
The Effectiveness of Anti-Apoptotic Agents to Preserve Primordial Follicles and Prevent Tissue Damage during Ovarian Tissue Cryopreservation and Xenotransplantation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
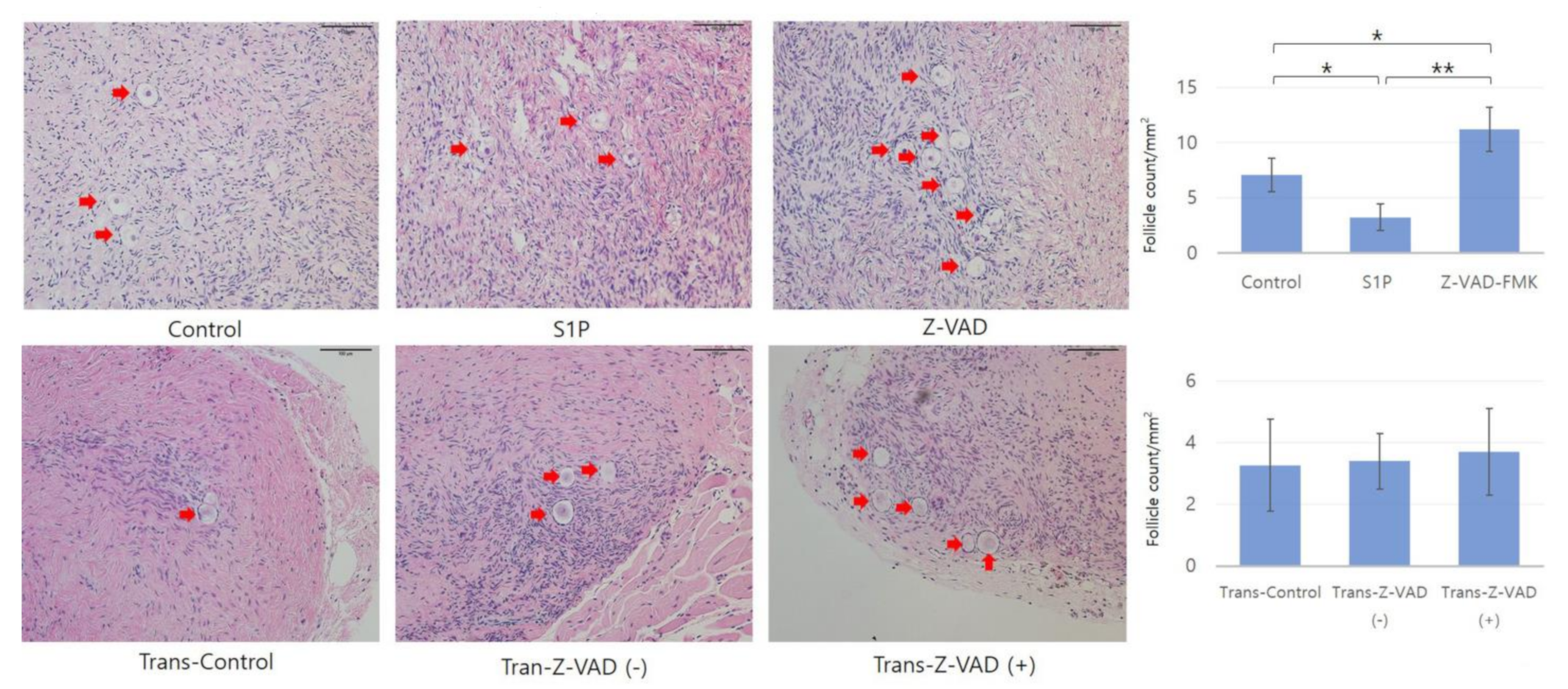
2.1. Histologic Evaluation and Primordial Follicular Counts
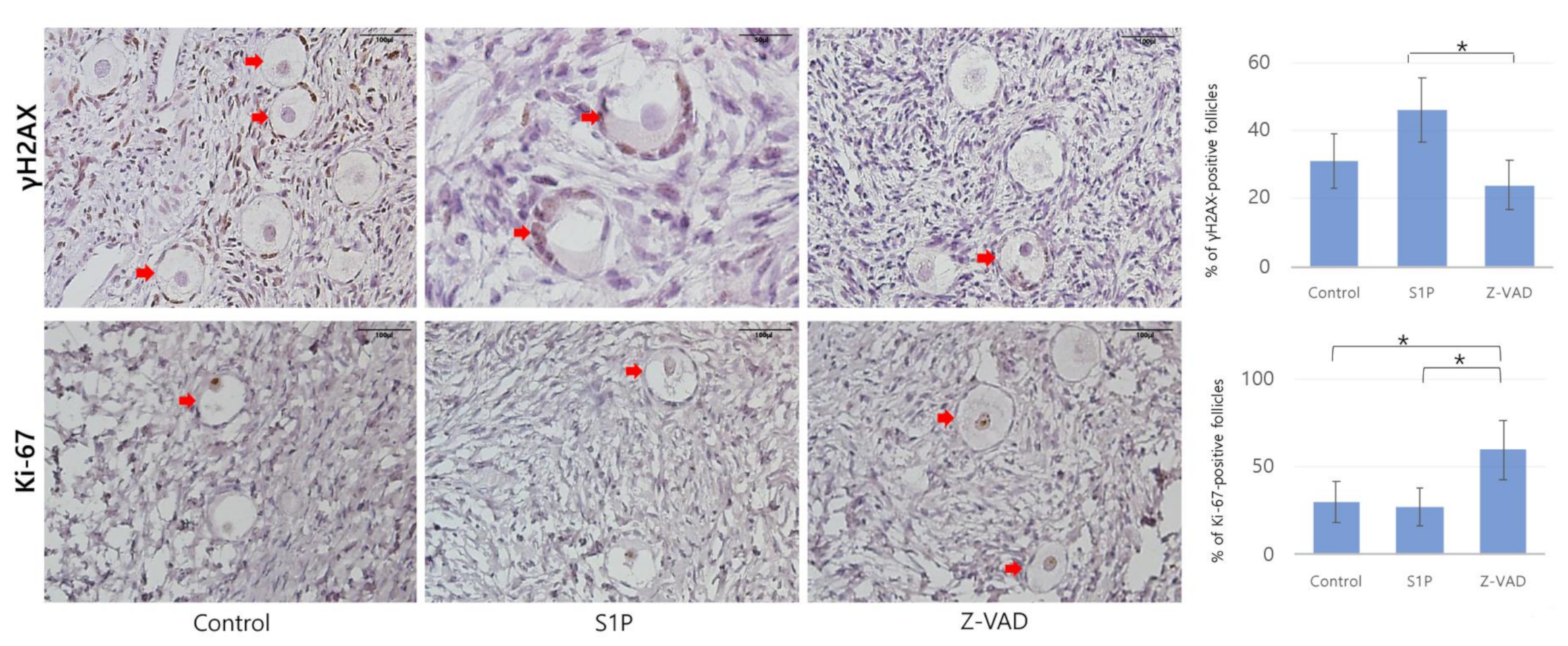
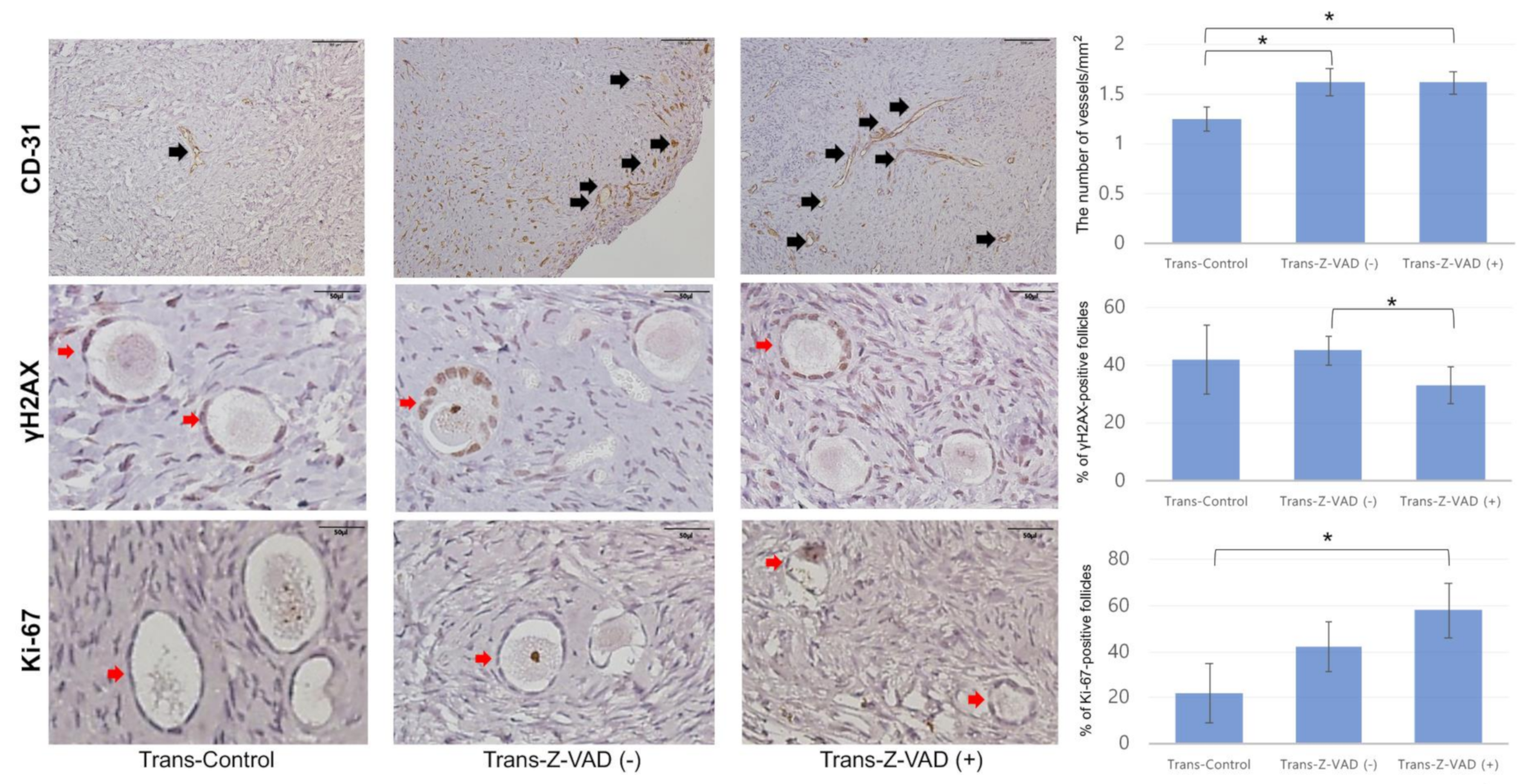
2.2. Follicular Cell Proliferation, Double-Strand DNA Damage, and Angiogenesis
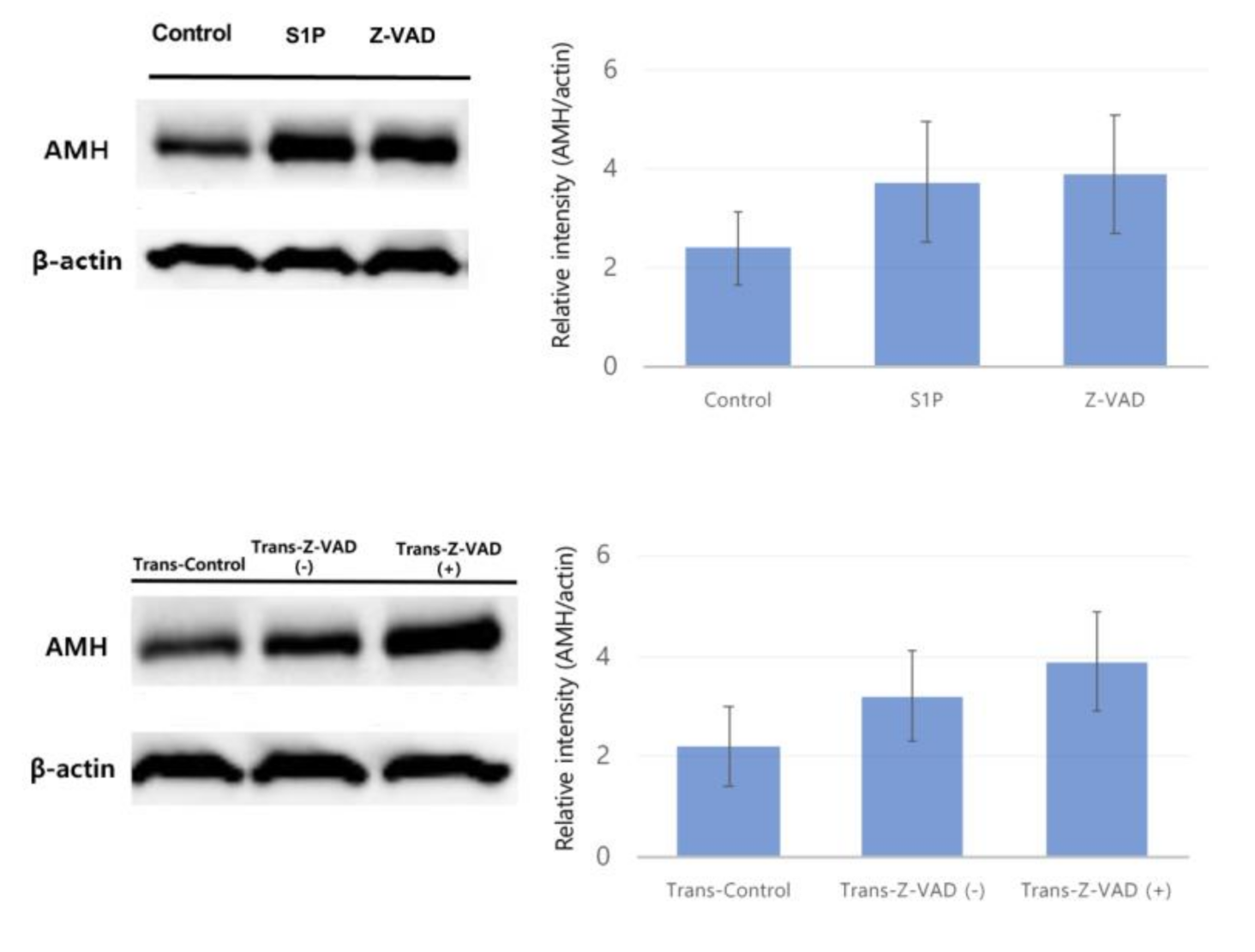
2.3. Western Blot
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Anti-Apoptotic Agents
4.3. Slow-Freezing Protocol
4.4. Slow Freezing–Thawing Protocol
4.5. Xenotransplantation into SCID Mice
4.6. Histologic Evaluation
4.7. Immunohistochemistry (IHC) Evaluation
4.8. Western Blot
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoekman, E.J.; Knoester, D.; Peters, A.A.W.; Jansen, F.W.; De Kroon, C.D.; Hilders, C.G.J.M. Ovarian survival after pelvic radiation: Transposition until the age of 35 years. Arch. Gynecol. Obstet. 2018, 298, 1001–1007. [Google Scholar] [CrossRef] [Green Version]
- Azim, A.A.; Costantini-Ferrando, M.; Oktay, K. Safety of Fertility Preservation by Ovarian Stimulation With Letrozole and Gonadotropins in Patients With Breast Cancer: A Prospective Controlled Study. J. Clin. Oncol. 2008, 26, 2630–2635. [Google Scholar] [CrossRef]
- Fertility preservation in patients undergoing gonadotoxic therapy or gonadectomy: A committee opinion. Fertil. Steril. 2019, 112, 1022–1033. [CrossRef]
- Demeestere, I.; Simon, P.; Dedeken, L.; Moffa, F.; Tsépélidis, S.; Brachet, C.; Delbaere, A.; Devreker, F.; Ferster, A. Live birth after autograft of ovarian tissue cryopreserved during childhood: Figure 1. Hum. Reprod. 2015, 30, 2107–2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salama, M.; Isachenko, V.; Isachenko, E.; Rahimi, G.; Mallmann, P. Updates in preserving reproductive potential of prepubertal girls with cancer: Systematic review. Crit. Rev. Oncol. 2016, 103, 10–21. [Google Scholar] [CrossRef]
- Donnez, J.; Dolmans, M.-M. Cryopreservation and Transplantation of Ovarian Tissue. Clin. Obstet. Gynecol. 2010, 53, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Grynberg, M.; Poulain, M.; Sebag-Peyrelevade, S.; Le Parco, S.; Fanchin, R.; Frydman, N.A. Ovarian tissue and follicle transplantation as an option for fertility preservation. Fertil. Steril. 2012, 97, 1260–1268. [Google Scholar] [CrossRef] [PubMed]
- Practice Committee of American Society for Reproductive Medicine Ovarian tissue cryopreservation: A committee opinion. Fertil. Steril. 2014, 101, 1237–1243. [CrossRef] [PubMed]
- Demeestere, I.; Simon, P.; Emiliani, S.; Delbaere, A.; Englert, Y. Orthotopic and heterotopic ovarian tissue transplantation. Hum. Reprod. Updat. 2009, 15, 649–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Eyck, A.-S.; Jordan, B.F.; Gallez, B.; Heilier, J.-F.; Van Langendonckt, A.; Donnez, J. Electron paramagnetic resonance as a tool to evaluate human ovarian tissue reoxygenation after xenografting. Fertil. Steril. 2009, 92, 374–381. [Google Scholar] [CrossRef]
- Van Eyck, A.-S.; Bouzin, C.; Feron, O.; Romeu, L.; Van Langendonckt, A.; Donnez, J.; Dolmans, M.-M. Both host and graft vessels contribute to revascularization of xenografted human ovarian tissue in a murine model. Fertil. Steril. 2010, 93, 1676–1685. [Google Scholar] [CrossRef]
- Soleimani, R.; Heytens, E.; Oktay, K. Enhancement of Neoangiogenesis and Follicle Survival by Sphingosine-1-Phosphate in Human Ovarian Tissue Xenotransplants. PLoS ONE 2011, 6, e19475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.-C.; Tzeng, C.-R.; Wang, C.-W.; Hsu, M.-I.; Tan, S.-J.; Chen, C.-H. Antiapoptotic Agent Sphingosine-1-Phosphate Protects Vitrified Murine Ovarian Grafts. Reprod. Sci. 2013, 21, 236–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzel, Y.; Bildik, G.; Oktem, O. Sphingosine-1-phosphate protects human ovarian follicles from apoptosis in vitro. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 222, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Fransolet, M.; Noël, L.; Henry, L.; Labied, S.; Blacher, S.; Nisolle, M.; Munaut, C. Evaluation of Z-VAD-FMK as an anti-apoptotic drug to prevent granulosa cell apoptosis and follicular death after human ovarian tissue transplantation. J. Assist. Reprod. Genet. 2019, 36, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Rimon, E.; Cohen, T.; Dantes, A.; Hirsh, L.; Amit, A.; Lessing, J.B.; Freimann, S.; Amsterdam, A.; Azem, F. Apoptosis in cryopreserved human ovarian tissue obtained from cancer patients: A tool for evaluating cryopreservation utility. Int. J. Oncol. 2005, 27, 345–353. [Google Scholar] [CrossRef]
- Fauque, P.; Ben Amor, A.; Joanne, C.; Agnani, G.; Bresson, J.L.; Roux, C. Use of trypan blue staining to assess the quality of ovarian cryopreservation. Fertil. Steril. 2007, 87, 1200–1207. [Google Scholar] [CrossRef]
- Xiao, Z.; Wang, Y.; Li, L.; Li, S.-W. Cryopreservation of the human ovarian tissue induces the expression of Fas system in morphologically normal primordial follicles. Cryo Lett. 2010, 31, 112–119. [Google Scholar]
- Liu, J.; Van Der Elst, J.; Broecke, R.V.D.; Dhont, M. Early massive follicle loss and apoptosis in heterotopically grafted newborn mouse ovaries. Hum. Reprod. 2002, 17, 605–611. [Google Scholar] [CrossRef]
- Yang, H.; Lee, H.H.; Lee, H.C.; Ko, D.S.; Kim, S.S. Assessment of vascular endothelial growth factor expression and apoptosis in the ovarian graft: Can exogenous gonadotropin promote angiogenesis after ovarian transplantation? Fertil. Steril. 2008, 90, 1550–1558. [Google Scholar] [CrossRef]
- Baird, D.T.; Webb, R.; Campbell, B.K.; Harkness, L.M.; Gosden, R.G. Long-Term Ovarian Function in Sheep after Ovariectomy and Transplantation of Autografts Stored at −196 C**This work was supported by Medical Research Council Program Grant 8929853. Endocrinology 1999, 140, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Cuvillier, O.; Pirianov, G.; Kleuser, B.; Vanek, P.G.; Coso, O.A.; Gutkind, J.S.; Spiegel, S. Suppression of ceramide-mediated programmed cell death by sphingosine-1-phosphate. Nat. Cell Biol. 1996, 381, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, S.; Kolesnick, R. Sphingosine 1-phosphate as a therapeutic agent. Leukemia 2002, 16, 1596–1602. [Google Scholar] [CrossRef]
- Morita, Y.; Perez, G.I.; Paris, F.; Miranda, S.R.; Ehleiter, D.; Haimovitz-Friedman, A.; Fuks, Z.; Xie, Z.; Reed, J.C.; Schuchman, E.H.; et al. Oocyte apoptosis is suppressed by disruption of the acid sphingomyelinase gene or by sphingosine -1-phosphate therapy. Nat. Med. 2000, 6, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Siess, W. Athero- and thrombogenic actions of lysophosphatidic acid and sphingosine-1-phosphate. Biochim. et Biophys. Acta (BBA) - Bioenerg. 2002, 1582, 204–215. [Google Scholar] [CrossRef]
- Roness, H.; Kashi, O.; Meirow, D. Prevention of chemotherapy-induced ovarian damage. Fertil. Steril. 2016, 105, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.R. Apoptosis in the ovary: Molecular mechanisms. Hum. Reprod. Updat. 2005, 11, 162–178. [Google Scholar] [CrossRef]
- Zhang, J.-M.; Li, L.-X.; Yang, Y.-X.; Liu, X.-L.; Wan, X.-P. Is caspase inhibition a valid therapeutic strategy in cryopreservation of ovarian tissue? J. Assist. Reprod. Genet. 2009, 26, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Pepper, A.R.; Bruni, A.; Pawlick, R.; Wink, J.; Rafiei, Y.; Gala-Lopez, B.; Bral, M.; Abualhassan, N.; Kin, T.; Shapiro, A.M.J. Engraftment Site and Effectiveness of the Pan-Caspase Inhibitor F573 to Improve Engraftment in Mouse and Human Islet Transplantation in Mice. Transplantation 2017, 101, 2321–2329. [Google Scholar] [CrossRef]
- Hancke, K.; Strauch, O.; Kissel, C.; Göbel, H.; Schäfer, W.; Denschlag, D. Sphingosine 1-phosphate protects ovaries from chemotherapy-induced damage in vivo. Fertil. Steril. 2007, 87, 172–177. [Google Scholar] [CrossRef]
- Vogler, R.; Sauer, B.; Kim, D.-S.; Schäfer-Korting, M.; Kleuser, B.; Sch, M. Sphingosine-1-Phosphate and Its Potentially Paradoxical Effects on Critical Parameters of Cutaneous Wound Healing. J. Investig. Dermatol. 2003, 120, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of Substances to Laboratory Animals: Routes of Administration and Factors to Consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar] [PubMed]
- Oktay, K.; Economos, K.; Kan, M.; Rucinski, J.; Veeck, L.; Rosenwaks, Z. Endocrine function and oocyte retrieval after autologous transplantation of ovarian cortical strips to the forearm. JAMA 2001, 286, 1490. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Ryu, K.-J.; Kim, B.; Kang, D.; Kim, Y.Y.; Kim, T. Comparison between Slow Freezing and Vitrification for Human Ovarian Tissue Cryopreservation and Xenotransplantation. Int. J. Mol. Sci. 2019, 20, 3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oktay, K.; Karlikaya, G.G.; A Aydin, B. Ovarian cryopreservation and transplantation: Basic aspects. Mol. Cell. Endocrinol. 2000, 169, 105–108. [Google Scholar] [CrossRef]
- Kim, B.; Ryu, K.-J.; Lee, S.; Kim, T. Changes in telomere length and senescence markers during human ovarian tissue cryopreservation. Sci. Rep. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Moretti, L.; Kim, K.W.; Jung, D.K.; Willey, C.D.; Lu, B. Radiosensitization of solid tumors by Z-VAD, a pan-caspase inhibitor. Mol. Cancer Ther. 2009, 8, 1270–1279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlüter, A.; Weller, P.; Kanaan, O.; Nel, I.; Heusgen, L.; Höing, B.; Haßkamp, P.; Zander, S.; Mandapathil, M.; Dominas, N.; et al. CD31 and VEGF are prognostic biomarkers in early-stage, but not in late-stage, laryngeal squamous cell carcinoma. BMC Cancer 2018, 18, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Cho, H.-W.; Kim, B.; Lee, J.K.; Kim, T. The Effectiveness of Anti-Apoptotic Agents to Preserve Primordial Follicles and Prevent Tissue Damage during Ovarian Tissue Cryopreservation and Xenotransplantation. Int. J. Mol. Sci. 2021, 22, 2534. https://doi.org/10.3390/ijms22052534
Lee S, Cho H-W, Kim B, Lee JK, Kim T. The Effectiveness of Anti-Apoptotic Agents to Preserve Primordial Follicles and Prevent Tissue Damage during Ovarian Tissue Cryopreservation and Xenotransplantation. International Journal of Molecular Sciences. 2021; 22(5):2534. https://doi.org/10.3390/ijms22052534
Chicago/Turabian StyleLee, Sanghoon, Hyun-Woong Cho, Boram Kim, Jae Kwan Lee, and Tak Kim. 2021. "The Effectiveness of Anti-Apoptotic Agents to Preserve Primordial Follicles and Prevent Tissue Damage during Ovarian Tissue Cryopreservation and Xenotransplantation" International Journal of Molecular Sciences 22, no. 5: 2534. https://doi.org/10.3390/ijms22052534