
Analysis of Sequence Divergence in Mammalian ABCGs Predicts a Structural Network of Residues That Underlies Functional Divergence

 ,
,  , , and
, , and 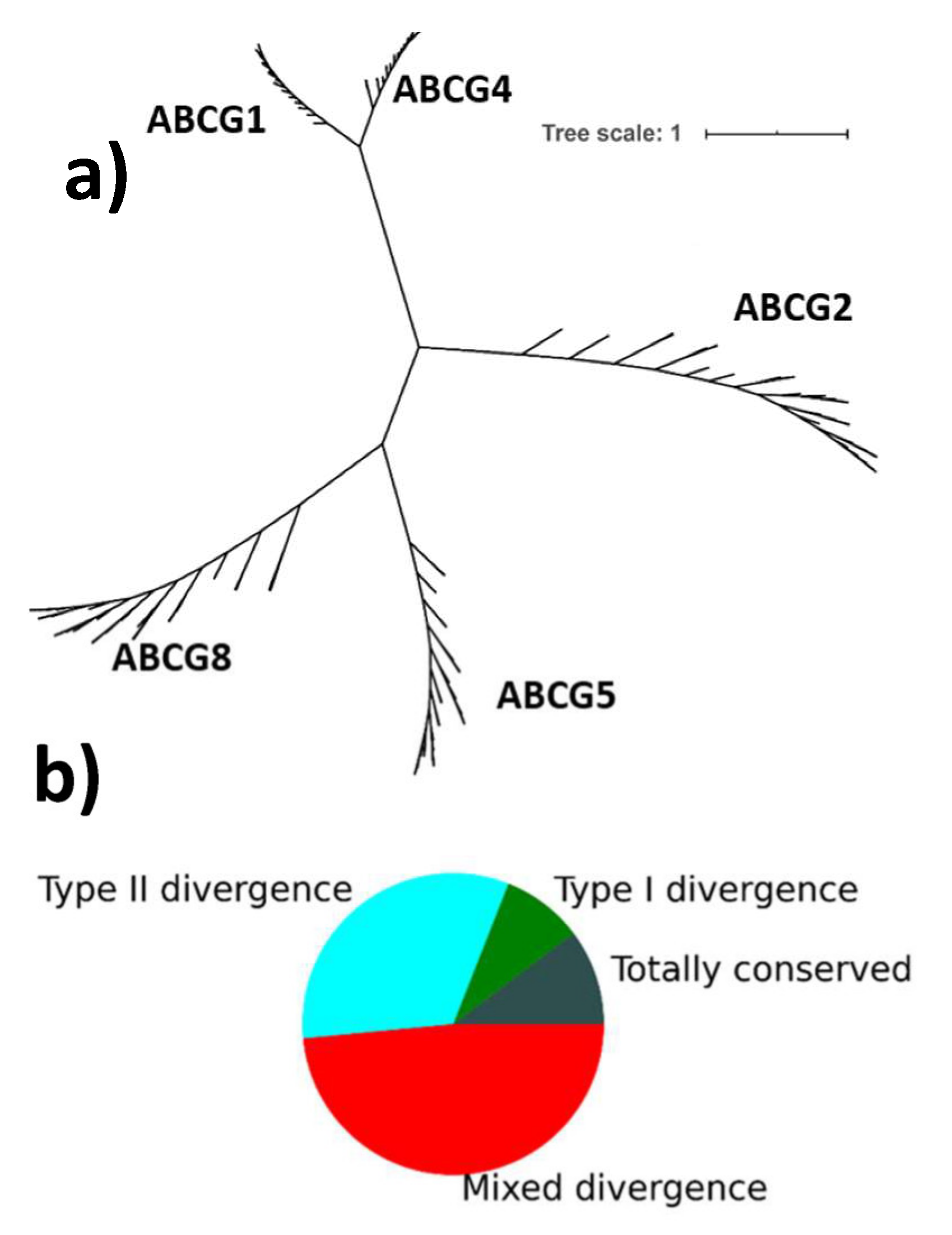{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Overall Conservation Patterns
2.2. Phylogenetically Significant Type II Divergence in ABCGs
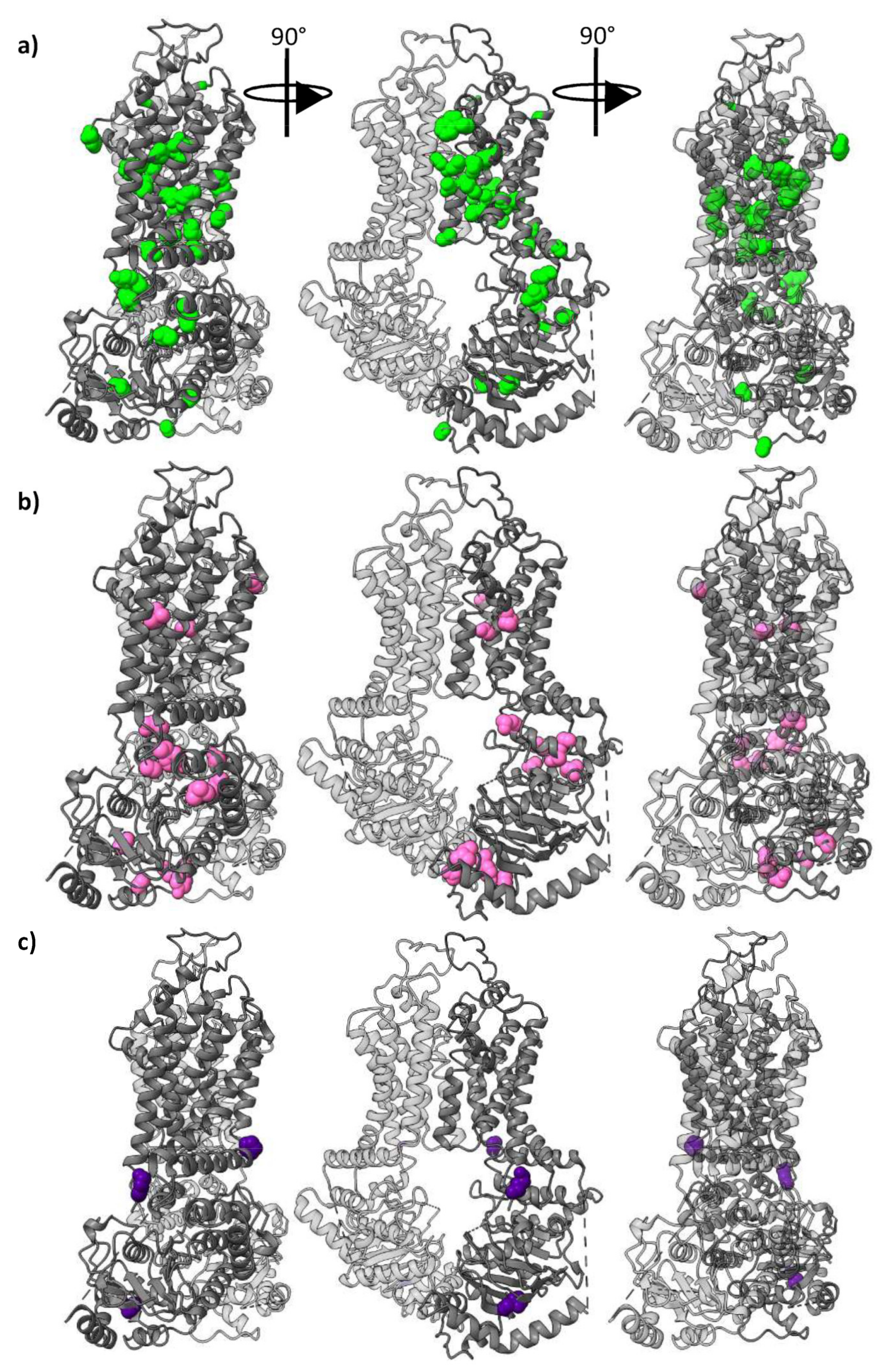
2.3. The Conservation Pattern (ABCG1, ABCG4), (ABCG2), (ABCG5), (ABCG8) Defines a Possible Allosteric Pathway in ABCG Proteins
2.4. Conservation of the Polar Relay
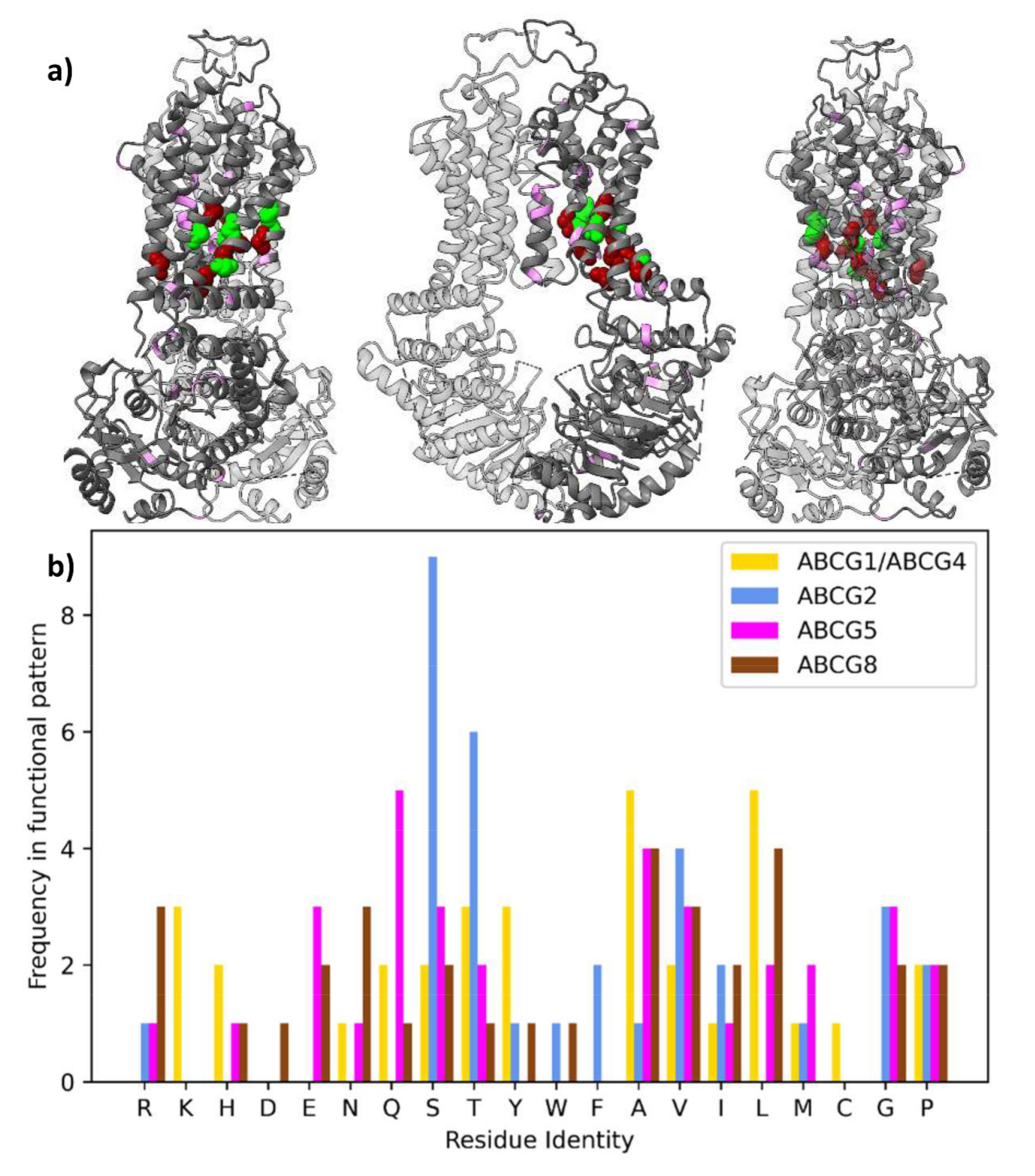
2.5. Sidechain Properties in the Allosteric Corkscrew
2.6. Conservation of Other Regions
2.7. Other Conservation Patterns
3. Discussion
4. Materials and Methods
4.1. Sequence Acquisition
4.2. Alignment and Tree Construction
4.3. Calculation of Conservation
4.4. Binding Pockets
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef]
- Kerr, I.D.; Haider, A.J.; Gelissen, I.C. The ABCG family of membrane-associated transporters: You don’t have to be big to be mighty. Br. J. Pharmacol. 2011, 164, 1767–1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaughan, A.M.; Oram, J.F. ABCA1 and ABCG1 or ABCG4 act sequentially to remove cellular cholesterol and generate cholesterol-rich HDL. J. Lipid Res. 2006, 47, 2433–2443. [Google Scholar] [CrossRef] [Green Version]
- Hegyi, Z.; Homolya, L. Functional Cooperativity between ABCG4 and ABCG1 Isoforms. PLoS ONE 2016, 11, e0156516. [Google Scholar] [CrossRef] [Green Version]
- Cserepes, J.; Szentpétery, Z.; Seres, L.; Özvegy-Laczka, C.; Langmann, T.; Schmitz, G.; Glavinas, H.; Klein, I.; Homolya, L.; Váradi, A.; et al. Functional expression and characterization of the human ABCG1 and ABCG4 proteins: Indications for heterodimerization. Biochem. Biophys. Res. Commun. 2004, 320, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-W.; Graf, G.A.; Gerard, R.D.; Cohen, J.C.; Hobbs, H.H. Functional Asymmetry of Nucleotide-binding Domains in ABCG5 and ABCG8. J. Biol. Chem. 2006, 281, 4507–4516. [Google Scholar] [CrossRef] [Green Version]
- Allikmets, R.; Schriml, L.M.; Hutchinson, A.; Romano-Spica, V.; Dean, M. A human placenta-specific ATP-binding cassette gene (ABCP) on chromosome 4q22 that is involved in multidrug resistance. Cancer Res. 1998, 58, 5337–5339. [Google Scholar] [PubMed]
- Doyle, L.A.; Yang, W.; Abruzzo, L.V.; Krogmann, T.; Gao, Y.; Rishi, A.K.; Ross, D.D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 15665–15670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-Y.; Kinch, L.N.; Borek, D.M.; Wang, J.; Wang, J.; Urbatsch, I.L.; Xie, X.-S.; Grishin, N.V.; Cohen, J.C.; Otwinowski, Z.; et al. Crystal structure of the human sterol transporter ABCG5/ABCG8. Nat. Cell Biol. 2016, 533, 561–564. [Google Scholar] [CrossRef] [Green Version]
- László, L.; Sarkadi, B.; Hegedűs, T. Jump into a New Fold—A Homology Based Model for the ABCG2/BCRP Multidrug Transporter. PLoS ONE 2016, 11, e0164426. [Google Scholar] [CrossRef] [Green Version]
- Clark, R.; Kerr, I.D.; Callaghan, R. Multiple drugbinding sites on the R482G isoform of the ABCG2 transporter. Br. J. Pharmacol. 2006, 149, 506–515. [Google Scholar] [CrossRef]
- Taylor, N.M.I.; Manolaridis, I.; Jackson, S.M.; Kowal, J.; Stahlberg, H.; Locher, K.P. Structure of the human multidrug transporter ABCG2. Nat. Cell Biol. 2017, 546, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Manolaridis, I.; Jackson, S.M.; Taylor, N.M.I.; Kowal, J.; Stahlberg, H.; Locher, K.P. Cryo-EM structures of a human ABCG2 mutant trapped in ATP-bound and substrate-bound states. Nat. Cell Biol. 2018, 563, 426–430. [Google Scholar] [CrossRef] [Green Version]
- Jackson, S.M.; Manolaridis, I.; Kowal, J.; Zechner, M.; Taylor, N.M.I.; Bause, M.; Bauer, S.; Bartholomaeus, R.; Bernhardt, G.; Koenig, B.; et al. Structural basis of small-molecule inhibition of human multidrug transporter ABCG2. Nat. Struct. Mol. Biol. 2018, 25, 333–340. [Google Scholar] [CrossRef]
- Orlando, B.J.; Liao, M. ABCG2 transports anticancer drugs via a closed-to-open switch. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Gu, X. Maximum-Likelihood Approach for Gene Family Evolution Under Functional Divergence. Mol. Biol. Evol. 2001, 18, 453–464. [Google Scholar] [CrossRef] [Green Version]
- Gu, X. Functional Divergence in Protein (Family) Sequence Evolution. Genetica 2003, 118, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Lopez, P.; Casane, D.; Philippe, H. Heterotachy, an Important Process of Protein Evolution. Mol. Biol. Evol. 2002, 19, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Xavier, B.M.; Zein, A.A.; Venes, A.; Wang, J.; Lee, J.-Y. Transmembrane Polar Relay Drives the Allosteric Regulation for ABCG5/G8 Sterol Transporter. bioRxiv 2020. [Google Scholar] [CrossRef]
- Tarr, P.T.; Edwards, P.A. ABCG1 and ABCG4 are coexpressed in neurons and astrocytes of the CNS and regulate cholesterol homeostasis through SREBP-2. J. Lipid Res. 2008, 49, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Pozza, A.; Pérez-Victoria, J.M.; Sardo, A.; Ahmed-Belkacem, A.; Di Pietro, A. Purification of breast cancer resistance protein ABCG2 and role of arginine-482. Cell. Mol. Life Sci. 2006, 63, 1912–1922. [Google Scholar] [CrossRef]
- Robey, R.W.; Honjo, Y.; Morisaki, K.; Nadjem, T.A.; Runge, S.; Risbood, M.; Poruchynsky, M.S.; Bates, S.E. Mutations at amino-acid 482 in the ABCG2 gene affect substrate and antagonist specificity. Br. J. Cancer 2003, 89, 1971–1978. [Google Scholar] [CrossRef] [PubMed]
- Khunweeraphong, N.; Mitchell-White, J.; Szöllősi, D.; Hussein, T.; Kuchler, K.; Kerr, I.D.; Stockner, T.; Lee, J. Picky ABCG5/G8 and promiscuous ABCG2—a tale of fatty diets and drug toxicity. FEBS Lett. 2020, 594, 4035–4058. [Google Scholar] [CrossRef]
- Zhou, F.; Cocco, M.J.; Russ, W.P.; Brunger, A.T.; Engelman, D.M. Interhelical hydrogen bonding drives strong interactions in membrane proteins. Nat. Genet. 2000, 7, 154–160. [Google Scholar] [CrossRef]
- Zhou, F.X.; Merianos, H.J.; Brunger, A.T.; Engelman, D.M. Polar residues drive association of polyleucine transmembrane helices. Proc. Natl. Acad. Sci. USA 2001, 98, 2250–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gratkowski, H.; Lear, J.D.; DeGrado, W.F. Polar side chains drive the association of model transmembrane peptides. Proc. Natl. Acad. Sci. USA 2001, 98, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Dawson, J.P.; Weinger, J.S.; Engelman, D.M. Motifs of serine and threonine can drive association of transmembrane helices. J. Mol. Biol. 2002, 316, 799–805. [Google Scholar] [CrossRef]
- North, B.; Cristian, L.; Stowell, X.F.; Lear, J.D.; Saven, J.G.; DeGrado, W.F. Characterization of a Membrane Protein Folding Motif, the Ser Zipper, Using Designed Peptides. J. Mol. Biol. 2006, 359, 930–939. [Google Scholar] [CrossRef]
- Gray, T.; Matthews, B. Intrahelical hydrogen bonding of serine, threonine and cysteine residues within α-helices and its relevance to membrane-bound proteins. J. Mol. Biol. 1984, 175, 75–81. [Google Scholar] [CrossRef]
- Ballesteros, J.A.; Deupi, X.; Olivella, M.; Haaksma, E.E.; Pardo, L. Serine and Threonine Residues Bend α-Helices in the χ1=g− Conformation. Biophys. J. 2000, 79, 2754–2760. [Google Scholar] [CrossRef] [Green Version]
- Deupi, X.; Edwards, P.; Singhal, A.; Nickle, B.; Oprian, D.; Schertler, G.; Standfuss, J. Stabilized G protein binding site in the structure of constitutively active metarhodopsin-II. Proc. Natl. Acad. Sci. USA 2011, 109, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Del Torrent, C.L.; Casajuana-Martin, N.; Pardo, L.; Tresadern, G.; Pérez-Benito, L. Mechanisms Underlying Allosteric Molecular Switches of Metabotropic Glutamate Receptor 5. J. Chem. Inf. Model. 2019, 59, 2456–2466. [Google Scholar] [CrossRef]
- Khunweeraphong, N.; Stockner, T.; Kuchler, K. The structure of the human ABC transporter ABCG2 reveals a novel mechanism for drug extrusion. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, P.; Briggs, D.A.; Cox, M.H.; Kerr, I.D. Disruption of the Unique ABCG-Family NBD:NBD Interface Impacts Both Drug Transport and ATP Hydrolysis. Int. J. Mol. Sci. 2020, 21, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gose, T.; Shafi, T.; Fukuda, Y.; Das, S.; Wang, Y.; Allcock, A.; McHarg, A.G.; Lynch, J.; Chen, T.; Tamai, I.; et al. ABCG2 requires a single aromatic amino acid to “clamp” substrates and inhibitors into the binding pocket. FASEB J. 2020, 34, 4890–4903. [Google Scholar] [CrossRef] [Green Version]
- Cox, M.H.; Kapoor, P.; Briggs, D.A.; Kerr, I.D. Residues contributing to drug transport by ABCG2 are localised to multiple drug-binding pockets. Biochem. J. 2018, 475, 1553–1567. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Bikádi, Z.; Cai, X.; Rosenberg, M.F.; Mao, Q. Transmembrane helices 1 and 6 of the human breast cancer resistance protein (BCRP/ABCG2): Identification of polar residues important for drug transport. Am. J. Physiol. Physiol. 2010, 299, C1100–C1109. [Google Scholar] [CrossRef] [Green Version]
- Polgar, O.; Ierano’, C.; Tamaki, A.; Stanley, B.; Ward, Y.; Xia, D.; Tarasova, N.; Robey, R.W.; Bates, S.E. Mutational Analysis of Threonine 402 Adjacent to the GXXXG Dimerization Motif in Transmembrane Segment 1 of ABCG2. Biochemistry 2010, 49, 2235–2245. [Google Scholar] [CrossRef] [Green Version]
- Tamura, A.; Wakabayashi, K.; Onishi, Y.; Takeda, M.; Ikegami, Y.; Sawada, S.; Tsuji, M.; Matsuda, Y.; Ishikawa, T. Re-evaluation and functional classification of non-synonymous single nucleotide polymorphisms of the human ATP-binding cassette transporter ABCG2. Cancer Sci. 2007, 98, 231–239. [Google Scholar] [CrossRef]
- Tamura, A.; Watanabe, M.; Saito, H.; Nakagawa, H.; Kamachi, T.; Okura, I.; Ishikawa, T. Functional Validation of the Genetic Polymorphisms of Human ATP-Binding Cassette (ABC) Transporter ABCG2: Identification of Alleles That Are Defective in Porphyrin Transport. Mol. Pharmacol. 2006, 70, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, H.; Tamura, A.; Wakabayashi, K.; Hoshijima, K.; Komada, M.; Yoshida, T.; Kometani, S.; Matsubara, T.; Mikuriya, K.; Ishikawa, T. Ubiquitin-mediated proteasomal degradation of non-synonymous SNP variants of human ABC transporter ABCG2. Biochem. J. 2008, 411, 623–631. [Google Scholar] [CrossRef] [Green Version]
- Deppe, S.; Ripperger, A.; Weiss, J.; Ergün, S.; Benndorf, R.A. Impact of genetic variability in the ABCG2 gene on ABCG2 expression, function, and interaction with AT1 receptor antagonist telmisartan. Biochem. Biophys. Res. Commun. 2014, 443, 1211–1217. [Google Scholar] [CrossRef]
- Sjöstedt, N.; Heuvel, J.J.M.W.V.D.; Koenderink, J.B.; Kidron, H. Transmembrane Domain Single-Nucleotide Polymorphisms Impair Expression and Transport Activity of ABC Transporter ABCG2. Pharm. Res. 2017, 34, 1626–1636. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, Y.; Mančíková, A.; Krylov, V.; Morimoto, K.; Pavelcová, K.; Bohatá, J.; Pavelka, K.; Pavlíková, M.; Suzuki, H.; Matsuo, H.; et al. Functional Characterization of Clinically-Relevant Rare Variants in ABCG2 Identified in a Gout and Hyperuricemia Cohort. Cells 2019, 8, 363. [Google Scholar] [CrossRef] [Green Version]
- Polgar, O.; Ediriwickrema, L.S.; Robey, R.W.; Sharma, A.; Hegde, R.S.; Li, Y.; Xia, D.; Ward, Y.; Dean, M.; Ozvegy-Laczka, C.; et al. Arginine 383 is a crucial residue in ABCG2 biogenesis. Biochim. Biophys. Acta Biomembr. 2009, 1788, 1434–1443. [Google Scholar] [CrossRef] [Green Version]
- Ni, Z.; Bikádi, Z.; Shuster, D.L.; Zhao, C.; Rosenberg, M.F.; Mao, Q. Identification of Proline Residues in or near the Transmembrane Helices of the Human Breast Cancer Resistance Protein (BCRP/ABCG2) That Are Important for Transport Activity and Substrate Specificity. Biochemistry 2011, 50, 8057–8066. [Google Scholar] [CrossRef] [Green Version]
- Keller, S.; Prechtl, D.; Aslanidis, C.; Ceglarek, U.; Thiery, J.; Schmitz, G.; Jahreis, G. Increased plasma plant sterol concentrations and a heterozygous amino acid exchange in ATP binding cassette transporter ABCG5: A case report. Eur. J. Med. Genet. 2011, 54, e458–e460. [Google Scholar] [CrossRef]
- Heimer, S.; Langmann, T.; Moehle, C.; Mauerer, R.; Dean, M.; Beil, F.-U.; Von Bergmann, K.; Schmitz, G. Mutations in the human ATP-binding cassette transportersABCG5 andABCG8 in sitosterolemia. Hum. Mutat. 2002, 20, 151. [Google Scholar] [CrossRef]
- Niu, D.-M.; Chong, K.-W.; Hsu, J.-H.; Wu, T.J.-T.; Yu, H.-C.; Huang, C.-H.; Lo, M.-Y.; Kwok, C.F.; Kratz, L.E.; Ho, L.-T. Clinical observations, molecular genetic analysis, and treatment of sitosterolemia in infants and children. J. Inherit. Metab. Dis. 2010, 33, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Heyes, N.; Kapoor, P.; Kerr, I.D. Polymorphisms of the Multidrug Pump ABCG2: A Systematic Review of Their Effect on Protein Expression, Function, and Drug Pharmacokinetics. Drug Metab. Dispos. 2018, 46, 1886–1899. [Google Scholar] [CrossRef] [Green Version]
- Abellán, R.; Mansego, M.L.; Martínez-Hervás, S.; Martín-Escudero, J.C.; Carmena, R.; Real, J.T.; Redon, J.; Castrodeza-Sanz, J.J.; Chaves, F.J. Association of selected ABC gene family single nucleotide polymorphisms with postprandial lipoproteins: Results from the population-based Hortega study. Atherosclerosis 2010, 211, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, M.; Lütjohann, D.; Schirin-Sokhan, R.; Villarroel, L.; Nervi, F.; Pimentel, F.; Lammert, F.; Miquel, J.F. Phytosterol and cholesterol precursor levels indicate increased cholesterol excretion and biosynthesis in gallstone disease. Hepatology 2012, 55, 1507–1517. [Google Scholar] [CrossRef]
- Hubacek, J.A.; Berge, K.E.; Cohen, J.C.; Hobbs, H.H. Mutations in ATP-cassette binding proteins G5 (ABCG5) and G8 (ABCG8) causing sitosterolemia. Hum. Mutat. 2001, 18, 359–360. [Google Scholar] [CrossRef]
- Lee, M.-H.; Lu, K.; Patel, S.B. Genetic basis of sitosterolemia. Curr. Opin. Lipidol. 2001, 12, 141–149. [Google Scholar] [CrossRef]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of Dietary Cholesterol in Sitosterolemia Caused by Mutations in Adjacent ABC Transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef] [Green Version]
- Pandit, B.; Ahn, G.-S.; Hazard, S.E.; Gordon, D.; Patel, S.B. A detailed Hapmap of the Sitosterolemia locus spanning 69 kb; differences between Caucasians and African-Americans. BMC Med. Genet. 2006, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- National Center for Biotechnology Information. The NCBI Handbook, 2nd ed.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- Rossum, G.V.; Drake, F.L. Python 3 Reference Manual; CreateSpace: Scotts Valley, CA, USA, 2009. [Google Scholar]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Capra, J.A.; Singh, M. Characterization and prediction of residues determining protein functional specificity. Bioinformatics 2008, 24, 1473–1480. [Google Scholar] [CrossRef] [Green Version]
- Tareen, A.; Kinney, J.B. Logomaker: Beautiful sequence logos in Python. Bioinformatics 2020, 36, 2272–2274. [Google Scholar] [CrossRef]
- Goddard, T.D.; Huang, C.C.; Meng, E.C.; Pettersen, E.F.; Couch, G.S.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Meeting modern challenges in visualization and analysis. Protein Sci. 2018, 27, 14–25. [Google Scholar] [CrossRef]
- Haider, A.J.; Cox, M.H.; Jones, N.; Goode, A.J.; Bridge, K.S.; Wong, K.; Briggs, D.; Kerr, I.D. Identification of residues in ABCG2 affecting protein trafficking and drug transport, using co-evolutionary analysis of ABCG sequences. Biosci. Rep. 2015, 35, e00241. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, T.A.; Klett, E.L.; Gylling, H.; Isoniemi, H.; Patel, S.B. Liver Transplantation in a Patient with Sitosterolemia and Cirrhosis. Gastroenterology 2006, 130, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Lee, M.-H.; Hazard, S.; Brooks-Wilson, A.; Hidaka, H.; Kojima, H.; Ose, L.; Stalenhoef, A.F.; Mietinnen, T.; Bjorkhem, I.; et al. Two Genes That Map to the STSL Locus Cause Sitosterolemia: Genomic Structure and Spectrum of Mutations Involving Sterolin-1 and Sterolin-2, Encoded by ABCG5 and ABCG8, Respectively. Am. J. Hum. Genet. 2001, 69, 278–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, T.; Doyle, L.A.; Hassel, B.; Wei, Y.; Bauer, K.S.; Wu, S.; Pumplin, D.W.; Fang, H.-B.; Ross, U.D. Functional Characterization of Human Breast Cancer Resistance Protein (BCRP, ABCG2) Expressed in the Oocytes of Xenopus laevis. Mol. Pharmacol. 2003, 64, 1452–1462. [Google Scholar] [CrossRef] [PubMed]
- Buch, S.; Schafmayer, C.; Völzke, H.; Becker, C.; Franke, A.; Von Eller-Eberstein, H.; Kluck, C.; Bässmann, I.; Brosch, M.; Lammert, F.; et al. A genome-wide association scan identifies the hepatic cholesterol transporter ABCG8 as a susceptibility factor for human gallstone disease. Nat. Genet. 2007, 39, 995–999. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mitchell-White, J.I.; Stockner, T.; Holliday, N.; Briddon, S.J.; Kerr, I.D. Analysis of Sequence Divergence in Mammalian ABCGs Predicts a Structural Network of Residues That Underlies Functional Divergence. Int. J. Mol. Sci. 2021, 22, 3012. https://doi.org/10.3390/ijms22063012
Mitchell-White JI, Stockner T, Holliday N, Briddon SJ, Kerr ID. Analysis of Sequence Divergence in Mammalian ABCGs Predicts a Structural Network of Residues That Underlies Functional Divergence. International Journal of Molecular Sciences. 2021; 22(6):3012. https://doi.org/10.3390/ijms22063012
Chicago/Turabian StyleMitchell-White, James I., Thomas Stockner, Nicholas Holliday, Stephen J. Briddon, and Ian D. Kerr. 2021. "Analysis of Sequence Divergence in Mammalian ABCGs Predicts a Structural Network of Residues That Underlies Functional Divergence" International Journal of Molecular Sciences 22, no. 6: 3012. https://doi.org/10.3390/ijms22063012