

Pyrite-Based Autotrophic Denitrifying Microorganisms Derived from Paddy Soils: Effects of Organic Co-Substrate Addition
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Materials
2.2. Experimental Procedure
2.3. Analysis
2.4. DNA Extraction and Illumina MiSeq Sequencing
3. Results and Discussion
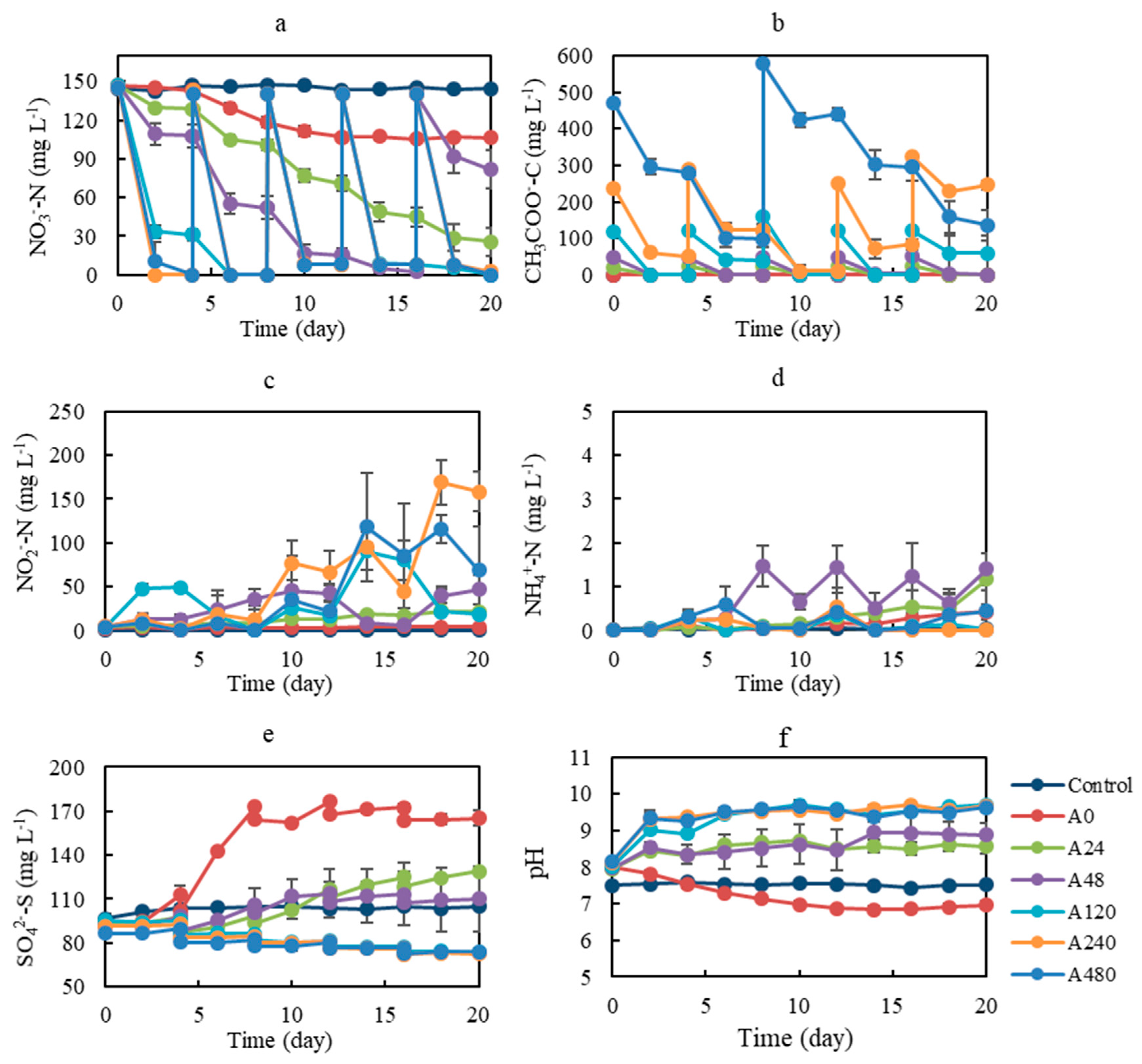
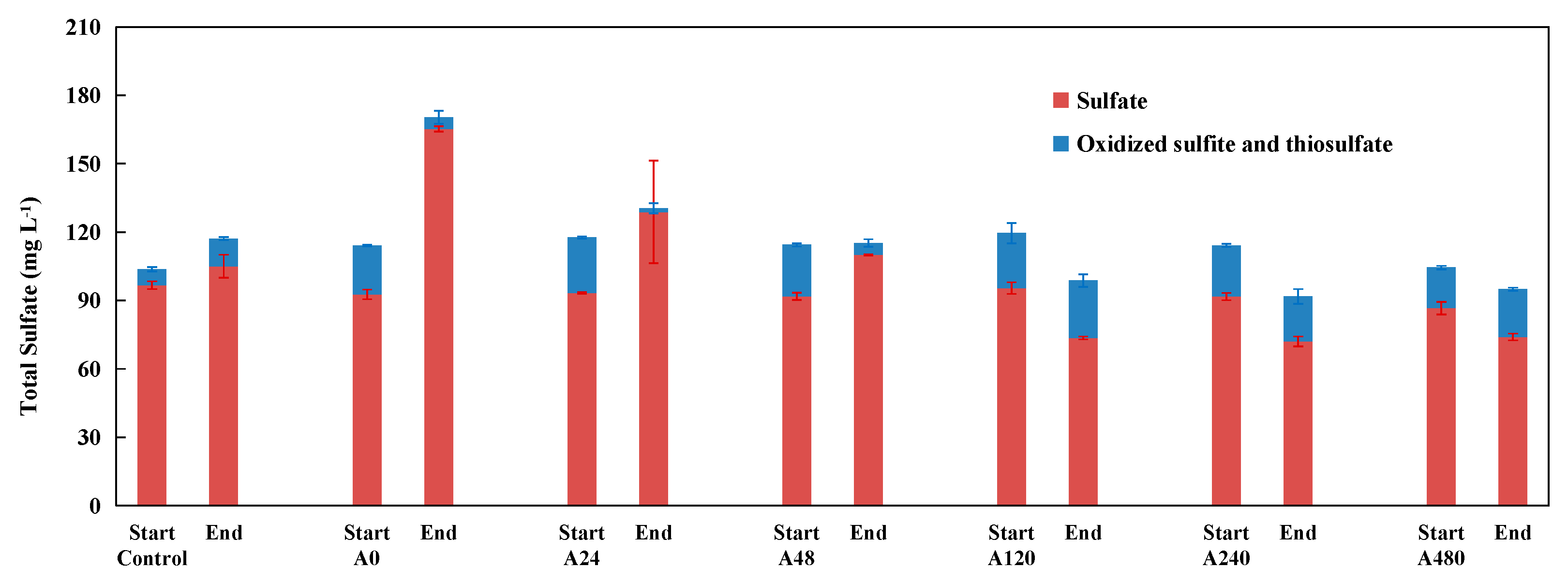
3.1. Effect of Organic Co-Substrate Addition on Denitrification Performance
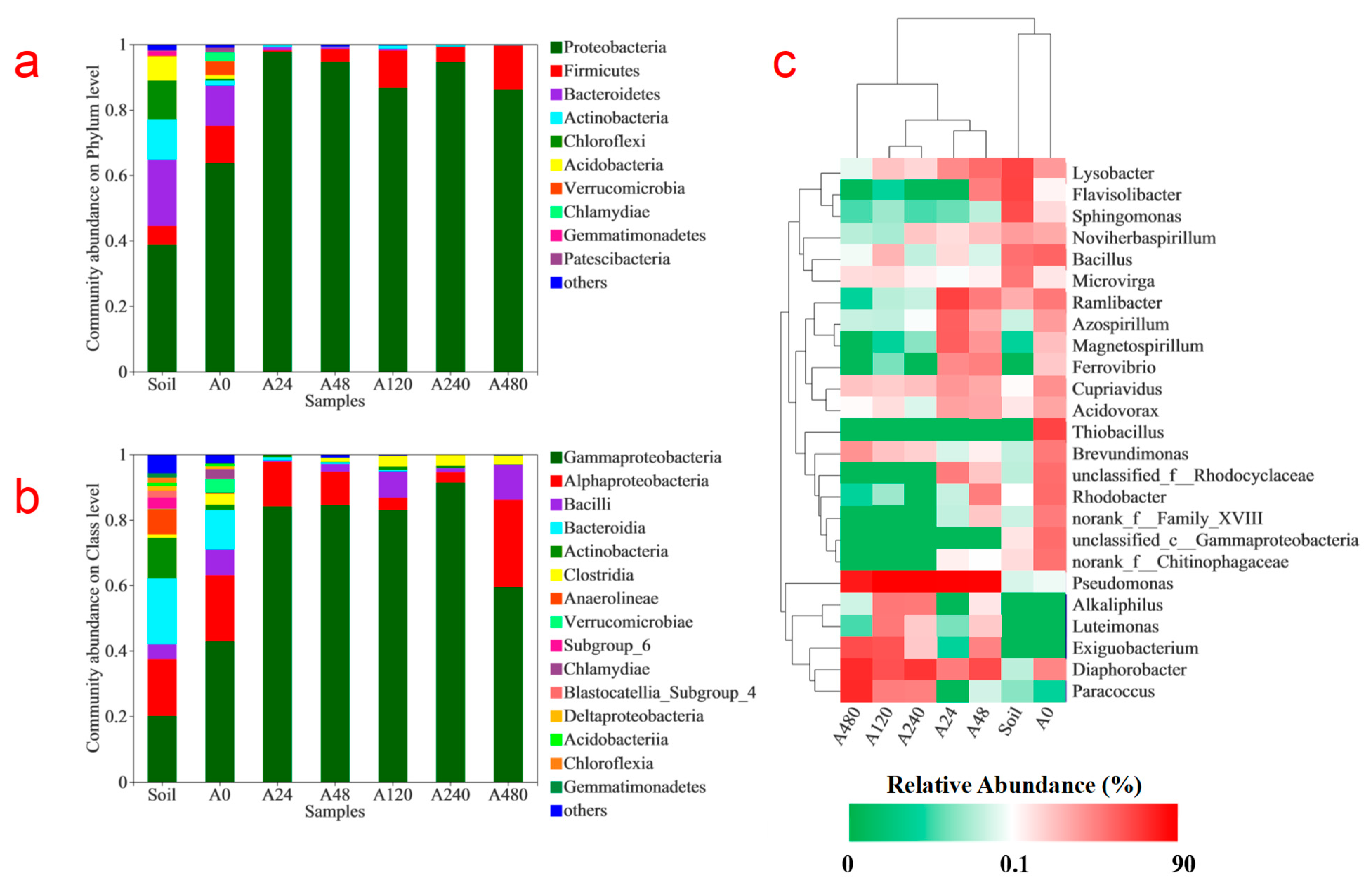
3.2. Microbial Community Analyses with the Addition of Different Concentrations of Organic Co-Substrate in the Pyrite-Based Autotrophic Denitrification System
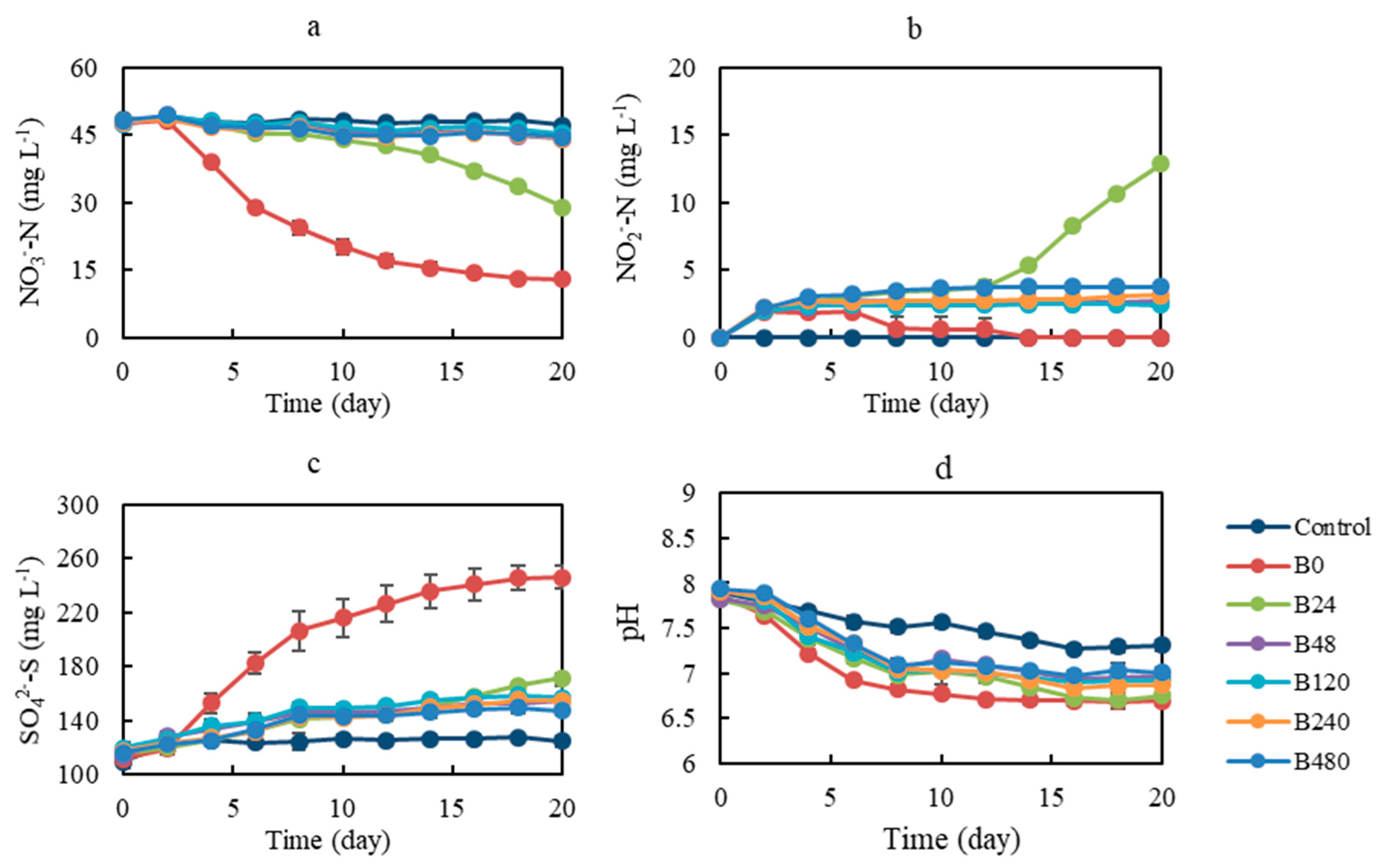
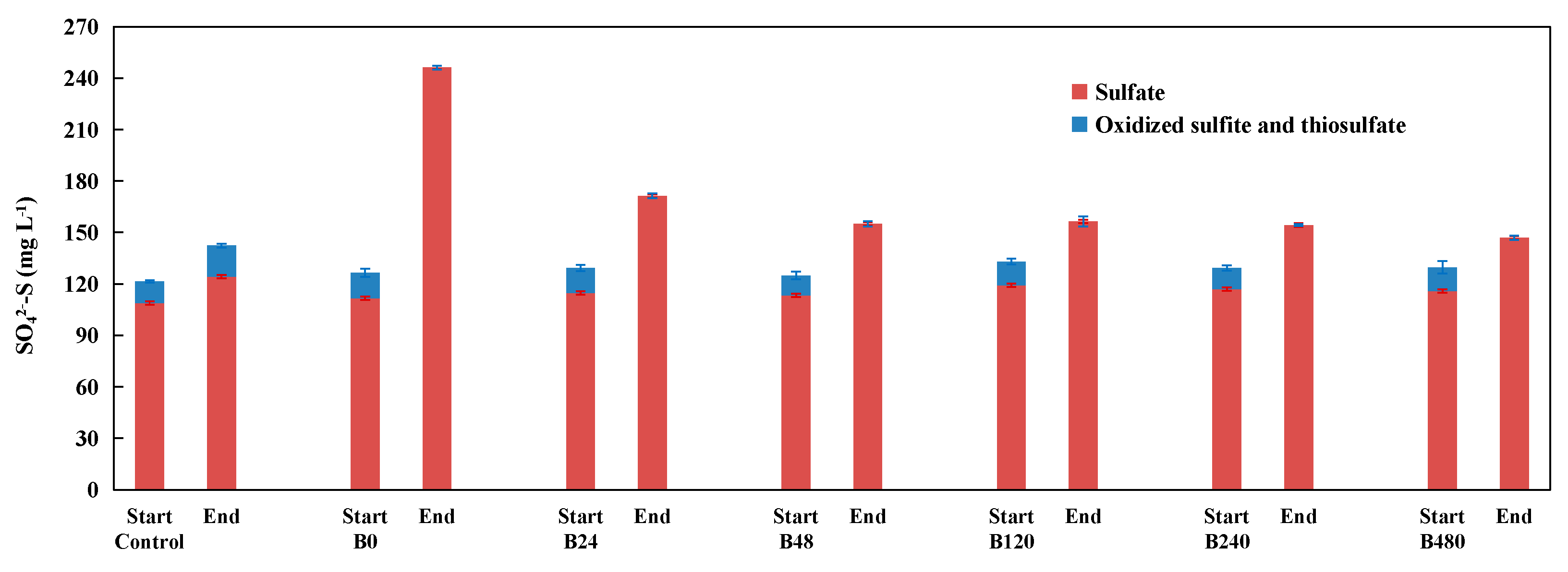
3.3. Performance of Pyrite-Based Denitrification with Inoculum from Denitrification Systems with the Addition of Different Concentrations of Organic Co-Substrate
3.4. Limitations and Outlook
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rivett, M.O.; Buss, S.R.; Morgan, P.; Smith, J.W.N.; Bemment, C.D. Nitrate attenuation in groundwater: A review of biogeochemical controlling processes. Water Res. 2008, 42, 4215–4232. [Google Scholar] [CrossRef] [PubMed]
- Ashok, V.; Hait, S. Remediation of nitrate-contaminated water by solid-phase denitrification process—A review. Env. Sci. Pollut. Res. Int. 2015, 22, 8075–8093. [Google Scholar] [CrossRef]
- Hu, Y.; Wu, G.; Li, R.; Xiao, L.; Zhan, X. Iron sulphides mediated autotrophic denitrification: An emerging bioprocess for nitrate pollution mitigation and sustainable wastewater treatment. Water Res. 2020, 179, 115914. [Google Scholar] [CrossRef]
- Li, R.; Feng, C.; Chen, N.; Zhang, B.; Hao, C.; Peng, T.; Zhu, X. A bench-scale denitrification wall for simulating the in-situ treatment of nitrate-contaminated groundwater. Ecol. Eng. 2014, 73, 536–544. [Google Scholar] [CrossRef]
- Pu, J.; Feng, C.; Liu, Y.; Li, R.; Kong, Z.; Chen, N.; Tong, S.; Hao, C.; Liu, Y. Pyrite-based autotrophic denitrification for remediation of nitrate contaminated groundwater. Bioresour. Technol. 2014, 173, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Tong, S.; Stocks, J.L.; Rodriguez-Gonzalez, L.C.; Feng, C.; Ergas, S.J. Effect of oyster shell medium and organic substrate on the performance of a particulate pyrite autotrophic denitrification (PPAD) process. Bioresour. Technol. 2017, 244, 296–303. [Google Scholar] [CrossRef]
- Xu, B.; Shi, L.; Zhong, H.; Wang, K. The performance of pyrite-based autotrophic denitrification column for permeable reactive barrier under natural environment. Bioresour. Technol. 2019, 290, 121763. [Google Scholar] [CrossRef]
- Schwientek, M.; Einsiedl, F.; Stichler, W.; St Gbauer, A.; Strauss, H.; Maloszewski, P. Evidence for denitrification regulated by pyrite oxidation in a heterogeneous porous groundwater system. Chem. Geol. 2008, 255, 60–67. [Google Scholar] [CrossRef]
- Juncher Jørgensen, C.; Jacobsen, O.S.; Elberling, B.; Aamand, J. Microbial Oxidation of Pyrite Coupled to Nitrate Reduction in Anoxic Groundwater Sediment. Environ. Sci. Technol. 2009, 43, 4851–4857. [Google Scholar] [CrossRef]
- Torrentó, C.; Cama, J.; Urmeneta, J.; Otero, N.; Soler, A. Denitrification of groundwater with pyrite and Thiobacillus denitrificans. Chem. Geol. 2010, 278, 80–91. [Google Scholar] [CrossRef] [Green Version]
- Torrentó, C.; Urmeneta, J.; Otero, N.; Soler, A.; Vi As, M.; Cama, J. Enhanced denitrification in groundwater and sediments from a nitrate-contaminated aquifer after addition of pyrite. Chem. Geol. 2011, 287, 90–101. [Google Scholar] [CrossRef]
- Kong, Z.; Li, L.; Feng, C.; Chen, N.; Dong, S.; Hu, W. Soil infiltration bioreactor incorporated with pyrite-based (mixotrophic) denitrification for domestic wastewater treatment. Bioresour. Technol. 2015, 187, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Kong, Z.; Li, L.; Feng, C.; Dong, S.; Chen, N. Comparative investigation on integrated vertical-flow biofilters applying sulfur-based and pyrite-based autotrophic denitrification for domestic wastewater treatment. Bioresour. Technol. 2016, 211, 125–135. [Google Scholar] [PubMed]
- Fei, X.; Sun, S.; He, S.; Huang, J.; Zhou, W. Application of a novel two-stage biofiltration system for simulated brackish aquaculture wastewater treatment. Environ. Sci. Pollut. Res. 2020, 27, 636–646. [Google Scholar] [CrossRef]
- Chen, Y.; Shao, Z.; Kong, Z.; Gu, L.; Fang, J.; Chai, H. Study of pyrite based autotrophic denitrification system for low-carbon source stormwater treatment. J. Water Process Eng. 2020, 37, 101414. [Google Scholar] [CrossRef]
- Zhang, W.; Huang, F.; Hu, W. Performance and mechanism of synchronous nitrate and phosphorus removal in constructed pyrite-based mixotrophic denitrification system from secondary effluent. Environ. Sci. Pollut. Res. 2020, 27, 36816–36825. [Google Scholar] [CrossRef]
- Zhao, L.; Xue, L.; Wang, L.; Liu, C.; Li, Y. Simultaneous heterotrophic and FeS2-based ferrous autotrophic denitrification process for low-C/N ratio wastewater treatment: Nitrate removal performance and microbial community analysis. Sci. Total Environ. 2022, 829, 154682. [Google Scholar] [CrossRef]
- Chu, Y.; Liu, W.; Tan, Q.; Yang, L.; Chen, J.; Ma, L.; Zhang, Y.; Wu, Z.; He, F. Vertical-flow constructed wetland based on pyrite intensification: Mixotrophic denitrification performance and mechanism. Bioresour. Technol. 2022, 347, 126710. [Google Scholar] [CrossRef]
- Weng, Z.; Ma, H.; Ma, J.; Kong, Z.; Shao, Z.; Yuan, Y.; Xu, Y.; Ni, Q.; Chai, H. Corncob-pyrite bioretention system for enhanced dissolved nutrient treatment: Carbon source release and mixotrophic denitrification. Chemosphere 2022, 306, 135534. [Google Scholar] [CrossRef]
- Pang, Y.; Hu, L.; Wang, J. Mixotrophic denitrification using pyrite and biodegradable polymer composite as electron donors. Bioresour. Technol. 2022, 351, 127011. [Google Scholar]
- Koenig, A.; Zhang, T.; Liu, L.; Fang, H.H.P. Microbial community and biochemistry process in autosulfurotrophic denitrifying biofilm. Chemosphere 2005, 58, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Straub, K.L.; Sch Nhuber, W.A.; Buchholz-Cleven, B.E.E.; Schink, B. Diversity of Ferrous Iron-Oxidizing, Nitrate-Reducing Bacteria and their Involvement in Oxygen-Independent Iron Cycling. Geomicrobiol. J. 2004, 21, 371–378. [Google Scholar] [CrossRef]
- Jamieson, J.; Prommer, H.; Kaksonen, A.H.; Sun, J.; Siade, A.J.; Yusov, A.; Bostick, B. Identifying and Quantifying the Intermediate Processes during Nitrate-Dependent Iron(II) Oxidation. Environ. Sci. Technol. 2018, 52, 5771–5781. [Google Scholar] [CrossRef]
- Xu, B.; Shi, L.; Zhong, H.; Wang, K. Investigation of Fe(II) and Mn(II) involved anoxic denitrification in agricultural soils with high manganese and iron contents. J. Soils Sediments 2021, 21, 452–468. [Google Scholar] [CrossRef]
- Chen, D.; Wei, L.; Zou, Z.; Yang, K.; Wang, H. Bacterial communities in a novel three-dimensional bioelectrochemical denitrification system: The effects of pH. Appl. Microbiol. Biotechnol. 2016, 100, 6805–6813. [Google Scholar] [CrossRef] [PubMed]
- Calderer, M.; Gibert, O.; Martí, V.; Rovira, M.; de Pablo, J.; Jordana, S.; Duro, L.; Guimerà, J.; Bruno, J. Denitrification in presence of acetate and glucose for bioremediation of nitrate-contaminated groundwater. Environ. Technol. 2010, 31, 799–814. [Google Scholar] [CrossRef]
- Glass, C.; Silverstein, J. Denitrification kinetics of high nitrate concentration water: pH effect on inhibition and nitrite accumulation. Water Res. 1998, 32, 831–839. [Google Scholar] [CrossRef]
- Ren, Y.; Yang, L.; Liang, X. The characteristics of a novel heterotrophic nitrifying and aerobic denitrifying bacterium, Acinetobacter junii YB. Bioresour. Technol. 2014, 171, 1–9. [Google Scholar]
- Qian, W.; Ma, B.; Li, X.; Zhang, Q.; Peng, Y. Long-term effect of pH on denitrification: High pH benefits achieving partial-denitrification. Bioresour. Technol. 2019, 278, 444–449. [Google Scholar] [CrossRef]
- Wang, T.; Dang, Q.; Liu, C.; Yan, J.; Fan, B.; Cha, D.; Yin, Y.; Zhang, Y. Heterotrophic nitrogen removal by a newly-isolated alkalitolerant microorganism, Serratia marcescens W5. Bioresour. Technol. 2016, 211, 618–627. [Google Scholar] [CrossRef]
- Zhang, Q.; Liu, Y.; Ai, G.; Miao, L.; Zheng, H.; Liu, Z. The characteristics of a novel heterotrophic nitrification–aerobic denitrification bacterium, Bacillus methylotrophicus strain L7. Bioresour. Technol. 2012, 108, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Wang, Y.; Chen, H.; Lyu, Y. Ammonium removal characteristics of an acid-resistant bacterium Acinetobacter sp. JR1 from pharmaceutical wastewater capable of heterotrophic nitrification-aerobic denitrification. Bioresour. Technol. 2019, 274, 56–64. [Google Scholar] [CrossRef]
- Sierra-Alvarez, R.; Beristain-Cardoso, R.; Salazar, M.; Gómez, J.; Razo-Flores, E.; Field, J.A. Chemolithotrophic denitrification with elemental sulfur for groundwater treatment. Water Res. 2007, 41, 1253–1262. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, N.; Tong, S.; Liang, J.; Yang, C.; Feng, C. Performance enhancement of H2S-based autotrophic denitrification with bio-gaseous CO2 as sole carbon source through new pH adjustment materials. J. Environ. Manag. 2020, 261, 110157. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Zhang, W.; Gao, J.; Hu, X.; Zhang, C.; He, Q.; Yang, F.; Wang, H.; Wang, X.; Zhan, X. A pilot-scale study on the treatment of landfill leachate by a composite biological system under low dissolved oxygen conditions: Performance and microbial community. Bioresour. Technol. 2020, 296, 122344. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Z. Assessing bacterial diversity in soil. J. Soils Sediments 2008, 8, 379–388. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing and clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Fierer, N.; Lauber, C.L.; Caporaso, J.G.; Knight, R.; Grogan, P. Soil bacterial diversity in the Arctic is not fundamentally different from that found in other biomes. Environ. Microbiol. 2010, 12, 2998–3006. [Google Scholar] [CrossRef]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three Genomes from the Phylum Acidobacteria Provide Insight into the Lifestyles of These Microorganisms in Soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef]
- Sun, B.; Gu, L.; Bao, L.; Zhang, S.; Wei, Y.; Bai, Z.; Zhuang, G.; Zhuang, X. Application of biofertilizer containing Bacillus subtilis reduced the nitrogen loss in agricultural soil. Soil Biol. Biochem. 2020, 148, 107911. [Google Scholar] [CrossRef]
- Zhao, Y.; Cao, X.; Song, X.; Zhao, Z.; Wang, Y.; Si, Z.; Lin, F.; Chen, Y.; Zhang, Y. Montmorillonite supported nanoscale zero-valent iron immobilized in sodium alginate (SA/Mt-NZVI) enhanced the nitrogen removal in vertical flow constructed wetlands (VFCWs). Bioresour. Technol. 2018, 267, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, Y.; Xu, X.; Sun, M.; Jiang, M.; Xue, G.; Li, X.; Liu, Z. How does iron facilitate the aerated biofilter for tertiary simultaneous nutrient and refractory organics removal from real dyeing wastewater? Water Res. 2019, 148, 344–358. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Zheng, Y.; Wu, S.; Yang, Z.; Zhao, F. Bacterial Community Structure of Autotrophic Denitrification Biocathode by 454 Pyrosequencing of the 16S rRNA Gene. Microb. Ecol. 2015, 69, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Wei, L.; Wu, Y.; Wang, Q.; Fu, X.; Zhang, X.; Chang, X.; Wang, L.; Pei, X. A comparative study on denitrifying sludge granulation with different electron donors: Sulfide, thiosulfate and organics. Chemosphere 2017, 186, 322–330. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Wang, J.; Song, Q.; Zhang, W.; He, Q.; Song, J.; Ma, F. Comparison of performance and microbial communities in a bioelectrochemical system for simultaneous denitrification and chromium removal: Effects of pH. Process Biochem. 2018, 73, 154–161. [Google Scholar] [CrossRef]
- Chen, C.; Ren, N.; Wang, A.; Yu, Z.; Lee, D. Microbial community of granules in expanded granular sludge bed reactor for simultaneous biological removal of sulfate, nitrate and lactate. Appl. Microbiol. Biotechnol. 2008, 79, 1071–1077. [Google Scholar] [CrossRef]
- Chen, D.; Wang, D.; Xiao, Z.; Wang, H.; Yang, K. Nitrate removal in a combined bioelectrochemical and sulfur autotrophic denitrification system under high nitrate concentration: Effects of pH. Bioprocess Biosyst. Eng. 2018, 41, 449–455. [Google Scholar] [CrossRef]
- Kwon, S.; Moon, E.; Kim, T.; Hong, S.; Park, H. Pyrosequencing Demonstrated Complex Microbial Communities in a Membrane Filtration System for a Drinking Water Treatment Plant. Microbes Environ. 2011, 26, 149–155. [Google Scholar] [CrossRef]
- He, Q.; Zhang, J.; Gao, S.; Chen, L.; Lyu, W.; Zhang, W.; Song, J.; Hu, X.; Chen, R.; Wang, H.; et al. A comprehensive comparison between non-bulking and bulking aerobic granular sludge in microbial communities. Bioresour. Technol. 2019, 294, 122151. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, C.; Hu, C.; Liu, H.; Qu, J. Denitrification of groundwater using a sulfur-oxidizing autotrophic denitrifying anaerobic fluidized-bed MBR: Performance and bacterial community structure. Appl. Microbiol. Biotechnol. 2015, 99, 2815–2827. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Y.; Wan, D.; Li, B.; Zhang, P.; Wang, H. Pilot-scale application of sulfur-limestone autotrophic denitrification biofilter for municipal tailwater treatment: Performance and microbial community structure. Bioresour. Technol. 2020, 300, 122682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhangzhu, G.; Wen, S.; Lu, H.; Wang, R.; Li, W.; Ding, S.; Ghulam, A.; Zheng, P. Chemolithotrophic denitrification by nitrate-dependent anaerobic iron oxidizing (NAIO) process: Insights into the evaluation of seeding sludge. Chem. Eng. J. 2018, 345, 345–352. [Google Scholar] [CrossRef]
- Liu, H.; Yan, Q.; Shen, W. Biohydrogen facilitated denitrification at biocathode in bioelectrochemical system (BES). Bioresour. Technol. 2014, 171, 187–192. [Google Scholar] [CrossRef]
- Kondaveeti, S.; Lee, S.; Park, H.; Min, B. Bacterial communities in a bioelectrochemical denitrification system: The effects of supplemental electron acceptors. Water Res. 2014, 51, 25–36. [Google Scholar] [CrossRef]
- Mao, Y.; Xia, Y.; Zhang, T. Characterization of Thauera-dominated hydrogen-oxidizing autotrophic denitrifying microbial communities by using high-throughput sequencing. Bioresour. Technol. 2013, 128, 703–710. [Google Scholar] [CrossRef]
- Verbaendert, I.; Boon, N.; De Vos, P.; Heylen, K. Denitrification is a common feature among members of the genus Bacillus. Syst. Appl. Microbiol. 2011, 34, 385–391. [Google Scholar] [CrossRef]
- Chen, C.; Xu, X.; Xie, P.; Yuan, Y.; Zhou, X.; Wang, A.; Lee, D.; Ren, N. Pyrosequencing reveals microbial community dynamics in integrated simultaneous desulfurization and denitrification process at different influent nitrate concentrations. Chemosphere 2017, 171, 294–301. [Google Scholar] [CrossRef]
- Jena, J.; Narwade, N.; Das, T.; Dhotre, D.; Sarkar, U.; Souche, Y. Treatment of industrial effluents and assessment of their impact on the structure and function of microbial diversity in a unique Anoxic-Aerobic sequential batch reactor (AnASBR). J. Environ. Manag. 2020, 261, 110241. [Google Scholar] [CrossRef]
- Hellman, M.; Bonilla-Rosso, G.; Widerlund, A.; Juhanson, J.; Hallin, S. External carbon addition for enhancing denitrification modifies bacterial community composition and affects CH4 and N2O production in sub-arctic mining pond sediments. Water Res. 2019, 158, 22–33. [Google Scholar] [CrossRef]
- Yamada, T.; Sekiguchi, Y.; Imachi, H.; Kamagata, Y.; Ohashi, A.; Harada, H. Diversity, Localization, and Physiological Properties of Filamentous Microbes Belonging to Chloroflexi Subphylum I in Mesophilic and Thermophilic Methanogenic Sludge Granules. Appl. Environ. Microbiol. 2005, 71, 7493–7503. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Sekiguchi, Y. Cultivation of Uncultured Chloroflexi Subphyla: Significance and Ecophysiology of Formerly Uncultured Chloroflexi ‘Subphylum I’ with Natural and Biotechnological Relevance. Microbes Environ. 2009, 24, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Keller, A.H.; Kleinsteuber, S.; Vogt, C. Anaerobic Benzene Mineralization by Nitrate-Reducing and Sulfate-Reducing Microbial Consortia Enriched From the Same Site: Comparison of Community Composition and Degradation Characteristics. Microb. Ecol. 2018, 75, 941–953. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Wu, Y.; Ren, N.; Zhang, K.; Xing, D. Microbial community structure in an integrated A/O reactor treating diluted livestock wastewater during start-up period. J. Environ. Sci. 2010, 22, 656–662. [Google Scholar] [CrossRef]
- Ligi, T.; Oopkaup, K.; Truu, M.; Preem, J.; N Lvak, H.; Mitsch, W.J.; Mander, Ü.; Truu, J. Characterization of bacterial communities in soil and sediment of a created riverine wetland complex using high-throughput 16S rRNA amplicon sequencing. Ecol. Eng. 2014, 72, 56–66. [Google Scholar] [CrossRef]
- Huang, X.; Liu, L.; Wen, T.; Zhang, J.; Wang, F.; Cai, Z. Changes in the soil microbial community after reductive soil disinfestation and cucumber seedling cultivation. Appl. Microbiol. Biotechnol. 2016, 100, 5581–5593. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Peng, J.; Feng, C.; Fang, F.; Chen, S.; Xu, Y.; Wang, X. Evaluation of simultaneous autotrophic and heterotrophic denitrification processes and bacterial community structure analysis. Appl. Microbiol. Biotechnol. 2015, 99, 6527–6536. [Google Scholar] [CrossRef]
- Ding, X.; Wei, D.; Guo, W.; Wang, B.; Meng, Z.; Feng, R.; Du, B.; Wei, Q. Biological denitrification in an anoxic sequencing batch biofilm reactor: Performance evaluation, nitrous oxide emission and microbial community. Bioresour. Technol. 2019, 285, 121359. [Google Scholar] [CrossRef]
- Lang, X.; Li, Q.; Xu, Y.; Ji, M.; Yan, G.; Guo, S. Aerobic denitrifiers with petroleum metabolizing ability isolated from caprolactam sewage treatment pool. Bioresour. Technol. 2019, 290, 121719. [Google Scholar] [CrossRef]
- Rout, P.R.; Bhunia, P.; Dash, R.R. Simultaneous removal of nitrogen and phosphorous from domestic wastewater using Bacillus cereus GS-5 strain exhibiting heterotrophic nitrification, aerobic denitrification and denitrifying phosphorous removal. Bioresour. Technol. 2017, 244, 484–495. [Google Scholar] [CrossRef]
- Chen, S.; Qi, G.; Ma, G.; Zhao, X. Biochar amendment controlled bacterial wilt through changing soil chemical properties and microbial community. Microbiol. Res. 2020, 231, 126373. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gerrity, S.; Collins, G.; Chen, T.; Li, R.; Xie, S.; Zhan, X. Enrichment and characterization of autotrophic Thiobacillus denitrifiers from anaerobic sludge for nitrate removal. Process Biochem. 2018, 68, 165–170. [Google Scholar] [CrossRef]
- Luan, Y.; Yin, Y.; An, Y.; Zhang, F.; Wang, X.; Zhao, F.; Xiao, Y.; Liu, C. Investigation of an intermittently-aerated moving bed biofilm reactor in rural wastewater treatment under low dissolved oxygen and C/N condition. Bioresour. Technol. 2022, 358, 127405. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhao, J.; Zhang, H.; Chi, C.; Li, B.; Wu, X. Bacterial community shift along with the changes in operational conditions in a membrane-aerated biofilm reactor. Appl. Microbiol. Biotechnol. 2015, 99, 3279–3290. [Google Scholar] [CrossRef]
- Dong, H.; Wang, W.; Song, Z.; Dong, H.; Wang, J.; Sun, S.; Zhang, Z.; Ke, M.; Zhang, Z.; Wu, W.; et al. A high-efficiency denitrification bioreactor for the treatment of acrylonitrile wastewater using waterborne polyurethane immobilized activated sludge. Bioresour. Technol. 2017, 239, 472–481. [Google Scholar] [CrossRef]
- Munoz, R.; Rosselló-Móra, R.; Amann, R. Revised phylogeny of Bacteroidetes and proposal of sixteen new taxa and two new combinations including Rhodothermaeota phyl. nov. Syst. Appl. Microbiol. 2016, 39, 281–296. [Google Scholar] [CrossRef]
- Zuo, X.; Zhang, H.; Yu, J. Microbial diversity for the improvement of nitrogen removal in stormwater bioretention cells with three aquatic plants. Chemosphere 2020, 244, 125626. [Google Scholar] [CrossRef]
- Xu, X.; Wang, J.; Yu, T.; Nian, H.; Zhang, H.; Wang, G.; Li, F. Characterization of a novel aryloxyphenoxypropionate herbicide-hydrolyzing carboxylesterase with R-enantiomer preference from Brevundimonas sp. QPT-2. Process Biochem. 2019, 82, 102–109. [Google Scholar] [CrossRef]
- Tsubouchi, T.; Koyama, S.; Mori, K.; Shimane, Y.; Usui, K.; Tokuda, M.; Tame, A.; Uematsu, K.; Maruyama, T.; Hatada, Y. Brevundimonas denitrificans sp. nov., a denitrifying bacterium isolated from deep subseafloor sediment. Int. J. Syst. Evol. Microbiol. 2014, 64 Pt 11, 3709–3716. [Google Scholar] [CrossRef]
- Zhao, L.; Fu, G.; Tang, J.; Wu, J.; Pang, W.; Guo, Z. Efficient nitrogen removal of mangrove constructed wetlands: Enhancing heterotrophic nitrification-aerobic denitrification microflora through quorum sensing. Chem. Eng. J. 2022, 430, 133048. [Google Scholar] [CrossRef]
- Luo, X.; Su, J.; Shao, P.; Liu, H.; Luo, X. Efficient autotrophic denitrification performance through integrating the bio-oxidation of Fe(II) and Mn(II). Chem. Eng. J. 2018, 348, 669–677. [Google Scholar] [CrossRef]
- Li, Y.; Katzmann, E.; Borg, S.; Schüler, D. The Periplasmic Nitrate Reductase Nap Is Required for Anaerobic Growth and Involved in Redox Control of Magnetite Biomineralization in Magnetospirillum gryphiswaldense. J. Bacteriol. 2012, 194, 4847–4856. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florio, A.; Bréfort, C.; Gervaix, J.; Bérard, A.; Le Roux, X. The responses of NO2−- and N2O-reducing bacteria to maize inoculation by the PGPR Azospirillum lipoferum CRT1 depend on carbon availability and determine soil gross and net N2O production. Soil Biol. Biochem. 2019, 136, 107524. [Google Scholar] [CrossRef]
- Jang, J.; Sakai, Y.; Senoo, K.; Ishii, S.; Vieille, C. Potentially Mobile Denitrification Genes Identified in Azospirillum sp. Strain TSH58. Appl. Environ. Microbiol. 2019, 85, e02474-18. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.; Li, X.; Fan, W.; Wang, J. Denitrification performance and microbial community of bioreactor packed with PHBV/PLA/rice hulls composite. Sci. Total Environ. 2022, 803, 150033. [Google Scholar] [CrossRef] [PubMed]
- Sorokina, A.Y.; Chernousova, E.Y.; Dubinina, G.A. Ferrovibrio denitrificans gen. nov., sp. nov., a novel neutrophilic facultative anaerobic Fe(II)-oxidizing bacterium. FEMS Microbiol. Lett. 2012, 335, 19–25. [Google Scholar] [CrossRef]
- Huang, C.; Li, Z.; Chen, F.; Liu, Q.; Zhao, Y.; Zhou, J.; Wang, A. Microbial community structure and function in response to the shift of sulfide/nitrate loading ratio during the denitrifying sulfide removal process. Bioresour. Technol. 2015, 197, 227–234. [Google Scholar] [CrossRef]
- Cao, X.; Liu, X.; Dong, X. Alkaliphilus crotonatoxidans sp. nov., a strictly anaerobic, crotonate-dismutating bacterium isolated from a methanogenic environment. Int. J. Syst. Evol. Microbiol. 2003, 53, 971–975. [Google Scholar] [CrossRef]
- Luo, X.; Peng, C.; Shao, P.; Tang, A.; Huang, A.; Wu, Q.; Sun, L.; Yang, L.; Shi, H.; Luo, X. Enhancing nitrate removal from wastewater by integrating heterotrophic and autotrophic denitrification coupled manganese oxidation process (IHAD-MnO): Internal carbon utilization performance. Environ. Res. 2021, 194, 110744. [Google Scholar] [CrossRef]
- Robertson, E.K.; Roberts, K.L.; Burdorf, L.D.W.; Cook, P.; Thamdrup, B. Dissimilatory nitrate reduction to ammonium coupled to Fe(II) oxidation in sediments of a periodically hypoxic estuary. Limnol. Oceanogr. 2016, 61, 365–381. [Google Scholar] [CrossRef]
- Sievert, S.M.; Scott, K.M.; Klotz, M.G.; Chain, P.G.; Hauser, L.J.; Hemp, J.; Hügler, M.; Land, M.; Lapidus, A.; Larimer, F.; et al. Genome of the epsilonproteobacterial chemolithoautotroph Sulfurimonas denitrificans. Appl. Environ. Microbiol. 2008, 74, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Dong, G.; Gong, L.; Li, Q.; Wang, Y. The Role of Low-Molecular-Weight Organic Carbons in Facilitating the Mobilization and Biotransformation of As(V)/Fe(III) from a Realgar Tailing Mine Soil. Geomicrobiol. J. 2018, 35, 555–563. [Google Scholar] [CrossRef]
- Corsini, A.; Cavalca, L.; Crippa, L.; Zaccheo, P.; Andreoni, V. Impact of glucose on microbial community of a soil containing pyrite cinders: Role of bacteria in arsenic mobilization under submerged condition. Soil Biol. Biochem. 2010, 42, 699–707. [Google Scholar] [CrossRef]
- Luo, S.; Guo, W.; Nealson, K.H.; Feng, X.; He, Z. 13C Pathway Analysis for the Role of Formate in Electricity Generation by Shewanella Oneidensis MR-1 Using Lactate in Microbial Fuel Cells. Sci. Rep. 2016, 6, 20941. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Soil | Nitrate-N (mg L−1) | Acetate-C (mg L−1) | NaHCO3 (mg L−1) | Pyrite (g) |
|---|---|---|---|---|---|
| Control | 0 | 140 | 0 | 840 | 30 |
| A0 | 0.1% | 140 | 0 | 840 | 30 |
| A24 | 0.1% | 140 | 24 | 840 | 30 |
| A48 | 0.1% | 140 | 48 | 840 | 30 |
| A120 | 0.1% | 140 | 120 | 840 | 30 |
| A240 | 0.1% | 140 | 240 | 840 | 30 |
| A480 | 0.1% | 140 | 480 | 840 | 30 |
| Treatment | Microbial Source | Nitrate-N (mg L−1) | NaHCO3 (mg L−1) | Pyrite (g) |
|---|---|---|---|---|
| Control | Physiological saline | 50 | 400 | 30 |
| B0 | A0 | 50 | 400 | 30 |
| B24 | A24 | 50 | 400 | 30 |
| B48 | A48 | 50 | 400 | 30 |
| B120 | A120 | 50 | 400 | 30 |
| B240 | A240 | 50 | 400 | 30 |
| B480 | A480 | 50 | 400 | 30 |
| Sample ID | Reads | OTUs 2 | Ace | Chao1 | Coverage | Shannon | Simpson |
|---|---|---|---|---|---|---|---|
| Soil 1 | 55,928 | 1180 | 1189.804 | 1185.194 | 0.999392 | 5.025932 | 0.028366 |
| A0 | 52,525 | 378 | 386.8607 | 387.7308 | 0.999562 | 4.330376 | 0.024486 |
| A24 | 64,104 | 133 | 188.8802 | 166 | 0.999485 | 1.714763 | 0.38743 |
| A48 | 62,599 | 169 | 216.1763 | 238.4615 | 0.999313 | 1.886757 | 0.388363 |
| A120 | 64,316 | 107 | 135.5286 | 141.3636 | 0.999565 | 1.310092 | 0.509113 |
| A240 | 61,244 | 81 | 97.28298 | 96.3 | 0.999706 | 1.026962 | 0.564491 |
| A480 | 54,153 | 82 | 104.8487 | 105.0769 | 0.999538 | 1.601675 | 0.250963 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, B.; Yang, X.; Li, Y.; Yang, K.; Xiong, Y.; Yuan, N. Pyrite-Based Autotrophic Denitrifying Microorganisms Derived from Paddy Soils: Effects of Organic Co-Substrate Addition. Int. J. Environ. Res. Public Health 2022, 19, 11763. https://doi.org/10.3390/ijerph191811763
Xu B, Yang X, Li Y, Yang K, Xiong Y, Yuan N. Pyrite-Based Autotrophic Denitrifying Microorganisms Derived from Paddy Soils: Effects of Organic Co-Substrate Addition. International Journal of Environmental Research and Public Health. 2022; 19(18):11763. https://doi.org/10.3390/ijerph191811763
Chicago/Turabian StyleXu, Baokun, Xiaoxia Yang, Yalong Li, Kejun Yang, Yujiang Xiong, and Niannian Yuan. 2022. "Pyrite-Based Autotrophic Denitrifying Microorganisms Derived from Paddy Soils: Effects of Organic Co-Substrate Addition" International Journal of Environmental Research and Public Health 19, no. 18: 11763. https://doi.org/10.3390/ijerph191811763