
Roscovitine, a Cyclin-Dependent Kinase-5 Inhibitor, Decreases Phosphorylated Tau Formation and Death of Retinal Ganglion Cells of Rats after Optic Nerve Crush

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Changes of Phosphorylated Tau after Optic Nerve Crush
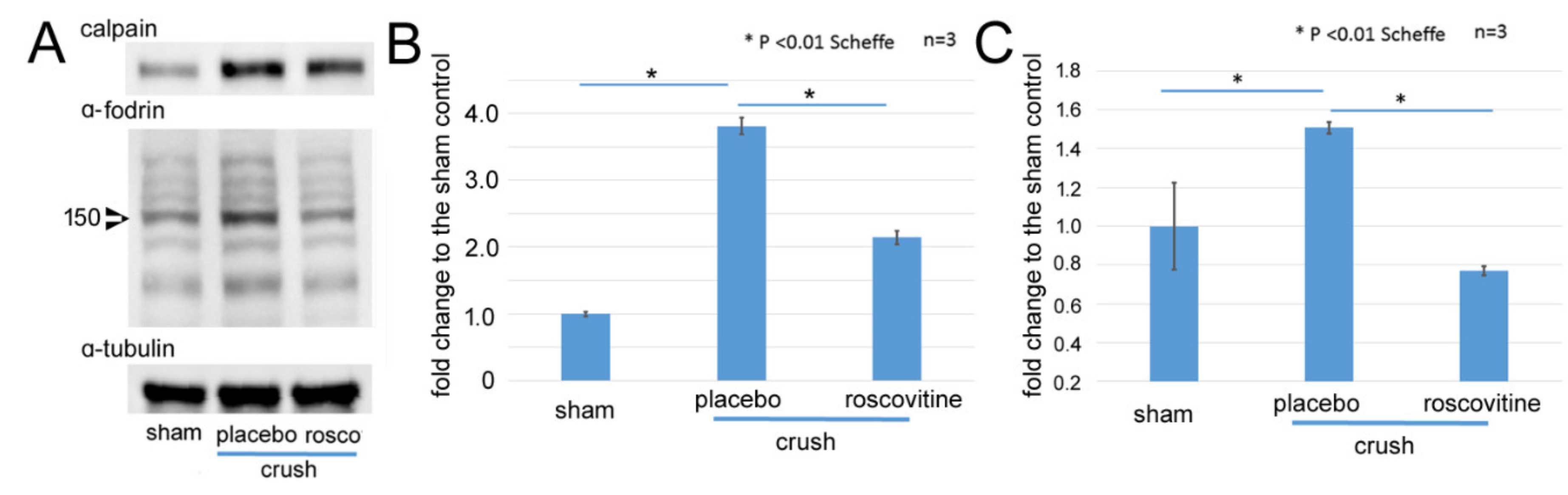
2.2. Changes of Calpain-1 and α-Fodrin after Optic Nerve Crush
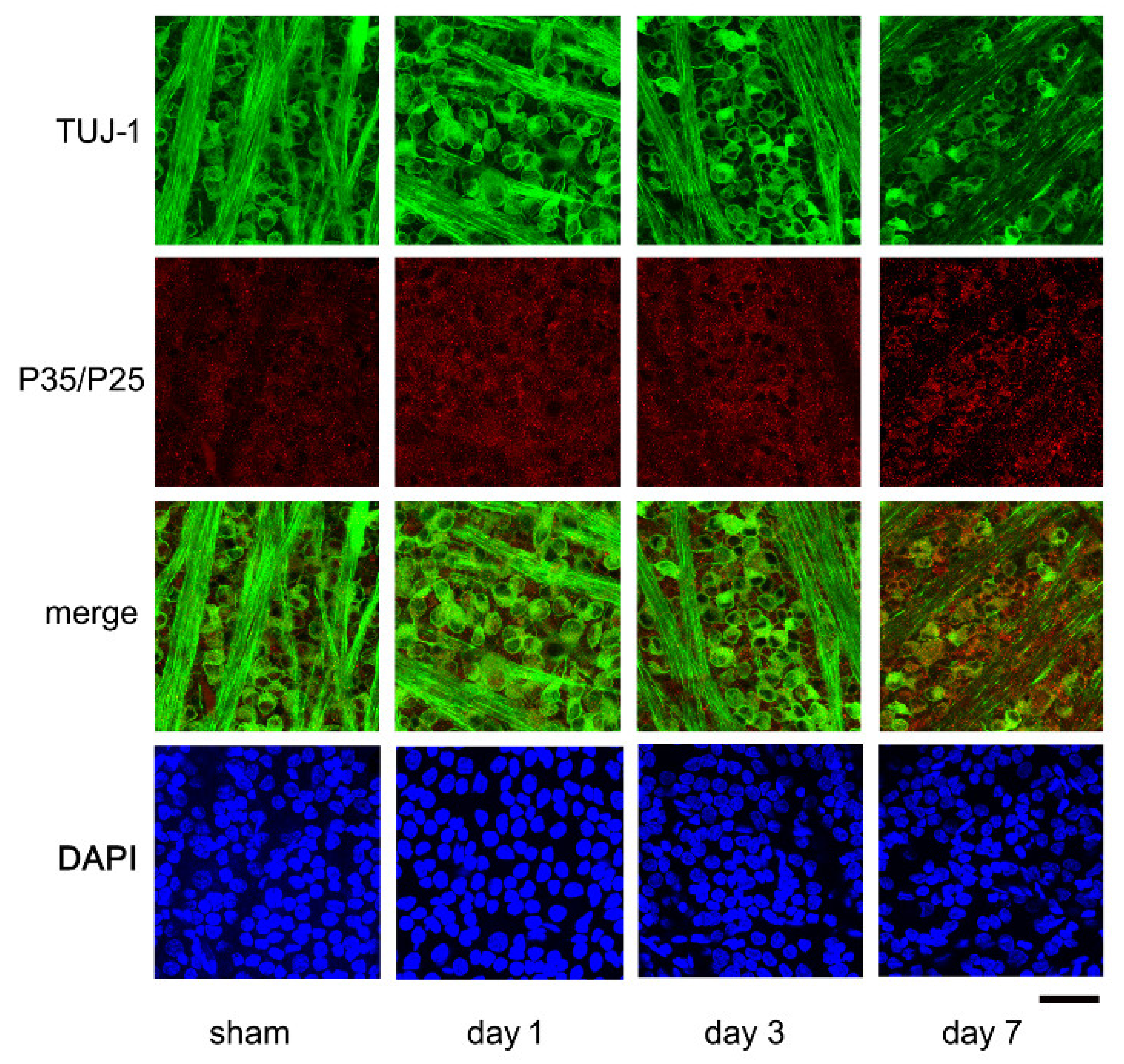
2.3. Expression of P35/P25 by Immunohistochemistry
2.4. Effects of Roscovitine and Calpain Inhibitor on Survival of RGCs after Optic Nerve Crush
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals
4.3. Anesthesia and Euthanasia
4.4. Optic Nerve Crush
4.5. Immunohistochemistry
4.6. Protein Extraction
4.7. Immunoblotting for Expression of Phosphorylated Tau after Optic Nerve Crush
4.8. Statistical Analyses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Goedert, M.; Eisenberg, D.S.; Crowther, R.A. Propagation of Tau Aggregates and Neurodegeneration. Annu. Rev. Neurosci. 2017, 40, 189–210. [Google Scholar] [CrossRef]
- Arendt, T.; Stieler, J.; Holzer, M. Tau and tauopathies. Brain Res. Bull. 2016, 126, 238–292. [Google Scholar] [CrossRef] [PubMed]
- Mroczko, B.; Groblewska, M.; Litman-Zawadzka, A. The Role of Protein Misfolding and Tau Oligomers (TauOs) in Alzheimer’s Disease (AD). Int. J. Mol. Sci. 2019, 20, 4661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oku, H.; Kida, T.; Horie, T.; Taki, K.; Mimura, M.; Kojima, S.; Ikeda, T. Tau Is Involved in Death of Retinal Ganglion Cells of Rats from Optic Nerve Crush. Investig. Opthalmol. Vis. Sci. 2019, 60, 2380–2387. [Google Scholar] [CrossRef]
- Malumbres, M. Cyclin-dependent kinases. Genome Biol. 2014, 15, 122. [Google Scholar] [CrossRef] [Green Version]
- Cruz, J.C.; Tseng, H.-C.; Goldman, J.A.; Shih, H.; Tsai, L.-H. Aberrant Cdk5 Activation by p25 Triggers Pathological Events Leading to Neurodegeneration and Neurofibrillary Tangles. Neuron 2003, 40, 471–483. [Google Scholar] [CrossRef] [Green Version]
- Lopes, J.P.; Agostinho, P. Cdk5: Multitasking between physiological and pathological conditions. Prog. Neurobiol. 2011, 94, 49–63. [Google Scholar] [CrossRef] [Green Version]
- Patrick, G.N.; Zukerberg, L.; Nikolic, M.; de la Monte, S.; Dikkes, P.; Tsai, L.H. Conversion of p35 to p25 deregulates Cdk5 activity and promotes neurodegeneration. Nature 1999, 402, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Ryu, M.; Yasuda, M.; Shi, D.; Shanab, A.Y.; Watanabe, R.; Himori, N.; Omodaka, K.; Yokoyama, Y.; Takano, J.; Saido, T.; et al. Critical role of calpain in axonal damage-induced retinal ganglion cell death. J. Neurosci. Res. 2011, 90, 802–815. [Google Scholar] [CrossRef]
- Taniguchi, S.; Fujita, Y.; Hayashi, S.; Kakita, A.; Takahashi, H.; Murayama, S.; Saido, T.; Hisanaga, S.; Iwatsubo, T.; Hasegawa, M. Calpain-mediated degradation of p35 to p25 in postmortem human and rat brains. FEBS Lett. 2001, 489, 46–50. [Google Scholar] [CrossRef] [Green Version]
- Meijer, L.; Borgne, A.; Mulner, O.; Chong, J.; Blow, J.J.; Inagaki, N.; Inagaki, M.; Delcros, J.-G.; Moulinoux, J.-P. Biochemical and Cellular Effects of Roscovitine, a Potent and Selective Inhibitor of the Cyclin-Dependent Kinases cdc2, cdk2 and cdk5. JBIC J. Biol. Inorg. Chem. 1997, 243, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Chiasseu, M.; Cueva Vargas, J.L.; Destroismaisons, L.; Vande Velde, C.; Leclerc, N.; Di Polo, A. Tau Accumulation, Altered Phosphorylation, and Missorting Promote Neurodegeneration in Glaucoma. J. Neurosci. 2016, 36, 5785–5798. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.K.; Netland, P.A. Early born lineage of retinal neurons express class III beta-tubulin isotype. Brain Res. 2007, 1176, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Dureau, P.; Bonnel, S.; Menasche, M.; Dufier, J.L.; Abitbol, M. Quantitative analysis of intravitreal injections in the rat. Curr. Eye Res. 2001, 22, 74–77. [Google Scholar] [CrossRef]
- Miao, Y.; Dong, L.D.; Chen, J.; Hu, X.C.; Yang, X.L.; Wang, Z. Involvement of calpain/p35-p25/Cdk5/NMDAR signaling pathway in glutamate-induced neurotoxicity in cultured rat retinal neurons. PLoS ONE 2012, 7, e42318. [Google Scholar] [CrossRef]
- Yarotskyy, V.; Elmslie, K.S. Roscovitine Inhibits CaV3.1 (T-Type) Channels by Preferentially Affecting Closed-State Inactivation. J. Pharmacol. Exp. Ther. 2011, 340, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Menn, B.; Bach, S.; Blevins, T.L.; Campbell, M.; Meijer, L.; Timsit, S. Delayed treatment with systemic (S)-roscovitine provides neuroprotection and inhibits in vivo CDK5 activity increase in animal stroke models. PLoS ONE 2010, 5, e12117. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.R.; Nakajima, T.R.; Fukiage, C.R.; Sakai, O.R.; Yoshida, Y.R.; Azuma, M.R.; Shearer, T.R. Involvement of calpain isoforms in ischemia-reperfusion injury in rat retina. Curr. Eye Res. 2000, 21, 571–580. [Google Scholar] [CrossRef]
- McKernan, D.P.; Guerin, M.B.; O’Brien, C.J.; Cotter, T.G. A Key Role for Calpains in Retinal Ganglion Cell Death. Investig. Opthalmol. Vis. Sci. 2007, 48, 5420–5430. [Google Scholar] [CrossRef]
- Maekawa, S.; Sato, K.; Fujita, K.; Daigaku, R.; Tawarayama, H.; Murayama, N.; Moritoh, S.; Yabana, T.; Shiga, Y.; Omodaka, K.; et al. The neuroprotective effect of hesperidin in NMDA-induced retinal injury acts by suppressing oxidative stress and excessive calpain activation. Sci. Rep. 2017, 7, 6885. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Weimer, R.M.; Kallop, D.; Olsen, O.; Wu, Z.; Renier, N.; Uryu, K.; Tessier-Lavigne, M. Regulation of Axon Degeneration after Injury and in Development by the Endogenous Calpain Inhibitor Calpastatin. Neuron 2013, 80, 1175–1189. [Google Scholar] [CrossRef] [Green Version]
- Hung, K.-S.; Hwang, S.-L.; Liang, C.-L.; Chen, Y.-J.; Lee, T.-H.; Liu, J.-K.; Howng, S.-L.; Wang, C.-H. Calpain Inhibitor Inhibits p35-p25-Cdk5 Activation, Decreases Tau Hyperphosphorylation, and Improves Neurological Function after Spinal Cord Hemisection in Rats. J. Neuropathol. Exp. Neurol. 2005, 64, 15–26. [Google Scholar] [CrossRef]
- Saido, T.C.; Yokota, M.; Nagao, S.; Yamaura, I.; Tani, E.; Tsuchiya, T.; Suzuki, K.; Kawashima, S. Spatial resolution of fodrin proteolysis in postischemic brain. J. Biol. Chem. 1993, 268, 25239–25243. [Google Scholar] [CrossRef]
- Donovan, M.; Cotter, T. Caspase-independent photoreceptor apoptosis in vivo and differential expression of apoptotic protease activating factor-1 and caspase-3 during retinal development. Cell Death Differ. 2002, 9, 1220–1231. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Ishiguro, K.; Hisanaga, S.-I. Physiological and pathological phosphorylation of tau by Cdk5. Front. Mol. Neurosci. 2014, 7, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weishaupt, J.H. Inhibition of CDK5 is protective in necrotic and apoptotic paradigms of neuronal cell death and prevents mitochondrial dysfunction. Mol. Cell. Neurosci. 2003, 24, 489–502. [Google Scholar] [CrossRef]
- Chen, J.; Miao, Y.; Wang, X.-H.; Wang, Z. Elevation of p-NR2AS1232 by Cdk5/p35 contributes to retinal ganglion cell apoptosis in a rat experimental glaucoma model. Neurobiol. Dis. 2011, 43, 455–464. [Google Scholar] [CrossRef]
- Lefèvre, K.; Clarke, P.G.; Danthe, E.E.; Castagné, V. Involvement of cyclin-dependent kinases in axotomy-induced retinal ganglion cell death. J. Comp. Neurol. 2002, 447, 72–81. [Google Scholar] [CrossRef]
- Hilton, G.D.; Stoica, B.A.; Byrnes, K.; Faden, A.I. Roscovitine Reduces Neuronal Loss, Glial Activation, and Neurologic Deficits after Brain Trauma. Br. J. Pharmacol. 2008, 28, 1845–1859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahani-Asl, A.; Huang, E.; Irrcher, I.; Rashidian, J.; Ishihara, N.; Lagace, D.C.; Slack, R.S.; Park, D.S. CDK5 phosphorylates DRP1 and drives mitochondrial defects in NMDA-induced neuronal death. Hum. Mol. Genet. 2015, 24, 4573–4583. [Google Scholar] [CrossRef] [Green Version]
- Fraser, M.M.; Zhu, X.; Kwon, C.-H.; Uhlmann, E.J.; Gutmann, D.; Baker, S.J. Pten Loss Causes Hypertrophy and Increased Proliferation of Astrocytes In vivo. Cancer Res. 2004, 64, 7773–7779. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Yang, J.; Xu, Y.; Guo, G.; Cai, L.; Wu, H.; Zhao, Y.; Zhang, X. Roscovitine, a CDK5 Inhibitor, Alleviates Sevoflurane-Induced Cognitive Dysfunction via Regulation Tau/GSK3β and ERK/PPARγ/CREB Signaling. Cell. Physiol. Biochem. 2017, 44, 423–435. [Google Scholar] [CrossRef] [Green Version]
- Tonari, M.; Kurimoto, T.; Horie, T.; Sugiyama, T.; Ikeda, T.; Oku, H. Blocking Endothelin-B Receptors Rescues Retinal Ganglion Cells from Optic Nerve Injury through Suppression of Neuroinflammation. Investig. Opthalmol. Vis. Sci. 2012, 53, 3490–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berkelaar, M.; Clarke, D.; Wang, Y.; Bray, G.; Aguayo, A. Axotomy results in delayed death and apoptosis of retinal ganglion cells in adult rats. J. Neurosci. 1994, 14, 4368–4374. [Google Scholar] [CrossRef] [Green Version]
- Winkler, B.S. The electroretinogram of the isolated rat retina. Vis. Res. 1972, 12, 1183–1198. [Google Scholar] [CrossRef]
- Jiang, S.-M.; Zeng, L.-P.; Zeng, J.-H.; Tang, L.; Chen, X.-M.; Wei, X. β-III-Tubulin: A reliable marker for retinal ganglion cell labeling in experimental models of glaucoma. Int. J. Ophthalmol. 2015, 8, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Abraha, A.; Ghoshal, N.; Gamblin, T.C.; Cryns, V.; Berry, R.W.; Kuret, J.; Binder, L.I. C-terminal inhibition of tau assembly in vitro and in Alzheimer’s disease. J. Cell Sci. 2000, 113 Pt 21, 3737–3745. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirokawa, T.; Horie, T.; Fukiyama, Y.; Mimura, M.; Takai, S.; Kida, T.; Oku, H. Roscovitine, a Cyclin-Dependent Kinase-5 Inhibitor, Decreases Phosphorylated Tau Formation and Death of Retinal Ganglion Cells of Rats after Optic Nerve Crush. Int. J. Mol. Sci. 2021, 22, 8096. https://doi.org/10.3390/ijms22158096
Hirokawa T, Horie T, Fukiyama Y, Mimura M, Takai S, Kida T, Oku H. Roscovitine, a Cyclin-Dependent Kinase-5 Inhibitor, Decreases Phosphorylated Tau Formation and Death of Retinal Ganglion Cells of Rats after Optic Nerve Crush. International Journal of Molecular Sciences. 2021; 22(15):8096. https://doi.org/10.3390/ijms22158096
Chicago/Turabian StyleHirokawa, Takahisa, Taeko Horie, Yurie Fukiyama, Masashi Mimura, Shinji Takai, Teruyo Kida, and Hidehiro Oku. 2021. "Roscovitine, a Cyclin-Dependent Kinase-5 Inhibitor, Decreases Phosphorylated Tau Formation and Death of Retinal Ganglion Cells of Rats after Optic Nerve Crush" International Journal of Molecular Sciences 22, no. 15: 8096. https://doi.org/10.3390/ijms22158096