The Temperature-Dependent Retention of Introns in GPI8 Transcripts Contributes to a Drooping and Fragile Shoot Phenotype in Rice


Abstract
:1. Introduction
2. Results
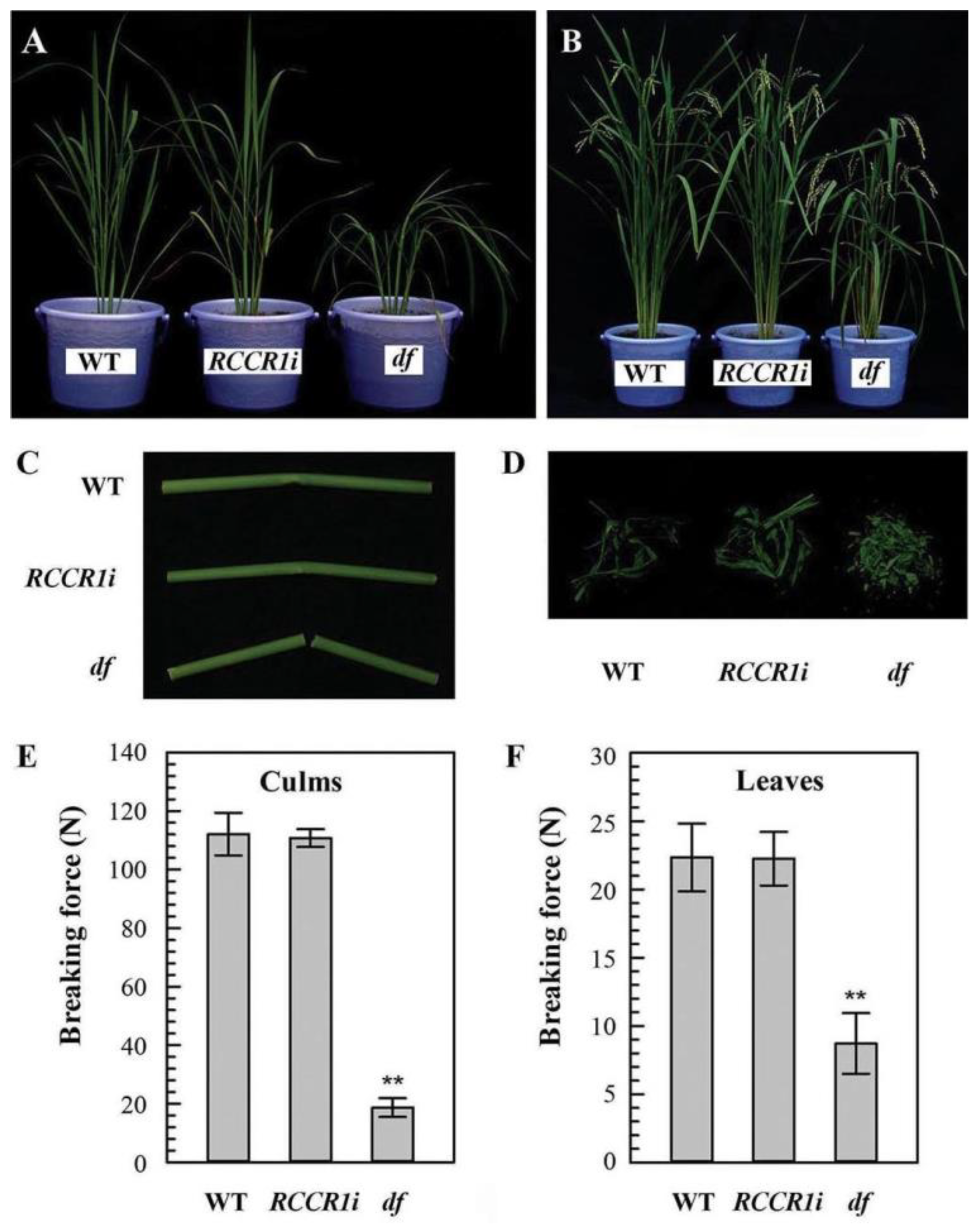
2.1. Isolation of the Drooping and Fragile Shoot Rice Mutant
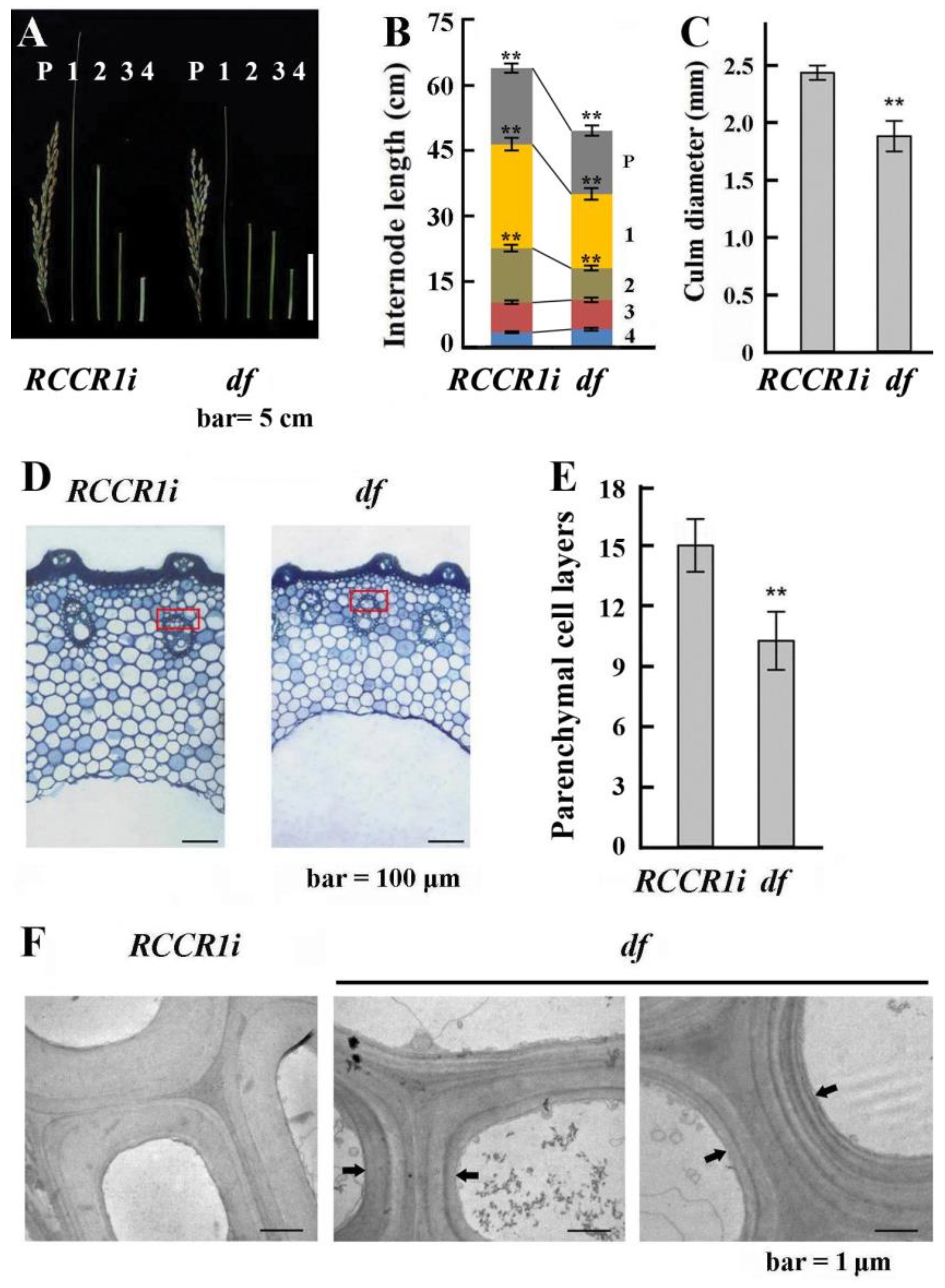
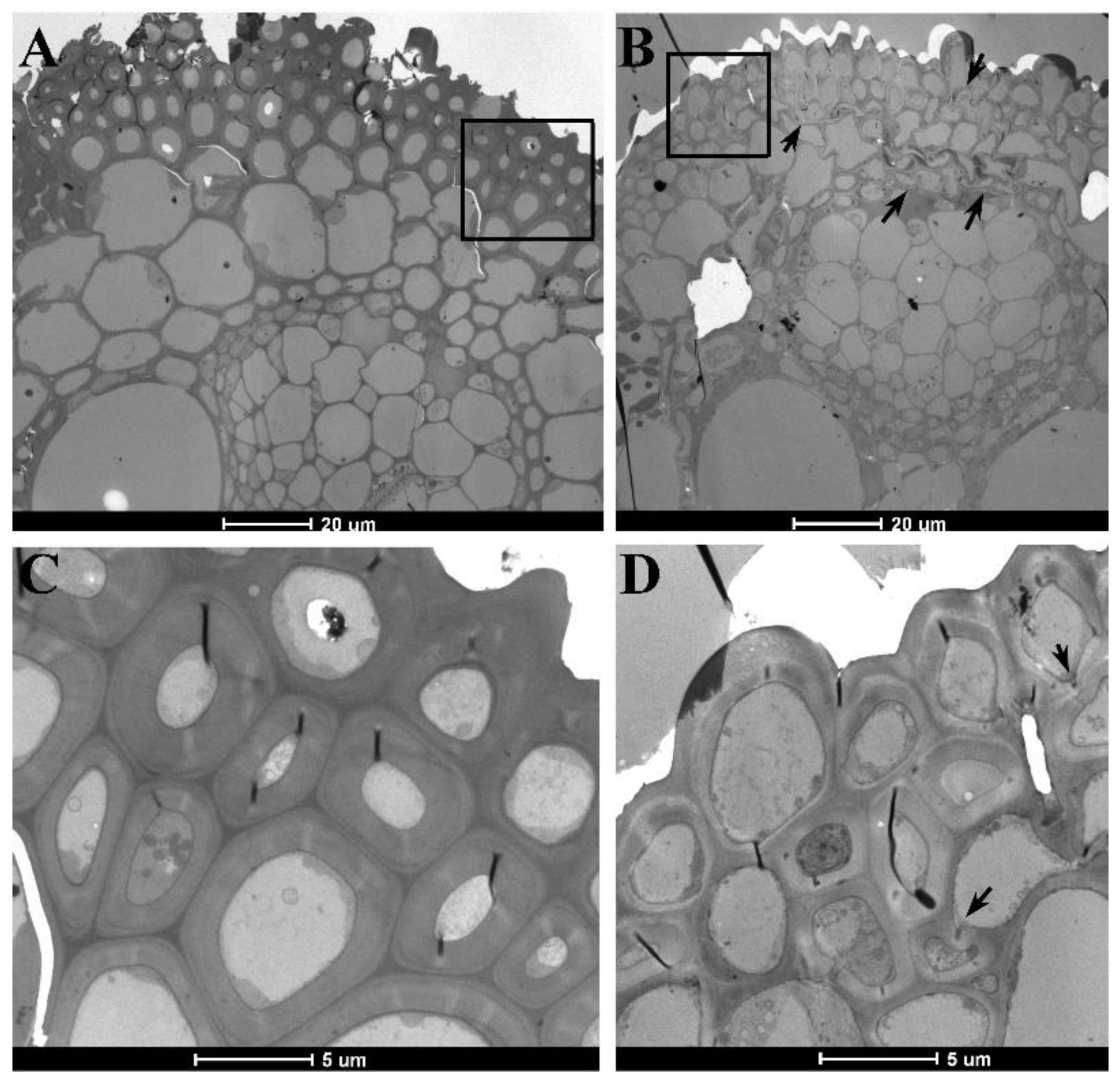
2.2. Changes in Morphological and Anatomical Structures of Culms
2.3. Changes in the Morphological and Anatomical Structures of Leaves
2.4. Differences in Cell Wall Composition
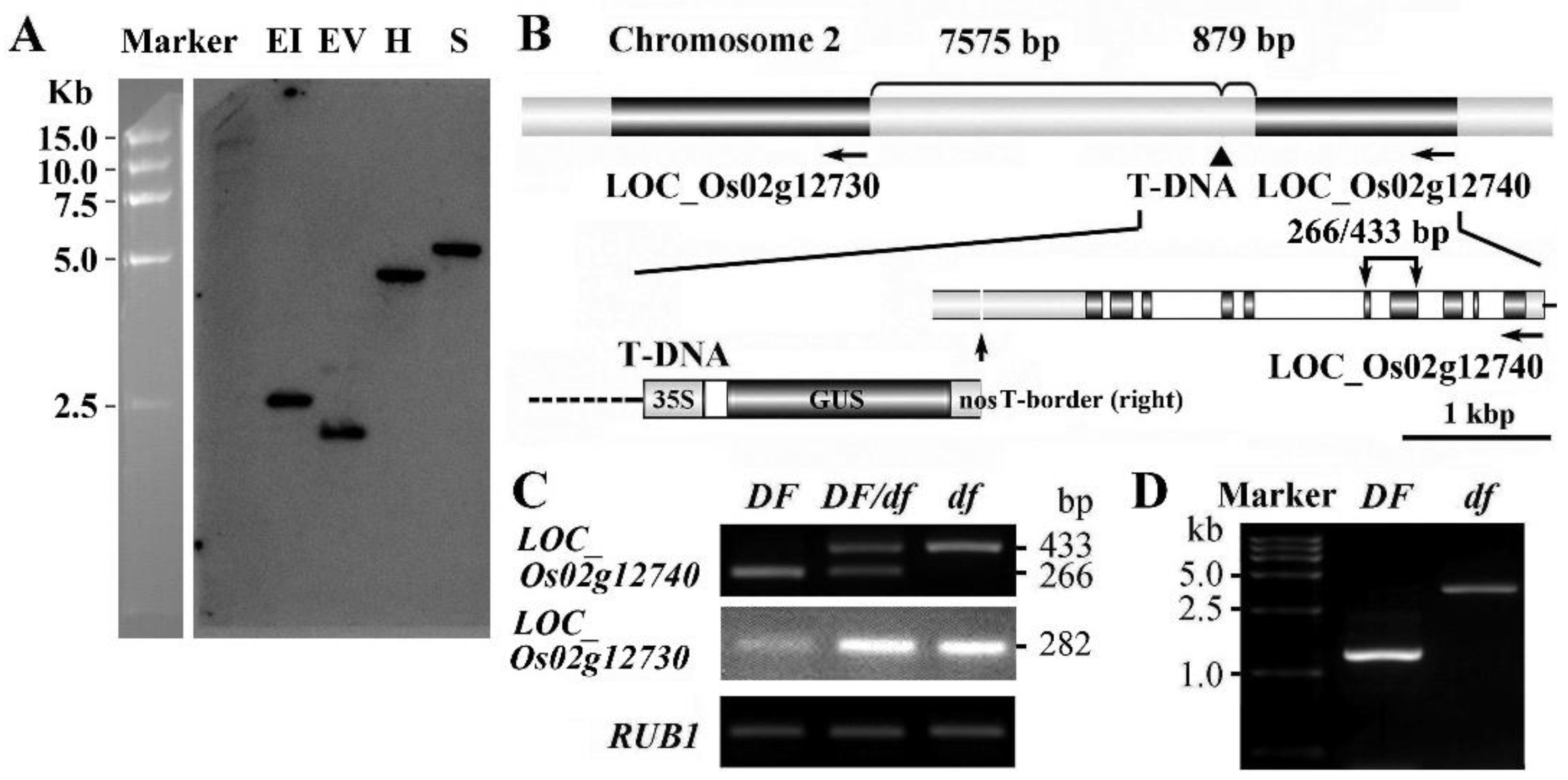
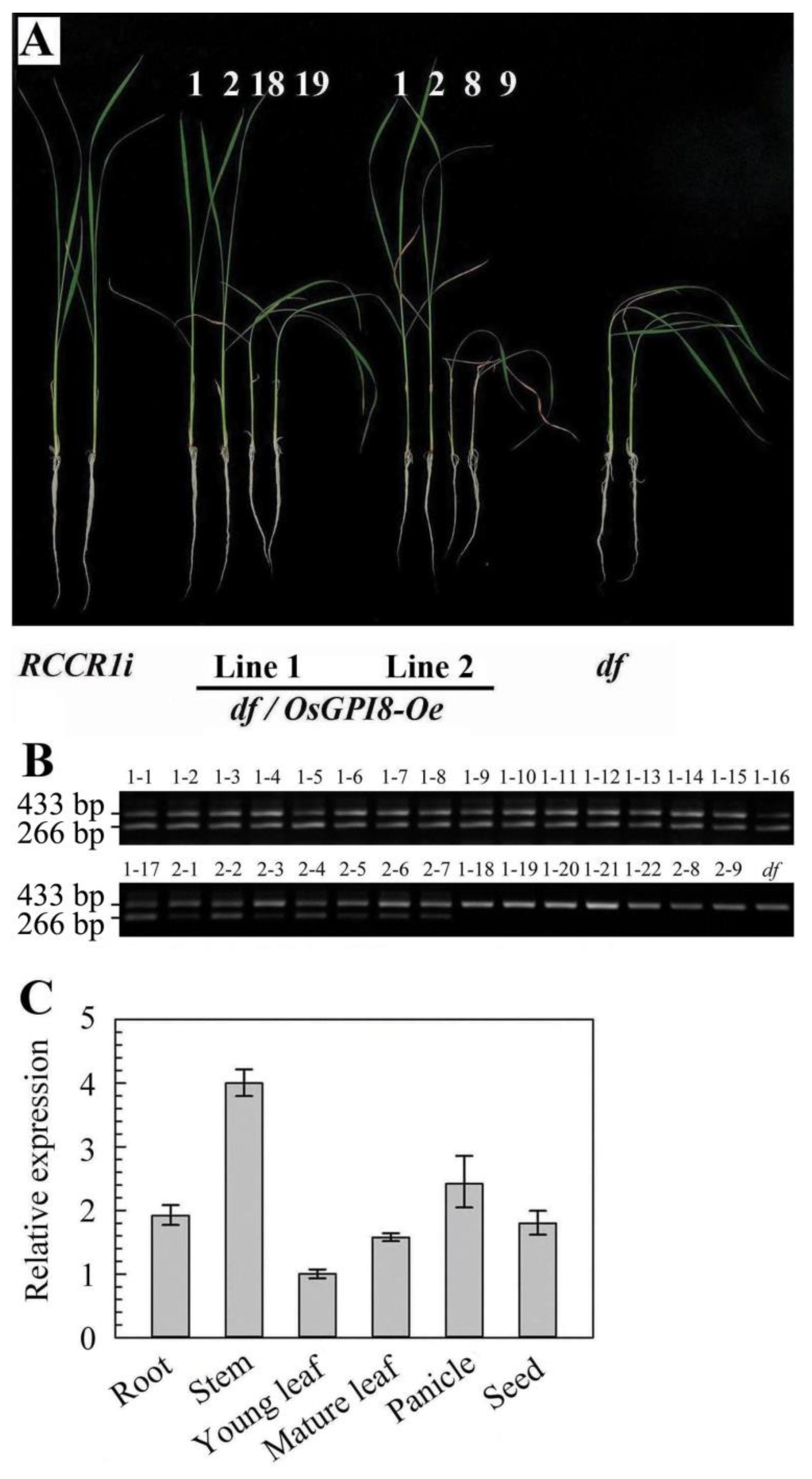
2.5. Cloning of the Gene
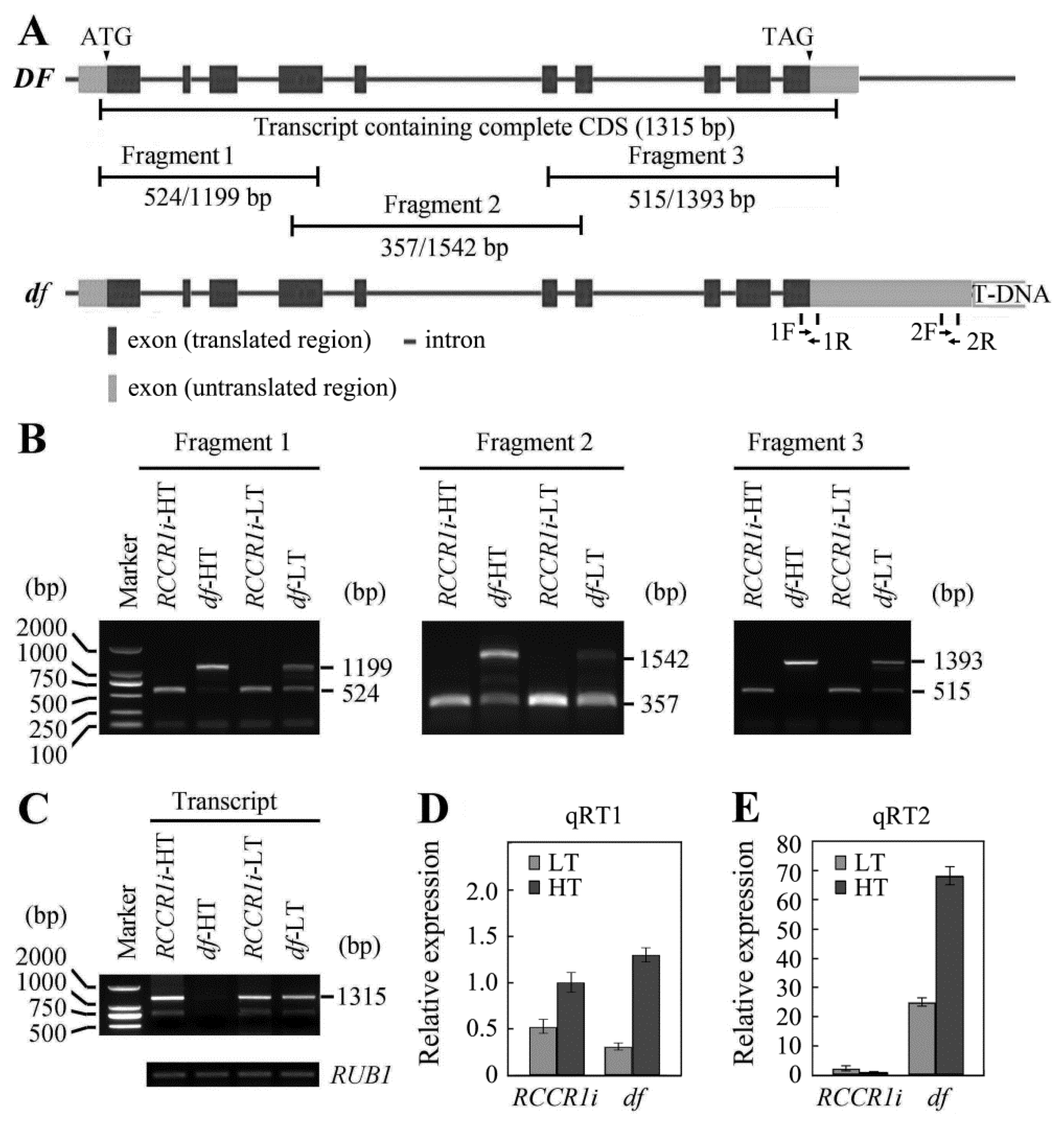
2.6. Temperature Influences the Pre-mRNA Processing of OsGPI8 Transcripts in df Plants
2.7. Change in Length of 3′-UTR of OsGPI8 Transcripts
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Cloning of the df Gene and Complementation Test
4.3. RNA Isolation and Expression Analysis
4.4. Southern Blot Analysis
4.5. Mechanical Strength Test
4.6. Microscopic Analysis
4.7. 3′-RACE PCR
4.8. Cell Wall Component Measurement
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AIR | Lcohol insoluble residue |
| At | Arabidopsis thaliana |
| CaMV | Cauliflower mosaic virus |
| df | Drooping and fragile |
| EtNP | Ethanolamine phosphate |
| GC-MS | Gas Chromatography-Mass Spectrometer |
| GlcA | Glucosamine |
| GNA | Glucosamine-6-P acetyltransferase |
| GPI | Glycosylphosphatidylinositol |
| GPI-Aps | GPI anchor proteins |
| GPI-T | GPI transamidase |
| GPIs | Glycosylphosphatidylinositols |
| HT | High temperature |
| LT | Low temperature |
| Os | Oryza sativa |
| PCR | Polymerase Chain Reaction |
| RACE | Rapid amplification of cDNA ends |
| RCCR | Red chlorophyll catabolite reductase |
| SD | Standard deviation |
| TAIL-PCR | Thermal asymmetric interlaced PCR |
| UDP-GlcNAc | UDP-N-acetylglucosamine |
| 3′-UTR | 3′-Untranslated region |
References
- Eisenhaber, B.; Bork, P.; Eisenhaber, F. Post-translational GPI lipid anchor modification of proteins in kingdoms of life: Analysis of protein sequence data from complete genomes. Protein Eng. 2001, 14, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, M.A.J. The structure, biosynthesis and functions of glycosylphosphatidylinositol anchors, and the contributions of trypanosome research. J. Cell Sci. 1999, 112, 2799–2809. [Google Scholar] [PubMed]
- Kinoshita, T. Enzymatic mechanism of GPI anchor attachment clarified. Cell Cycle 2014, 13, 1838–1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenhaber, B.; Eisenhaber, S.; Kwang, T.Y.; Grüber, G.; Eisenhaber, F. Transamidase subunit GAA1/GPAA1 is a M28 family metallo-peptidesynthetase that catalyzes the peptide bond formation between the substrate protein’s omega-site and the GPI lipid anchor’s phosphoethanolamine. Cell Cycle 2014, 13, 1912–1917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacks, M.; Garg, A.N. Recent developments in the molecular, biochemical and functional characterization of GPI8 and the GPI-anchoring mechanism. Mol. Membr. Biol. 2006, 23, 209–225. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Inoue, N.; Westfall, B.; Taron, C.H.; Orlean, P.; Takeda, J.; Kinoshita, T. The first step of glycosylphosphatidylinositol biosynthesis is mediated by a complex of PIG-A, PIG-H, PIG-C and GPI1. EMBO J. 1998, 17, 877–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Wang, S.; Dang, L.; Wang, S.; Chen, H.; Wu, Y.; Jiang, X.; Wu, P. A novel short-root gene encodes a glucosamine-6-phosphate acetyltransferase required for maintaining normal root cell shape in rice. Plant Physiol. 2005, 138, 232–242. [Google Scholar] [CrossRef] [Green Version]
- Nozaki, M.; Sugiyama, M.; Duan, J.; Uematsu, H.; Genda, T.; Sato, Y. A missense mutation in the glucosamine-6-phosphate N-acetyltransferase-encoding gene causes temperature-dependent growth defects and ectopic lignin deposition in Arabidopsis. Plant Cell 2012, 24, 3366–3379. [Google Scholar] [CrossRef] [Green Version]
- Lalanne, E.; Honys, D.; Johnson, A.; Borner, G.H.; Lilley, K.S.; Dupree, P.; Grossniklaus, U.; Twell, D. SETH1 and SETH2, two components of the glycosylphosphatidylinositol anchor biosynthetic pathway, are required for pollen germination and tube growth in Arabidopsis. Plant Cell 2004, 16, 229–240. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.R.; Gao, X.Q.; Chen, G.H.; Tang, L.L.; Wang, H.; Zhang, X.S. ABNORMAL POLLEN TUBE GUIDANCE1, an endoplasmic reticulum-localized mannosyltransferase homolog of GLYCOSYLPHOSPHATIDYLINOSITOL10 in yeast and PHOSPHATIDYLINOSITOL GLYCAN ANCHOR BIOSYNTHESIS B in human, is required for arabidopsis pollen tube micropylar guidance and embryo development. Plant Physiol. 2014, 165, 1544–1556. [Google Scholar] [CrossRef]
- Gillmor, C.S.; Lukowitz, W.; Brininstool, G.; Sedbrook, J.C.; Hamann, T.; Poindexter, P.; Somerville, C. Glycosylphosphatidylinositol-anchored proteins are required for cell wall synthesis and morphogenesis in Arabidopsis. Plant Cell 2005, 17, 1128–1140. [Google Scholar] [CrossRef] [PubMed]
- Bundy, M.G.R.; Kosentka, P.Z.; Willet, A.H.; Zhang, L.; Miller, E.; Shpak, E.D. A mutation in the catalytic subunit of the glycosylphosphatidylinositol transamidase disrupts growth, fertility, and stomata formation. Plant Physiol. 2016, 171, 974–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borner, G.H.; Lilley, K.S.; Stevens, T.J.; Dupree, P. Identification of glycosylphosphatidylinositol-anchored proteins in Arabidopsis. A proteomic and genomic analysis. Plant Physiol. 2003, 132, 568–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelman, G.; Morikami, A.; Jung, J.; Baskin, T.I.; Carpita, N.C.; Derbyshire, P.; McCann, M.C.; Benfey, P.N. COBRA encodes a putative GPI-anchored protein, which is polarly localized and necessary for oriented cell expansion in Arabidopsis. Genes Dev. 2001, 15, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Shi, H.; Kim, Y.; Guo, Y.; Stevenson, B.; Zhu, J. The Arabidopsis SOS5 locus encodes a putative cell surface adhesion protein and is required for normal cell expansion. Plant Cell 2003, 15, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Ishii, T.; Matsunaga, T.; Tominaga, R.; Kuromori, T.; Wada, T.; Shinozaki, K.; Hirayama, T. The glycerophosphoryl diester phosphodiesterase-like proteins SHV3 and its homologs play important roles in cell wall organization. Plant Cell Physiol. 2008, 49, 1522–1535. [Google Scholar] [CrossRef] [Green Version]
- MacMillan, C.P.; Mansfield, S.D.; Stachurski, Z.H.; Evans, R.; Southerton, S.G. Fasciclin-like arabinogalactan proteins: Specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus. Plant J. 2010, 62, 689–703. [Google Scholar] [CrossRef]
- Sedbrook, J.C.; Carroll, K.L.; Hung, K.F.; Masson, P.H.; Somerville, C.R. The Arabidopsis SKU5 gene encodes an extracellular glycosyl phosphatidylinositol-anchored glycoprotein involved in directional root growth. Plant Cell 2002, 14, 1635–1648. [Google Scholar] [CrossRef] [Green Version]
- Coimbra, S.; Costa, M.; Jones, B.; Mendes, M.A.; Pereira, L.G. Pollen grain development is compromised in Arabidopsis agp6 agp11 null mutants. J. Exp. Bot. 2009, 60, 3133–3142. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Zhang, Y.; Wang, W.; Zhao, K.; Liu, C.; Bai, L.; Li, R.; Guo, Y. Two membrane-anchored aspartic proteases contribute to pollen and ovule development. Plant Physiol. 2017, 173, 219–239. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Guo, X.; Cyprys, P.; Zhang, Y.; Bleckmann, A.; Cai, L.; Huang, Q.; Luo, Y.; Gu, H.; Dresselhaus, T.; et al. Maternal ENODLs are required for pollen tube reception in Arabidopsis. Curr. Biol. 2016, 26, 2343–2350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capron, A.; Gourgues, M.; Neiva, L.S.; Faure, J.E.; Berger, F.; Pagnussat, G.; Krishnan, A.; Alvarez-Mejia, C.; Vielle-Calzada, J.P.; Lee, Y.R.; et al. Maternal control of male-gamete delivery in Arabidopsis involves a putative GPI-anchored protein encoded by the LORELEI gene. Plant Cell 2008, 20, 3038–3049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeats, T.H.; Bacic, A.; Johnson, K.L. Plant glycosylphosphatidylinositol anchored proteins at the plasma membrane-cell wall nexus. J. Integr. Plant Biol. 2018, 60, 649–669. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, Q.; Zhou, Y.; Yan, M.; Sun, L.; Zhang, M.; Fu, Z.; Wang, Y.; Han, B.; Pang, X.; et al. BRITTLE CULM1, which encodes a COBRA-like protein, affects the mechanical properties of rice plants. Plant Cell 2003, 15, 2020–2031. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; You, C.; Chen, G.; Li, X.; Zhang, Q.; Wu, C. OsBC1L4 encodes a COBRA-like protein that affects cellulose synthesis in rice. Plant Mol. Biol. 2011, 75, 333–345. [Google Scholar] [CrossRef]
- Xiang, J.; Zhang, G.; Qian, Q.; Xue, H. SEMI-ROLLED LEAF1 encodes a putative glycosylphosphatidylinositol-anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells. Plant Physiol. 2012, 159, 1488–1500. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Zhang, M.; Gan, P.; Qiao, L.; Yang, S.; Miao, H.; Wang, G.; Zhang, M.; Liu, W.; Li, H.; et al. CLD1/SRL1 modulates leaf rolling by affecting cell wall formation, epidermis integrity and water homeostasis in rice. Plant J. 2017, 92, 904–923. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Li, M.; Chen, Y.; Wu, P.; Wu, G.; Jiang, H. Knockdown of OsPAO and OsRCCR1 cause different plant death phenotypes in rice. J. Plant Physiol. 2011, 168, 1952–1959. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, Y. High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences. Biotechniques 2007, 43, 649–656. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Li, Q.; Liu, Z.; Lu, W.; Liu, M. Interplay between alternative splicing and alternative polyadenylation defines the expression outcome of the plant unique OXIDATIVE TOLERANT-6 gene. Sci. Rep. 2017, 7, 2052. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Brendel, V. Genomewide comparative analysis of alternative splicing in plants. Proc. Natl. Acad. Sci. USA 2006, 103, 7175–7180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, A.S.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the alternative splicing landscape in plants. Plant Cell 2013, 25, 3657–3683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, A.B.; Syed, N.H.; Bordage, S.; Marshall, J.; Nimmo, G.A.; Jenkins, G.I.; Herzyk, P.; Brown, J.W.; Nimmo, H.G. Alternative splicing mediates responses of the Arabidopsis circadian clock to temperature changes. Plant Cell 2012, 24, 961–981. [Google Scholar] [CrossRef] [Green Version]
- Capovilla, G.; Pajoro, A.; Immink, R.G.; Schmid, M. Role of alternative pre-mRNA splicing in temperature signaling. Curr. Opin. Plant Biol. 2015, 27, 97–103. [Google Scholar] [CrossRef]
- Proudfoot, N.J.; Furger, A.; Dye, M.J. Integrating mRNA processing with transcription. Cell 2002, 108, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Ullah, F.; Hamilton, M.; Reddy, A.S.; Ben-Hur, A. Exploring the relationship between intron retention and chromatin accessibility in plants. BMC Genomics 2018, 19, 21. [Google Scholar] [CrossRef] [Green Version]
- Mazo, A.; Hodgson, J.W.; Petruk, S.; Sedkov, Y.; Brock, H.W. Transcriptional interference: An unexpected layer of complexity in gene regulation. J. Cell Sci. 2007, 120, 2755–2761. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Suzuki, R.; Nishikubo, N.; Takenouchi, S.; Ito, S.; Nakano, Y.; Nakaba, S.; Sano, Y.; Funada, R.; Kajita, S.; et al. Isolation of a novel cell wall architecture mutant of rice with defective Arabidopsis COBL4 ortholog BC1 required for regulated deposition of secondary cell wall components. Planta 2010, 232, 257–270. [Google Scholar] [CrossRef]
- Liu, L.; Shang-Guan, K.; Zhang, B.; Liu, X.; Yan, M.; Zhang, L.; Shi, Y.; Zhang, M.; Qian, Q.; Li, J.; et al. Brittle Culm1, a COBRA-like protein, functions in cellulose assembly through binding cellulose microfibrils. PLoS Genet. 2013, 9, e1003704. [Google Scholar] [CrossRef]
- Dai, X.; You, C.; Wang, L.; Chen, G.; Zhang, Q.; Wu, C. Molecular characterization, expression pattern, and function analysis of the OsBC1L family in rice. Plant Mol. Biol. 2009, 71, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Vaddepalli, P.; Fulton, L.; Wieland, J.; Wassmer, K.; Schaeffer, M.; Ranf, S.; Schneitz, K. The cell wall-localized atypical β-1,3 glucanase ZERZAUST controls tissue morphogenesis in Arabidopsis thaliana. Development 2017, 144, 2259–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, H.; Wang, Y.; Cloix, C.; Li, K.; Jenkins, G.I.; Wang, S.; Shang, Z.; Shi, Y.; Yang, S.; Li, X. The Arabidopsis RCC1 family protein TCF1 regulates freezing tolerance and cold acclimation through modulating lignin biosynthesis. PLoS Genet. 2015, 11, e1005471. [Google Scholar] [CrossRef] [PubMed]
- Nieuwland, J.; Feron, R.; Huisman, B.A.; Fasolino, A.; Hilbers, C.W.; Derksen, J.; Mariani, C. Lipid transfer proteins enhance cell wall extension in tobacco. Plant Cell 2005, 17, 2009–2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Sardar, H.S.; McGovern, K.R.; Zhang, Y.; Showalter, A.M. A lysine-rich arabinogalactan protein in Arabidopsis is essential for plant growth and development, including cell division and expansion. Plant J. 2007, 49, 629–640. [Google Scholar] [CrossRef]
- Yoshida, S.; Forno, D.A.; Cock, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd ed.; International Rice Research Institute: Manila, Philippines, 1976; pp. 61–64. [Google Scholar]
- Harholt, J.; Jensen, J.K.; Sorensen, S.O.; Orfila, C.; Pauly, M.; Scheller, H.V. ARABINAN DEFICIENT 1 is a putative arabinosyltransferase involved in biosynthesis of pectic arabinan in Arabidopsis. Plant Physiol. 2006, 140, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Xiong, G.; Li, R.; Cui, J.; Tang, D.; Zhang, B.; Pauly, M.; Cheng, Z.; Zhou, Y. Rice cellulose synthase-like D4 is essential for normal cell-wall biosynthesis and plant growth. Plant J. 2009, 60, 1055–1069. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Zhou, Y. Carbohydrate composition analysis in xylem. Methods Mol. Biol. 2017, 1544, 213–222. [Google Scholar] [CrossRef]
- Updegraff, D.M. Semimicro determination of cellulose in biological materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Fukushima, R.S.; Hatfield, R.D. Extraction and isolation of lignin for utilization as a standard to determine lignin concentration using the acetyl bromide spectrophotometric method. J. Agric. Food Chem. 2001, 49, 3133–3139. [Google Scholar] [CrossRef]
- Köhle, H.; Jeblick, W.; Blaschek, F.; Kauss, H. Chitosan-elicited callose synthesis in soybean cells as a Ca2+-dependent process. Plant Physiol. 1985, 77, 544–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, D.B. Multiple range and multiple F tests. Int. Biom. Soc. 1955, 11, 1–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound (μg mg−1 AIR) | Culms RCCR1i | RCCR1i df | Leaf Blades RCCR1i | Leaf Blades df |
|---|---|---|---|---|
| Cellulose | 453.74 ± 8.13 | 331.9 3 ± 4.79 ** | 326.91 ± 5.62 | 221.21 ± 7.06 ** |
| Lignin | 142.41 ± 3.84 | 104.45 ± 1.01 ** | 98.72 ± 0.99 | 85.54 ± 1.42 ** |
| Neutral Sugars | ||||
| Rhamnose | 2.15 ± 0.08 | 2.44 ± 0.05 * | 2.76 ± 0.09 | 2.97 ± 0.08 |
| Fucose | 1.20 ± 0.02 | 1.18 ± 0.02 | 1.41 ± 0.04 | 1.51 ± 0.01 |
| Arabinose | 32.58 ± 0.70 | 34.97 ± 0.65 * | 41.79 ± 0.88 | 45.40 ± 0.79 * |
| Xylose | 212.59 ± 3.74 | 317.89 ± 5.00 ** | 219.57 ± 4.83 | 229.71 ± 3.58 * |
| Mannose | 2.19 ± 0.06 | 2.16 ± 0.04 | 2.56 ± 0.07 | 2.79 ± 0.03 |
| Galactose | 24.24 ± 0.296 | 19.89 ± 0.27 ** | 17.10 ± 0.30 | 18.56 ± 0.09 ** |
| Glucose | 135.75 ± 0.83 | 87.60 ± 1.58 ** | 27.86 ± 0.52 | 26.11 ± 0.31 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, B.; Tang, Y.; Zhang, B.; Wu, P.; Li, M.; Xu, X.; Wu, G.; Jiang, H.; Chen, Y. The Temperature-Dependent Retention of Introns in GPI8 Transcripts Contributes to a Drooping and Fragile Shoot Phenotype in Rice. Int. J. Mol. Sci. 2020, 21, 299. https://doi.org/10.3390/ijms21010299
Zhao B, Tang Y, Zhang B, Wu P, Li M, Xu X, Wu G, Jiang H, Chen Y. The Temperature-Dependent Retention of Introns in GPI8 Transcripts Contributes to a Drooping and Fragile Shoot Phenotype in Rice. International Journal of Molecular Sciences. 2020; 21(1):299. https://doi.org/10.3390/ijms21010299
Chicago/Turabian StyleZhao, Bo, Yongyan Tang, Baocai Zhang, Pingzhi Wu, Meiru Li, Xinlan Xu, Guojiang Wu, Huawu Jiang, and Yaping Chen. 2020. "The Temperature-Dependent Retention of Introns in GPI8 Transcripts Contributes to a Drooping and Fragile Shoot Phenotype in Rice" International Journal of Molecular Sciences 21, no. 1: 299. https://doi.org/10.3390/ijms21010299