Ubiquitin Pathway Is Associated with Worsening Left Ventricle Function after Mitral Valve Repair: A Global Gene Expression Study


 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
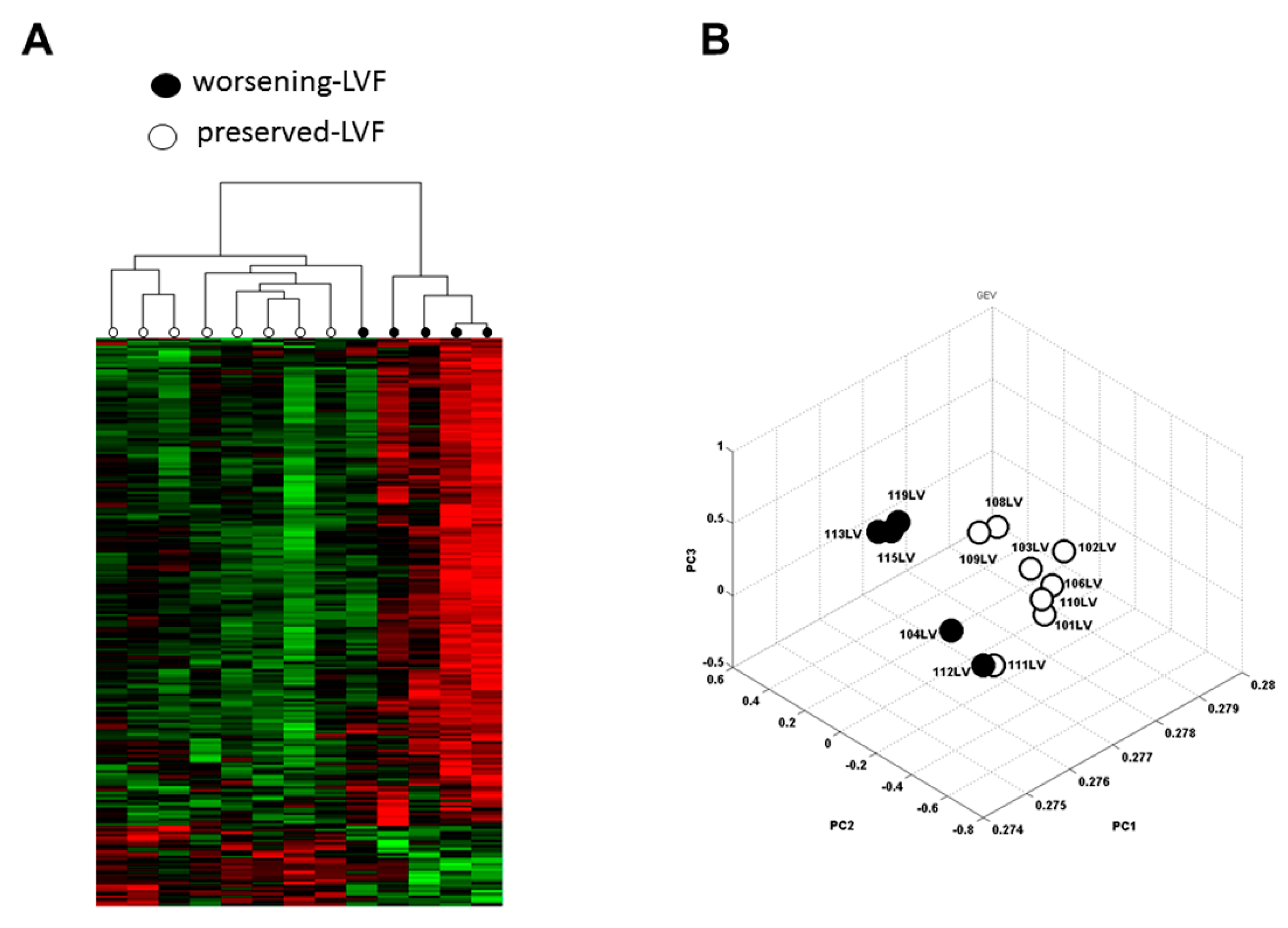
2.2. Microarray
2.3. Validation of Microarray with q-PCR
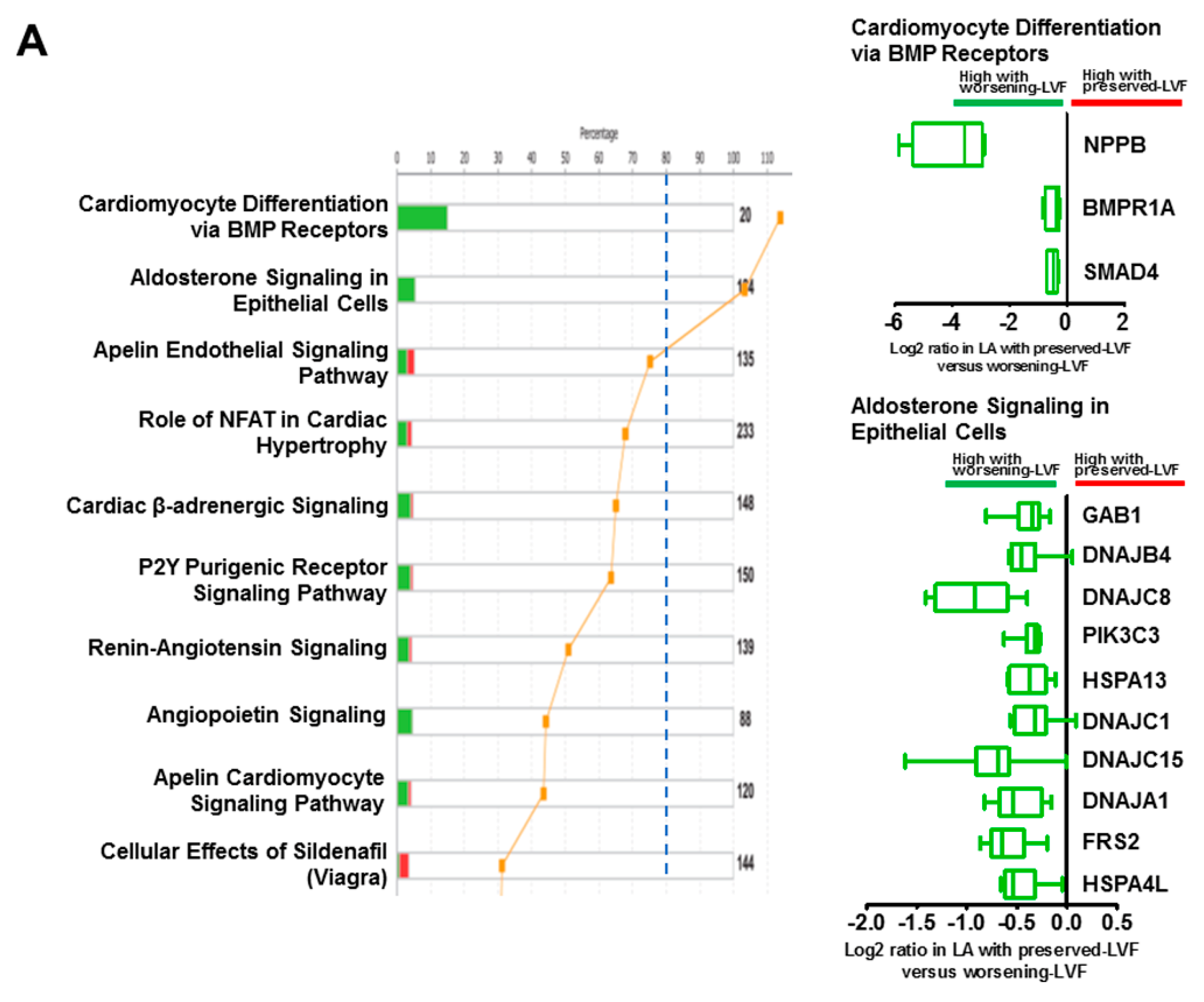
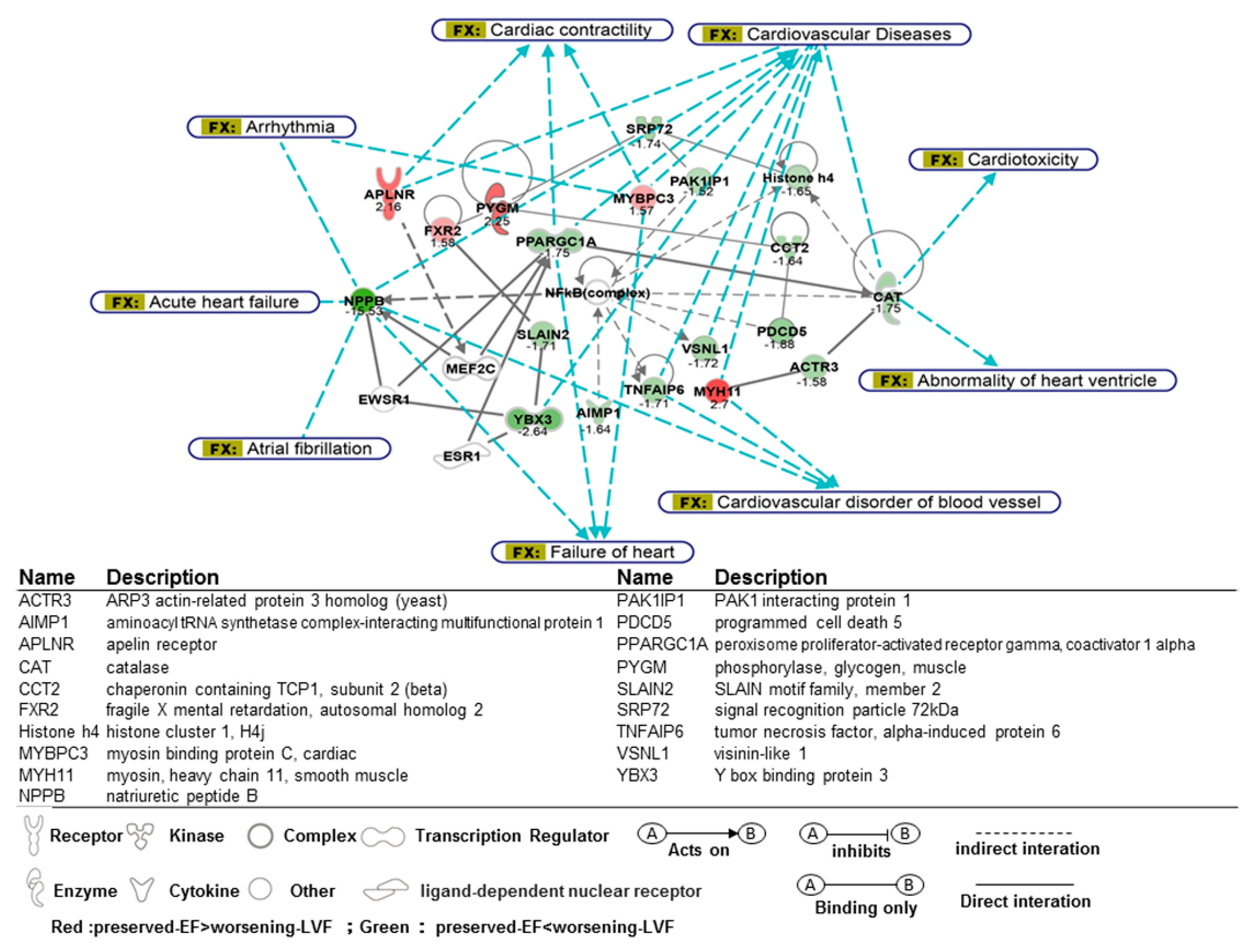
2.4. Functional Analysis of Microarray
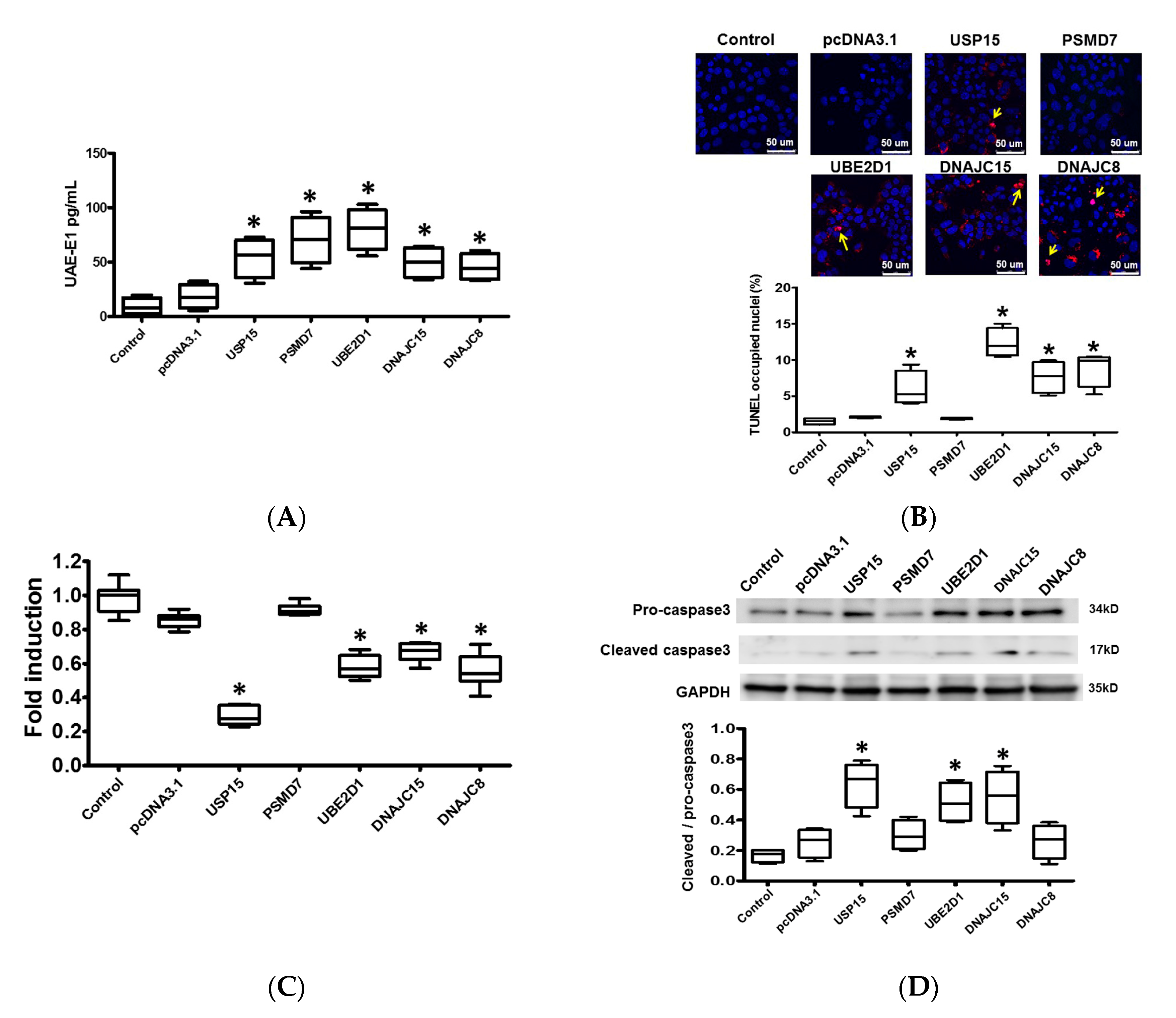
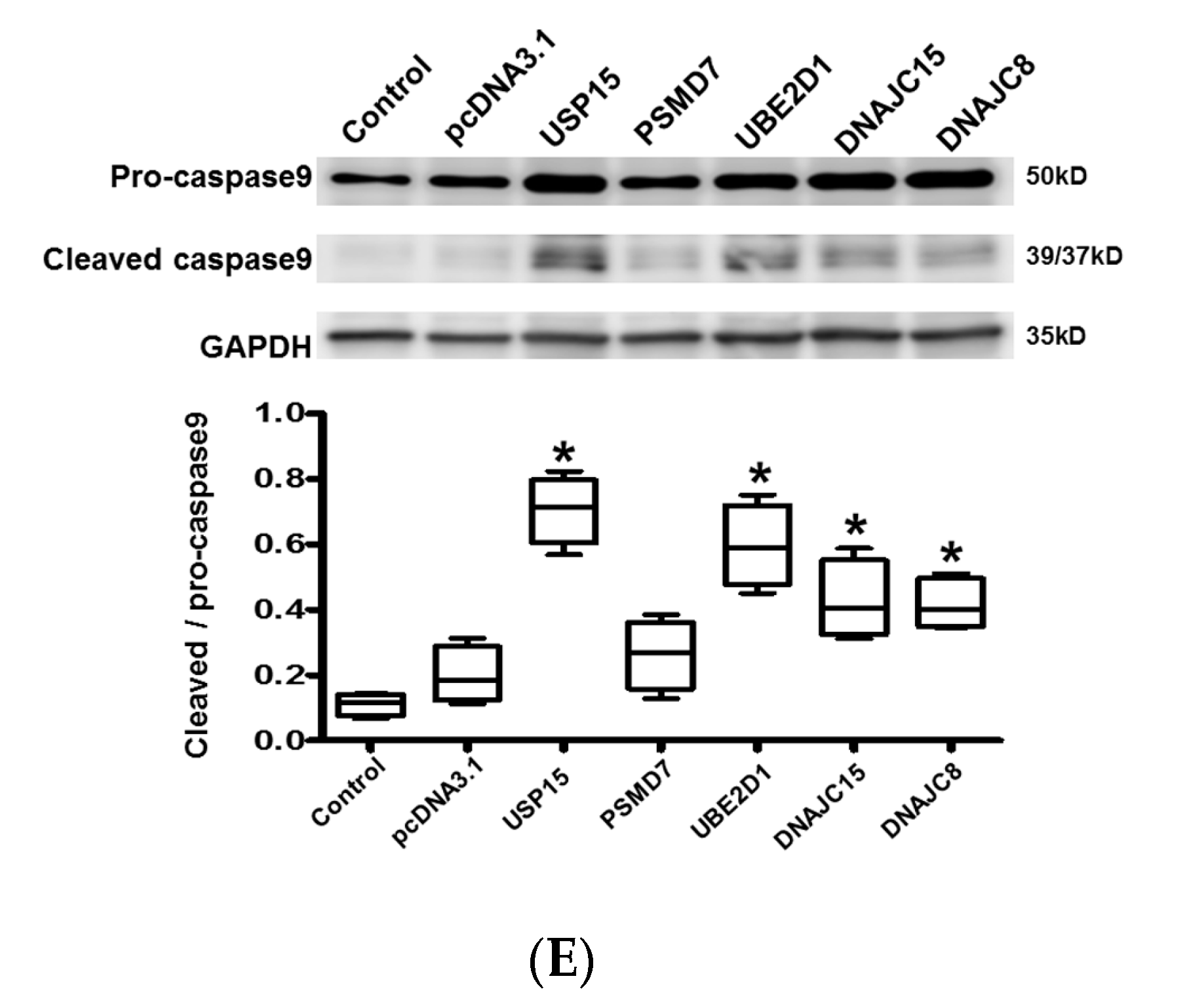
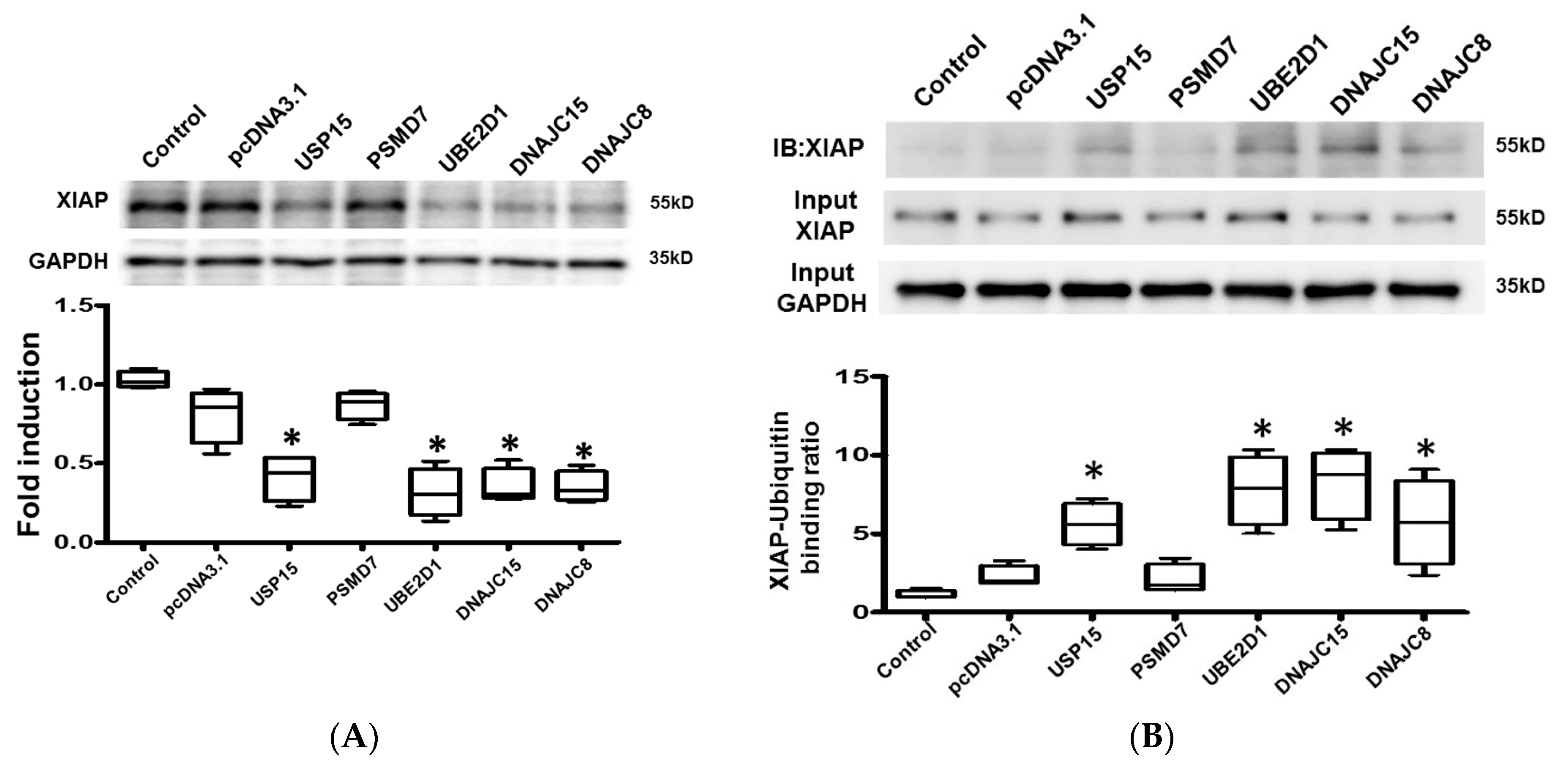
2.5. Protein Ubiquitination in Cardiomyocytes
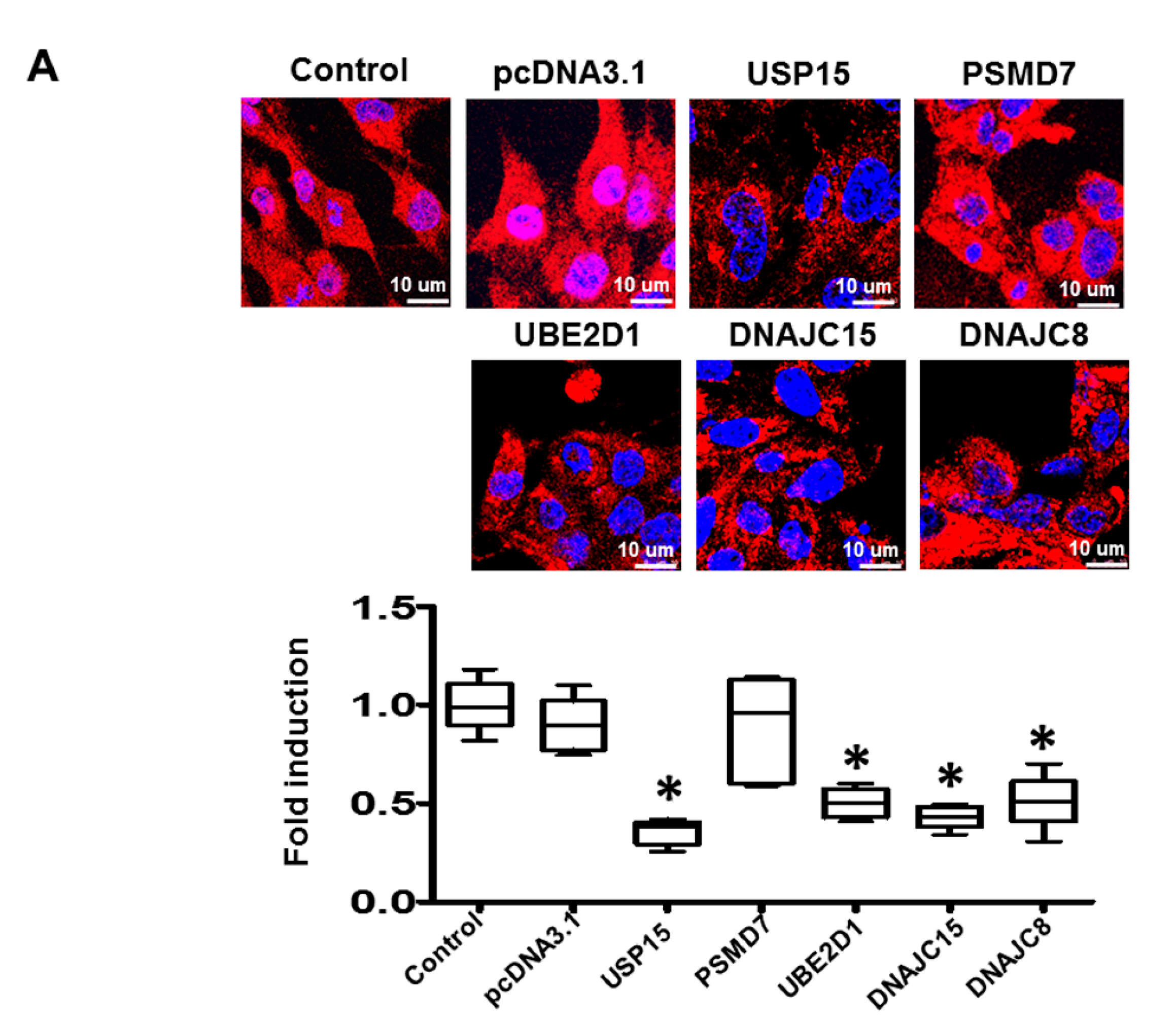
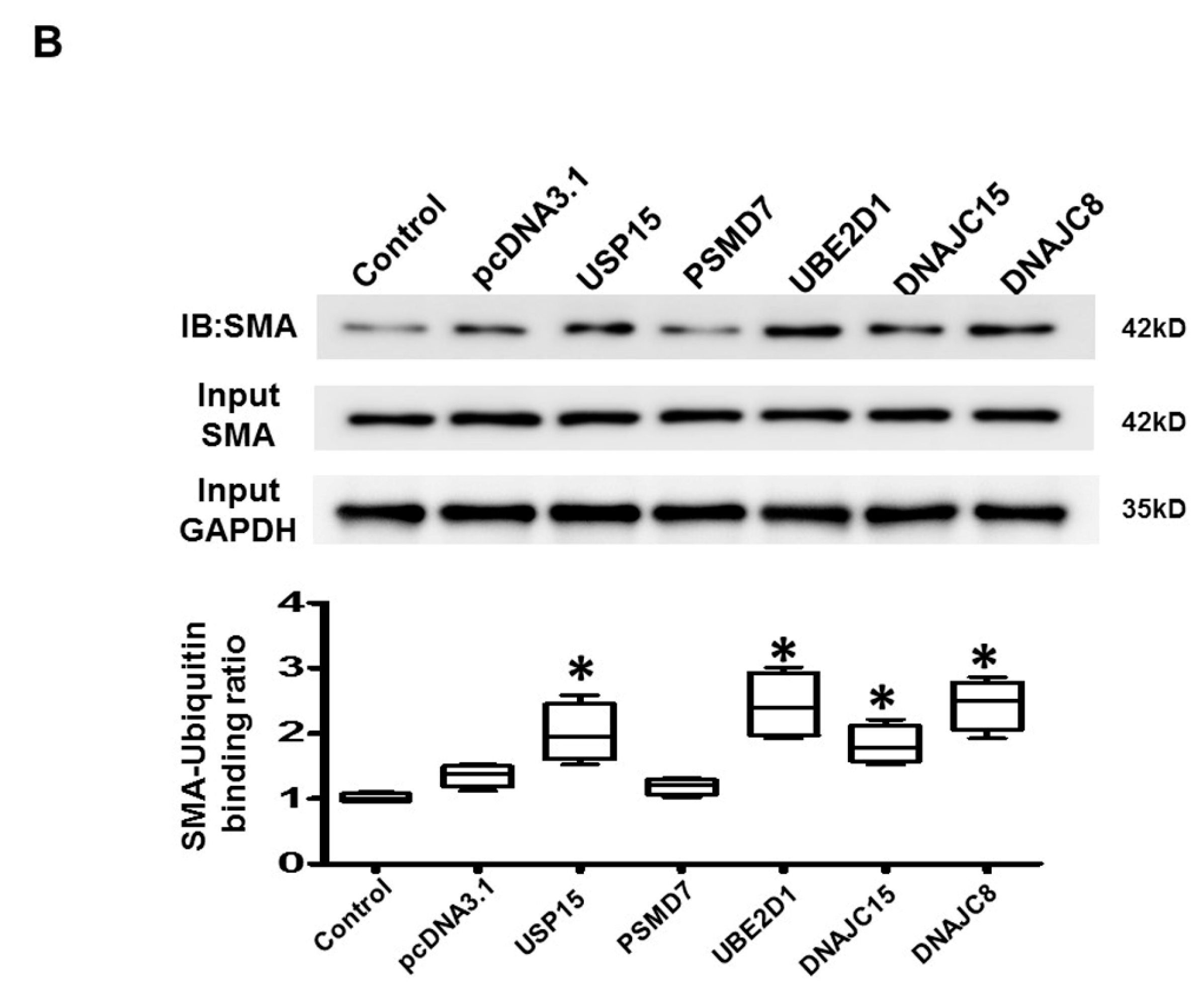
2.6. Apoptosis and Intracellular Contractile Protein Degradation in Cardiomyocytes
3. Discussion
3.1. Main Findings
3.2. Chronic, Severe MR-Related LV Remodeling and Dysfunction
3.3. Ubiquitin-ProteasomeSystem (UPS) and Heart Failure
3.4. Signaling Pathways and Molecules Associated with Worsening LV Function
3.5. Limitations
4. Conclusions
5. Materials and Methods
5.1. Study Subjects
5.2. Microarray Studies
5.3. Pathway and Network Analysis
5.4. Quantitative Polymerase Chain Reaction (q-PCR)
5.5. HL-1 Cell Culture
5.6. Expression Vectors and Transfection
5.7. Determination of Ubiquitin Activating Enzyme E1(UAE-E1) Concentration
5.8. TUNEL Staining
5.9. Cell Viability Assay
5.10. Western Blot
5.11. Co-Immunoprecipitation
5.12. Immunohistochemistry
5.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Glossary of Abbreviations
| APLNR | Apelin receptor |
| BMP | Bone morphogenetic protein |
| DNAJC15 | DnaJ heat shock protein family (Hsp40) member C15 |
| DNAJC8 | DnaJ heat shock protein family (Hsp40) member C8 |
| EST | Expressed sequence tag |
| IPA | Ingenuity Pathway Analysis |
| LV EF | Left ventricular ejection fraction |
| LVEDD | Left ventricular end-diastolic diameter |
| MHC | Myosin heavy chain |
| MYBPC3 | Myosin binding protein C |
| MYH11 | Myosin heavy chain-11 |
| MV | Mitral valve |
| MR | Mitral regurgitation |
| NPPB | Natriuretic peptide B |
| PCA | Principal components analysis |
| PPARGC1A | Peroxisome proliferator-activated receptor gamma coactivator 1-alpha |
| preserved-LVF | Preserved LV function |
| PSMD7 | Proteasome 26S subunit, non-ATPase-7 |
| q-PCR | Quantitative polymerase chain reaction |
| SMA | α-smooth muscle actin |
| TNFAIP6 | Tumor necrosis factor, α-induced protein 6 |
| UAE-E1 | Ubiquitin activating enzyme E1 |
| UBE2D1 | Ubiquitin conjugating enzyme E2 D1 |
| UPS | Ubiquitin-proteasome system |
| USP15 | Ubiquitin-specific peptidase-15 |
| worsening-LVF | Progressive LV remodeling |
| XIAP | X-linked inhibitor of apoptosis protein |
References
- Nishimura, R.A.; Otto, C.M.; Bonow, R.O.; Carabello, B.A.; Erwin, J.P.; Guyton, R.A., 3rd; O’Gara, P.T.; Ruiz, C.E.; Skubas, N.J.; Sorajja, P.; et al. 2014 AHA/ACC guideline for the management of patients with valvular heart disease: A report of the American College of Cardiology/American Heart Association Task Force on Practice Guidelines. J. Thorac. Cardiovasc. Surg. 2014, 148, e1–e132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enriquez-Sarano, M.; Suri, R.M.; Clavel, M.A.; Mantovani, F.; Michelena, H.I.; Pislaru, S.; Mahoney, D.W.; Schaff, H.V. Is there an outcome penalty linked to guideline-based indications for valvular surgery? Early and long-term analysis of patients with organic mitral regurgitation. J. Thorac. Cardiovasc. Surg. 2015, 150, 50–58. [Google Scholar] [CrossRef] [Green Version]
- Kitai, T.; Okada, Y.; Shomura, Y.; Tani, T.; Kaji, S.; Kita, T.; Furukawa, Y. Timing of valve repair for severe degenerative mitral regurgitation and long-term left ventricular function. J. Thorac. Cardiovasc. Surg. 2014, 148, 1978–1982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balachandran, P.; Schaff, H.V.; Lahr, B.D.; Nguyen, A.; Daly, R.C.; Maltais, S.; Pislaru, S.V.; Dearani, J.A. Preoperative left atrial volume index is associated with postoperative outcomes in mitral valve repair for chronic mitral regurgitation. J. Thorac. Cardiovasc. Surg. 2019. [Google Scholar] [CrossRef] [PubMed]
- Naber, C.K.; Prendergast, B.; Thomas, M.; Vahanian, A.; Iung, B.; Rosenhek, R.; Tornos, P.; Otto, C.M.; Antunes, M.J.; Kappetein, P.; et al. An interdisciplinary debate initiated by the European Society of Cardiology Working Group on Valvular Heart Disease. EuroIntervention 2012, 7, 1257–1274. [Google Scholar] [CrossRef] [PubMed]
- Quintana, E.; Suri, R.M.; Thalji, N.M.; Daly, R.C.; Dearani, J.A.; Burkhart, H.M.; Li, Z.; Enriquez-Sarano, M.; Schaff, H.V. Left ventricular dysfunction after mitral valve repair—The fallacy of “normal” preoperative myocardial function. J. Thorac. Cardiovasc. Surg. 2014, 148, 2752–2760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, I.; Singh, K.; Varshney, N.K.; Khan, S. Delineating Crosstalk Mechanisms of the Ubiquitin Proteasome System That Regulate Apoptosis. Front. Cell Dev. Biol. 2018, 6, 11. [Google Scholar] [CrossRef]
- Zheng, J.; Yancey, D.M.; Ahmed, M.I.; Wei, C.C.; Powell, P.C.; Shanmugam, M.; Gupta, H.; Lloyd, S.G.; McGiffin, D.C.; Schiros, C.G.; et al. Increased sarcolipin expression and adrenergic drive in humans with preserved left ventricular ejection fraction and chronic isolated mitral regurgitation. Circ. Heart Fail. 2014, 7, 194–202. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Chen, Y.; Pat, B.; Dell’italia, L.A.; Tillson, M.; Dillon, A.R.; Powell, P.C.; Shi, K.; Shah, N.; Denney, T.; et al. Microarray identifies extensive downregulation of noncollagen extracellular matrix and profibrotic growth factor genes in chronic isolated mitral regurgitation in the dog. Circulation 2009, 119, 2086–2095. [Google Scholar] [CrossRef] [Green Version]
- Childers, R.C.; Sunyecz, I.; West, T.A.; Cismowski, M.J.; Lucchesi, P.A.; Gooch, K.J. Role of the cytoskeleton in the development of a hypofibrotic cardiac fibroblast phenotype in volume overload heart failure. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H596–H608. [Google Scholar] [CrossRef]
- Ahmed, M.I.; Guichard, J.L.; Soorappan, R.N.; Ahmad, S.; Mariappan, N.; Litovsky, S.; Gupta, H.; Lloyd, S.G.; Denney, T.S.; Powell, P.C.; et al. Disruption of desmin-mitochondrial architecture in patients with regurgitant mitral valves and preserved ventricular function. J. Thorac. Cardiovasc. Surg. 2016, 152, 1059–1070e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCutcheon, K.; Dickens, C.; van Pelt, J.; Dix-Peek, T.; Grinter, S.; McCutcheon, L.; Patel, A.; Hale, M.; Tsabedze, N.; Vachiat, A.; et al. Dynamic Changes in the Molecular Signature of Adverse Left Ventricular Remodeling in Patients With Compensated and Decompensated Chronic Primary Mitral Regurgitation. Circ. Heart Fail. 2019, 12, e005974. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.R.; Herrmann, J.; Lerman, A.; Patterson, C.; Wang, X. The ubiquitin-proteasome system and cardiovascular disease. Prog. Mol. Biol. Transl. Sci. 2012, 109, 295–346. [Google Scholar]
- Spanig, S.; Kellermann, K.; Dieterlen, M.T.; Noack, T.; Lehmann, S.; Borger, M.A.; Garbade, J.; Barac, Y.D.; Emrich, F. The Ubiquitin Proteasome System in Ischemic and Dilated Cardiomyopathy. Int. J. Mol. Sci. 2019, 20, 6354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drews, O.; Taegtmeyer, H. Targeting the ubiquitin-proteasome system in heart disease: The basis for new therapeutic strategies. Antioxid. Redox Signal. 2014, 21, 2322–2343. [Google Scholar] [CrossRef] [Green Version]
- Depre, C.; Wang, Q.; Yan, L.; Hedhli, N.; Peter, P.; Chen, L.; Hong, C.; Hittinger, L.; Ghaleh, B.; Sadoshima, J.; et al. Activation of the cardiac proteasome during pressure overload promotes ventricular hypertrophy. Circulation 2006, 114, 1821–1828. [Google Scholar] [CrossRef] [Green Version]
- Piacentino, V.; Milano, C.A., 3rd; Bolanos, M.; Schroder, J.; Messina, E.; Cockrell, A.S.; Jones, E.; Krol, A.; Bursac, N.; Mao, L.; et al. X-linked inhibitor of apoptosis protein-mediated attenuation of apoptosis, using a novel cardiac-enhanced adeno-associated viral vector. Hum. Gene Ther. 2012, 23, 635–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Q.; Cheng, C.; Wei, Y.; Yang, J.; Zhou, W.; Song, Q.; Ke, M.; Yan, W.; Zheng, L.; Zhang, Y.; et al. USP15 potentiates NF-kappaB activation by differentially stabilizing TAB2 and TAB3. FEBS J. 2020. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, T.; Zhu, X.; Yang, C.; Wang, Y.; Zhou, N.; Ju, B.; Zhou, T.; Deng, G.; Qiu, C. Hyperoside Induces Breast Cancer Cells Apoptosis via ROS-Mediated NF-kappaB Signaling Pathwa. Int. J. Mol. Sci. 2019, 21, 131. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Sato, C.; Kadowaki, A.; Watanabe, H.; Ho, L.; Ishida, J.; Yamaguchi, T.; Kimura, A.; Fukamizu, A.; Penninger, J.M.; et al. ELABELA-APJ axis protects from pressure overload heart failure and angiotensin II-induced cardiac damage. Cardiovasc. Res. 2017, 113, 760–769. [Google Scholar] [CrossRef]
- Japp, A.G.; Cruden, N.L.; Barnes, G.; van Gemeren, N.; Mathews, J.; Adamson, J.; Johnston, N.R.; Denvir, M.A.; Megson, I.L.; Flapan, A.D.; et al. Acute cardiovascular effects of apelin in humans: Potential role in patients with chronic heart failure. Circulation 2010, 121, 1818–1827. [Google Scholar] [CrossRef] [PubMed]
- Kulikova, T.G.; Stepanova, O.V.; Voronova, A.D.; Valikhov, M.P.; Sirotkin, V.N.; Zhirov, I.V.; Tereshchenko, S.N.; Masenko, V.P.; Samko, A.N.; Sukhikh, G.T. Pathological Remodeling of the Myocardium in Chronic Heart Failure: Role of PGC-1alpha. Bull. Exp. Biol. Med. 2018, 164, 794–797. [Google Scholar] [CrossRef]
- Yeh, Y.H.; Kuo, C.T.; Lee, Y.S.; Lin, Y.M.; Nattel, S.; Tsai, F.C.; Chen, W.J. Region-specific gene expression profiles in the left atria of patients with valvular atrial fibrillation. Heart Rhythm 2013, 10, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.C.; Lin, Y.C.; Chang, S.H.; Chang, G.J.; Hsu, Y.J.; Lin, Y.M.; Lee, Y.S.; Wang, C.L.; Yeh, Y.H. Differential left-to-right atria gene expression ratio in human sinus rhythm and atrial fibrillation: Implications for arrhythmogenesis and thrombogenesis. Int. J. Cardiol. 2016, 222, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.H.; Kuo, C.T.; Chang, G.J.; Chen, Y.H.; Lai, Y.J.; Cheng, M.L.; Chen, W.J. Rosuvastatin suppresses atrial tachycardia-induced cellular remodeling via Akt/Nrf2/heme oxygenase-1 pathway. J. Mol. Cell Cardiol. 2015, 82, 84–92. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Worsening-LVF (n = 5) | Preserved-LVF (n = 8) | |
|---|---|---|
| Demographic Features | ||
| Age (years old) | 71.8 ± 14.0 | 62.4 ± 4.4 |
| Sex, M:F | 3:2 | 5:3 |
| Clinical Features | ||
| Pre-op LV EF (%) | 63.6 ± 11.1 | 62.9 ± 9.9 |
| Post-op LV EF (%) | 50.4 ± 4.5 | 64.9 ± 7.2 |
| Left atrialLA size (mm) | 60.0 ± 9.6 | 49.0 ± 8.9 |
| Rheumatic heart disease | 1 | 4 |
| Hypertension | 0 | 2 |
| Diabetes | 0 | 0 |
| Coronary artery disease | 0 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, F.-C.; Chang, G.-J.; Lai, Y.-J.; Chang, S.-H.; Chen, W.-J.; Yeh, Y.-H. Ubiquitin Pathway Is Associated with Worsening Left Ventricle Function after Mitral Valve Repair: A Global Gene Expression Study. Int. J. Mol. Sci. 2020, 21, 5073. https://doi.org/10.3390/ijms21145073
Tsai F-C, Chang G-J, Lai Y-J, Chang S-H, Chen W-J, Yeh Y-H. Ubiquitin Pathway Is Associated with Worsening Left Ventricle Function after Mitral Valve Repair: A Global Gene Expression Study. International Journal of Molecular Sciences. 2020; 21(14):5073. https://doi.org/10.3390/ijms21145073
Chicago/Turabian StyleTsai, Feng-Chun, Gwo-Jyh Chang, Ying-Ju Lai, Shang-Hung Chang, Wei-Jan Chen, and Yung-Hsin Yeh. 2020. "Ubiquitin Pathway Is Associated with Worsening Left Ventricle Function after Mitral Valve Repair: A Global Gene Expression Study" International Journal of Molecular Sciences 21, no. 14: 5073. https://doi.org/10.3390/ijms21145073