New Insights on the Effect of TNF Alpha Blockade by Gene Silencing in Noise-Induced Hearing Loss

 ,
,  ,
, 
 , and
, and
Abstract
:1. Introduction
2. Results
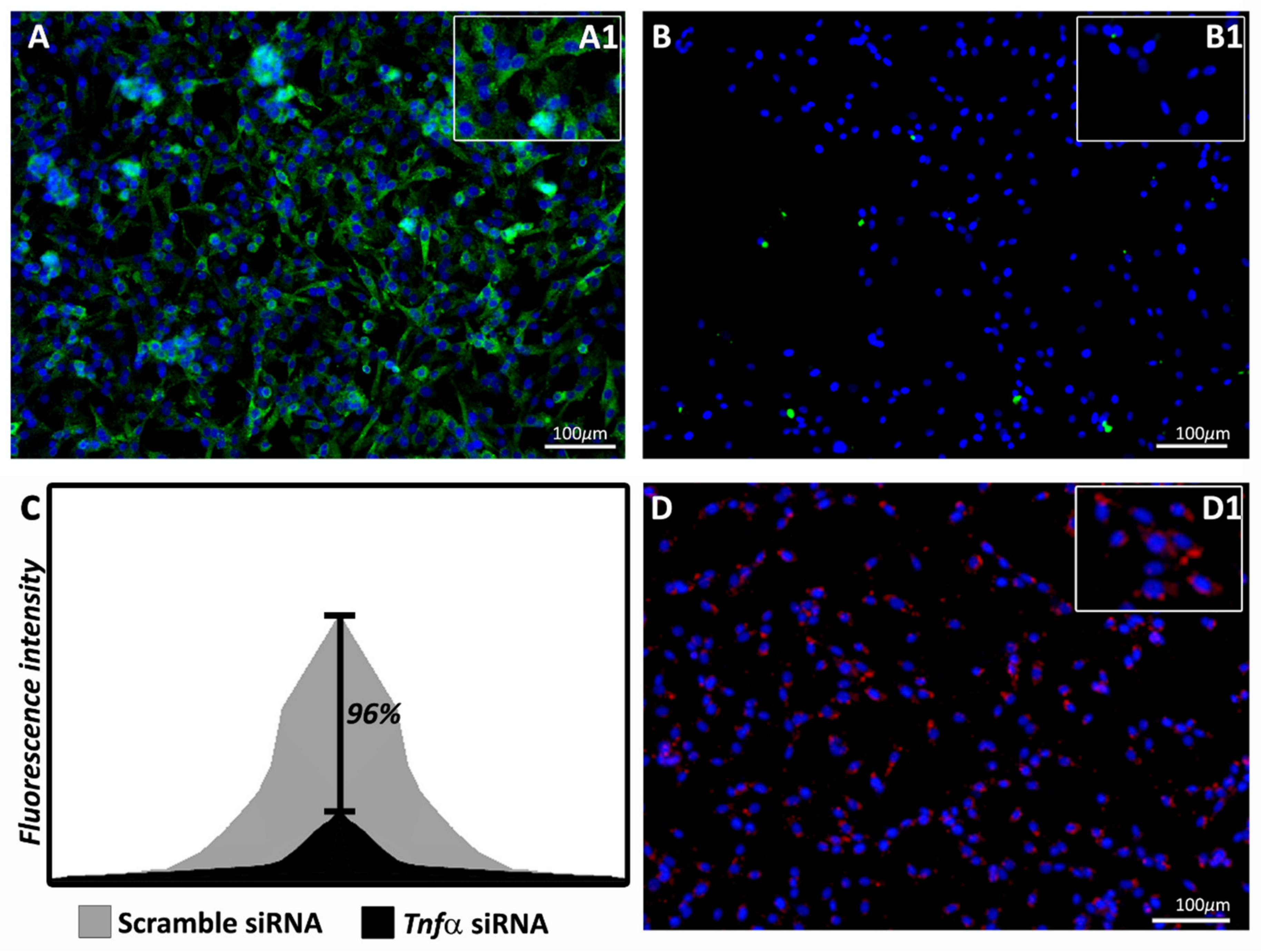
2.1. Tnf alpha siRNA Silencer Was Able to Reduce TNFα Expression
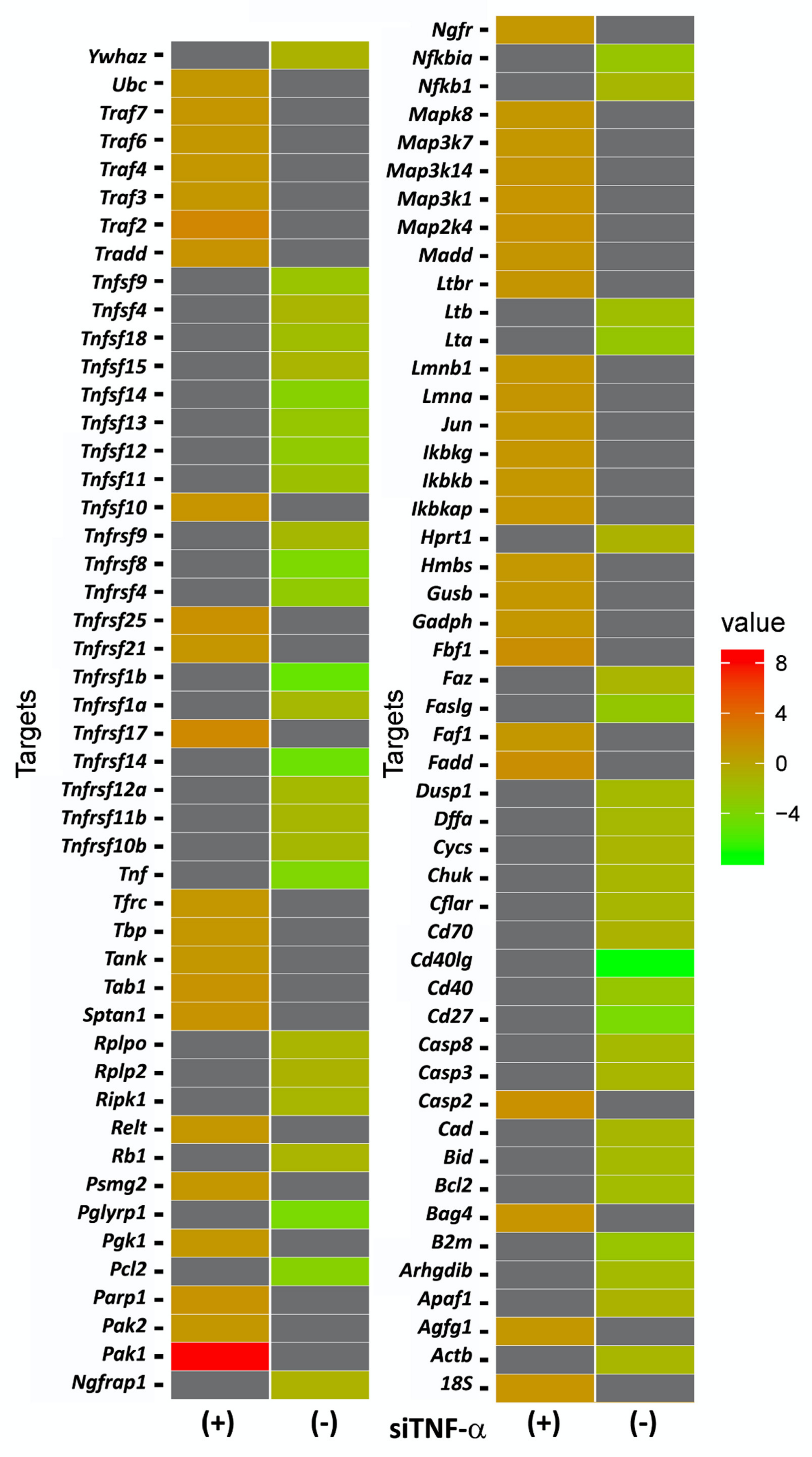
2.2. Tnf alpha siRNA Administration Elicited a Differential Gene Expression in Rats Submitted to Noise Exposure
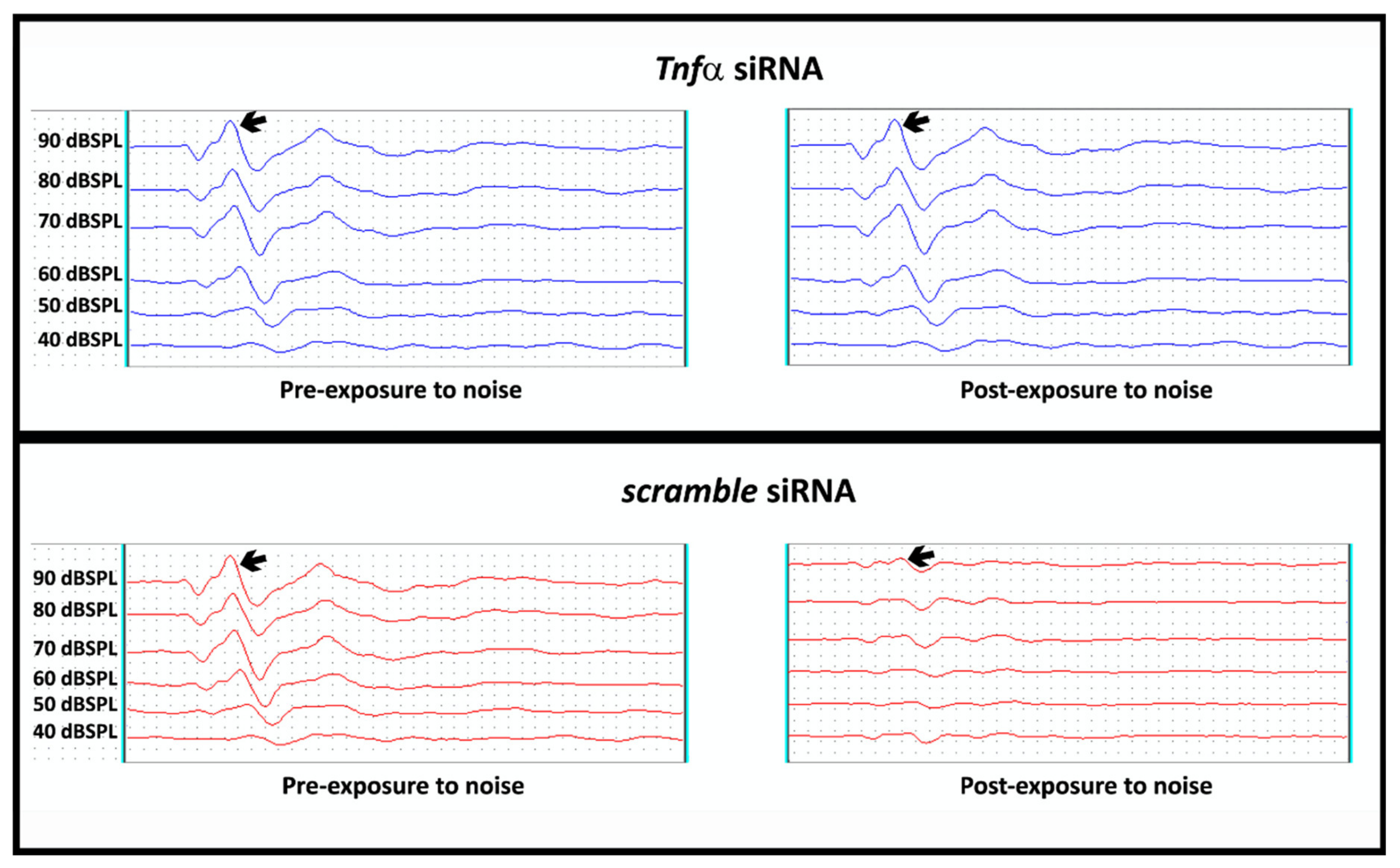
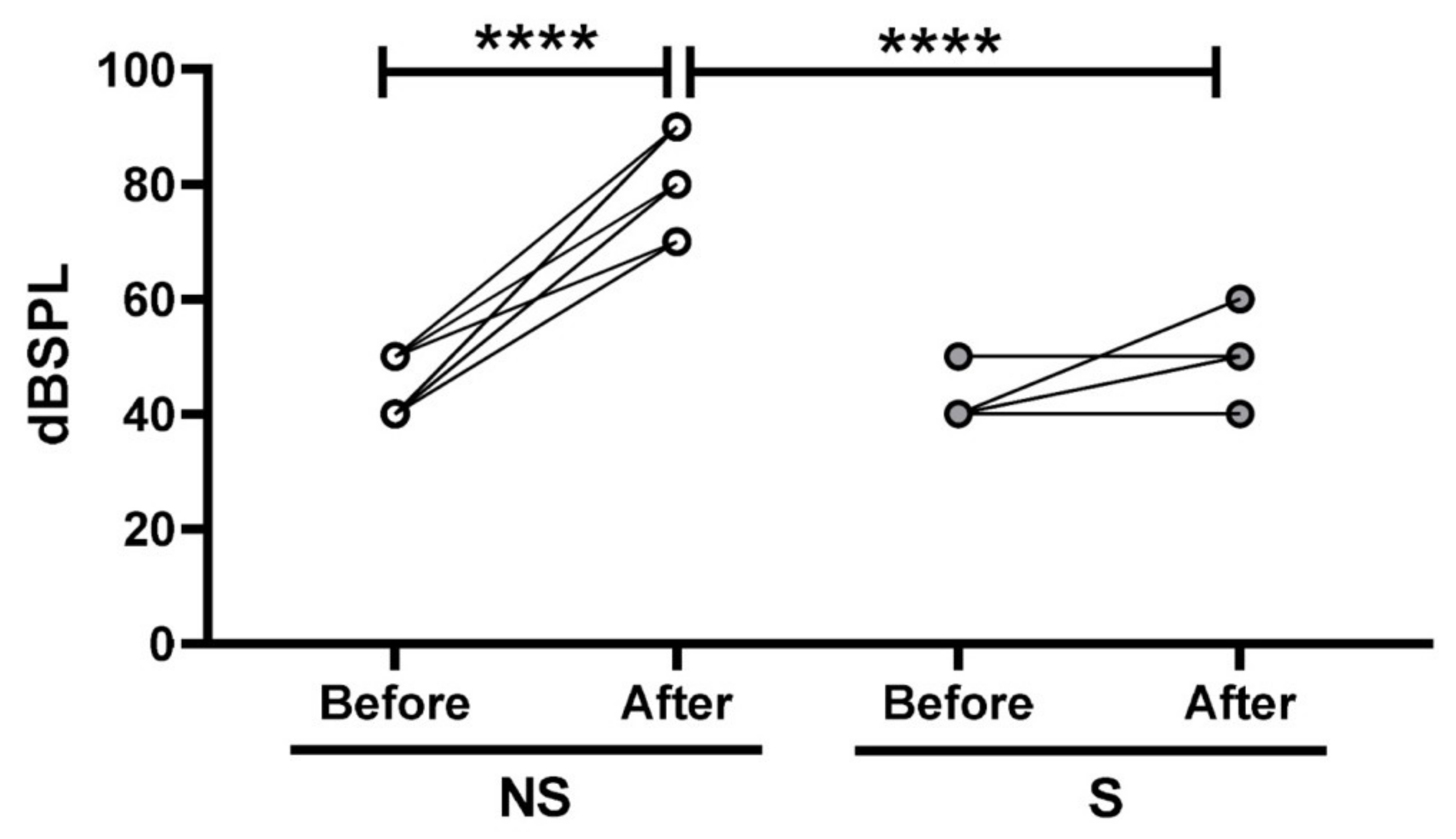
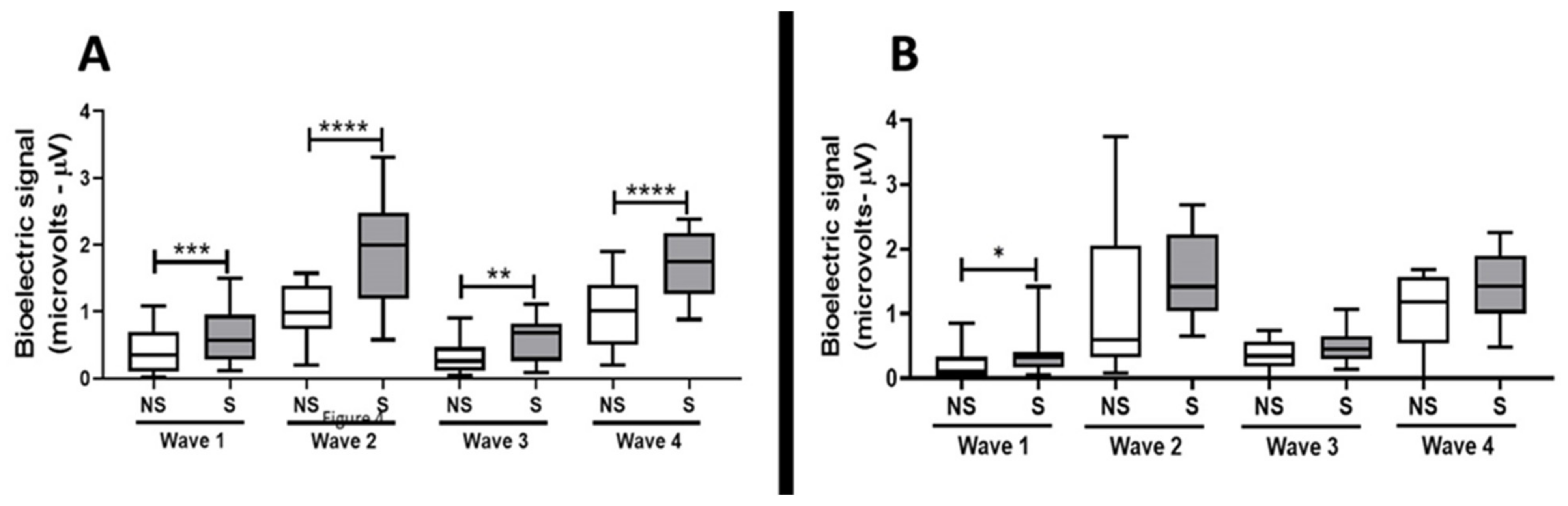
2.3. TNF alpha Blockade by Gene Silence Was Able to Preserve the Hearing in Ears of Rats Submitted to Noise Exposure
3. Discussion
4. Material and Methods
4.1. Materials
4.2. Animal Preparation
4.3. Auditory Brainstem Response
4.4. Trans-Tympanic Injection
4.5. Loud Noise Exposure
4.6. RNA Isolation and REAL-TIME -qPCR
4.7. In Vitro Silencing Analysis with High-Content Screening
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Landegger, L.D.; Vasilijic, S.; Fujita, T.; Soares, V.Y.; Seist, R.; Xu, L.; Stankovic, K.M. Cytokine levels in inner ear fluid of young and aged mice as molecular biomarkers of noise-induced hearing loss. Front. Neurol. 2019, 10, 977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novis, K.; Bell, S. Objective comparison of the quality and reliability of auditory brainstem response features elicited by click and speech sounds. Ear Hear. 2019, 40, 447–457. [Google Scholar] [CrossRef]
- Skoe, E.; Tufts, J. Evidence of noise-induced subclinical hearing loss using auditory brainstem responses and objective measures of noise exposure in humans. Hear. Res. 2018, 361, 80–91. [Google Scholar] [CrossRef]
- Kamogashira, T.; Fujimoto, C.; Yamasoba, T. Reactive oxygen species, apoptosis, and mitochondrial dysfunction in hearing loss. Biomed. Res. Int. 2015, 2015, 617207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coordes, A.; Gröschel, M.; Ernst, A.; Basta, D. Apoptotic cascades in the central auditory pathway after noise exposure. J. Neurotrauma 2012, 29, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Gröschel, M.; Basta, D.; Ernst, A.; Mazurek, B.; Szczepek, A.J. Acute noise exposure is associated with intrinsic apoptosis in murine central auditory pathway. Front. Neurosci. 2018, 12, 312. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, M.; Kanzaki, S.; Okano, H.J.; Masuda, M.; Ogawa, K.; Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J. Neurosci. Res. 2006, 83, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Arpornchayanon, W.; Canis, M.; Ihler, F.; Settevendemie, C.; Strieth, S. TNF alpha inhibition using etanercept prevents noise-induced hearing loss by improvement of cochlear blood flow in vivo. Int. J. Audiol. 2013, 52, 545–552. [Google Scholar] [CrossRef]
- Akash, M.S.H.; Rehman, K.; Liaqat, A. Tumor necrosis factor-alpha: Role in development of insulin resistance and pathogenesis of type 2 diabetes mellitus. J. Cell. Biochem. 2018, 119, 105–110. [Google Scholar] [CrossRef]
- Ehlers, S. Role of tumour necrosis factor (TNF) in host defence against tuberculosis: Implications for immunotherapies targeting TNF. Ann. Rheum. Dis. 2003, 62, ii37–ii42. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.; Maier, O. Interrelation of oxidative stress and inflammation in neurodegenerative disease: Role of TNF. Oxidative Med. Cell. Longev. 2015, 2015, 610813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, J.; Pyykkö, I.; Sutinen, P.; Toppila, E. Vibration induced hearing loss in guinea pig cochlea: Expression of TNF-α and VEGF. Hear. Res. 2005, 202, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Lee, M.H.; Kang, S.U.; Hwang, H.S.; Park, K.; Choung, Y.H.; Kim, C.H. Nitric oxide mediates TNF-α–induced apoptosis in the auditory cell line. Laryngoscope 2012, 122, 2256–2264. [Google Scholar] [CrossRef]
- Wu, Q.; Wang, G.P.; Xie, J.; Guo, J.Y.; Gong, S.S. Tumor necrosis factor-α-induced ototoxicity in mouse cochlear organotypic culture. PLoS ONE 2015, 10, e0127703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eshraghi, A.A.; Gupta, C.; Van De Water, T.R.; Bohorquez, J.E.; Garnham, C.; Bas, E.; Talamo, V.M. Molecular mechanisms involved in cochlear implantation trauma and the protection of hearing and auditory sensory cells by inhibition of c-jun-N-terminal kinase signaling. Laryngoscope 2013, 123, S1–S14. [Google Scholar] [CrossRef] [PubMed]
- Elbashir, S.M.; Lendeckel, W.; Tuschl, T. RNA interference is mediated by 21-and 22-nucleotide RNAs. Genes Dev. 2001, 15, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Van De Water, T.R.; Dinh, C.T.; Vivero, R.; Hoosien, G.; Eshraghi, A.A.; Balkany, T.J. Mechanisms of hearing loss from trauma and inflammation: Otoprotective therapies from the laboratory to the clinic. Acta Oto-Laryngol. 2010, 130, 308–311. [Google Scholar] [CrossRef]
- Marshall, A.; Celentano, A.; Cirillo, N.; Mirams, M.; McCullough, M.; Porter, S. Immune receptors CD40 andCD86 in oral keratinocytes and implications for oral lichen planus. J. Oral Sci. 2017, 59, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Moon, S.K.; Woo, J.I.; Lim, D.J. Involvement of TNF-α and IFN-γ in inflammation-mediated cochlear injury. Ann. Otol. Rhinol. Laryngol. 2019, 128, 8S–15S. [Google Scholar] [CrossRef]
- Arpornchayanon, W. Effects of TNF-Alpha Inhibition on Inner Ear Microcirculation and Hearing Functionafter Acute Loud Noise In Vivo. Ph.D. Thesis, Medical Faculty. Ludwig Maximilians University, Munich, Germany, 2010. [Google Scholar]
- Ruigrok, M.J.; Xian, J.L.; Frijlink, H.W.; Melgert, B.N.; Hinrichs, W.L.; Olinga, P. siRNA-mediated proteinknockdown in precision-cut lung slices. Eur. J. Pharm. Biopharm. 2018, 133, 339–348. [Google Scholar] [CrossRef]
- Mukherjea, D.; Jajoo, S.; Whitworth, C.; Bunch, J.R.; Turner, J.G.; Rybak, L.P.; Ramkumar, V. Short interfering RNA against transient receptor potential vanilloid 1 attenuates cisplatin-induced hearing loss in the rat. J. Neurosci. 2008, 28, 13056–13065. [Google Scholar] [CrossRef] [Green Version]
- Mukherjea, D.; Jajoo, S.; Kaur, T.; Sheehan, K.E.; Ramkumar, V.; Rybak, L.P. Trans-tympanic administration of short interfering (si) RNA for the NOX3 isoform of NADPH oxidase protects against cisplatin-inducedhearing loss in the rat. Antioxid. Redox Signal. 2010, 13, 589–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjea, D.; Jajoo, S.; Sheehan, K.; Kaur, T.; Sheth, S.; Bunch, J.; Perro, C.; Rybak, L.P.; Ramkumar, V. NOX3 NADPH oxidase couples transient receptor potential vanilloid 1 to signal transducer and activator of transcription 1-mediated inflammation and hearing loss. Antioxid. Redox Signal. 2011, 14, 999–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Yuan, H.; Talaska, A.E.; Hill, K.; Sha, S.H. Increased sensitivity to noise-induced hearing loss by blockade of endogenous PI3K/Akt signaling. J. Assoc. Res. Otolaryngol. 2015, 16, 347–356. [Google Scholar] [CrossRef] [Green Version]
- Sakon, S.; Xue, X.; Takekawa, M.; Sasazuki, T.; Okazaki, T.; Kojima, Y.; Piao, J.H.; Yagita, H.; Okumura, K.; Doi, T.; et al. NF-κB inhibits TNF-induced accumulation of ROS that mediate prolonged MAPK activationand necrotic cell death. EMBO J. 2003, 22, 3898–3909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devin, A.; Cook, A.; Lin, Y.; Rodriguez, Y.; Kelliher, M.; Liu, Z.G. The distinct roles of TRAF2 and RIP in IKK activation by TNF-R1: TRAF2 recruits IKK to TNF-R1 while RIP mediates IKK activation. Immunity 2000, 12, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Schürmann, A.; Mooney, A.F.; Sanders, L.C.; Sells, M.A.; Wang, H.G.; Reed, J.C.; Bokoch, G.M. p21-activated kinase 1phosphorylates the death agonist bad and protects cells from apoptosis. Mol. Cell Biol. 2000, 20, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Frost, J.A.; Swantek, J.L.; Stippec, S.; Yin, M.J.; Gaynor, R.; Cobb, M.H. Stimulation of NFκB activity by multiplesignaling pathways requires PAK1. J. Biol. Chem. 2000, 275, 19693–19699. [Google Scholar] [CrossRef] [Green Version]
- Portt, L.; Norman, G.; Clapp, C.; Greenwood, M.; Greenwood, M.T. Anti-apoptosis and cell survival: A review. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2011, 1813, 238–259. [Google Scholar] [CrossRef] [Green Version]
- Wajant, H.; Pfizenmaier, K.; Scheurich, P. Tumor necrosis factor signaling. Cell Death Differ. 2003, 10, 45–65. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Goeddel, D.V. TNF-R1 signaling: A beautiful pathway. Science 2002, 296, 1634–1635. [Google Scholar] [CrossRef] [Green Version]
- Durrant, J.D.; Boston, J.R. Stimuli for auditory evoked potential assessment. In Auditory Evoked Potentials, Basic Principle and Clinical Application; Burkard, R.F., Eggermont, J.J., Don, M., Eds.; Lippincott Williams and Wilkins: Baltimore, MD, USA, 2007. [Google Scholar]
- Overbeck, G.W.; Church, M.W. Effects of tone burst frequency and intensity on the auditory brainstem response (ABR) from albino and pigmented rats. Hear. Res. 1992, 59, 129–137. [Google Scholar] [CrossRef]
- Alvarado, J.C.; Fuentes-Santamaría, V.; Jareño-Flores, T.; Blanco, J.L.; Juiz, J.M. Normal variation in the morphology of auditory brainstem response (ABR) waveform: A study in wistar rats. Neurosci. Res. 2012, 73, 302–311. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Adding insult to injury: Cochlear nerve degeneration after “temporary” noise-induced hearing loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamper, G.C.; Johnson, T.A. Auditory function in normal-hearing, noise-exposed human ears. Ear Hear. 2015, 36, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syka, J. Plastic changes in the central auditory system after hearing loss, restoration of function, and during learning. Physiol. Rev. 2002, 82, 601–636. [Google Scholar] [CrossRef]
- Popelar, J.; Grecova, J.; Rybalko, N.; Syka, J. Comparison of noise-induced changes of auditory brainstem and middle latency response amplitudes in rats. Hear. Res. 2008, 245, 82–91. [Google Scholar] [CrossRef]
- Oishi, N.; Chen, F.Q.; Zheng, H.W.; Sha, S.H. Intra-tympanic delivery of short interfering RNA into the adult mouse cochlea. Hear. Res. 2013, 296, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C. Effect of TNF-A Gene Silencing On Auditory Preservation in Wistar Rats Exposed to Noise and Analysis of Gene Expression of This Metabolic Pathway. Ph.D. Thesis, School of Medicine, University of São Paulo, São Paulo, Brazil, 2018. [Google Scholar]
- Melgar-Rojas, P.; Alvarado, J.C.; Fuentes-Santamaría, V.; Gabaldón-Ull, M.C.; Juiz, J.M. Validation of reference genes for RT–qPCR analysis in noise–induced hearing loss: A study in wistar rat. PLoS ONE 2015, 10, e0138027. [Google Scholar] [CrossRef]
- Yu, J.; Xiang, M.; Wu, H.; Shen, C. Intratympanic injection of shRNA-expressing lentivirus causes gene silencing in the inner ear in chicken. Neurosci. Lett. 2012, 510, 132–137. [Google Scholar] [CrossRef]
- Murillo-Cuesta, S.; Vallecillo, N.; Cediel, R.; Celaya, A.M.; Lassaletta, L.; Varela-Nieto, I.; Contreras, J. A comparative study of drug delivery methods targeted to the mouse inner ear: Bullostomy versus trans-tympanic injection. JoVE J. Vis. Exp. 2017, 8, e54951. [Google Scholar]
- Pan, M.; Ni, J.; He, H.; Gao, S.; Duan, X. New paradigms on siRNA local application. BMB Rep. 2015, 48, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.H.; Cai, Q.; Manohar, S.; Jiang, H.; Ding, D.; Coling, D.E.; Zheng, G.; Salvi, R. Differential expression of apoptosis-related genes in the cochlea of noise-exposed rats. Neuroscience 2009, 161, 915–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.H.; Cai, Q. Acoustic overstimulation modifies Mcl-1 expression in cochlear sensory epithelial cells. J. Neurosci. Res. 2010, 88, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.; Hu, Z.; Bard, J.; Jamison, J.; Cai, Q.; Hu, B.H. Transcriptome characterization by RNA-Seq reveals the involvement of the complement components in noise-traumatized rat cochleae. Neuroscience 2013, 248, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C.; Brito Neto, R.V. RNA extraction from wistar rat cochlea for qRT-PCR. Bio-Protoc. 2017, Bio101, e2621. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR andhe 2 (-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101. [Google Scholar] [CrossRef]
- Yang, W.P.; Henderson, D.; Hu, B.H.; Nicotera, T.M. Quantitative analysis of apoptotic and necrotic outer haircells after exposure to different levels of continuous noise. Hear. Res. 2004, 196, 69–76. [Google Scholar] [CrossRef]
- Kirkwood, B.R.; Sterne, J.A.C. Essential Medical Statistics., 2nd ed.; Blackwell Science: Malden, MA, USA, 2006. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fold Change Tnfα siRNA/Scramble | Fold Change Noise/No Noise | Fold Chang eTnfα siRNA/Scramble | Fold Change Noise/No Noise | ||
|---|---|---|---|---|---|
| Bcl Family Intrinsic pathway | Caspase Family | ||||
| Bcl2 | −1.73 | −1.64 | Casp 3 | −1.26 | 1.24 |
| BID | −1.57 | 1.22 | Casp 8 | −1.64 | 1.30 |
| Cytochrome c | −1.18 | −1.36 | |||
| Apaf 1 | −1.09 | −1.19 | TNF ligand Family | ||
| Cd40L | −7.63 | −1.75 | |||
| TNF receptors Family | Tnfa | −3.85 | 2.065 | ||
| Cd27 | −4.18 | −1.46 | Tnfsf4 | −1.18 | −1.23 |
| Cd40 | −2.61 | 1.25 | Tnfsf9 | −2.36 | 1.03 |
| Lta | −2.54 | 1.17 | Tnfsf10 | 1.23 | −1.89 |
| Ltb | −1.96 | −1.56 | Tnfsf11 | −2.11 | −1.03 |
| Ltbr | 1.19 | −1.08 | Tnfsf12 | −2.88 | 1.0 |
| Tnfrsf1A | −1.52 | 1.18 | Tnfsf13 | −2.5 | −1.31 |
| Tnfrsf1B | −5.18 | −1.15 | Tnfsf14 | −3.5 | 1.549 |
| Tnfrsf4 | −2.84 | 1.09 | Tnfsf15 | −1.21 | −1.38 |
| Tnfrsf8 | −4.07 | 1.18 | Tnfsf18 | −1.8 | −1.70 |
| Tnfrsf9 | −1.49 | −1.15 | FasL | −1.15 | −1.24 |
| Tnfrsf10B | −1.48 | −1.05 | |||
| Tnfrsf11B | −1.39 | −1.09 | Traf Family | ||
| Tnfrsf12A | −1.56 | 1.1 | Traf2 | 2.293 | −1.28 |
| Tnfrsf14 | −4.84 | −1.36 | Traf3 | 1.03 | −1.29 |
| Tnfrsf 17 | 2.09 | −5.02 | Traf4 | 1.048 | −1.2 |
| Tnfrsf21 | 1.16 | −1.01 | Traf6 | −1.03 | −1.19 |
| Tnfrsf25 | 1.47 | −1.09 | Traf7 | 1.142 | −1.07 |
| Fas | −2.68 | 1.04 | |||
| Fbf1 | 1.61 | −1.17 | |||
| Death domain Family | MapK Family | ||||
| Fadd | 1.65 | −1.43 | Map3K1 | 1.253 | 4.52 |
| Tradd | 1.35 | −1.53 | Map2K4 | 1.263 | −1.23 |
| Map3K7 | 1.011 | 4.52 | |||
| NfkB Family | Mapk8 | −1.07 | −1.38 | ||
| NfkB1 | −1.3 | −1.31 | Map3k14 | 1.18 | −5.02 |
| NfkBia | −2.51 | −2.60 | Jun | 1.07 | 1.07 |
| Ikbkg | 1.11 | −1.10 | |||
| Ikbkb | 1.02 | −1.10 | Anti-apoptotic genes | ||
| Ikbkap | 1.158 | −1.19 | Pak1 | 8.640 | −1.05 |
| Tradd | 1.35 | −1.53 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, J.C.; Bachi, A.L.L.; Silva, G.A.V.; Rossi, M.; do Amaral, J.B.; Lezirovitz, K.; de Brito, R. New Insights on the Effect of TNF Alpha Blockade by Gene Silencing in Noise-Induced Hearing Loss. Int. J. Mol. Sci. 2020, 21, 2692. https://doi.org/10.3390/ijms21082692
Rodrigues JC, Bachi ALL, Silva GAV, Rossi M, do Amaral JB, Lezirovitz K, de Brito R. New Insights on the Effect of TNF Alpha Blockade by Gene Silencing in Noise-Induced Hearing Loss. International Journal of Molecular Sciences. 2020; 21(8):2692. https://doi.org/10.3390/ijms21082692
Chicago/Turabian StyleRodrigues, Janaína C., André L. L. Bachi, Gleiciele A. V. Silva, Marcelo Rossi, Jonatas B. do Amaral, Karina Lezirovitz, and Rubens de Brito. 2020. "New Insights on the Effect of TNF Alpha Blockade by Gene Silencing in Noise-Induced Hearing Loss" International Journal of Molecular Sciences 21, no. 8: 2692. https://doi.org/10.3390/ijms21082692