
Research Advances in Heterotrimeric G-Protein α Subunits and Uncanonical G-Protein Coupled Receptors in Plants

Abstract
:1. Introduction
2. Self-Activation and Recycling of Gα in Plants
3. Research Progress on the Gα in Plants
4. GPCR, a G-Protein-Coupled Receptor
4.1. Structure and Classification of Canonical GPCRs
4.2. Prediction of Putative GPCR in Plants
5. Research Progress and New Definition of Uncanonical Plant GPCRs
5.1. Putative Plant GPCRs
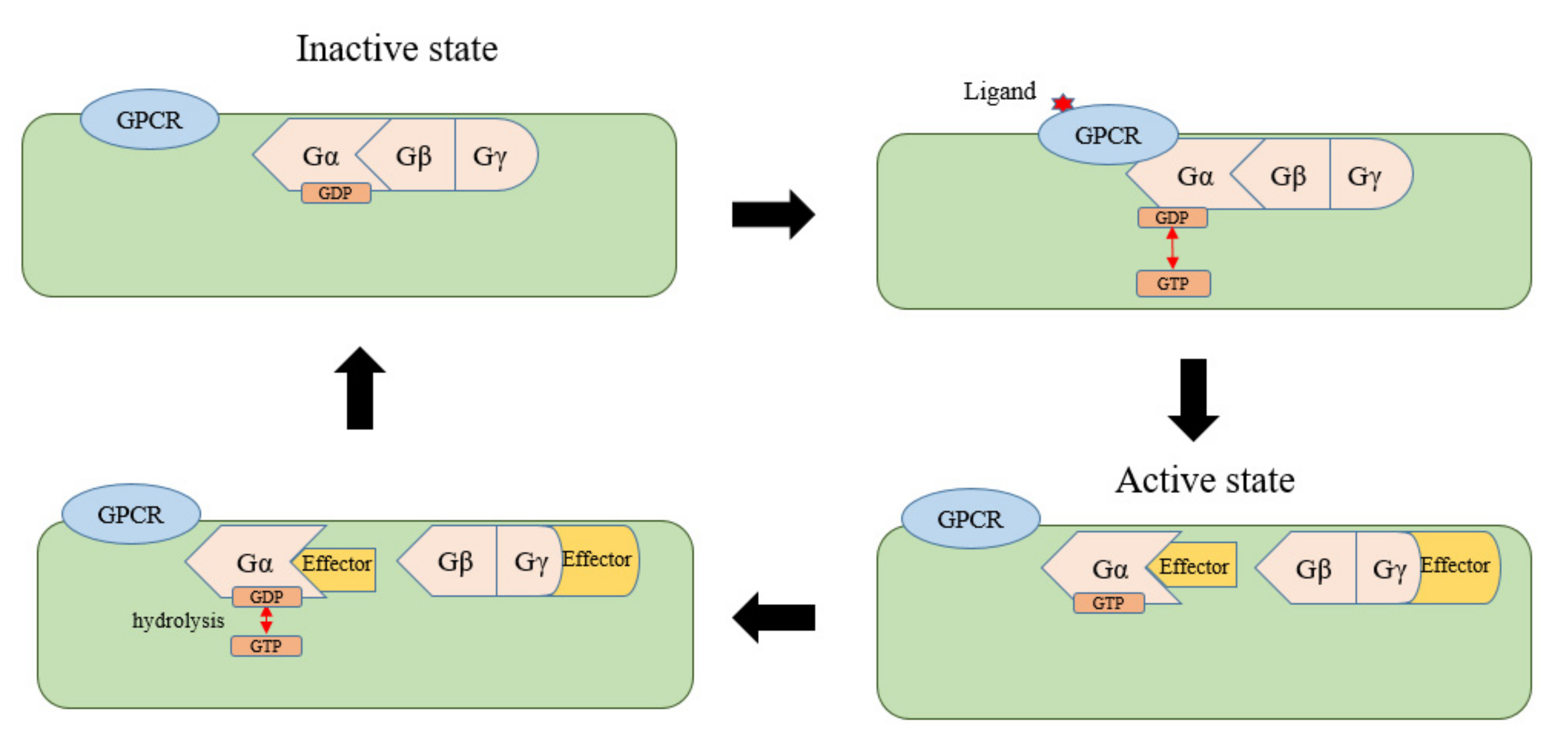
5.2. New Definition of GPCRs and a Proposed Model of the G-Protein Cycle in Plants
6. Summary and Prospect
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCudden, C.R.; Hains, M.D.; Kimple, R.J.; Siderovski, D.P.; Willard, F.S. G-protein signaling: Back to the future. Cell Mol. Life Sci. 2005, 62, 551–577. [Google Scholar] [CrossRef] [Green Version]
- Stateczny, D.; Oppenheimer, J.; Bommert, P. G protein signaling in plants: Minus times minus equals plus. Curr. Opin. Plant Biol. 2016, 34, 127–135. [Google Scholar] [CrossRef]
- Chen, J.G.; Willard, F.S.; Huang, J.; Liang, J.; Chasse, S.A.; Jones, A.M.; Siderovski, D.P. A seven-transmembrane RGS protein that modulates plant cell proliferation. Science 2003, 301, 1728–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.C.; Duffy, J.W.; Machius, M.; Temple, B.R.; Dohlman, H.G.; Jones, A.M. The crystal structure of a self-activating G protein alpha subunit reveals its distinct mechanism of signal initiation. Sci. Signal 2011, 4, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sprang, S.R. G protein mechanisms: Insights from structural analysis. Annu. Rev. Biochem. 1997, 66, 639–678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldham, W.M.; Hamm, H.E. Heterotrimeric G protein activation by G-protein-coupled receptors. Nat. Rev. Mol. Cell Biol. 2008, 9, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Neubig, R.R.; Siderovski, D.P. Regulators of G-protein signalling as new central nervous system drug targets. Nat. Rev. Drug Discov. 2002, 1, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Siderovski, D.P.; Willard, F.S. The GAPs, GEFs, and GDIs of heterotrimeric G-protein alpha subunits. Int. J. Biol. Sci. 2005, 1, 51–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackenberg, D.; Perroud, P.F.; Quatrano, R.; Pandey, S. Sporophyte Formation and Life Cycle Completion in Moss Requires Heterotrimeric G-Proteins. Plant Physiol. 2016, 172, 1154–1166. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.C.; Jones, A.M.; Temple, B.R.; Dohlman, H.G. Differences in intradomain and interdomain motion confer distinct activation properties to structurally similar Galpha proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 7275–7279. [Google Scholar] [CrossRef] [Green Version]
- Urano, D.; Chen, J.G.; Botella, J.R.; Jones, A.M. Heterotrimeric G protein signalling in the plant kingdom. Open Biol. 2013, 3, 120186. [Google Scholar] [CrossRef] [Green Version]
- Johnston, C.A.; Taylor, J.P.; Gao, Y.; Kimple, A.J.; Grigston, J.C.; Chen, J.G.; Siderovski, D.P.; Jones, A.M.; Willard, F.S. GTPase acceleration as the rate-limiting step in Arabidopsis G protein-coupled sugar signaling. Proc. Natl. Acad. Sci. USA 2007, 104, 17317–17322. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, S.R.; Westfall, C.S.; Pandey, S. The RGS proteins add to the diversity of soybean heterotrimeric G-protein signaling. Plant Signal. Behav. 2012, 7, 1114–1117. [Google Scholar] [CrossRef] [Green Version]
- Bhatnagar, N.; Pandey, S. Heterotrimeric G-Protein Interactions Are Conserved Despite Regulatory Element Loss in Some Plants. Plant Physiol. 2020, 184, 1941–1954. [Google Scholar] [CrossRef]
- Roy Choudhury, S.; Pandey, S. Recently duplicated plant heterotrimeric Galpha proteins with subtle biochemical differences influence specific outcomes of signal-response coupling. J. Biol. Chem. 2017, 292, 16188–16198. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S. Heterotrimeric G-protein regulatory circuits in plants: Conserved and novel mechanisms. Plant Signal. Behav. 2017, 12, e1325983. [Google Scholar] [CrossRef] [Green Version]
- Roy Choudhury, S.; Wang, Y.; Pandey, S. Soya bean Galpha proteins with distinct biochemical properties exhibit differential ability to complement Saccharomyces cerevisiae gpa1 mutant. Biochem. J. 2014, 461, 75–85. [Google Scholar] [CrossRef]
- Maruta, N.; Trusov, Y.; Chakravorty, D.; Urano, D.; Assmann, S.M.; Botella, J.R. Nucleotide exchange-dependent and nucleotide exchange-independent functions of plant heterotrimeric GTP-binding proteins. Sci. Signal. 2019, 12. [Google Scholar] [CrossRef]
- Jose, J.; Roy Choudhury, S. Heterotrimeric G-proteins mediated hormonal responses in plants. Cell Signal. 2020, 76, 109799. [Google Scholar] [CrossRef]
- Lapik, Y.R.; Kaufman, L.S. The Arabidopsis cupin domain protein AtPirin1 interacts with the G protein alpha-subunit GPA1 and regulates seed germination and early seedling development. Plant Cell 2003, 15, 1578–1590. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.G.; Pandey, S.; Huang, J.; Alonso, J.M.; Ecker, J.R.; Assmann, S.M.; Jones, A.M. GCR1 can act independently of heterotrimeric G-protein in response to brassinosteroids and gibberellins in Arabidopsis seed germination. Plant Physiol. 2004, 135, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Kato, C.; Mizutani, T.; Tamaki, H.; Kumagai, H.; Kamiya, T.; Hirobe, A.; Fujisawa, Y.; Kato, H.; Iwasaki, Y. Characterization of heterotrimeric G protein complexes in rice plasma membrane. Plant J. 2004, 38, 320–331. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Fujisawa, Y.; Kobayashi, M.; Ashikari, M.; Iwasaki, Y.; Kitano, H.; Matsuoka, M. Rice dwarf mutant d1, which is defective in the alpha subunit of the heterotrimeric G protein, affects gibberellin signal transduction. Proc. Natl. Acad. Sci. USA 2000, 97, 11638–11643. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Qi, C.; Ren, H.; Huang, A.; Hei, S.; She, X. Ethylene mediates brassinosteroid-induced stomatal closure via Galpha protein-activated hydrogen peroxide and nitric oxide production in Arabidopsis. Plant J. 2015, 82, 280–301. [Google Scholar] [CrossRef]
- Wang, X.Q.; Ullah, H.; Jones, A.M.; Assmann, S.M. G protein regulation of ion channels and abscisic acid signaling in Arabidopsis guard cells. Science 2001, 292, 2070–2072. [Google Scholar] [CrossRef] [Green Version]
- Coursol, S.; Fan, L.M.; Le Stunff, H.; Spiegel, S.; Gilroy, S.; Assmann, S.M. Sphingolipid signalling in Arabidopsis guard cells involves heterotrimeric G proteins. Nature 2003, 423, 651–654. [Google Scholar] [CrossRef]
- Ullah, H.; Chen, J.G.; Wang, S.; Jones, A.M. Role of a heterotrimeric G protein in regulation of Arabidopsis seed germination. Plant Physiol. 2002, 129, 897–907. [Google Scholar] [CrossRef] [Green Version]
- Yan, S.; Luo, S.; Dong, S.; Zhang, T.; Sun, J.; Wang, N.; Yao, H.; Shen, Y. Heterotrimeric G-proteins involved in the MeJA regulated ion flux and stomatal closure in Arabidopsis thaliana. Funct. Plant Biol. 2015, 42, 126–135. [Google Scholar] [CrossRef]
- Steffens, B.; Sauter, M. Heterotrimeric G protein signaling is required for epidermal cell death in rice. Plant Physiol. 2009, 151, 732–740. [Google Scholar] [CrossRef] [Green Version]
- Ge, X.M.; Cai, H.L.; Lei, X.; Zhou, X.; Yue, M.; He, J.M. Heterotrimeric G protein mediates ethylene-induced stomatal closure via hydrogen peroxide synthesis in Arabidopsis. Plant J. 2015, 82, 138–150. [Google Scholar] [CrossRef]
- Ullah, H.; Chen, J.G.; Young, J.C.; Im, K.H.; Sussman, M.R.; Jones, A.M. Modulation of cell proliferation by heterotrimeric G protein in Arabidopsis. Science 2001, 292, 2066–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullah, H.; Chen, J.G.; Temple, B.; Boyes, D.C.; Alonso, J.M.; Davis, K.R.; Ecker, J.R.; Jones, A.M. The beta-subunit of the Arabidopsis G protein negatively regulates auxin-induced cell division and affects multiple developmental processes. Plant Cell 2003, 15, 393–409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, S.; Bahn, S.C.; Qu, G.; Qin, H.; Hong, Y.; Xu, Q.; Zhou, Y.; Hong, Y.; Wang, X. Increased expression of phospholipase Dalpha1 in guard cells decreases water loss with improved seed production under drought in Brassica napus. Plant Biotechnol. J. 2013, 11, 380–389. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.S.; Pandey, S. The role of PLDalpha1 in providing specificity to signal-response coupling by heterotrimeric G-protein components in Arabidopsis. Plant J. 2016, 86, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, R.S.; Pandey, S. Phosphatidic acid binding inhibits RGS1 activity to affect specific signaling pathways in Arabidopsis. Plant J. 2017, 90, 466–477. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Zhang, Y.; Chen, L.; Wang, W.; Liu, R.; Gao, R.; Zhou, Y.; Li, H. G protein and PLDdelta are involved in JA to regulate osmotic stress responses in Arabidopsis thaliana. Biochem. Biophys. Rep. 2021, 26, 100952. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, R.; Zhou, Y.; Wang, S.; Zhang, B.; Kong, J.; Zheng, S.; Yang, N. PLDalpha1 and GPA1 are involved in the stomatal closure induced by Oridonin in Arabidopsis thaliana. Funct. Plant Biol. 2021. [Google Scholar] [CrossRef]
- Hong, Z.; Ueguchi-Tanaka, M.; Umemura, K.; Uozu, S.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 2003, 15, 2900–2910. [Google Scholar] [CrossRef] [Green Version]
- Thung, L.; Trusov, Y.; Chakravorty, D.; Botella, J.R. Ggamma1+Ggamma2+Ggamma3=Gbeta: The search for heterotrimeric G-protein gamma subunits in Arabidopsis is over. J. Plant Physiol. 2012, 169, 542–545. [Google Scholar] [CrossRef]
- Chen, J.G.; Gao, Y.; Jones, A.M. Differential roles of Arabidopsis heterotrimeric G-protein subunits in modulating cell division in roots. Plant Physiol. 2006, 141, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Fujisawa, Y.; Kato, T.; Ohki, S.; Ishikawa, A.; Kitano, H.; Sasaki, T.; Asahi, T.; Iwasaki, Y. Suppression of the heterotrimeric G protein causes abnormal morphology, including dwarfism, in rice. Proc. Natl. Acad. Sci USA 1999, 96, 7575–7580. [Google Scholar] [CrossRef] [Green Version]
- Urano, D.; Jackson, D.; Jones, A.M. A G protein alpha null mutation confers prolificacy potential in maize. J. Exp. Bot. 2015, 66, 4511–4515. [Google Scholar] [CrossRef] [Green Version]
- Bommert, P.; Je, B.I.; Goldshmidt, A.; Jackson, D. The maize Galpha gene COMPACT PLANT2 functions in CLAVATA signalling to control shoot meristem size. Nature 2013, 502, 555–558. [Google Scholar] [CrossRef]
- Zhu, R.; Dong, X.; Hao, W.; Gao, W.; Zhang, W.; Xia, S.; Liu, T.; Shang, Z. Heterotrimeric G Protein-Regulated Ca(2+) Influx and PIN2 Asymmetric Distribution Are Involved in Arabidopsis thaliana Roots’ Avoidance Response to Extracellular ATP. Front Plant Sci. 2017, 8, 1522. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Li, J.; Zhang, L.; Zhang, Z.; He, P.; Wang, W.; Wang, M.; Wang, A.; Zhu, J. Heterotrimeric G-protein alpha subunit (LeGPA1) confers cold stress tolerance to processing tomato plants (Lycopersicon esculentum Mill). BMC Plant Biol. 2020, 20, 394. [Google Scholar] [CrossRef]
- Kumar, R.; Bisht, N.C. Heterotrimeric Galpha subunit regulates plant architecture, organ size and seed weight in the oilseed Brassica juncea. Plant Mol. Biol. 2020, 104, 549–560. [Google Scholar] [CrossRef]
- Sun, H.; Qian, Q.; Wu, K.; Luo, J.; Wang, S.; Zhang, C.; Ma, Y.; Liu, Q.; Huang, X.; Yuan, Q.; et al. Heterotrimeric G proteins regulate nitrogen-use efficiency in rice. Nat. Genet. 2014, 46, 652–656. [Google Scholar] [CrossRef]
- Yan, Y.; Sun, M.; Li, Y.; Wang, J.; He, C.; Yu, X. The CsGPA1-CsAQPs module is essential for salt tolerance of cucumber seedlings. Plant Cell Rep. 2020, 39, 1301–1316. [Google Scholar] [CrossRef]
- Choudhury, S.R.; Pandey, S. Phosphorylation-Dependent Regulation of G-Protein Cycle during Nodule Formation in Soybean. Plant Cell 2015, 27, 3260–3276. [Google Scholar] [CrossRef] [Green Version]
- Llorente, F.; Alonso-Blanco, C.; Sanchez-Rodriguez, C.; Jorda, L.; Molina, A. ERECTA receptor-like kinase and heterotrimeric G protein from Arabidopsis are required for resistance to the necrotrophic fungus Plectosphaerella cucumerina. Plant J. 2005, 43, 165–180. [Google Scholar] [CrossRef]
- Gookin, T.E.; Assmann, S.M. Cantil: A previously unreported organ in wild-type Arabidopsis regulated by FT, ERECTA and heterotrimeric G proteins. Development 2021, 148. [Google Scholar] [CrossRef]
- Xue, J.; Gong, B.Q.; Yao, X.; Huang, X.; Li, J.F. BAK1-mediated phosphorylation of canonical G protein alpha during flagellin signaling in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 690–701. [Google Scholar] [CrossRef]
- Xu, L.; Yao, X.; Zhang, N.; Gong, B.Q.; Li, J.F. Dynamic G protein alpha signaling in Arabidopsis innate immunity. Biochem. Biophys Res. Commun. 2019, 516, 1039–1045. [Google Scholar] [CrossRef]
- Rosenbaum, D.M.; Rasmussen, S.G.; Kobilka, B.K. The structure and function of G-protein-coupled receptors. Nature 2009, 459, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.A.; Kobilka, B.K.; Strader, D.J.; Benovic, J.L.; Dohlman, H.G.; Frielle, T.; Bolanowski, M.A.; Bennett, C.D.; Rands, E.; Diehl, R.E.; et al. Cloning of the gene and cDNA for mammalian beta-adrenergic receptor and homology with rhodopsin. Nature 1986, 321, 75–79. [Google Scholar] [CrossRef]
- Felder, C.C.; Briley, E.M.; Axelrod, J.; Simpson, J.T.; Mackie, K.; Devane, W.A. Anandamide, an endogenous cannabimimetic eicosanoid, binds to the cloned human cannabinoid receptor and stimulates receptor-mediated signal transduction. Proc. Natl. Acad. Sci. USA 1993, 90, 7656–7660. [Google Scholar] [CrossRef] [Green Version]
- Masu, Y.; Nakayama, K.; Tamaki, H.; Harada, Y.; Kuno, M.; Nakanishi, S. cDNA cloning of bovine substance-K receptor through oocyte expression system. Nature 1987, 329, 836–838. [Google Scholar] [CrossRef]
- Jones, A. G-protein-coupled signaling in Arabidopsis. Curr. Opin. Plant Biol. 2002, 5, 402–407. [Google Scholar] [CrossRef]
- Krebs, A.; Villa, C.; Edwards, P.C.; Schertler, G.F. Characterisation of an improved two-dimensional p22121 crystal from bovine rhodopsin. J. Mol. Biol. 1998, 282, 991–1003. [Google Scholar] [CrossRef]
- Lefkowitz, R.J. Historical review: A brief history and personal retrospective of seven-transmembrane receptors. Trends Pharmacol. Sci. 2004, 25, 413–422. [Google Scholar] [CrossRef]
- Venkatakrishnan, A.J.; Deupi, X.; Lebon, G.; Tate, C.G.; Schertler, G.F.; Babu, M.M. Molecular signatures of G-protein-coupled receptors. Nature 2013, 494, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, P.; Park, J.H.; Hildebrand, P.W.; Kim, Y.J.; Krauss, N.; Choe, H.W.; Hofmann, K.P.; Ernst, O.P. Crystal structure of opsin in its G-protein-interacting conformation. Nature 2008, 455, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Lagerstrom, M.C.; Schioth, H.B. Structural diversity of G protein-coupled receptors and significance for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Katritch, V.; Cherezov, V.; Stevens, R.C. Diversity and modularity of G protein-coupled receptor structures. Trends Pharmacol. Sci. 2012, 33, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Samama, P.; Cotecchia, S.; Costa, T.; Lefkowitz, R.J. A mutation-induced activated state of the beta 2-adrenergic receptor. Extending the ternary complex model. J. Biol. Chem. 1993, 268, 4625–4636. [Google Scholar] [CrossRef]
- Blumer, K.J.; Reneke, J.E.; Thorner, J. The STE2 gene product is the ligand-binding component of the alpha-factor receptor of Saccharomyces cerevisiae. J. Biol. Chem. 1988, 263, 10836–10842. [Google Scholar] [CrossRef]
- Anctil, M.; Hayward, D.C.; Miller, D.J.; Ball, E.E. Sequence and expression of four coral G protein-coupled receptors distinct from all classifiable members of the rhodopsin family. Gene 2007, 392, 14–21. [Google Scholar] [CrossRef]
- Carre-Pierrat, M.; Baillie, D.; Johnsen, R.; Hyde, R.; Hart, A.; Granger, L.; Segalat, L. Characterization of the Caenorhabditis elegans G protein-coupled serotonin receptors. Invert Neurosci 2006, 6, 189–205. [Google Scholar] [CrossRef]
- Stafflinger, E.; Hansen, K.K.; Hauser, F.; Schneider, M.; Cazzamali, G.; Williamson, M.; Grimmelikhuijzen, C.J. Cloning and identification of an oxytocin/vasopressin-like receptor and its ligand from insects. Proc. Natl. Acad. Sci. USA 2008, 105, 3262–3267. [Google Scholar] [CrossRef] [Green Version]
- Nathans, J.; Hogness, D.S. Isolation and nucleotide sequence of the gene encoding human rhodopsin. Proc. Natl. Acad. Sci. USA 1984, 81, 4851–4855. [Google Scholar] [CrossRef] [Green Version]
- Rompler, H.; Rohland, N.; Lalueza-Fox, C.; Willerslev, E.; Kuznetsova, T.; Rabeder, G.; Bertranpetit, J.; Schoneberg, T.; Hofreiter, M. Nuclear gene indicates coat-color polymorphism in mammoths. Science 2006, 313, 62. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.; Paiva, A.C.; Vriend, G. A low resolution model for the interaction of G proteins with G protein-coupled receptors. Protein Eng. 1999, 12, 1087–1095. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Moriyama, E.N.; Warr, C.G.; Clyne, P.J.; Carlson, J.R. Identification of novel multi-transmembrane proteins from genomic databases using quasi-periodic structural properties. Bioinformatics 2000, 16, 767–775. [Google Scholar] [CrossRef] [Green Version]
- Gookin, T.E.; Kim, J.; Assmann, S.M. Whole proteome identification of plant candidate G-protein coupled receptors in Arabidopsis, rice, and poplar: Computational prediction and in-vivo protein coupling. Genome. Biol. 2008, 9, R120. [Google Scholar] [CrossRef] [Green Version]
- Josefsson, L.G.; Rask, L. Cloning of a putative G-protein-coupled receptor from Arabidopsis thaliana. Eur. J. Biochem. 1997, 249, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S.; Assmann, S.M. The Arabidopsis putative G protein-coupled receptor GCR1 interacts with the G protein alpha subunit GPA1 and regulates abscisic acid signaling. Plant Cell 2004, 16, 1616–1632. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, N.; Sharma, P.; Kanyuka, K.; Pathak, R.R.; Choudhury, D.; Hooley, R.A.; Raghuram, N. Transcriptome analysis of Arabidopsis GCR1 mutant reveals its roles in stress, hormones, secondary metabolism and phosphate starvation. PLoS ONE 2015, 10, e0117819. [Google Scholar] [CrossRef]
- Chakraborty, N.; Kanyuka, K.; Jaiswal, D.K.; Kumar, A.; Arora, V.; Malik, A.; Gupta, N.; Hooley, R.; Raghuram, N. GCR1 and GPA1 coupling regulates nitrate, cell wall, immunity and light responses in Arabidopsis. Sci. Rep. 2019, 9, 5838. [Google Scholar] [CrossRef]
- Liu, X.; Yue, Y.; Li, B.; Nie, Y.; Li, W.; Wu, W.H.; Ma, L. A G protein-coupled receptor is a plasma membrane receptor for the plant hormone abscisic acid. Science 2007, 315, 1712–1716. [Google Scholar] [CrossRef]
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D.; et al. COLD1 confers chilling tolerance in rice. Cell 2015, 160, 1209–1221. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Huan, Q.; Xu, Y.; Qian, W.; Chong, K.; Zhang, J. Integrated global analysis reveals a vitamin E-vitamin K1 sub-network, downstream of COLD1, underlying rice chilling tolerance divergence. Cell Rep. 2021, 36, 109397. [Google Scholar] [CrossRef]
- Wei, J.; Li, D.X.; Zhang, J.R.; Shan, C.; Rengel, Z.; Song, Z.B.; Chen, Q. Phytomelatonin receptor PMTR1-mediated signaling regulates stomatal closure in Arabidopsis thaliana. J. Pineal Res. 2018, 65, e12500. [Google Scholar] [CrossRef]
- Lu, P.; Magwanga, R.O.; Lu, H.; Kirungu, J.N.; Wei, Y.; Dong, Q.; Wang, X.; Cai, X.; Zhou, Z.; Wang, K.; et al. A Novel G-Protein-Coupled Receptors Gene from Upland Cotton Enhances Salt Stress Tolerance in Transgenic Arabidopsis. Genes 2018, 9, 209. [Google Scholar] [CrossRef] [Green Version]
- Lu, P.; Magwanga, R.O.; Kirungu, J.N.; Dong, Q.; Cai, X.; Zhou, Z.; Wang, X.; Xu, Y.; Hou, Y.; Peng, R.; et al. Genome-wide analysis of the cotton G-coupled receptor proteins (GPCR) and functional analysis of GTOM1, a novel cotton GPCR gene under drought and cold stress. BMC Genom. 2019, 20, 651. [Google Scholar] [CrossRef] [Green Version]
- Urano, D.; Jones, A.M. “Round up the usual suspects”: A comment on nonexistent plant G protein-coupled receptors. Plant Physiol. 2013, 161, 1097–1102. [Google Scholar] [CrossRef] [Green Version]
- Strader, C. Structure and Function of G Protein-Coupled Receptors. Annu. Rev. Biochem. 1994, 63, 101–132. [Google Scholar] [CrossRef]
- Wess, J. Molecular Basis of Receptor/G-Protein-Coupling Selectivity. Pharmacol. Ther. 1998, 80, 231–264. [Google Scholar] [CrossRef]
- Jeon, B.W.; Hwang, J.U.; Hwang, Y.; Song, W.Y.; Fu, Y.; Gu, Y.; Bao, F.; Cho, D.; Kwak, J.M.; Yang, Z.; et al. The Arabidopsis small G protein ROP2 is activated by light in guard cells and inhibits light-induced stomatal opening. Plant Cell 2008, 20, 75–87. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Plant Hormone | Arabidopsis gpa1 | Rice rga1 | Reference |
|---|---|---|---|
| GA | Longer seed dormancy; hyposensitive to GA | Lower expression and activity of α-amylase induced by GA | [20,21,23] |
| BR | Hyposensitive to stomata closure, seed germination, hypocotyl, and root elongation induced by EBR | Weaken effect of root growth, and coleoptile and second coleoptile elongation stimulated by BR | [24,38] |
| ABA | Hyposensitive to ABA inhibition of stomatal opening; weaker activity of inward potassium channels; inhibition of elongation of primary roots | n. d. | [25,27] |
| MeJA | Hyposensitive to MeJA inhibition of stomatal opening; decreased ROS accumulation | n. d. | [28] |
| ET | Hyposensitive to ET promotion of stomatal closure | Hyposensitive to ET | [29,30] |
| Auxin | Reduced cell division in developing hypocotyls and leaves | n. d. | [31,32] |
| Phenotypes | Shorter Hypocotyl; less lateral root; weaker root avoidance of ATP | Shorter rod; round grain | [22,23,31,39,40] |
| Putative Plant GPCRs | Evidence of Interaction with Gα | Transmembrane Structure | Signal Pathways Involved | Reference |
|---|---|---|---|---|
| GCR1 | Pull-down assays; yeast two-hybrid analysis; co-immunoprecipitation (Co-IP) assays | 7-TM | Negative regulator of the GPA1-mediated ABA response; regulation of seed germination by GA and BR | [76,78] |
| GCR2 | Bimolecular fluorescence complementation (BiFC); co-immunoprecipitation (Co-IP) assays | 7-TM | ABA signal transduction | [79] |
| COLD1 | Co-immunoprecipitation (Co-IP) assays; yeast two-hybrid analysis; bimolecular fluorescence complementation assays | 9-TM | Confers chilling tolerance in japonica rice | [80] |
| CAND2 | Bimolecular fluorescence complementation; yeast two-hybrid analysis | 7-TM | Receptor of phytomelatonin | [82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Wang, X.; Dong, D.; Guo, L.; Dong, X.; Leng, J.; Zhao, B.; Guo, Y.-D.; Zhang, N. Research Advances in Heterotrimeric G-Protein α Subunits and Uncanonical G-Protein Coupled Receptors in Plants. Int. J. Mol. Sci. 2021, 22, 8678. https://doi.org/10.3390/ijms22168678
Liu Y, Wang X, Dong D, Guo L, Dong X, Leng J, Zhao B, Guo Y-D, Zhang N. Research Advances in Heterotrimeric G-Protein α Subunits and Uncanonical G-Protein Coupled Receptors in Plants. International Journal of Molecular Sciences. 2021; 22(16):8678. https://doi.org/10.3390/ijms22168678
Chicago/Turabian StyleLiu, Ying, Xiaoyun Wang, Danhui Dong, Luqin Guo, Xiaonan Dong, Jing Leng, Bing Zhao, Yang-Dong Guo, and Na Zhang. 2021. "Research Advances in Heterotrimeric G-Protein α Subunits and Uncanonical G-Protein Coupled Receptors in Plants" International Journal of Molecular Sciences 22, no. 16: 8678. https://doi.org/10.3390/ijms22168678