
Effects of Whole-Body Adenylyl Cyclase 5 (Adcy5) Deficiency on Systemic Insulin Sensitivity and Adipose Tissue

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
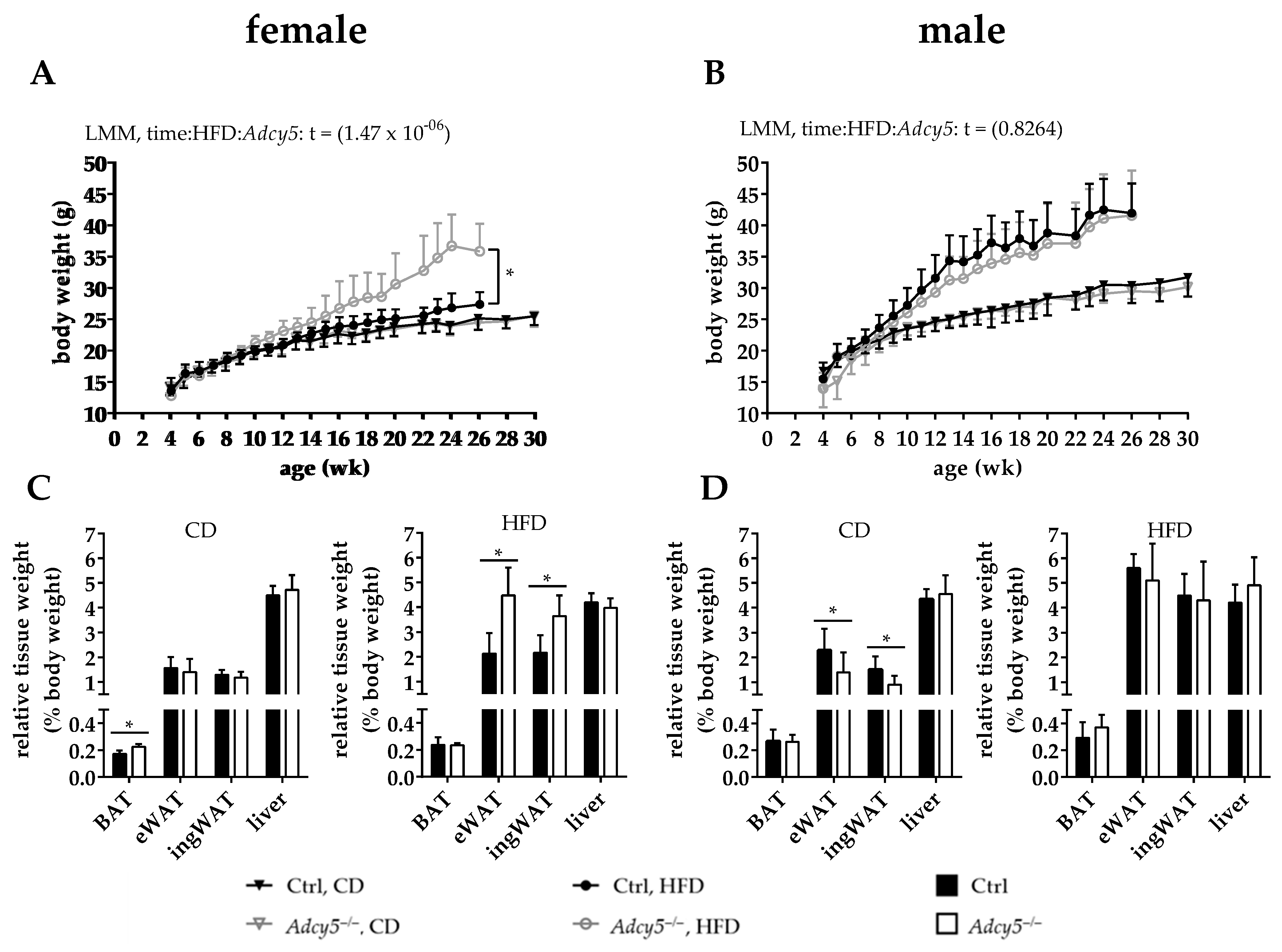
2.1. Adcy5–/– Mice Are Not Protected against Obesity
2.2. Adcy5 Deletion Effects on Glucose Tolerance, Insulin Sensitivity and Serum Parameters
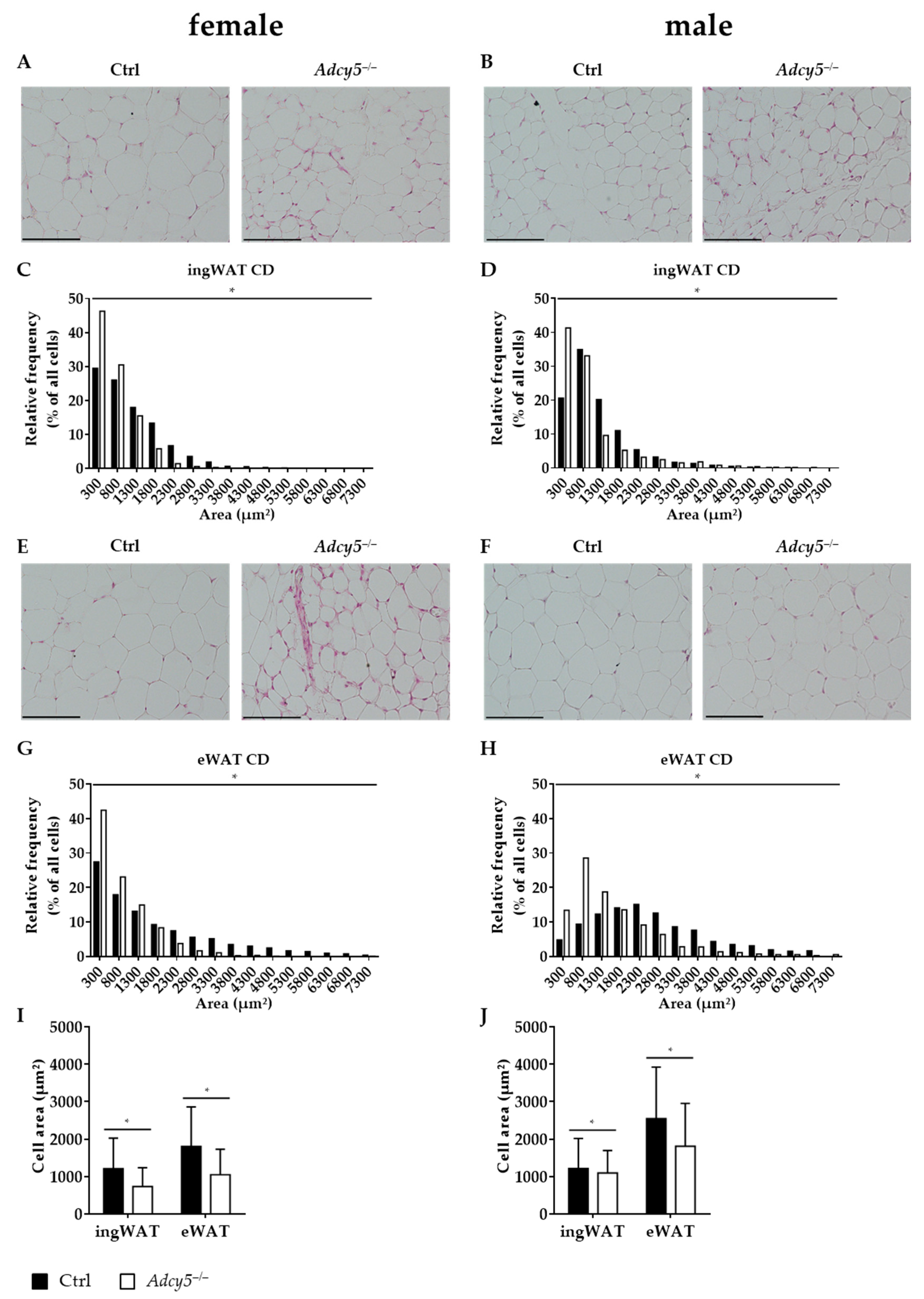
2.3. Consequences of Adcy5–/– on AT Morphology
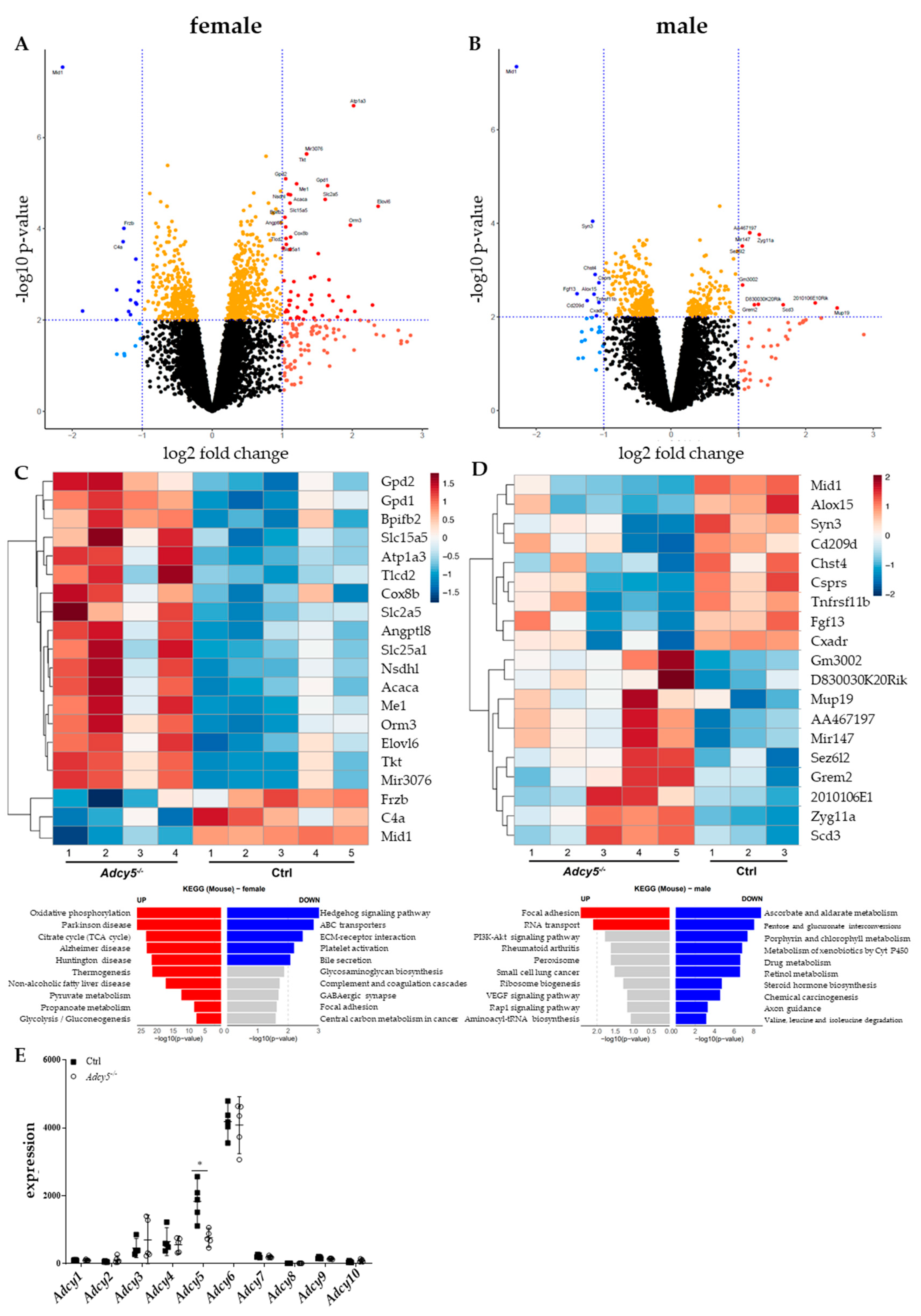
2.4. Adcy5–/– Mice Are Characterized by a Distinct EWAT Gene Expression Signature
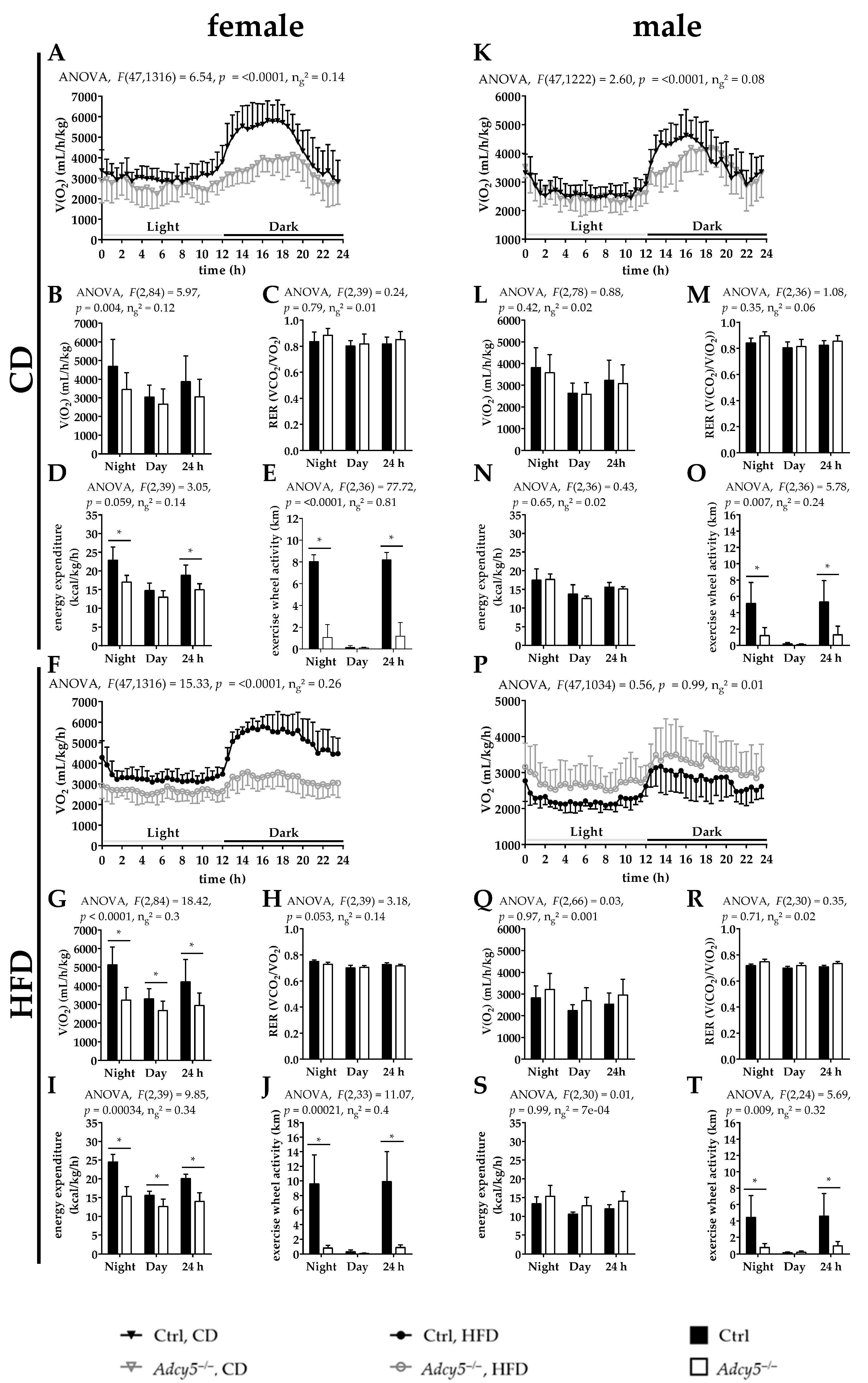
2.5. Adcy5–/– Affects Activity and Energy Expenditure
3. Discussion
4. Materials and Methods
4.1. Animal Studies
4.2. Generation of Adenylyl Cyclase Type 5 Knockout Mice
4.3. Phenotypic Characterization
4.4. Adipose Tissue Histology
4.5. RNA Isolation and Tissue-Specific mRNA Expression
4.6. Adipose Tissue Microarray Analyses
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 36b4 | Rplp0, ribosomal protein, large, P0 |
| ACC | Acetyl-CoA carboxylase |
| Adcy | Adenylyl cyclase |
| AMPK | AMP-activated protein kinase |
| AT | Adipose tissue |
| Atp1a3 | Sodium/potassium-transporting ATPase subunit alpha-3 |
| BAT | Brown adipose tissue |
| BMI | Body mass index |
| cAMP | Cyclic adenosine monophosphate |
| CD | Chow diet |
| CD11c, CD40, CD68, CD163 | Cluster of differentiation 11c, 40, 68 and 163 |
| Ctrl | Control |
| DIO | Diet-induced obesity |
| EDTA | Ethylenediaminetetraacetic acid |
| EE | Energy expenditure |
| Elovl6 | Elongation of very long chain fatty acids protein 6 |
| eWAT | Epigonadal white adipose tissue |
| Fasn | Fatty acid synthase |
| Gpd1 | Glycerol-3-phosphate dehydrogenase |
| GTT | Glucose tolerance test |
| HbA1c | Glycated hemoglobin A1c |
| HDL | High density lipoprotein |
| HFD | High-fat diet |
| i.p. | Intraperitoneal |
| IGF-1 | Insulin-like growth factor 1 |
| IL-17f | Interleukin 17f |
| ingWAT | Inguinal white adipose tissue |
| ITT | Insulin tolerance test |
| LDL | Low density lipoprotein |
| LIMMA | Linear models for microarray data |
| Mid1 | Midline 1 |
| Mid1ip1 | Mid1-interacting protein 1, also Mig12 or Spot14R |
| Nnt | Nicotinamide nucleotide transhydrogenase |
| PKA | Protein kinase A |
| PPARg | Peroxisome proliferator-activated receptor gamma |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| RER | Respiratory exchange ratio |
| RING | Really interesting new gene |
| RMA | Robust multichip average |
| Slc2a5 | Solute carrier family 2, facilitated glucose transporter member 5 |
| SNP | Single nucleotide polymorphism |
| T2D | Type 2 diabetes mellitus |
| TAC | Transcriptome analysis console |
| Thrsp | Thyroid hormone-inducible hepatic protein |
| Tkt | Transketolase |
| TRIM | Tripartite motive family |
References
- Dupuis, J.; DIAGRAM Consortium; Langenberg, C.; Prokopenko, I.; Saxena, R.; Soranzo, N.; Jackson, A.U.; Wheeler, E.; Glazer, N.L.; Bouatia-Naji, N.; et al. New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat. Genet. 2010, 42, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Hodson, D.J.; Mitchell, R.K.; Marselli, L.; Pullen, T.J.; Brias, S.G.; Semplici, F.; Everett, K.L.; Cooper, D.M.; Bugliani, M.; Marchetti, P.; et al. ADCY5 Couples Glucose to Insulin Secretion in Human Islets. Diabetes 2014, 63, 3009–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.K.; Gutierrez, D.A.; Hasty, A.H. Adipose tissue recruitment of leukocytes. Curr. Opin. Lipidol. 2010, 21, 172–177. [Google Scholar] [CrossRef]
- Huopio, H.; Cederberg, H.; Vangipurapu, J.; Hakkarainen, H.; Pääkkönen, M.; Kuulasmaa, T.; Heinonen, S.; Laakso, M. Association of risk variants for type 2 diabetes and hyperglycemia with gestational diabetes. Eur. J. Endocrinol. 2013, 169, 291–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Windholz, J.; Kovacs, P.; Tönjes, A.; Dittrich, K.; Blüher, S.; Kiess, W.; Stumvoll, M.; Körner, A. Effects of Genetic Variants in ADCY5, GIPR, GCKR and VPS13C on Early Impairment of Glucose and Insulin Metabolism in Children. PLoS ONE 2011, 6, e22101. [Google Scholar] [CrossRef] [Green Version]
- Rees, S.D.; Hydrie, M.Z.I.; O’Hare, J.P.; Kumar, S.; Shera, A.S.; Basit, A.; Barnett, A.H.; Kelly, M.A. Effects of 16 Genetic Variants on Fasting Glucose and Type 2 Diabetes in South Asians: ADCY5 and GLIS3 Variants May Predispose to Type 2 Diabetes. PLoS ONE 2011, 6, e24710. [Google Scholar] [CrossRef]
- Wagner, R.; Dudziak, K.; Herzberg-Schäfer, S.A.; Machicao, F.; Stefan, N.; Staiger, H.; Häring, H.-U.; Fritsche, A. Glucose-Raising Genetic Variants in MADD and ADCY5 Impair Conversion of Proinsulin to Insulin. PLoS ONE 2011, 6, e23639. [Google Scholar] [CrossRef]
- Lin, R.; Yuan, Z.; Zhang, C.; Ju, H.; Sun, Y.; Huang, N.; Chen, L.; Jin, L. Common genetic variants in ADCY5 and gestational glycemic traits. PLoS ONE 2020, 15, e0230032. [Google Scholar] [CrossRef] [Green Version]
- Roman, T.S.; Cannon, M.E.; Vadlamudi, S.; Buchkovich, M.L.; Wolford, B.N.; Welch, R.P.; Morken, M.A.; Kwon, G.J.; Varshney, A.; Kursawe, R.; et al. A Type 2 Diabetes–Associated Functional Regulatory Variant in a Pancreatic Islet Enhancer at the ADCY5 Locus. Diabetes 2017, 66, 2521–2530. [Google Scholar] [CrossRef] [Green Version]
- Cash, T.P.; Pita, G.; Domínguez, O.; Alonso, M.R.; Moreno, L.T.; Borrás, C.; Rodríguez-Mañas, L.; Santiago, C.; Garatachea, N.; Lucia, A.; et al. Exome sequencing of three cases of familial exceptional longevity. Aging Cell 2014, 13, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Vatner, D.E.; O’Connor, J.P.; Ivessa, A.; Ge, H.; Chen, W.; Hirotani, S.; Ishikawa, Y.; Sadoshima, J.; Vatner, S.F. Type 5 Adenylyl Cyclase Disruption Increases Longevity and Protects Against Stress. Cell 2007, 130, 247–258. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M.; Dodson Michael, M.; Peroni, O.D.; Ueki, K.; Carter, N.; Kahn, B.B.; Kahn, C. Adipose Tissue Selective Insulin Receptor Knockout Protects against Obesity and Obesity-Related Glucose Intolerance. Dev. Cell 2002, 3, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.; Zhao, X.; Yan, L.; Yuan, C.; Zong, H.; Vatner, D.E.; Pessin, J.E.; Vatner, S.F. Adenylyl Cyclase Type 5 Deficiency Protects Against Diet-Induced Obesity and Insulin Resistance. Diabetes 2015, 64, 2636–2645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumura, S.; Kawabe, J.; Yatani, A.; Takagi, G.; Lee, M.-C.; Hong, C.; Liu, J.; Takagi, I.; Sadoshima, J.; Vatner, D.E.; et al. Type 5 Adenylyl Cyclase Disruption Alters Not Only Sympathetic But Also Parasympathetic and Calcium-Mediated Cardiac Regulation. Circ. Res. 2003, 93, 364–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwatsubo, K.; Bravo, C.; Uechi, M.; Baljinnyam, E.; Nakamura, T.; Umemura, M.; Lai, L.; Gao, S.; Yan, L.; Zhao, X.; et al. Prevention of heart failure in mice by an antiviral agent that inhibits type 5 cardiac adenylyl cyclase. Am. J. Physiol. Circ. Physiol. 2012, 302, H2622–H2628. [Google Scholar] [CrossRef]
- Vatner, S.F.; Park, M.; Yan, L.; Lee, G.J.; Lai, L.; Iwatsubo, K.; Ishikawa, Y.; Pessin, J.; Vatner, D.E. Adenylyl cyclase type 5 in cardiac disease, metabolism, and aging. Am. J. Physiol. Circ. Physiol. 2013, 305, H1–H8. [Google Scholar] [CrossRef] [Green Version]
- Knigge, A.; Klöting, N.; Schön, M.R.; Dietrich, A.; Fasshauer, M.; Gärtner, D.; Lohmann, T.; Dreßler, M.; Stumvoll, M.; Kovacs, P.; et al. ADCY5 Gene Expression in Adipose Tissue Is Related to Obesity in Men and Mice. PLoS ONE 2015, 10, e0120742. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Park, J.Y.; Dillinger, J.-G.; De Lorenzo, M.S.; Yuan, C.; Lai, L.; Wang, C.; Ho, D.; Tian, B.; Stanley, W.C.; et al. Common mechanisms for calorie restriction and adenylyl cyclase type 5 knockout models of longevity. Aging Cell 2012, 11, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Blüher, M.; Kahn, B.B.; Kahn, C.R. Extended Longevity in Mice Lacking the Insulin Receptor in Adipose Tissue. Science 2003, 299, 572–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiesenborn, D.S.; Menon, V.; Zhi, X.; Do, A.; Gesing, A.; Wang, Z.; Bartke, A.; Altomare, D.A.; Masternak, M.M. The effect of calorie restriction on insulin signaling in skeletal muscle and adipose tissue of Ames dwarf mice. Aging 2014, 6, 900–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weindruch, R.; Walford, R. Dietary restriction in mice beginning at 1 year of age: Effect on life-span and spontaneous cancer incidence. Science 1982, 215, 1415–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walford, R.L.; Crew, M. How dietary restriction retards aging: An integrative hypothesis. Growth Dev. Aging 1989, 53, 139–140. [Google Scholar] [PubMed]
- Holzenberger, M.; Dupont, J.; Ducos, B.; Leneuve, P.; Géloën, A.; Even, P.C.; Cervera, P.; Le Bouc, Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nat. Cell Biol. 2002, 421, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Okumura, S.; Takagi, G.; Kawabe, J.-I.; Yang, G.; Lee, M.-C.; Hong, C.; Liu, J.; Vatner, D.E.; Sadoshima, J.; Vatner, S.F.; et al. Disruption of type 5 adenylyl cyclase gene preserves cardiac function against pressure overload. Proc. Natl. Acad. Sci. USA 2003, 100, 9986–9990. [Google Scholar] [CrossRef] [Green Version]
- Hong, J.; Stubbins, R.E.; Smith, R.R.; Harvey, A.E.; Núñez, N.P. Differential susceptibility to obesity between male, female and ovariectomized female mice. Nutr. J. 2009, 8, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stubbins, R.E.; Holcomb, V.B.; Hong, J.; Núñez, N.P. Estrogen modulates abdominal adiposity and protects female mice from obesity and impaired glucose tolerance. Eur. J. Nutr. 2011, 51, 861–870. [Google Scholar] [CrossRef]
- NCBI. Adcy5 Adenylate Cyclase 5 [Mus Musculus (House Mouse)]—Gene. Available online: https://www.ncbi.nlm.nih.gov/gene/224129 (accessed on 19 October 2020).
- Simpson, E.R.; Mahendroo, M.S.; Means, G.D.; Kilgore, M.W.; Hinshelwood, M.M.; Graham-Lorence, S.; Amarneh, B.; Ito, Y.; Fisher, C.R.; Michael, M.D.; et al. Aromatase Cytochrome P450, The Enzyme Responsible for Estrogen Biosynthesis. Endocr. Rev. 1994, 15, 342–355. [Google Scholar] [CrossRef]
- Freeman, H.C.; Hugill, A.; Dear, N.T.; Ashcroft, F.M.; Cox, R.D. Deletion of Nicotinamide Nucleotide Transhydrogenase: A New Quantitive Trait Locus Accounting for Glucose Intolerance in C57BL/6J Mice. Diabetes 2006, 55, 2153–2156. [Google Scholar] [CrossRef] [Green Version]
- Kunath, A.; Heiker, J.T.; Kern, M.; Kosacka, J.; Flehmig, G.; Stumvoll, M.; Kovacs, P.; Blüher, M.; Klöting, N. Nicotinamide Nucleotide Transhydrogenase (Nnt) is Related to Obesity in Mice. Horm. Metab. Res. 2020, 52, 877–881. [Google Scholar]
- Heiker, J.T.; Kunath, A.; Kosacka, J.; Flehmig, G.; Knigge, A.; Kern, M.; Stumvoll, M.; Kovacs, P.; Blüher, M.; Klöting, N. Identification of genetic loci associated with different responses to high-fat diet-induced obesity in C57BL/6N and C57BL/6J substrains. Physiol. Genom. 2014, 46, 377–384. [Google Scholar] [CrossRef]
- Madsen, L.; Myrmel, L.S.; Fjære, E.; Liaset, B.; Kristiansen, K. Links between Dietary Protein Sources, the Gut Microbiota, and Obesity. Front. Physiol. 2017, 8, 1047. [Google Scholar] [CrossRef] [Green Version]
- Fjære, E.; Myrmel, L.S.; Lützhøft, D.O.; Andersen, H.; Holm, J.B.; Kiilerich, P.; Hannisdal, R.; Liaset, B.; Kristiansen, K.; Madsen, L. Effects of exercise and dietary protein sources on adiposity and insulin sensitivity in obese mice. J. Nutr. Biochem. 2019, 66, 98–109. [Google Scholar] [CrossRef]
- Roberts, R.; Hodson, L.; Dennis, A.L.; Neville, M.J.; Humphreys, S.M.; Harnden, K.E.; Micklem, K.J.; Frayn, K.N. Markers of de novo lipogenesis in adipose tissue: Associations with small adipocytes and insulin sensitivity in humans. Diabetologia 2009, 52, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Cotillard, A.; Poitou, C.; Torcivia, A.; Bouillot, J.-L.; Dietrich, A.; Klöting, N.; Grégoire, C.; Lolmede, K.; Blüher, M.; Clément, K. Adipocyte Size Threshold Matters: Link with Risk of Type 2 Diabetes and Improved Insulin Resistance After Gastric Bypass. J. Clin. Endocrinol. Metab. 2014, 99, E1466–E1470. [Google Scholar] [CrossRef]
- Tang, W.; Ma, W.; Ding, H.; Lin, M.; Xiang, L.; Lin, G.; Zhang, Z. Adenylyl cyclase 1 as a major isoform to generate cAMP signaling for apoA-1-mediated cholesterol efflux pathway. J. Lipid Res. 2018, 59, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Bergman, B.C.; Jensen, D.R.; Pulawa, L.K.; Ferreira, L.D.-B.; Eckel, R.H. Fasting decreases free fatty acid turnover in mice overexpressing skeletal muscle lipoprotein lipase. Metabolism 2006, 55, 1481–1487. [Google Scholar] [CrossRef] [PubMed]
- Baldini, R.; Mascaro, M.; Meroni, G. The MID1 gene product in physiology and disease. Gene 2020, 747, 144655. [Google Scholar] [CrossRef] [PubMed]
- Berti, C.; Fontanella, B.; Ferrentino, R.; Meroni, G. Mig12, a novel Opitz syndrome gene product partner, is expressed in the embryonic ventral midline and co-operates with Mid1 to bundle and stabilize microtubules. BMC Cell Biol. 2004, 5, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.J.; Sim, D.Y.; Lee, H.M.; Lee, H.-J.; Kim, S.-H. Hypolipogenic Effect of Shikimic Acid Via Inhibition of MID1IP1 and Phosphorylation of AMPK/ACC. Int. J. Mol. Sci. 2019, 20, 582. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMP-Activated Protein Kinase: Maintaining Energy Homeostasis at the Cellular and Whole-Body Levels. Annu. Rev. Nutr. 2014, 34, 31–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, D.G.; Pan, D.A. Regulation of fatty acid synthesis and oxidation by the AMP-activated protein kinase. Biochem. Soc. Trans. 2002, 30, 1064–1070. [Google Scholar] [CrossRef]
- Papandreou, A.; Danti, F.R.; Spaull, R.; Leuzzi, V.; McTague, A.; A Kurian, M. The expanding spectrum of movement disorders in genetic epilepsies. Dev. Med. Child Neurol. 2019, 62, 178–191. [Google Scholar] [CrossRef] [PubMed]
- Carecchio, M.; Mencacci, N.E.; Iodice, A.; Pons, R.; Panteghini, C.; Zorzi, G.; Zibordi, F.; Bonakis, A.; Dinopoulos, A.; Jankovic, J.; et al. ADCY5-related movement disorders: Frequency, disease course and phenotypic variability in a cohort of paediatric patients. Park. Relat. Disord. 2017, 41, 37–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carapito, R.; Paul, N.; Untrau, M.; Le Gentil, M.; Ott, L.; Alsaleh, G.; Jochem, P.; Radosavljevic, M.; Le Caignec, C.; David, A.; et al. A de novo ADCY5 mutation causes early-onset autosomal dominant chorea and dystonia. Mov. Disord. 2015, 30, 423–427. [Google Scholar] [CrossRef]
- Vijiaratnam, N.; Newby, R.; Kempster, P.A. Depression and psychosis in ADCY5-related dyskinesia—Part of the phenotypic spectrum? J. Clin. Neurosci. 2018, 57, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, Y.; Park, J.-Y.; Kim, J.-E.; Kim, T.-K.; Choi, J.; Lee, J.-E.; Lee, E.-H.; Kim, D.; Kim, K.-S.; et al. Loss of Adenylyl Cyclase Type-5 in the Dorsal Striatum Produces Autistic-Like Behaviors. Mol. Neurobiol. 2017, 54, 7994–8008. [Google Scholar] [CrossRef]
- Kern, M.; Kosacka, J.; Hesselbarth, N.; Brückner, J.; Heiker, J.T.; Flehmig, G.; Klöting, I.; Kovacs, P.; Matz-Soja, M.; Gebhardt, R.; et al. Liver-Restricted Repin1 Deficiency Improves Whole-Body Insulin Sensitivity, Alters Lipid Metabolism, and Causes Secondary Changes in Adipose Tissue in Mice. Diabetes 2014, 63, 3295–3309. [Google Scholar] [CrossRef] [Green Version]
- Heiker, J.T.; Klöting, N.; Kovacs, P.; Kuettner, E.B.; Sträter, N.; Schultz, S.; Kern, M.; Stumvoll, M.; Blüher, M.; Beck-Sickinger, A.G. Vaspin inhibits kallikrein 7 by serpin mechanism. Cell. Mol. Life Sci. 2013, 70, 2569–2583. [Google Scholar] [CrossRef] [Green Version]
- Kern, M.; Klöting, N.; Niessen, H.G.; Thomas, L.; Stiller, D.; Mark, M.; Klein, T.; Blüher, M. Linagliptin improves insulin sensitivity and hepatic steatosis in diet-induced obesity. PLoS ONE 2012, 7, e38744. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, B.S.; Irizarry, R.A. A framework for oligonucleotide microarray preprocessing. Bioinformatics 2010, 26, 2363–2367. [Google Scholar] [CrossRef]
- Irizarry, R.A.; Hobbs, B.; Collin, F.; Beazer-Barclay, Y.D.; Antonellis, K.J.; Scherf, U.; Speed, T.P. Exploration, normalization, and summaries of high density oligonucleotide array probe level data. In Selected Works of Terry Speed; Springer: New York, NY, USA, 2012; Volume 4, pp. 601–616. ISBN 9781461413479. [Google Scholar]
- Huber, W.; Carey, V.J.; Gentleman, R.; Anders, S.; Carlson, M.; Carvalho, B.S.; Bravo, H.C.; Davis, S.; Gatto, L.; Girke, T.; et al. Orchestrating high-throughput genomic analysis with Bioconductor. Nat. Methods 2015, 12, 115–121. [Google Scholar] [CrossRef]
- Kauffmann, A.; Gentleman, R.; Huber, W. arrayQualityMetrics—a bioconductor package for quality assessment of microarray data. Bioinformatics 2008, 25, 415–416. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kassambara, A. Pipe-Friendly Framework for Basic Statistical Tests—Rstatix. Available online: https://rpkgs.datanovia.com/rstatix/ (accessed on 12 March 2021).
- Haynes, W. Bonferroni Correction. In Encyclopedia of Systems Biology; Springer: New York, NY, USA, 2013; p. 154. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Mauchly, J.W. Significance Test for Sphericity of a Normal n-Variate Distribution. Ann. Math. Stat. 1940, 11, 204–209. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CD | HFD | |||||
|---|---|---|---|---|---|---|
| Ctrl | Adcy5–/– | n | Ctrl | Adcy5–/– | n | |
| Serum Lipids | ||||||
| Triglycerides (mmol/L) | 1.07 ± 0.32 | 1.10 ± 0.20 | 4 vs. 5 | 1.03 ± 0.47 | 1.31 ± 0.22 | 5 vs. 5 |
| Cholesterol (mmol/L) | 2.16 ± 0.22 | 1.53 ± 0.23 | 5 vs. 5 | 2.98 ± 0.28 | 3.50 ± 0.30 | 5 vs. 5 |
| HDL-cholesterol (mmol/L) | 1.67 ± 0.20 | 1.21 ± 0.20 | 5 vs. 5 | 2.23 ± 0.27 | 2.78 ± 0.26 | 5 vs. 5 |
| LDL-cholesterol (mmol/L) | 0.35 ± 0.07 | 0.18 ± 0.03 | 5 vs. 5 | 0.73 ± 0.07 | 0.58 ± 0.07 | 5 vs. 5 |
| FFA (mmol/L) | 1.50 ± 0.30 | 1.67 ± 0.35 | 5 vs. 5 | 1.71 ± 0.23 | 1.82 ± 0.26 | 5 vs. 5 |
| OLINK Protein serum Analysis (Normalized Protein Expression (AU)) | ||||||
| Il17f | 3.43 ± 0.20 | 1.84 ± 0.26 | 3 vs. 4 | 5.05 ± 0.80 | 2.79 ± 0.23 | 3 vs. 3 |
| Glucose Homeostasis | ||||||
| C-peptide (ng/mL) | 0.60 ± 0.10 | 0.40 ± 0.19 | 5 vs. 5 | 1.18 ± 0.53 | 1.11 ± 0.35 | 5 vs. 5 |
| Insulin (µg/L) | 0.28 ± 0.12 | 0.19 ± 0.03 | 5 vs. 5 | 0.69 ± 0.63 | 0.65 ± 0.48 | 5 vs. 5 |
| Adiponectin (µg/mL) | 208.5 ± 56.1 | 242.5 ± 105.7 | 5 vs. 5 | 140.1 ± 8.3 | 145.6 ± 4.3 | 5 vs. 5 |
| Leptin (ng/mL) | 3.51 ± 1.12 | 3.96 ± 5.55 | 5 vs. 5 | 7.02 ± 2.83 | 22.80 ± 10.61 | 5 vs. 5 |
| Leptin/body weight (ng/mL/g) | 0.15 ± 0.04 | 0.17 ± 0.24 | 5 vs. 5 | 0.27 ± 0.10 | 0.69 ± 0.31 | 5 vs. 5 |
| Fasting glucose (mmol/L) | 4.57 ± 1.14 | 4.23 ± 1.34 | 12 vs. 12 | 4.96 ± 1.43 | 6.10 ± 1.85 | 14 vs. 11 |
| HbA1c (%) | 4.20 ± 0.12 | 3.94 ± 0.18 | 12 vs. 11 | 4.09 ± 0.14 | 4.09 ± 0.14 | 13 vs. 11 |
| CD | HFD | |||||
|---|---|---|---|---|---|---|
| Ctrl | Adcy5–/– | n | Ctrl | Adcy5–/– | n | |
| Serum Lipids | ||||||
| Triglycerides (mmol/L) | 1.08 ± 0.09 | 1.11 ± 0.22 | 5 vs. 5 | 1.54 ± 0.10 | 5 vs. 5 | |
| Cholesterol (mmol/L) | 2.62 ± 0.49 | 2.07 ± 0.10 | 5 vs. 5 | 4.30 ± 0.15 | 5.05 ± 0.40 | 5 vs. 5 |
| HDL-cholesterol (mmol/L) | 2.07 ± 0.33 | 1.77 ± 0.09 | 5 vs. 5 | 3.28 ± 0.14 | 3.77 ± 0.20 | 5 vs. 5 |
| LDL-cholesterol (mmol/L) | 0.39 ± 0.10 | 0.15 ± 0.03 | 5 vs. 5 | 0.83 ± 0.20 | 1.04 ± 0.15 | 5 vs. 5 |
| FFA (mmol/L) | 1.53 ± 0.09 | 1.47 ± 0.26 | 5 vs. 5 | 1.86 ± 0.08 | 1.99 ± 0.19 | 5 vs. 5 |
| OLINK Protein Serum Analysis (Normalized Protein Expression (AU)) | ||||||
| Il17f | 3.17 ± 0.28 | 1.56 ± 0.23 | 3 vs. 4 | 5.21 ± 1.39 | 3.14 ± 0.89 | 3 vs. 4 |
| Glucose Homeostasis | ||||||
| C-peptide (ng/mL) | 1.08 ± 0.41 | 0.45 ± 0.11 | 5 vs. 5 | 2.28 ± 1.37 | 4.56 ± 1.47 | 5 vs. 5 |
| Insulin (µg/L) | 0.34 ± 0.15 | 0.17 ± 0.06 | 5 vs. 5 | 1.47 ± 1.05 | 1.81 ± 0.75 | 4 vs. 5 |
| Adiponectin (µg/mL) | 135.2 ± 13.1 | 107.9 ± 20.7 | 5 vs. 5 | 88.3 ± 7.9 | 85.4 ± 10.7 | 5 vs. 5 |
| Leptin (ng/mL) | 7.69 ± 4.40 | 2.80 ± 1.26 | 5 vs. 4 | 39.8 ± 1.30 | 40.4 ± 1.17 | 5 vs. 5 |
| Leptin/body weight (ng/mL/g) | 0.25 ± 0.12 | 0.10 ± 0.05 | 5 vs. 4 | 1.00 ± 0.05 | 0.92 ± 0.02 | 5 vs. 5 |
| Fasting glucose (mmol/L) | 4.94 ± 1.44 | 3.44 ± 0.55 | 12 vs. 12 | 5.28 ± 2.20 | 6.18 ± 1.37 | 14 vs. 13 |
| HbA1c (%) | 4.48 ± 0.15 | 4.29 ± 0.13 | 12 vs. 12 | 4.31 ± 0.17 | 4.34 ± 0.15 | 8 vs. 13 |
| HOMA-β (%) | 86.6 ± 38.3 | 190.2 ± 15.3 | 5 vs. 5 | 318 ± 265 | 281 ± 118 | 4 vs. 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dommel, S.; Hoffmann, A.; Berger, C.; Kern, M.; Klöting, N.; Kannt, A.; Blüher, M. Effects of Whole-Body Adenylyl Cyclase 5 (Adcy5) Deficiency on Systemic Insulin Sensitivity and Adipose Tissue. Int. J. Mol. Sci. 2021, 22, 4353. https://doi.org/10.3390/ijms22094353
Dommel S, Hoffmann A, Berger C, Kern M, Klöting N, Kannt A, Blüher M. Effects of Whole-Body Adenylyl Cyclase 5 (Adcy5) Deficiency on Systemic Insulin Sensitivity and Adipose Tissue. International Journal of Molecular Sciences. 2021; 22(9):4353. https://doi.org/10.3390/ijms22094353
Chicago/Turabian StyleDommel, Sebastian, Anne Hoffmann, Claudia Berger, Matthias Kern, Nora Klöting, Aimo Kannt, and Matthias Blüher. 2021. "Effects of Whole-Body Adenylyl Cyclase 5 (Adcy5) Deficiency on Systemic Insulin Sensitivity and Adipose Tissue" International Journal of Molecular Sciences 22, no. 9: 4353. https://doi.org/10.3390/ijms22094353