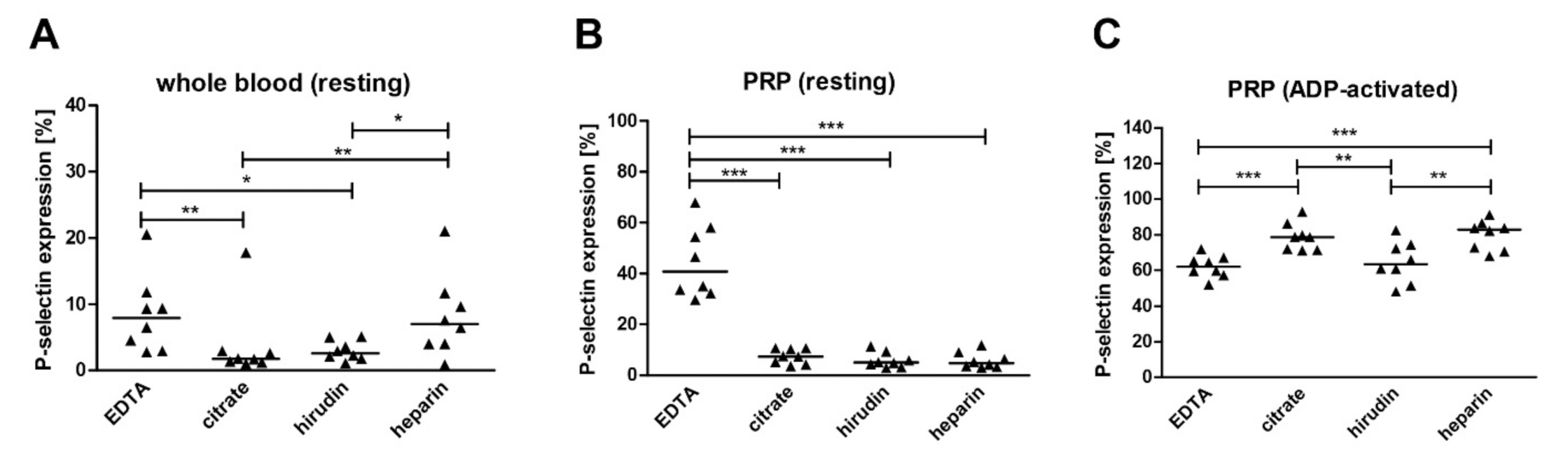
2.1. The Impact of Anticoagulants on Platelet Activation and Mitochondrial Respiration
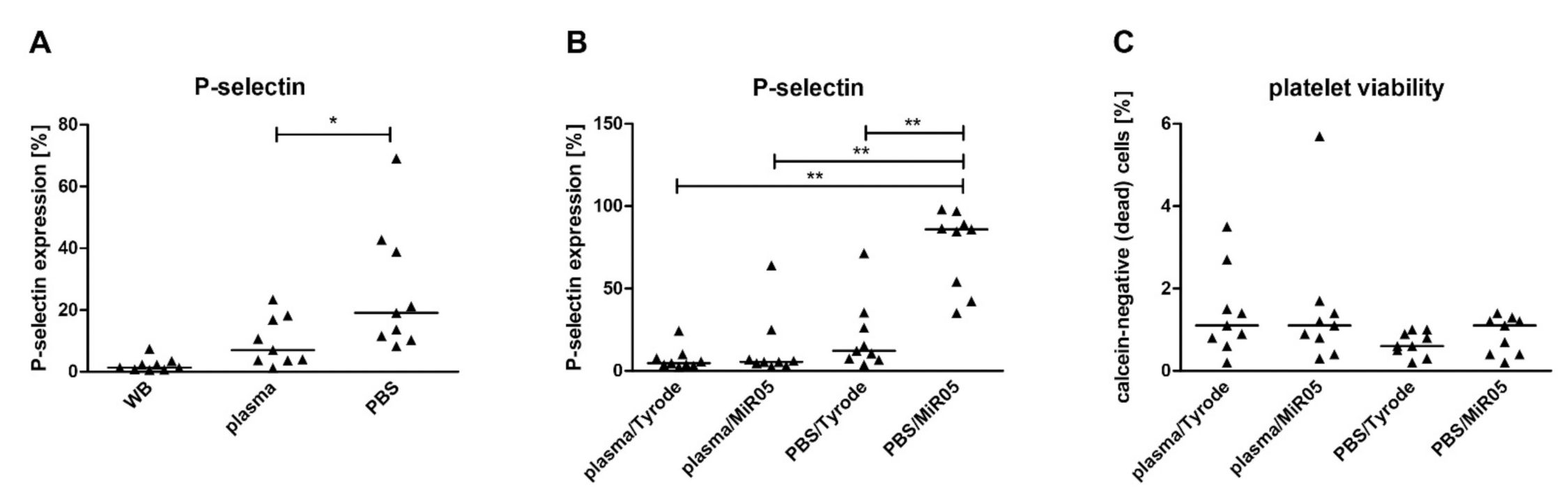
2.2. The Impact of Media and Respiration Buffers on Platelet Activation and Mitochondrial Respiration
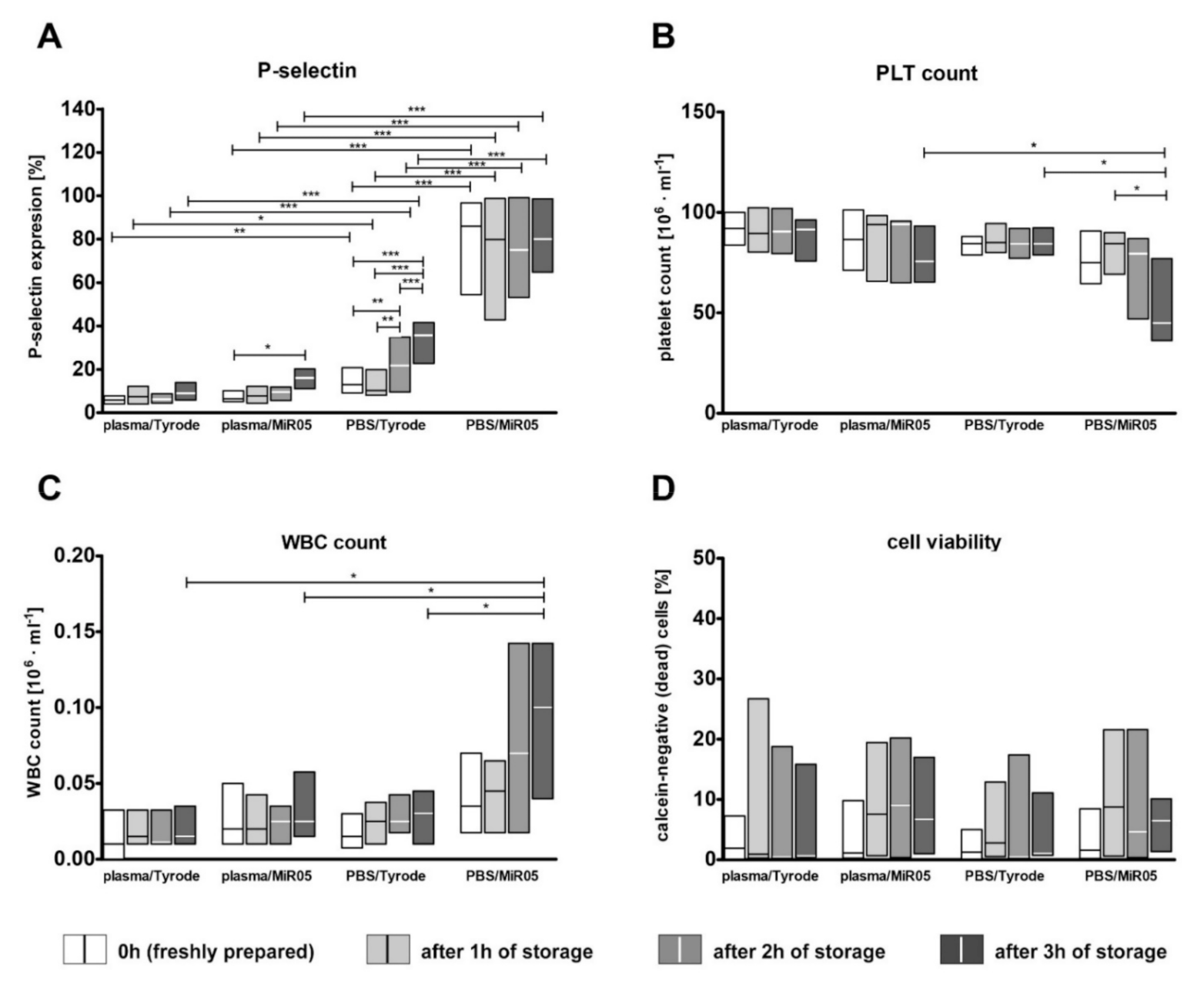
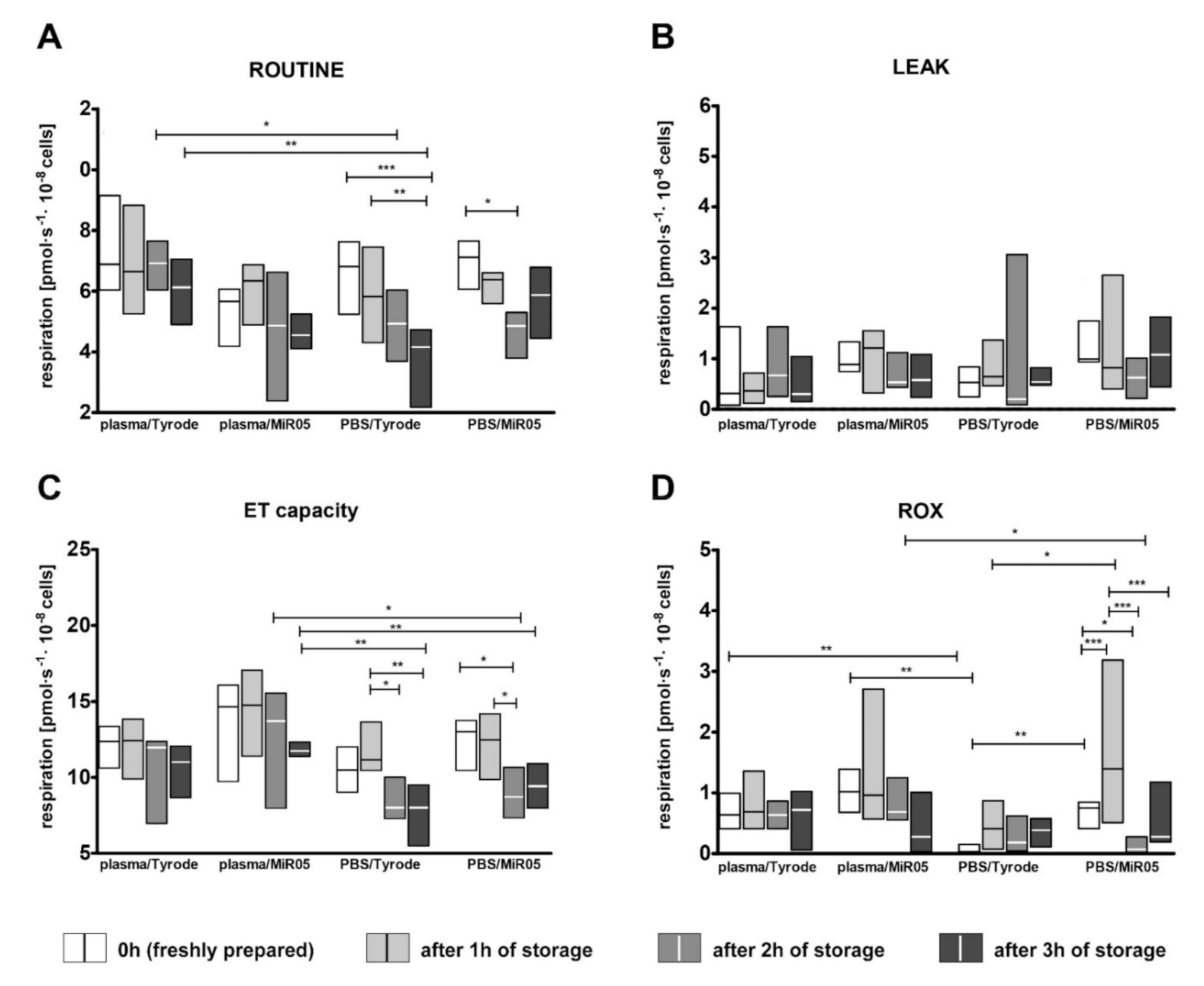
2.3. The Impact of Storage Time on Platelet Viability, Artefactual Activation and Mitochondrial Respiration
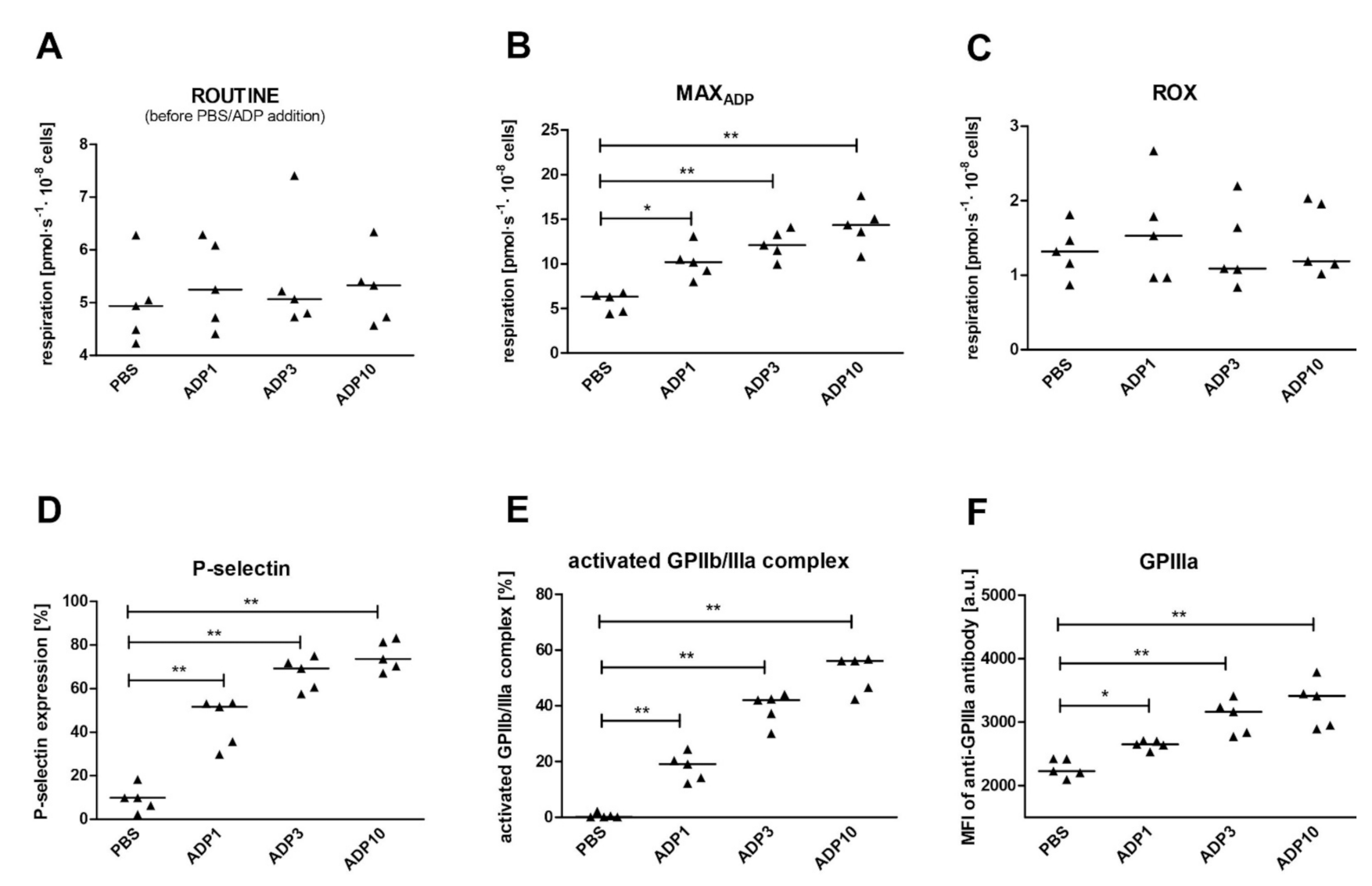
2.4. The Impact of Platelet Activation on Mitochondrial Respiration
2.5. Predictors of Platelet Functioning during Activation: Multivariate Analyses
3. Discussion
3.1. Effect of Anticoagulants on Platelet Activation and Mitochondrial Respiration
3.2. Effect of Media and Respiration Buffers on Platelet Activation and Mitochondrial Respiration
3.3. The Effect of the Short-Term Storage of Platelet Suspension on Platelet Activation and Mitochondrial Respiration
3.4. The Effect of Platelet Activation with ADP on Mitochondrial Respiration
3.5. The Impact of Platelet Activation in the Course of Sample Preparation for Oxygraphic Measurements on Platelet Mitochondrial Respiration
3.6. Study Limitations
4. Materials and Methods
4.1. Chemicals
4.2. Subjects
4.3. Blood Collection and Sample Preparation
4.4. Flow Cytometric Analysis of Platelet Activation and Viability
4.5. Measurement of Mitochondrial Respiratory Parameters
4.6. Study Design
- (a)
- Commonly used anticoagulants (EDTA, sodium citrate, hirudin, heparin): for this purpose, blood was collected using four different anticoagulants from each subject (n = 8). Experiments were conducted in intact platelets suspended in autologous plasma. As blood platelets in plasma appeared to be less sensitive to the uncoupler, for these measurements, FCCP was titrated in 2.25 µM aliquots.
- (b)
- The type of medium in which platelets are resuspended after isolation to prepare stock of cells, and the type of respiration buffer in which platelets are suspended for the measurements: for this purpose, blood was collected using sodium citrate, centrifuged to PRP and the platelet concentration was assessed. Next, PRP was evenly divided into two tubes and centrifuged to obtain platelet sediment, which was suspended in a small volume of autologous plasma or PBS to obtain a concentration close to 20 × 108 platelets/mL. The blood platelet concentration was validated again and the necessary number of platelets were transferred to the respiration buffer (Tyrode’s buffer with 0.3% of BSA or MiR05) to obtain 1 × 108 platelets/mL. The influence of two factors (medium and buffer) was examined at the same time in the following combinations: plasma/Tyrode, plasma/MiR05, PBS/Tyrode, PBS/MiR05; all of these were prepared from the blood of one subject (n = 9). Additionally, changes in the viability of the blood platelets were assessed.
- (c)
- The effect of storage time: we evaluated whether the results of platelet mitochondrial respiration will differ over time; measurements were taken every hour during three hours of storage from the sample preparation step. For this purpose, blood was collected using sodium citrate (n = 12). Isolated platelets were resuspended in autologous plasma or in PBS with 5 mM glucose and stored in the dark at 37 °C with gentle mixing by inversion every half an hour. At each time point, platelets were transferred to the respiration buffer (Tyrode’s buffer with 0.3% BSA or MIR05) to obtain 1 × 108 platelets/mL. As the blood platelets appeared to be more sensitive to FCCP at the one- to three-hour time points, for these measurements, FCCP was added in 0.125 µM aliquots instead of 0.25 µM aliquots. Additionally, changes in the viability of platelets during storage were verified.
- (d)
- The effect of platelet activation: for this purpose, blood was collected using sodium citrate (n = 10). Isolated platelets were suspended in plasma/MiR05, as in point c above. ROUTINE respiration was allowed to stabilize and then either ADP (1 µM, 3 µM, 10 µM, final concentrations) or PBS was added. After ADP addition to the oxygraphic chamber, an immediate increase in oxygen consumption was observed; this quickly reached the maximum rate (MAXADP) and began to decrease. When respiration stabilized, ROUTINE respiration in ADP-activated platelets were measured (ROUTINEADP) and the standard protocol was continued. Oxygen consumption in ROUTINE state, MAXADP, ROUTINEADP, LEAK state and for ET capacity was ROX-corrected. In addition, to better assess the differences in mitochondrial functioning between the tested variants, the following ratios were calculated: MAXADP/E, RADP/E, netROUTINEADP ((RADP − L)/E) and (RADP − R)/R × 100.
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fisar, Z.; Jirak, R.; Zverova, M.; Setnicka, V.; Habartova, L.; Hroudova, J.; Vanickova, Z.; Raboch, J. Plasma amyloid beta levels and platelet mitochondrial respiration in patients with Alzheimer’s disease. Clin. Biochem. 2019, 72, 71–80. [Google Scholar] [CrossRef]
- Cardenes, N.; Corey, C.; Geary, L.; Jain, S.; Zharikov, S.; Barge, S.; Novelli, E.M.; Shiva, S. Platelet bioenergetic screen in sickle cell patients reveals mitochondrial complex V inhibition, which contributes to platelet activation. Blood 2014, 123, 2864–2872. [Google Scholar] [CrossRef] [Green Version]
- Protti, A.; Fortunato, F.; Artoni, A.; Lecchi, A.; Motta, G.; Mistraletti, G.; Novembrino, C.; Comi, G.P.; Gattinoni, L. Platelet mitochondrial dysfunction in critically ill patients: Comparison between sepsis and cardiogenic shock. Crit. Care 2015, 19, 39. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Cardenes, N.; Corey, C.; Erzurum, S.C.; Shiva, S. Platelets from Asthmatic Individuals Show Less Reliance on Glycolysis. PLoS ONE 2015, 10, e0132007. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, T.; Mallick, S.K.; Chakraborty, A.; Maiti, A.; Chowdhury, S.; Bhattacharyya, M. Signature biomarkers in diabetes mellitus and associated cardiovascular diseases. Clin. Hemorheol. Microcirc. 2015, 59, 67–81. [Google Scholar] [CrossRef]
- Yamakawa, K.; Ogura, H.; Koh, T.; Ogawa, Y.; Matsumoto, N.; Kuwagata, Y.; Shimazu, T. Platelet mitochondrial membrane potential correlates with severity in patients with systemic inflammatory response syndrome. J. Trauma Acute Care Surg. 2013, 74, 411–417; discussion 418. [Google Scholar] [CrossRef]
- Sjovall, F.; Morota, S.; Hansson, M.J.; Friberg, H.; Gnaiger, E.; Elmer, E. Temporal increase of platelet mitochondrial respiration is negatively associated with clinical outcome in patients with sepsis. Crit. Care 2010, 14, R214. [Google Scholar] [CrossRef] [Green Version]
- Boudreau, L.H.; Duchez, A.C.; Cloutier, N.; Soulet, D.; Martin, N.; Bollinger, J.; Pare, A.; Rousseau, M.; Naika, G.S.; Levesque, T.; et al. Platelets release mitochondria serving as substrate for bactericidal group IIA-secreted phospholipase A2 to promote inflammation. Blood 2014, 124, 2173–2183. [Google Scholar] [CrossRef] [Green Version]
- Marcoux, G.; Duchez, A.C.; Rousseau, M.; Levesque, T.; Boudreau, L.H.; Thibault, L.; Boilard, E. Microparticle and mitochondrial release during extended storage of different types of platelet concentrates. Platelets 2017, 28, 272–280. [Google Scholar] [CrossRef]
- Ahnadi, C.E.; Sabrinah Chapman, E.; Lepine, M.; Okrongly, D.; Pujol-Moix, N.; Hernandez, A.; Boughrassa, F.; Grant, A.M. Assessment of platelet activation in several different anticoagulants by the Advia 120 Hematology System, fluorescence flow cytometry, and electron microscopy. Thromb. Haemost. 2003, 90, 940–948. [Google Scholar] [CrossRef]
- Soderstrom, A.C.; Nybo, M.; Nielsen, C.; Vinholt, P.J. The effect of centrifugation speed and time on pre-analytical platelet activation. Clin. Chem. Lab. Med. 2016, 54, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Maurer-Spurej, E.; Pfeiler, G.; Maurer, N.; Lindner, H.; Glatter, O.; Devine, D.V. Room temperature activates human blood platelets. Lab. Investig. A J. Tech. Methods Pathol. 2001, 81, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akkerman, J.W.; Holmsen, H. Interrelationships among platelet responses: Studies on the burst in proton liberation, lactate production, and oxygen uptake during platelet aggregation and Ca2+ secretion. Blood 1981, 57, 956–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barile, C.J.; Herrmann, P.C.; Tyvoll, D.A.; Collman, J.P.; Decreau, R.A.; Bull, B.S. Inhibiting platelet-stimulated blood coagulation by inhibition of mitochondrial respiration. Proc. Natl. Acad. Sci. USA 2012, 109, 2539–2543. [Google Scholar] [CrossRef] [Green Version]
- Tomasiak, M.; Stelmach, H.; Rusak, T.; Wysocka, J. Nitric oxide and platelet energy metabolism. Acta Biochim. Pol. 2004, 51, 789–803. [Google Scholar] [CrossRef] [Green Version]
- Petrus, A.T.; Lighezan, D.L.; Danila, M.D.; Duicu, O.M.; Sturza, A.; Muntean, D.M.; Ionita, I. Assessment of platelet respiration as emerging biomarker of disease. Physiol. Res. 2019, 68, 347–363. [Google Scholar] [CrossRef]
- Golebiewska, E.M.; Poole, A.W. Platelet secretion: From haemostasis to wound healing and beyond. Blood Rev. 2015, 29, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Siewiera, K.; Kassassir, H.; Talar, M.; Wieteska, L.; Watala, C. Higher mitochondrial potential and elevated mitochondrial respiration are associated with excessive activation of blood platelets in diabetic rats. Life Sci. 2016, 148, 293–304. [Google Scholar] [CrossRef]
- Ehinger, J.K.; Morota, S.; Hansson, M.J.; Paul, G.; Elmer, E. Mitochondrial Respiratory Function in Peripheral Blood Cells from Huntington’s Disease Patients. Mov. Disord. Clin. Pract. 2016, 3, 472–482. [Google Scholar] [CrossRef] [Green Version]
- Puskarich, M.A.; Kline, J.A.; Watts, J.A.; Shirey, K.; Hosler, J.; Jones, A.E. Early alterations in platelet mitochondrial function are associated with survival and organ failure in patients with septic shock. J. Crit. Care 2016, 31, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Golanski, J.; Pietrucha, T.; Baj, Z.; Greger, J.; Watala, C. Molecular insights into the anticoagulant-induced spontaneous activation of platelets in whole blood-various anticoagulants are not equal. Thromb. Res. 1996, 83, 199–216. [Google Scholar] [CrossRef]
- Ritchie, J.L.; Alexander, H.D.; Rea, I.M. Flow cytometry analysis of platelet P-selectin expression in whole blood--methodological considerations. Clin. Lab. Haematol. 2000, 22, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Gatterer, H.; Menz, V.; Salazar-Martinez, E.; Sumbalova, Z.; Garcia-Souza, L.F.; Velika, B.; Gnaiger, E.; Burtscher, M. Exercise Performance, Muscle Oxygen Extraction and Blood Cell Mitochondrial Respiration after Repeated-Sprint and Sprint Interval Training in Hypoxia: A Pilot Study. J. Sports Sci. Med. 2018, 17, 339–347. [Google Scholar]
- Karlsson, M.; Ehinger, J.K.; Piel, S.; Sjovall, F.; Henriksnas, J.; Hoglund, U.; Hansson, M.J.; Elmer, E. Changes in energy metabolism due to acute rotenone-induced mitochondrial complex I dysfunction—An in vivo large animal model. Mitochondrion 2016, 31, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Vevera, J.; Fisar, Z.; Nekovarova, T.; Vrablik, M.; Zlatohlavek, L.; Hroudova, J.; Singh, N.; Raboch, J.; Vales, K. Statin-induced changes in mitochondrial respiration in blood platelets in rats and human with dyslipidemia. Physiol. Res. 2016, 65, 777–788. [Google Scholar] [CrossRef]
- Malinow, A.M.; Schuh, R.A.; Alyamani, O.; Kim, J.; Bharadwaj, S.; Crimmins, S.D.; Galey, J.L.; Fiskum, G.; Polster, B.M. Platelets in preeclamptic pregnancies fail to exhibit the decrease in mitochondrial oxygen consumption rate seen in normal pregnancies. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, D.H.; Khatri, U.G.; Mudan, A.; Love, J.S.; Owiredu, S.; Eckmann, D.M. Translational Application of Measuring Mitochondrial Functions in Blood Cells Obtained from Patients with Acute Poisoning. J. Med. Toxicol. Off. J. Am. Coll. Med. Toxicol. 2018, 14, 144–151. [Google Scholar] [CrossRef]
- Kramer, P.A.; Chacko, B.K.; Ravi, S.; Johnson, M.S.; Mitchell, T.; Darley-Usmar, V.M. Bioenergetics and the oxidative burst: Protocols for the isolation and evaluation of human leukocytes and platelets. J. Vis. Exp. 2014, 85, e51301. [Google Scholar] [CrossRef] [Green Version]
- Vagdatli, E.; Gounari, E.; Lazaridou, E.; Katsibourlia, E.; Tsikopoulou, F.; Labrianou, I. Platelet distribution width: A simple, practical and specific marker of activation of coagulation. Hippokratia 2010, 14, 28–32. [Google Scholar]
- Hechler, B.; Dupuis, A.; Mangin, P.H.; Gachet, C. Platelet preparation for function testing in the laboratory and clinic: Historical and practical aspects. Res. Pract. Thromb. Haemost. 2019, 3, 615–625. [Google Scholar] [CrossRef] [Green Version]
- Sjovall, F.; Ehinger, J.K.; Marelsson, S.E.; Morota, S.; Frostner, E.A.; Uchino, H.; Lundgren, J.; Arnbjornsson, E.; Hansson, M.J.; Fellman, V.; et al. Mitochondrial respiration in human viable platelets--methodology and influence of gender, age and storage. Mitochondrion 2013, 13, 7–14. [Google Scholar] [CrossRef]
- Wollenman, L.C.; Vander Ploeg, M.R.; Miller, M.L.; Zhang, Y.; Bazil, J.N. The effect of respiration buffer composition on mitochondrial metabolism and function. PLoS ONE 2017, 12, e0187523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, T.; Tanaka, S.; Hori, Y.; Hirayama, F.; Sato, E.F.; Inoue, M. Role of mitochondria in the maintenance of platelet function during in vitro storage. Transfus. Med. 2011, 21, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Perales Villarroel, J.P.; Figueredo, R.; Guan, Y.; Tomaiuolo, M.; Karamercan, M.A.; Welsh, J.; Selak, M.A.; Becker, L.B.; Sims, C. Increased platelet storage time is associated with mitochondrial dysfunction and impaired platelet function. J. Surg. Res. 2013, 184, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sowton, A.P.; Millington-Burgess, S.L.; Murray, A.J.; Harper, M.T. Rapid kinetics of changes in oxygen consumption rate in thrombin-stimulated platelets measured by high-resolution respirometry. Biochem. Biophys. Res. Commun. 2018, 503, 2721–2727. [Google Scholar] [CrossRef]
- Przygodzki, T.; Luzak, B.; Kassassir, H.; Mnich, E.; Boncler, M.; Siewiera, K.; Kosmalski, M.; Szymanski, J.; Watala, C. Diabetes and Hyperglycemia Affect Platelet GPIIIa Expression. Effects on Adhesion Potential of Blood Platelets from Diabetic Patients under In Vitro Flow Conditions. Int. J. Mol. Sci. 2020, 21, 3222. [Google Scholar] [CrossRef]
- Rywaniak, J.; Luzak, B.; Podsedek, A.; Dudzinska, D.; Rozalski, M.; Watala, C. Comparison of cytotoxic and anti-platelet activities of polyphenolic extracts from Arnica montana flowers and Juglans regia husks. Platelets 2015, 26, 168–176. [Google Scholar] [CrossRef]
- Gnaiger, E. Polarographic Oxygen Sensors, the Oxygraph, and High-Resolution Respirometry to Assess Mitochondrial Function. Drug-Induc. Mitochondrial Dysfunct. 2008, 327, 325–352. [Google Scholar] [CrossRef]
- Gnaiger, E.; Renner-Sattler, K. High-resolution respirometry and phosphorylation control protocol with intact cells: ROUTINE, LEAK, ETS, ROX. Mitochondrial Physiol. Netw. 2014, 8, 1–8. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anticoagulant | ROUTINE R | LEAK L | ET E | ROX | R/E | L/E | netROUTINE (R − L)/E |
|---|---|---|---|---|---|---|---|
| (pmol × s−1 × 10−8 cells) | (a.u.) | ||||||
| citrate | 7.50; 6.62–7.86 | 0.34; 0.14–0.46 | 13.43; 12.33–18.30 | 3.35 2.61–3.92 | 0.51; 0.51–0.56 | 0.03; 0.00–0.03 | 0.50; 0.48–0.56 |
| hirudin | 6.14; 5.72–7.56 | 0.32; 0.23–0.46 | 13.97; 12.15–16.73 | 3.57; 3.36–5.06 | 0.48; 0.43–0.53 | 0.02; 0.01–0.04 | 0.43; 0.4–0.48 |
| heparin | 6.65; 5.98–7.48 | 0.36; 0.03–0.81 | 12.94; 12.09–16.03 | 3.98; 3.18–4.42 | 0.50; 0.42–0.54 | 0.03; 0.00–0.07 | 0.46; 0.42–0.50 |
| EDTA | 9.96; 8.4–16.60 | 2.20; 1.04–7.58 | 14.45; 13.25–17.67 | 2.95; 2.72–4.24 | 0.77; 0.59–0.85 | 0.19; 0.07–0.41 | 0.49; 0.47–0.65 |
| Significance by inference tests | * p= 0.003 EDTA vs. citrate, hirudin, heparin #P1α < 0.03 | * p = 0.013 EDTA vs. citrate # p1α = 0.023 EDTA vs. hirudin # p1α = 0.049 EDTA vs. heparin # p1α = 0.034 | ns. | ns. | * p = 0.014 EDTA vs. citrate # p1α = 0.048 EDTA vs. hirudin # p1α = 0.026 EDTA vs. heparin # p1α = 0.028 | * p = 0.020 EDTA vs. citrate # p1α = 0.026 EDTA vs. hirudin # p1α = 0.04 EDTA vs. heparin # p1α = 0.075 | ns. |
| Anticoagulant | ROUTINE R | LEAK L | ET E | ROX | R/E | L/E | netROUTINE (R − L)/E |
|---|---|---|---|---|---|---|---|
| Bootstrap-boosted Spearman RS in groups, P1-sided | |||||||
| citrate | −0.183, ns. | 0.235, ns. | 0.691, p < 0.0001 | −0.838, p < 0.0001 | −0.828, p < 0.0001 | 0.038, ns. | −0.826, p < 0.0001 |
| hirudin | 0.002, ns. | 0.206, ns. | 0.315, p = 0.040 | 0.316, p = 0.039 | −0.144, ns. | 0.191, ns. | −0.114 ns. |
| heparin | −0.194, ns. | −0.565, p = 0.0004 | 0.062, ns. | 0.121, ns. | −0.051, ns. | −0.511, p = 0.001 | 0.259, ns. |
| EDTA | −0.339, p = 0.029 | −0.407, p = 0.010 | −0.046, ns. | −0.496, p = 0.002 | −0.087, ns. | −0.332, p = 0.032 | 0.195, ns. |
| Overall@ and partial & (in parentheses) Spearman RS, P1-sided | @ 0.428, p = 0.007 & (0.314, p = 0.040) | @ 0.321, p = 0.037 & (0.108, ns.) | @ 0.284, ns. & (0.273, ns.) | @ −0.207, ns. & (−0.237, ns.) | @ 0.264. ns. & (0.089, ns.) | @ 0.273. ns. & (0.048, ns.) | @ 0.020. ns. & (0.034, ns.) |
| Condition Medium/Buffer | ROUTINE R | LEAK L | ET E | ROX |
|---|---|---|---|---|
| (pmol × s−1 × 10−8 cells) | ||||
| plasma/MiR05 | 5.05; 4.32–6.06 | 0.61; 0.28–1.16 | 13.17; 11.02–13.93 | 1.26; 0.50–3.43 |
| plasma/Tyrode | 6.07; 5.10–6.76 | 0.19; 0.09–0.73 | 9.95; 9.29–11.41 | 0.66; 0.25–1.04 |
| PBS/MiR05 | 6.38; 5.30–7.12 | 0.77; 0.37–1.14 | 13.27; 10.64–15.14 | 1.27; 1.19–2.43 |
| PBS/Tyrode | 6.05; 3.92–7.79 | 0.29; 0.09–0.38 | 11.33; 9.15–13.96 | 0.68; 0.38–1.21 |
| Statistical significance of the effect of medium: plasma vs. PBS | ns. | ns. | ns. | ns. |
| Statistical significance of the effect of buffer: MiR05 vs. Tyrode | ns. | p < 0.05 | p < 0.05 | p = 0.0706 |
| Statistical significance of the differences between the four tested combinations | p < 0.05 plasma/MiR05 vs. plasma/Tyrode | p < 0.05 PBS/MiR05 vs. plasma/Tyrode, plasma/MiR05 vs. plasma/Tyrode | p < 0.05 plasma/MiR05 vs. plasma/Tyrode | |
| Statistical significance according to bootstrap-boosted test | p < 0.05 plasma/MiR05 vs. plasma/Tyrode | p < 0.01 plasma/MiR05 vs. plasma/Tyrode, p < 0.001 PBS/MiR05 vs. plasma/Tyrode, | p< 0.05 plasma/MiR05 vs. plasma/Tyrode | |
| ROUTINE R | MAXADP | ROUTINEADP RADP | LEAK L | ET E | ROX | R/E | RADP/E | netROUTINEADP (RADP − L)/E | MAXADP/E | (RADP − R)/R*100 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| (pmol × s−1 × 10−8 cells) | (a.u.) | (%) | |||||||||
| resting (PBS) | 6.19; 4.86–6.87 | 6.80; 5.61–7.92 | 5.93; 5.26–7.13 | 0.69; 0.28–0.98 | 12.40; 11.01–14.00 | 1.11; 0.71–1.30 | 0.49; 0.40–0.53 | 0.47; 0.44–0.53 | 0.40; 0.36–0.50 | 0.50; 0.48–0.63 | 5.77; 8.15–13.20 |
| ADP 1 μM | 5.49; 4.86–6.74 | 12.18; 11.75–12.87 | 7.72; 6.70–8.18 | 0.37; 0.18–0.54 | 11.45; 10.24–12.77 | 1.20; 0.82–1.57 | 0.51; 0.43–0.56 | 0.64; 0.59–0.69 | 0.62; 0.54–0.67 | 1.03; 0.94–1.16 | 30.01; 17.91–39.13 |
| ADP 3 μM | 5.43; 5.04–5.94 | 12.72; 11.50–13.74 | 8.18; 6.77–8.38 | 0.27; 0.09–0.55 | 11.51; 10.66–12.80 | 1.33; 0.94–1.67 | 0.47; 0.43–0.54 | 0.66; 0.63–0.71 | 0.63; 0.60–0.66 | 1.07; 0.99–1.17 | 37.73; 30.69–53.52 |
| ADP 10 μM | 5.66; 5.09–6.26 | 13.29; 12.04–16.35 | 7.72; 7.17–8.40 | 0.31; 0.21–0.78 | 11.98; 10.42–13.01 | 1.30; 0.97–1.47 | 0.47; 0.45–0.55 | 0.68; 0.61–0.70 | 0.60; 0.57–0.66 | 1.10; 1.03–1.31 | 35.93; 30.57–41.14 |
| Statistical significance | # ns. | # p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | @ p < 0.05 PBS vs. ADP3, PBS vs. ADP10 | # ns. | # p < 0.05 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | #ns. | # ns. | # p< 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | # p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | # p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | # p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 |
| Statistical significance by bootstrap-boosted | p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | p < 0.01 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | p < 0.01 PBS vs. ADP3, PBS vs. ADP10 | p = 0.001 PBS vs. ADP1; p < 0.001 PBS vs. ADP3, PBS vs. ADP10 | p = 0.002 PBS vs. ADP1; p < 0.001 PBS vs. ADP3, PBS vs. ADP10 | p < 0.001 PBS vs. ADP1, PBS vs. ADP3, PBS vs. ADP10 | p = 0.002 PBS vs. ADP1; p < 0.001 PBS vs. ADP3, PBS vs. ADP10 | ||||
| Variable | P-Selectin (%) | Activated GPIIb/IIIa Complex (%) | GPIIIa (a.u.) |
|---|---|---|---|
| MAXADP | 0.580, p < 0.0001 (−0.050, ns.) | 0.624, p < 0.0001 (0.142, ns.) | 0.587, p < 0.0001 (0.168, ns.) |
| ROUTINEADP | 0.402, p = 0.008 (0.172, ns.) | 0.428, p = 0.005 (0.239, ns.) | 0.372, p = 0.013 (0.170, ns) |
| ET capacity | −0.011, ns. (0.204, ns.) | 0.024, ns. (0.225, ns.) | 0.149, ns. (0.348, p = 0.019) |
| RADP/E | 0.358, p = 0.016 (−0.142, ns.) | 0.391, p = 0.009 (−0.017, ns.) | 0.297, p = 0.039 (−0.094, ns.) |
| netROUTINEADP | 0.320, p = 0.028 (−0.133, ns.) | 0.403, p = 0.007 (0.072, ns.) | 0.236, ns. (−0.137, ns.) |
| (RADP − R)/R*100 | 0.641, p < 0.0001 (0.294, p = 0.041) | 0.734, p < 0.0001 (0.513, p = 0.001) | 0.544, p < 0.0001 (0.208, ns.) |
| MAXADP/E | 0.503, p = 0.001 (−0.264, ns.) | 0.579, p < 0.0001 (0.026, ns.) | 0.522, p = 0.001 (0.040, ns.) |
| Resting/Activated Cells | Platelet Reactivity/Activation (Set 1) | Extracted Variance [%] | Total REDUNDANCE [%] | Explanatory Variables (Set 2) | Extracted Variance [%] | Total Redundance [%] | Canonical Correlation [R] | Canonical Determination [R2] | p | Wilks’ Lambda | Best Contributors |
|---|---|---|---|---|---|---|---|---|---|---|---|
| resting platelets | cytometric | 100.0% | 52.8% | mitochondrial | 58.1% | 38.2% | 0.960 | 0.921 | 0.000 | 0.008 | P-selectin, GPIIb/IIIaADP, (RADP −R)/R × 100, MAXADP, ROUTINEADP |
| activated platelets | 100.0% | 35.3% | 34.5% | 11.6% | 0.698 | 0.487 | 0.039 | 0.320 | |||
| all platelets | 100.0% | 51.8% | 66.8% | 31.1% | 0.826 | 0.681 | 0.003 | 0.228 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siewiera, K.; Labieniec-Watala, M.; Wolska, N.; Kassassir, H.; Watala, C. Sample Preparation as a Critical Aspect of Blood Platelet Mitochondrial Respiration Measurements—The Impact of Platelet Activation on Mitochondrial Respiration. Int. J. Mol. Sci. 2021, 22, 9332. https://doi.org/10.3390/ijms22179332
Siewiera K, Labieniec-Watala M, Wolska N, Kassassir H, Watala C. Sample Preparation as a Critical Aspect of Blood Platelet Mitochondrial Respiration Measurements—The Impact of Platelet Activation on Mitochondrial Respiration. International Journal of Molecular Sciences. 2021; 22(17):9332. https://doi.org/10.3390/ijms22179332
Chicago/Turabian StyleSiewiera, Karolina, Magdalena Labieniec-Watala, Nina Wolska, Hassan Kassassir, and Cezary Watala. 2021. "Sample Preparation as a Critical Aspect of Blood Platelet Mitochondrial Respiration Measurements—The Impact of Platelet Activation on Mitochondrial Respiration" International Journal of Molecular Sciences 22, no. 17: 9332. https://doi.org/10.3390/ijms22179332