Increased Pro-Thrombotic Platelet Activity Associated with Thrombin/PAR1-Dependent Pathway Disorder in Patients with Secondary Progressive Multiple Sclerosis

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
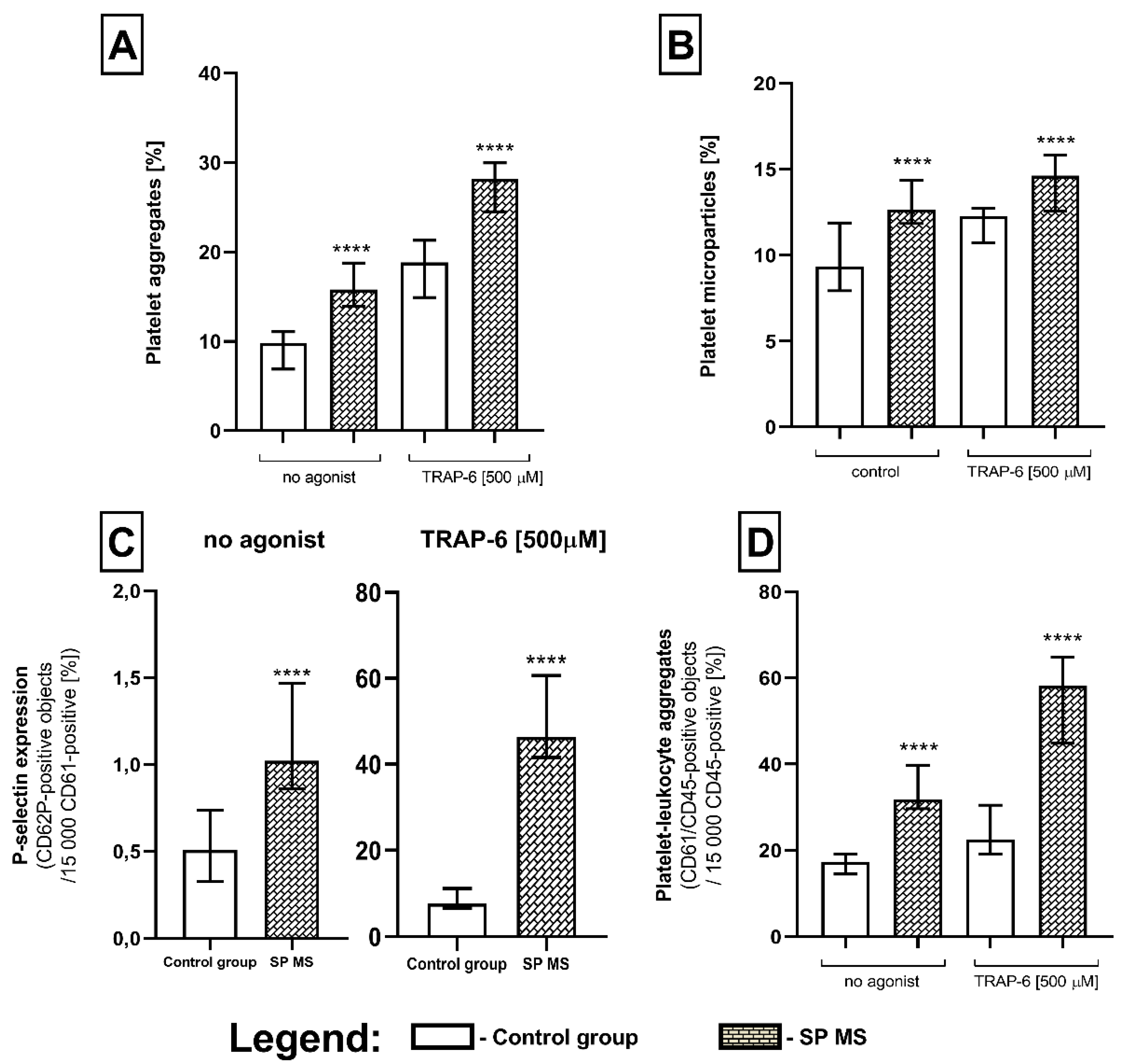
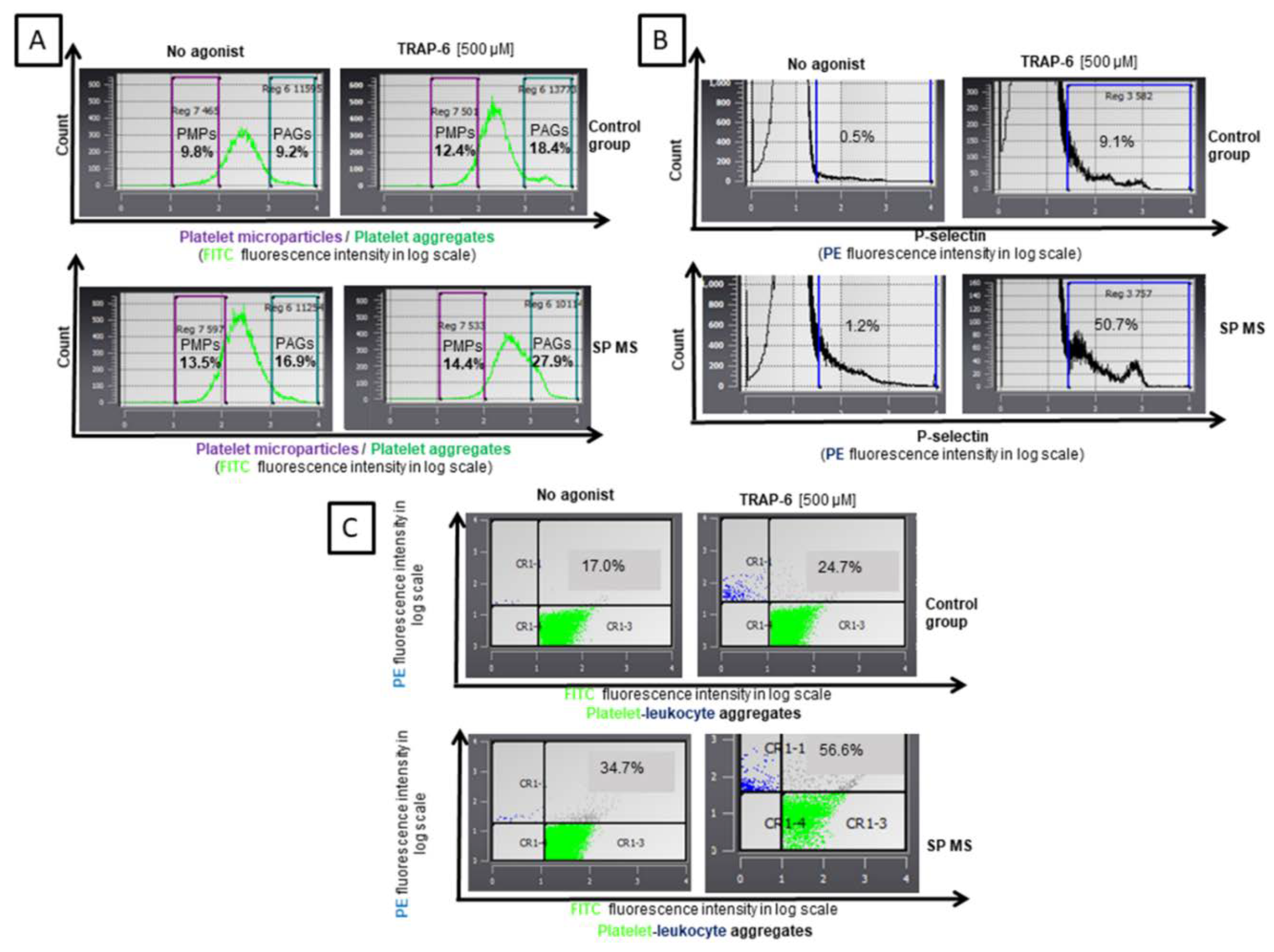
2.1. The Level of Blood Platelet’s Markers of Activation
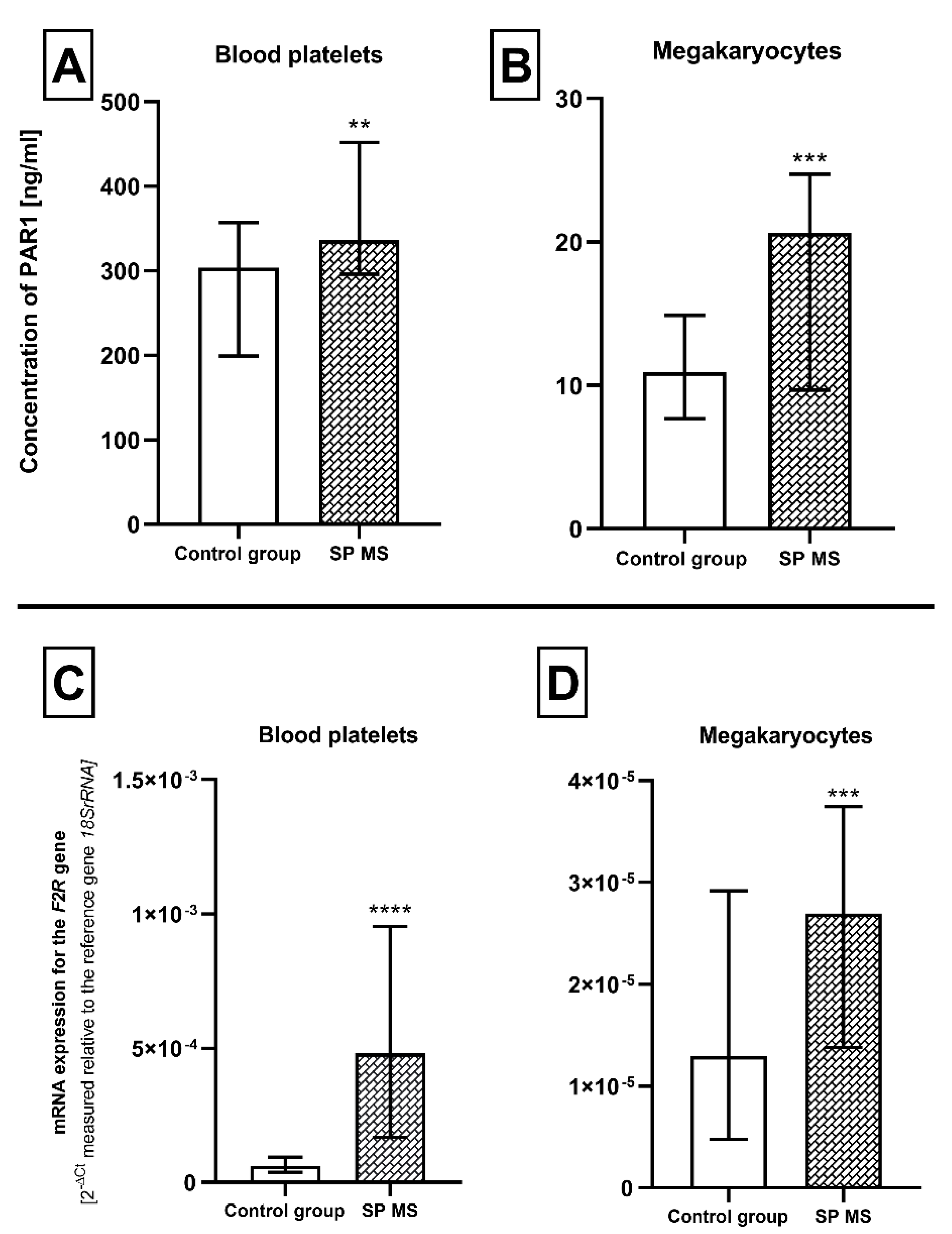
2.2. The Expression for PAR1 at Protein and mRNA Level in Blood Platelets and Megakaryocytes
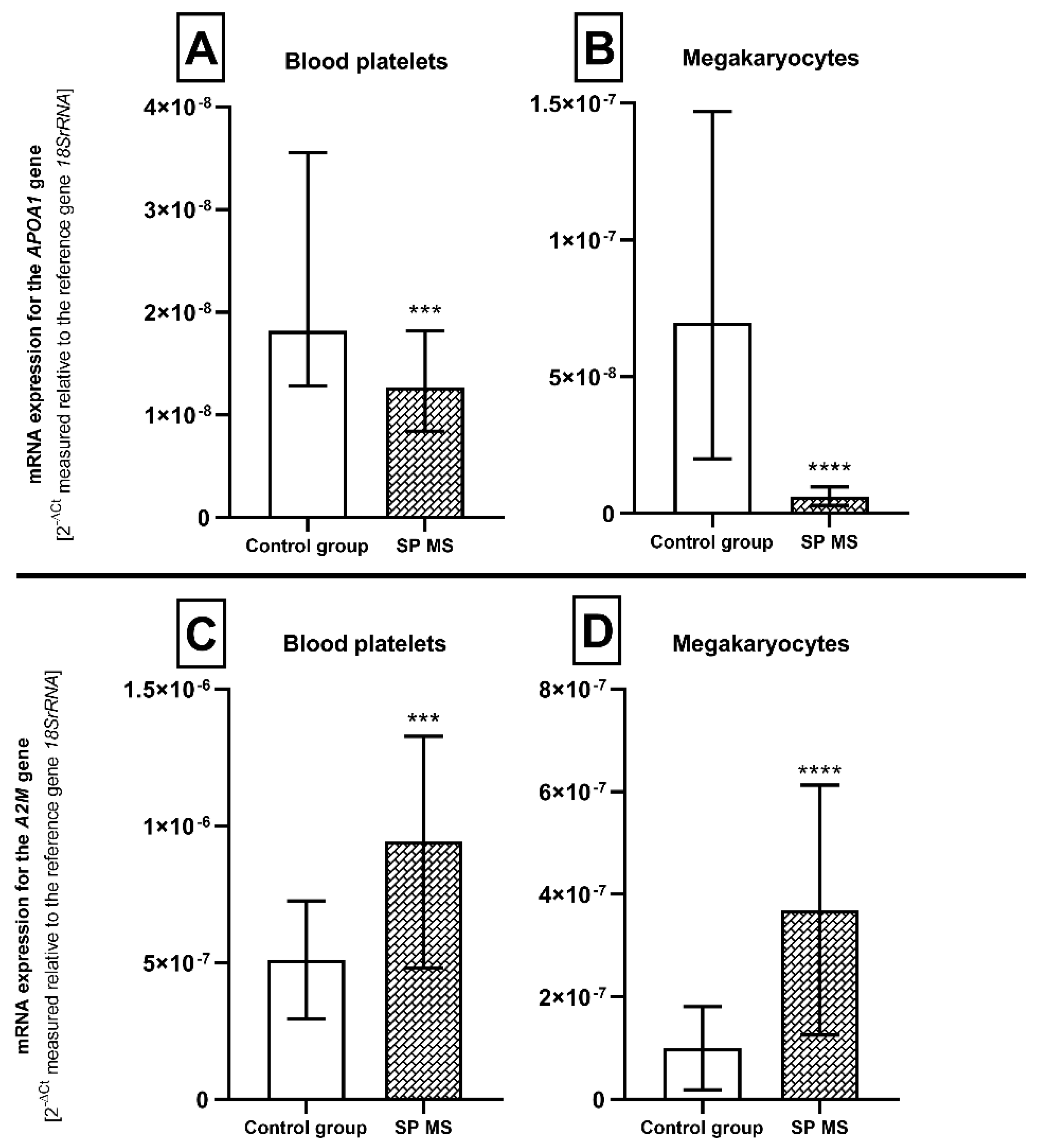
2.3. The Relative mRNA Expression of Genes Encoding Atherosclerosis Biomarkers—APOA1 and A2M in Blood Platelets and Megakaryocytes
3. Discussion
4. Materials and Methods
4.1. Clinical Characteristics of MS Patients and Control Group
4.2. Cytometric Measurement of Characteristic Markers of Platelet Activation
4.3. Isolation of Blood Platelets
4.4. Isolation of Megakaryocytes
4.5. Isolation of mRNA and cDNA Synthesis
4.6. Expression of the F2R, APOA1 and A2M Genes at the mRNA Level in Blood Platelets and Megakaryocytes
4.7. Concentration of PAR1 Receptor in Blood Platelets and Megakaryocytes
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 2-DE | 2-dimensional electrophoresis |
| ADP | adenosine diphosphate |
| ApoA1 | apolipoprotein A1 |
| BBB | blood-brain barrier |
| BDI | Beck’s depression inventory |
| BMI | body mass index |
| CIMT | carotid intima-media thickness |
| CIS | clinically isolated syndrome |
| CNS | central nervous system |
| CPDA-1 | citrate phosphate dextrose adenine-1 |
| CRP | C-reactive protein |
| CSF | cerebrospinal fluid |
| CVD | cardiovascular disease |
| EDSS | expanded disability status scale |
| GPCRs | G-protein-coupled protease-activated receptors |
| GPIIb/IIIa | glycoprotein IIb/IIIa |
| HDL | high-density lipoprotein |
| IgM | immunoglobulin M |
| IL-6 | interleukin-6 |
| IQR | interquartile range |
| LDL | low-density lipoprotein |
| MALDI-TOF/TOF | matrix-assisted laser desorption/ionization time-of-flight/time-of-flight |
| miRNA | microRNA |
| MMP | matrix metalloproteinase |
| MS | multiple sclerosis |
| PAGs | platelet aggregates |
| PAP | plasmin–anti-plasmin |
| PAR1 | protease-activated receptor 1 |
| PBMCs | peripheral blood mononuclear cells |
| PF4 | platelet factor 4 |
| PLAs | platelet-leukocyte aggregates |
| PMPs | platelet microparticles |
| PP MS | primary progressive multiple sclerosis |
| PRP | platelet-rich plasma |
| PSGL-1 | P-selectin glycoprotein ligand 1 |
| Rho | Spearman’s rank correlation |
| RR MS | relapsing-remitting multiple sclerosis |
| RT-qPCR | quantitative Real-Time polymerase chain reaction |
| SP MS | secondary progressive multiple sclerosis |
| TAT | thrombin–anti-thrombin |
| TNF-α | tumor necrosis factor α |
| TRAP-6 | thrombin receptor-activating peptide 6 |
| vWF | von Willebrand factor |
| α2M | α-2-macroglobulin |
| β-TG | β-thromboglobulin |
References
- Lassmann, H.; van Horssen, J. The Molecular Basis of Neurodegeneration in Multiple Sclerosis. FEBS Lett. 2011, 585, 3715–3723. [Google Scholar] [CrossRef] [Green Version]
- D’Haeseleer, M.; Cambron, M.; Vanopdenbosch, L.; De Keyser, J. Vascular aspects of multiple sclerosis. Lancet Neurol. 2011, 10, 657–666. [Google Scholar] [CrossRef]
- Lublin, F.D. New multiple sclerosis phenotypic classification. Eur. Neurol. 2014, 72 (Suppl. 1), 1–5. [Google Scholar] [CrossRef]
- Christiansen, C.F.; Christensen, S.; Farkas, D.K.; Miret, M.; Sørensen, H.T.; Pedersen, L. Risk of arterial cardiovascular diseases in patients with multiple sclerosis: A population-based cohort study. Neuroepidemiology 2010, 35, 267–274. [Google Scholar] [CrossRef]
- Jadidi, E.; Mohammadi, M.; Moradi, T. High risk of cardiovascular diseases after diagnosis of multiple sclerosis. Mult. Scler. J. 2013, 19, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Bronnum-Hansen, H.; Koch-Henriksen, N.; Stenager, E. Trends in survival and cause of death in Danish patients with multiple sclerosis. Brain 2004, 127 Pt 4, 844–850. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.; Farkas, D.K.; Pedersen, L.; Miret, M.; Christiansen, C.F.; Sørensen, H.T. Multiple sclerosis and risk of venous thromboembolism: A population-based cohort study. Neuroepidemiology 2012, 38, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Peeters, P.J.H.L.; Bazelier, M.T.; Uitdehaag, B.M.J.; Leufkens, H.G.M.; De Bruin, M.L.; de Vries, F. The risk of venous thromboembolism in patients with multiple sclerosis: The Clinical Practice Research Datalink. J. Thromb. Haemost. 2014, 12, 444–451. [Google Scholar] [CrossRef]
- Tseng, C.H.; Huang, W.S.; Lin, C.L.; Chang, Y.J. Increased risk of ischaemic stroke among patients with multiple sclerosis. Eur. J. Neurol. 2015, 22, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Persson, R.; Lee, S.; Yood, M.U.; Wagner, M.; Minton, N.; Niemcryk, S.; Lindholm, A.; Evans, A.; Jick, S. Incident cardiovascular disease in patients diagnosed with multiple sclerosis: A multi-database study. Mult. Scler. Relat. Disord. 2019, 37, 101423. [Google Scholar] [CrossRef]
- Hong, Y.; Tang, H.R.; Ma, M.; Chen, N.; Xie, X.; He, L. Multiple sclerosis and stroke: A systematic review and meta-analysis. BMC Neurol. 2019, 19, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puthenparampil, M.; Rinaldi, F.; Federle, L.; Cazzola, C.; Perini, P.; Gallo, P. Decreased platelet number in multiple sclerosis during alemtuzumab infusion: A common, transient and clinically silent phenomenon. Ther. Adv. Neurol. Disord. 2017, 11, 1756285617741056. [Google Scholar] [CrossRef] [PubMed]
- Callea, L.; Arese, M.; Orlandini, A.; Bargnani, C.; Priori, A.; Bussolino, F. Platelet activating factor is elevated in cerebral spinal fluid and plasma of patients with relapsing-remitting multiple sclerosis. J. Neuroimmunol. 1999, 94, 212–221. [Google Scholar] [CrossRef]
- Kuenz, B.; Lutterotti, A.; Khali, M.; Ehling, R.; Gneiss, C.; Deisenhammer, F.; Reindl, M.; Berger, T. Plasma levels of soluble adhesion molecules sPECAM-1, sP-selectin and sE-selectin are associated with relapsing-remitting disease course of multiple sclerosis. J. Neuroimmunol. 2005, 167, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Dziedzic, A.; Morel, A.; Miller, E.; Bijak, M.; Sliwinski, T.; Synowiec, E.; Ceremuga, M.; Saluk-Bijak, J. Oxidative Damage of Blood Platelets Correlates with the Degree of Psychophysical Disability in Secondary Progressive Multiple Sclerosis. Oxidative Med. Cell. Longev. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Morel, A.; Rywaniak, J.; Bijak, M.; Miller, E.; Niwald, M.; Saluk, J. Flow cytometric analysis reveals the high levels of platelet activation parameters in circulation of multiple sclerosis patients. Mol. Cell. Biochem. 2017, 430, 69–80. [Google Scholar] [CrossRef] [Green Version]
- Morel, A.; Bijak, M.; Miller, E.; Rywaniak, J.; Miller, S.; Saluk, J. Relationship between the Increased Haemostatic Properties of Blood Platelets and Oxidative Stress Level in Multiple Sclerosis Patients with the Secondary Progressive Stage. Oxidative Med. Cell. Longev. 2015, 2015, 240918. [Google Scholar] [CrossRef] [Green Version]
- Bijak, M.; Olejnik, A.; Rokita, B.; Morel, A.; Dziedzic, A.; Miller, E.; Saluk-Bijak, J. Increased level of fibrinogen chains in the proteome of blood platelets in secondary progressive multiple sclerosis patients. J. Cell. Mol. Med. 2019, 23, 3476–3482. [Google Scholar] [CrossRef] [Green Version]
- Duvernay, M.; Young, S.; Gailani, D.; Schoenecker, J.; Hamm, H. Protease-activated receptor (PAR) 1 and PAR4 differentially regulate factor V expression from human platelets. Mol. Pharmacol. 2013, 83, 781–792. [Google Scholar] [CrossRef] [Green Version]
- Heuberger, D.M.; Schuepbach, R.A. Protease-activated receptors (PARs): Mechanisms of action and potential therapeutic modulators in PAR-driven inflammatory diseases. Thromb. J. 2019, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Kahn, M.L.; Nakanishi-Matsui, M.; Shapiro, M.J.; Ishihara, H.; Coughlin, S.R. Protease-activated receptors 1 and 4 mediate activation of human platelets by thrombin. J. Clin. Investig. 1999, 103, 879–887. [Google Scholar] [CrossRef] [Green Version]
- McNicol, A.; Gerrard, J.M. Post-receptor events associated with thrombin-induced platelet activation. Blood Coagul. Fibrinolysis 1993, 4, 975–991. [Google Scholar] [CrossRef]
- De Luca, C.; Vurtuoso, A.; Maggio, N.; Papa, M. Neuro-Coagulopathy: Blood Coagulation Factors in Central Nervous System Diseases. Int. J. Mol. Sci. 2017, 18, 2128. [Google Scholar] [CrossRef]
- Schoergenhofer, C.; Schwameis, M.; Gelbengger, G.; Buchtele, N.; Thaler, B.; Mussbacher, M.; Schabbauer, G.; Wojta, J.; Jilma-Stohlawets, P.; Jilma, B. Inhibition of Protease-Activated Receptor (PAR1) Reduces Activation of the Endothelium, Coagulation, Fibrinolysis and Inflammation during Human Endotoxemia. Thromb. Haemost. 2018, 118, 1176–1184. [Google Scholar] [CrossRef]
- Kent, S.; Hough, S.; Kelleher, A.D.; Law, M.G. Vorapaxar for HIV-associated inflammation and coagulopathy (ADVICE): A randomised, double-blind, placebo-controlled trial. Lancet HIV 2018, 5, e553–e559. [Google Scholar] [CrossRef]
- O’Donoghue, M.L.; Bhatt, D.L.; Flather, M.D.; Goto, S.; Angiolillo, D.J.; Goodman, S.G.; Zeymer, U.; Aylward, P.E.; Montalescot, G.; Ziecina, R.; et al. Atopaxar and its effects on markers of platelet activation and inflammation: Results from the LANCELOT CAD program. J. Thromb. Thrombolysis 2012, 34, 36–43. [Google Scholar] [CrossRef]
- Valavanis, A.; Tsitsipa, E.; Intzes, S.; Psoma, E.; Tegos, T. The impact of athrosclerosis on cognition and disability in multiple sclerosis patients: The ATHUS score. Hippokratia 2019, 23, 81–86. [Google Scholar]
- Florvall, G.; Basu, S.; Larsson, A. Apolipoprotein A1 Is a Stronger Prognostic Marker Than Are HDL and LDL Cholesterol for Cardiovascular Disease and Mortality in Elderly Men. J. Gerontol. Ser. A 2006, 61, 1262–1266. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, R.; Nezu, T.; Hosomi, N.; Aoki, S.; Sugimoto, T.; Kinoshita, N.; Ataki, M.; Takahashi, T.; Muruyama, T.; Matsumoto, M. Alpha-2-macroglobulin as a Promising Biological Marker of Endothelial Function. J. Atheroscler. Thromb. 2017, 25, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.A.; Wuescher, L.M.; Worth, R.G. Platelets: Essential components of the immune system. Curr. Trends Immunol. 2015, 16, 65–78. [Google Scholar]
- Yun, S.-H.; Sim, E.-H.; Goh, R.-Y.; Park, J.-I.; Han, J.-Y. Platelet Activation: The Mechanisms and Potential Biomarkers. BioMed Res. Int. 2016, 2016, 9060143. [Google Scholar] [CrossRef] [Green Version]
- Hoyer, F.F.; Nickenig, G.; Werner, N. Microparticles—Messengers of biological information. J. Cell. Mol. Med. 2010, 14, 2250–2256. [Google Scholar] [CrossRef]
- Weber, M.S.; Prod’homme, T.; Youssef, S.; Dunn, S.E.; Rundle, C.D.; Lee, L.; Patarroyo, J.C.; Stuve, O.; Sobel, R.A.; Steinman, L. Type II monocytes modulate T cell-mediated central nervous system autoimmune disease. Nat. Med. 2007, 13, 935–943. [Google Scholar] [CrossRef] [PubMed]
- Heppner, F.L.; Greter, M.; Marino, D.; Falsig, J.; Raivich, G.; Hovelmeyer, N.; Waisman, A.; Rulicke, T.; Prinz, M.; Priller, J.; et al. Experimental autoimmune encephalomyelitis repressed by microglial paralysis. Nat. Med. 2005, 11, 146–152. [Google Scholar] [CrossRef]
- Sheremata, W.A.; Jy, W.; Horstman, L.L.; Ahn, Y.S.; Alexander, J.S.; Minagar, A. Evidence of platelet activation in multiple sclerosis. J. Neuroinflammation 2008, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Han, M.H.; Hwang, S.-I.; Roy, D.B.; Lundgren, D.H.; Price, J.V.; Ousman, S.S.; Fernald, G.H.; Gerlitz, B.; Robinson, W.H.; Baranzini, S.E.; et al. Proteomic analysis of active multiple sclerosis lesions reveals therapeutic targets. Nature 2008, 451, 1076–1081. [Google Scholar] [CrossRef]
- Langer, H.F.; Choi, E.-Y.; Zhou, H.; Schleicher, R.; Chung, K.-J.; Tang, Z.; Gobel, K.; Bdeir, K.; Chatzigeorgiou, A.; Wong, C.; et al. Platelets contribute to the pathogenesis of experimental autoimmune encephalomyelitis. Circ. Res. 2012, 110, 1202–1210. [Google Scholar] [CrossRef] [Green Version]
- McMichael, M. New models of hemostasis. Top. Companion Anim. Med. 2012, 27, 40–45. [Google Scholar] [CrossRef]
- Bijak, M.; Gajak, A.; Nowak, P. Hemostatic and cellular functions of factor XIII. Pol. Merkur. Lek. 2013, 34, 71–74. [Google Scholar]
- Hoffman, M.; Monroe, D.M. Coagulation 2006: A modern view of hemostasis. Hematol./Oncol. Clin. North Am. 2007, 21, 1–11. [Google Scholar] [CrossRef]
- Andersen, H.; Greenberg, D.L.; Fujikawa, K.; Xu, W.; Chung, D.W.; Davie, E.W. Protease-activated receptor 1 is the primary mediator of thrombin-stimulated platelet procoagulant activity. Proc. Natl. Acad. Sci. USA 1999, 96, 11189. [Google Scholar] [CrossRef] [Green Version]
- Ofosu, F. Review: Laboratory markers quantifying prothrombin activation and actions of thrombin in venous and arterial thrombosis do not accurately assess disease severity or the effectiveness of treatment. Thromb. Haemost. 2006, 96, 568–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palladino, R.; Marrie, R.A.; Majeed, A.; Chataway, J. Evaluating the Risk of Macrovascular Events and Mortality among People with Multiple Sclerosis in England. JAMA Neurol. 2020, 77, 1–9. [Google Scholar] [CrossRef]
- Roshanisefat, H.; Bahmanyar, S.; Hillert, J.; Olsson, T.; Montgomery, S. Multiple sclerosis clinical course and cardiovascular disease risk—Swedish cohort study. Eur. J. Neurol. 2014, 21, 1353-e88. [Google Scholar] [CrossRef]
- Wei, L.; Mac Donald, T.M.; Walker, B.R. Taking glucocorticoids by prescription is associated with subsequent cardiovascular disease. Ann. Intern. Med. 2004, 141, 764–770. [Google Scholar] [CrossRef]
- Sonnenblick, M.; Rosin, A. Cardiotoxicity of interferon. A review of 44 cases. Chest 1991, 99, 557–561. [Google Scholar] [CrossRef] [Green Version]
- Hussein, W.I.; Reddy, S.S. Prevalence of diabetes in patients with multiple sclerosis. Diabetes Care 2006, 29, 1984–1985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, Hypertension, and Cardiovascular Disease: Clinical Insights and Vascular Mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef] [Green Version]
- Hargett, L.A.; Bauer, N.N. On the origin of microparticles: From “platelet dust” to mediators of intercellular communication. Pulm. Circ. 2013, 3, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Dye, J.R.; Ullal, A.J.; Pisetsky, D.S. The role of microparticles in the pathogenesis of rheumatoid arthritis and systemic lupus erythematosus. Scand. J. Immunol. 2013, 78, 140–148. [Google Scholar] [CrossRef]
- Sellam, J.; Proulle, V.; Jungel, A.; Ittah, M.; Richard, C.M.; Gottenberg, J.-E.; Toi, F.; Benssiano, J.; Gay, S.; Freyssinet, J.-M.; et al. Increased levels of circulating microparticles in primary Sjögren’s syndrome, systemic lupus erythematosus and rheumatoid arthritis and relation with disease activity. Arthritis Res. Ther. 2009, 11, R156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowert, N.S.; Donner, L.; Chattarjee, M.; Eisele, Y.S.; Towhid, S.t.; Munzer, P.; Walker, B.; Ogorek, I.; Borst, O.; Grandoch, M. Blood platelets in the progression of Alzheimer’s disease. PLoS ONE 2014, 9, e90523. [Google Scholar] [CrossRef] [Green Version]
- Peerschke, E.I.; Yin, W.; Ghebrehiwet, B. Complement activation on platelets: Implications for vascular inflammation and thrombosis. Mol. Immunol. 2010, 47, 2170–2175. [Google Scholar] [CrossRef] [Green Version]
- Waubant, E. Biomarkers indicative of blood-brain barrier disruption in multiple sclerosis. Dis. Markers 2006, 22, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Flaumenhaft, R.; Mairuhu, A.T.; Italiano, J.E. Platelet- and megakaryocyte-derived microparticles. Semin. Thromb. Hemost. 2010, 36, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Frenette, P.S.; Denis, C.V.; Weiss, L.; Jurk, K.; Subbarao, S.; Kehrel, B.; Hartwig, J.H.; Vestweber, D.; Wagner, D.D. P-Selectin glycoprotein ligand 1 (PSGL-1) is expressed on platelets and can mediate platelet-endothelial interactions in vivo. J. Exp. Med. 2000, 191, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; de la Motte, C.; Reyes, B.M.R.; Sans, M.; Levine, A.D.; Fiocchi, C. Cutting edge: T cells trigger CD40-dependent platelet activation and granular RANTES release: A novel pathway for immune response amplification. J. Immunol. 2004, 172, 2011–2015. [Google Scholar] [CrossRef] [Green Version]
- Ziliotto, N.; Bernardi, F.; Jakimovski, D.; Zivadinov, R. Coagulation Pathways in Neurological Diseases: Multiple Sclerosis. Front. Neurol. 2019, 10, 409. [Google Scholar] [CrossRef]
- Bijak, M.; Saluk, J.; Ponczek, M.B.; Nowak, P.; Wachowicz, B. The synthesis of proteins in unnucleated blood platelets. Postepy Hig. Med. Dosw. (Online) 2013, 67, 672–679. [Google Scholar] [CrossRef]
- Schubert, P.; Devine, D.V. De novo protein synthesis in mature platelets: A consideration for transfusion medicine. Vox Sang. 2010, 99, 112–122. [Google Scholar] [CrossRef]
- Harrison, P.; Savidge, G.F.; Cramer, E.M. The origin and physiological relevance of alpha-granule adhesive proteins. Br. J. Haematol. 1990, 74, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Weyrich, A.S.; Schwertz, H.; Kraiss, L.W.; Zimmerman, G.A. Protein synthesis by platelets: Historical and new perspectives. J. Thromb. Haemost. 2009, 7, 241–246. [Google Scholar] [CrossRef] [Green Version]
- Yuksel, B.; Koc, P.; Goksu, E.O.; Karacay, E.; Kurtulus, F.; Cekin, Y.; Gamceli, Y.B. Is multiple sclerosis a risk factor for atherosclerosis? J. Neuroradiol. 2019. [CrossRef]
- Mincu, R.I.; Magda, L.S.; Florescu, M.; Velcea, A.; Mihaila, S.; Popescu, B.O.; Chiru, A.; Tiu, C.; Ciinteza, M.; Vinereanu, D.; et al. Cardiovascular Dysfunction in Multiple Sclerosis. Maedica 2015, 10, 364–370. [Google Scholar]
- Manfroi, W.; Zago, A.J.; Campos, M.; Alves, A.; Brisolara, L.; de Souza, J.; Candigo, R.H.; Krischnick, L.; Ribeiro, L.; Ordovas, K.; et al. Are apolipoproteins A and B better than lipoproteins for assessing risk of obstructive coronary heart disease? Arq. Bras. Cardiol. 1999, 72, 657–668. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, T.; Nguyen, T.T.; Hallaway, B.J.; Hodge, D.; Bailey, K.; Holmes, D.; Kottke, B.A. The Role of Lipoprotein A-I and Lipoprotein A-I/A-II in Predicting Coronary Artery Disease. Arterioscler. Thromb. Vasc. Biol. 1995, 15, 228–231. [Google Scholar] [CrossRef] [PubMed]
- Mucchiano, G.I.; Jonasson, J.; Haggqvist, B.; Einarsson, E.; Westermark, P. Apolipoprotein A-I-derived amyloid in atherosclerosis. Its association with plasma levels of apolipoprotein A-I and cholesterol. Am. J. Clin. Pathol. 2001, 115, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Murali, N.; Browne, R.W.; Maxwell, K.F.; Bodziak, M.L.; Jakimovski, D.; Hagemaier, J.; Bergsland, N.; Weinstock-Guttman, B.; Zivadinov, R.; Ramanathan, M. Cholesterol and neurodegeneration: Longitudinal changes in serum cholesterol biomarkers are associated with new lesions and gray matter atrophy in multiple sclerosis over 5 years of follow-up. Eur. J. Neurol. 2020, 27, 188-e4. [Google Scholar] [CrossRef]
- Jakimovski, D.; Gandhi, S.; Paunokoski, I.; Bergsland, N.; Hagemeier, J.; Ramasamy, D.P.; Hojnacki, D.; Kolb, C.; Benedict, R.H.B.; Weinstock-Guttman, B.; et al. Hypertension and heart disease are associated with development of brain atrophy in multiple sclerosis: A 5-year longitudinal study. Eur. J. Neurol. 2019, 26, 87-e8. [Google Scholar] [CrossRef] [Green Version]
- McComb, M.; Parambi, R.; Browne, R.W.; Bodziak, M.L.; Jakimovski, D.; Bergsland, N.; Maceski, A.; Weinstock-Guttman, B.; Kuhle, J.; Zivadinov, R.; et al. Apolipoproteins AI and E are associated with neuroaxonal injury to gray matter in multiple sclerosis. Mult. Scler. Relat. Disord. 2020, 45, 102389. [Google Scholar] [CrossRef]
- McComb, M.; Kirkheli, M.; Uher, T.; Browne, R.W.; Srpova, B.; Oechtering, J.; Maleska Maceski, A.; Tyblova, M.; Jakimovski, D.; Ramasamy, D.P.; et al. Neuroprotective associations of apolipoproteins A-I and A-II with neurofilament levels in early multiple sclerosis. J. Clin. Lipidol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, P.B.; Quigley, J.P. Alpha2-macroglobulin: An evolutionarily conserved arm of the innate immune system. Dev. Comp. Immunol. 1999, 23, 375–390. [Google Scholar] [CrossRef]
- Yoshino, S.; Fujimoto, K.; Takada, T.; Kawamura, S.; Ogawa, J.; Kamata, Y.; Kodera, Y.; Shichiri, M. Molecular form and concentration of serum α(2)-macroglobulin in diabetes. Sci. Rep. 2019, 9, 12927. [Google Scholar] [CrossRef] [PubMed]
- Larionov, S.; Dedeck, O.; Birkenmeier, G.; Thal, D.R. Expression of alpha2-macroglobulin, neutrophil elastase, and interleukin-1alpha differs in early-stage and late-stage atherosclerotic lesions in the arteries of the circle of Willis. Acta Neuropathol. 2007, 113, 33–43. [Google Scholar] [CrossRef]
- Mori, T.; Sasaki, J.; Kawaguchi, H.; Handa, K.; Takada, Y.; Matsynaga, A.; Kono, S.; Arakawa, K. Serum glycoproteins and severity of coronary atherosclerosis. Am. Heart J. 1995, 129, 234–238. [Google Scholar] [CrossRef]
- Saxena, R.; Talwar, K.K.; Kotte, V.K.; Rao, Y.V.; Salam, S.R.; Saray, A.K. Evaluation of factors predisposing to arterial thrombosis in coronary artery disease. Indian Heart J. 1990, 42, 433–435. [Google Scholar] [PubMed]
- Lublin, F.D.; Reingold, S.C. Defining the clinical course of multiple sclerosis: Results of an international survey. National Multiple Sclerosis Society (USA) Advisory Committee on Clinical Trials of New Agents in Multiple Sclerosis. Neurology 1996, 46, 907–911. [Google Scholar] [CrossRef]
- Nuttall, F.Q. Body Mass Index: Obesity, BMI, and Health: A Critical Review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Kurtzke, J.F. Rating neurologic impairment in multiple sclerosis: An expanded disability status scale (EDSS). Neurology 1983, 33, 1444–1452. [Google Scholar] [CrossRef] [Green Version]
- Beck, A.T.; Steer, R.A.; Ball, R.; Ranieri, W. Comparison of Beck Depression Inventories -IA and -II in psychiatric outpatients. J. Personal. Assess. 1996, 67, 588–597. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Correlated Parameters | Corrections | ||||
|---|---|---|---|---|---|
| Benjamini–Hochberg (Desired FDR (Q) = 5%) | Holm–Sidak (α = 0.05) | Bonferroni–Sidak (α = 0.05) | |||
| Rho | p-Value | Adjusted p1-Value | Adjusted p2-Value | Adjusted p3-Value | |
| mRNA Expression for the A2M gene in Control Blood Platelets vs. | |||||
| mRNA expression for the PAR1 gene | 0.5986 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAR1 (ng/mL) | 0.6525 | 0.0023 | 0.0038 | 0.0114 | 0.0228 |
| PAGs (%) (no agonist) | 0.6028 | 0.0005 | 0.001 | 0.003 | 0.005 |
| PAGs (%) (TRAP-6) | 0.5251 | 0.0032 | 0.0046 | 0.0127 | 0.0315 |
| PMPs (%) (no agonist) | 0.3925 | 0.0327 | 0.0327 | 0.0492 | 0.2828 |
| PMPs (%) (TRAP-6) | 0.6741 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| P-selectin (%) (no agonist) | 0.4104 | 0.0249 | 0.0277 | 0.0492 | 0.2229 |
| P-selectin (%) (TRAP-6) | 0.7294 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PLAs (%) (no agonist) | 0.4430 | 0.015 | 0.0188 | 0.0443 | 0.1403 |
| PLAs (%) (TRAP-6) | 0.6154 | 0.0004 | 0.001 | 0.0028 | 0.004 |
| mRNA expression for the A2M gene in SP MS Blood Platelets vs. | |||||
| mRNA expression for the PAR1 gene | 0.8454 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAR1 (ng/mL) | 0.5994 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (no agonist) | 0.6628 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (TRAP-6) | 0.4314 | 0.018 | 0.02 | 0.0357 | 0.1661 |
| PMPs (%) (no agonist) | 0.4003 | 0.0291 | 0.0291 | 0.0357 | 0.2557 |
| PMPs (%) (TRAP-6) | 0.4870 | 0.0069 | 0.0099 | 0.0273 | 0.0669 |
| P-selectin (%) (no agonist) | 0.6082 | 0.0005 | 0.0008 | 0.0025 | 0.005 |
| P-selectin (%) (TRAP-6) | 0.4724 | 0.009 | 0.0113 | 0.0273 | 0.0864 |
| PLAs (%) (no agonist) | 0.6367 | 0.0002 | 0.0004 | 0.0012 | 0.002 |
| PLAs (%) (TRAP-6) | 0.6935 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| mRNA expression for the A2M gene in Control Megakaryocytes vs. | |||||
| mRNA expression for the PAR1 gene | 0.4322 | 0.0033 | 0.0039 | 0.0099 | 0.0325 |
| PAR1 (ng/mL) | 0.6897 | 0.002 | 0.0029 | 0.008 | 0.0198 |
| PAGs (%) (no agonist) | 0.6175 | 0.0005 | 0.0013 | 0.0035 | 0.005 |
| PAGs (%) (TRAP-6) | 0.4474 | 0.0138 | 0.0138 | 0.0138 | 0.1297 |
| PMPs (%) (no agonist) | 0.5658 | 0.0013 | 0.0023 | 0.0078 | 0.0129 |
| PMPs (%) (TRAP-6) | 0.6806 | 0.0001 | 0.001 | 0.001 | 0.001 |
| P-selectin (%) (no agonist) | 0.5220 | 0.0035 | 0.0039 | 0.0099 | 0.0345 |
| P-selectin (%) (TRAP-6) | 0.6259 | 0.0003 | 0.001 | 0.0027 | 0.003 |
| PLAs (%) (no agonist) | 0.5639 | 0.0014 | 0.0023 | 0.0078 | 0.0139 |
| PLAs (%) (TRAP-6) | 0.6325 | 0.0003 | 0.001 | 0.0027 | 0.003 |
| mRNA expression for the A2M gene in SP MS Megakaryocytes vs. | |||||
| mRNA expression for the PAR1 gene | 0.7112 | 0.0006 | 0.0009 | 0.003 | 0.006 |
| PAR1 (ng/mL) | 0.7112 | 0.0006 | 0.0009 | 0.003 | 0.006 |
| PAGs (%) (no agonist) | 0.6533 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (TRAP-6) | 0.5111 | 0.0043 | 0.0043 | 0.0074 | 0.0422 |
| PMPs (%) (no agonist) | 0.7447 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PMPs (%) (TRAP-6) | 0.6565 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| P-selectin (%) (no agonist) | 0.6110 | 0.0004 | 0.0008 | 0.0028 | 0.004 |
| P-selectin (%) (TRAP-6) | 0.6157 | 0.0004 | 0.0008 | 0.0028 | 0.004 |
| PLAs (%) (no agonist) | 0.5728 | 0.0011 | 0.0014 | 0.0033 | 0.0109 |
| PLAs (%) (TRAP-6) | 0.5193 | 0.0037 | 0.0041 | 0.0074 | 0.0364 |
| mRNA expression for the APOA1 gene in Control Blood Platelets vs. | |||||
| mRNA expression for the PAR1 gene | −0.5779 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAR1 (ng/mL) | −0.4964 | 0.0276 | 0.0276 | 0.0276 | 0.2441 |
| PAGs (%) (no agonist) | −0.7107 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (TRAP-6) | −0.7035 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PMPs (%) (no agonist) | −0.4675 | 0.0098 | 0.0109 | 0.0253 | 0.0938 |
| PMPs (%) (TRAP-6) | −0.6024 | 0.0011 | 0.0024 | 0.0077 | 0.0109 |
| P-selectin (%) (no agonist) | −0.4835 | 0.0085 | 0.0106 | 0.0253 | 0.0818 |
| P-selectin (%) (TRAP-6) | −0.5713 | 0.0012 | 0.0024 | 0.0077 | 0.0119 |
| PLAs (%) (no agonist) | −0.5112 | 0.0051 | 0.0073 | 0.0202 | 0.0498 |
| PLAs (%) (TRAP-6) | −0.5412 | 0.0023 | 0.0038 | 0.0114 | 0.0228 |
| mRNA expression for the APOA1 gene in SP MS Blood Platelets vs. | |||||
| mRNA expression for the PAR1 gene | −0.4046 | 0.0062 | 0.0089 | 0.0246 | 0.0603 |
| PAR1 (ng/mL) | −0.7693 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (no agonist) | −0.6823 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PAGs (%) (TRAP-6) | −0.6809 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| PMPs (%) (no agonist) | −0.3627 | 0.0495 | 0.0495 | 0.0495 | 0.3981 |
| PMPs (%) (TRAP-6) | −0.6714 | 0.0001 | 0.0003 | 0.001 | 0.001 |
| P-selectin (%) (no agonist) | −0.4691 | 0.0096 | 0.012 | 0.0285 | 0.092 |
| P-selectin (%) (TRAP-6) | −0.5538 | 0.0018 | 0.0036 | 0.0108 | 0.0179 |
| PLAs (%) (no agonist) | −0.4546 | 0.0123 | 0.0137 | 0.0285 | 0.1164 |
| PLAs (%) (TRAP-6) | −0.5292 | 0.003 | 0.005 | 0.0149 | 0.0296 |
| mRNA expression for the APOA1 gene in Control Megakaryocytes vs. | |||||
| mRNA expression for the PAR1 gene | −0.4631 | 0.0106 | 0.0151 | 0.0417 | 0.1011 |
| PAR1 (ng/mL) | −0.7127 | 0.0006 | 0.0043 | 0.006 | 0.006 |
| PAGs (%) (no agonist) | −0.5555 | 0.0017 | 0.0043 | 0.0119 | 0.0169 |
| PAGs (%) (TRAP-6) | −0.5604 | 0.0015 | 0.0043 | 0.0119 | 0.0149 |
| PMPs (%) (no agonist) | −0.4843 | 0.013 | 0.0163 | 0.0417 | 0.1227 |
| PMPs (%) (TRAP-6) | −0.5156 | 0.004 | 0.008 | 0.0238 | 0.0393 |
| P-selectin (%) (no agonist) | −0.4129 | 0.0241 | 0.0241 | 0.0417 | 0.2165 |
| P-selectin (%) (TRAP-6) | −0.5700 | 0.0012 | 0.0043 | 0.0107 | 0.0119 |
| PLAs (%) (no agonist) | −0.4942 | 0.006 | 0.01 | 0.0296 | 0.0584 |
| PLAs (%) (TRAP-6) | −0.4320 | 0.0179 | 0.0199 | 0.0417 | 0.1652 |
| mRNA expression for the APOA1 gene in SP MS Megakaryocytes vs. | |||||
| mRNA expression for the PAR1 gene | −0.5478 | 0.0024 | 0.008 | 0.0205 | 0.0237 |
| PAR1 (ng/mL) | −0.5737 | 0.0092 | 0.0153 | 0.0452 | 0.0883 |
| PAGs (%) (no agonist) | −0.4983 | 0.0065 | 0.013 | 0.0392 | 0.0631 |
| PAGs (%) (TRAP-6) | −0.3698 | 0.049 | 0.049 | 0.0885 | 0.3949 |
| PMPs (%) (no agonist) | −0.5896 | 0.0023 | 0.008 | 0.0205 | 0.0228 |
| PMPs (%) (TRAP-6) | −0.5887 | 0.0057 | 0.013 | 0.0392 | 0.0556 |
| P-selectin (%) (no agonist) | −0.436 | 0.0304 | 0.038 | 0.0885 | 0.2656 |
| P-selectin (%) (TRAP-6) | −0.3928 | 0.0395 | 0.0439 | 0.0885 | 0.3317 |
| PLAs (%) (no agonist) | −0.6045 | 0.0017 | 0.008 | 0.0169 | 0.0169 |
| PLAs (%) (TRAP-6) | −0.5342 | 0.0114 | 0.0163 | 0.0452 | 0.1083 |
| Parameters with Reference Ranges | Control Group (n = 45) | SP MS (n = 45) |
|---|---|---|
| Age | 49 ± 11.2 | 48.6 ± 12.5 |
| Gender (F—Female; M—Male) | 28(F); 17(M) | 27(F); 18(M) |
| BMI (kg/m2) | 22.3 ± 5.1 | 21.5 ± 9.5 |
| <18.5—underweight | ||
| 18.5–24.9—normal weight | ||
| 25–29.9—overweight | ||
| 30 or greater—obesity [78] | ||
| EDSS (0–10 scale in 0.5 unit increment) | N/A | 5.5. ± 1.9 |
| 1–4.5—from normal to mid disability | ||
| 5–9.5—from moderate to severe disability | ||
| 10—death due to MS [79] | ||
| BDI (1–40 scale) | N/A | 9.6 ± 4.6 |
| 1–9—normal | ||
| 10–15—minimal depressive symptomatology | ||
| 16–31—mild depression | ||
| 32–47—moderate depression | ||
| over 47—severe depression [80] | ||
| Mean disease duration (years) | N/A | 14.5 ± 8.1 |
| Blood platelets (130–400 × 103/μL) | 258.9 ± 68.10 | 321.9 ± 43 |
| CRP (<5 mg/L) | 3.15 ± 0.9 | 12.5 ± 5.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziedzic, A.; Miller, E.; Bijak, M.; Przyslo, L.; Saluk-Bijak, J. Increased Pro-Thrombotic Platelet Activity Associated with Thrombin/PAR1-Dependent Pathway Disorder in Patients with Secondary Progressive Multiple Sclerosis. Int. J. Mol. Sci. 2020, 21, 7722. https://doi.org/10.3390/ijms21207722
Dziedzic A, Miller E, Bijak M, Przyslo L, Saluk-Bijak J. Increased Pro-Thrombotic Platelet Activity Associated with Thrombin/PAR1-Dependent Pathway Disorder in Patients with Secondary Progressive Multiple Sclerosis. International Journal of Molecular Sciences. 2020; 21(20):7722. https://doi.org/10.3390/ijms21207722
Chicago/Turabian StyleDziedzic, Angela, Elzbieta Miller, Michal Bijak, Lukasz Przyslo, and Joanna Saluk-Bijak. 2020. "Increased Pro-Thrombotic Platelet Activity Associated with Thrombin/PAR1-Dependent Pathway Disorder in Patients with Secondary Progressive Multiple Sclerosis" International Journal of Molecular Sciences 21, no. 20: 7722. https://doi.org/10.3390/ijms21207722