
N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Experimental Animals
- The first group (control) received a basal diet without any treatments;
- The second group (NAC) fed on a diet supplemented with NAC, 5.2 g/kg diet [21];
- The third group (DLM) fed on a diet contaminated with DLM 300 mg/kg diet [22];
- The fourth group (DLM+NAC) fed on a diet mixed with DLM and NAC, at the same rates of the above-mentioned doses.
2.3. Growth Parameters
2.4. Blood Samples
2.5. Hepato-Renal Function Tests
2.6. Determination of Oxidative Stress Markers
2.7. Real-Time PCR
2.8. Examination of Histopathology
2.9. Statistical Analysis
3. Results
3.1. Effect of DLM and/or NAC on Growth Performance
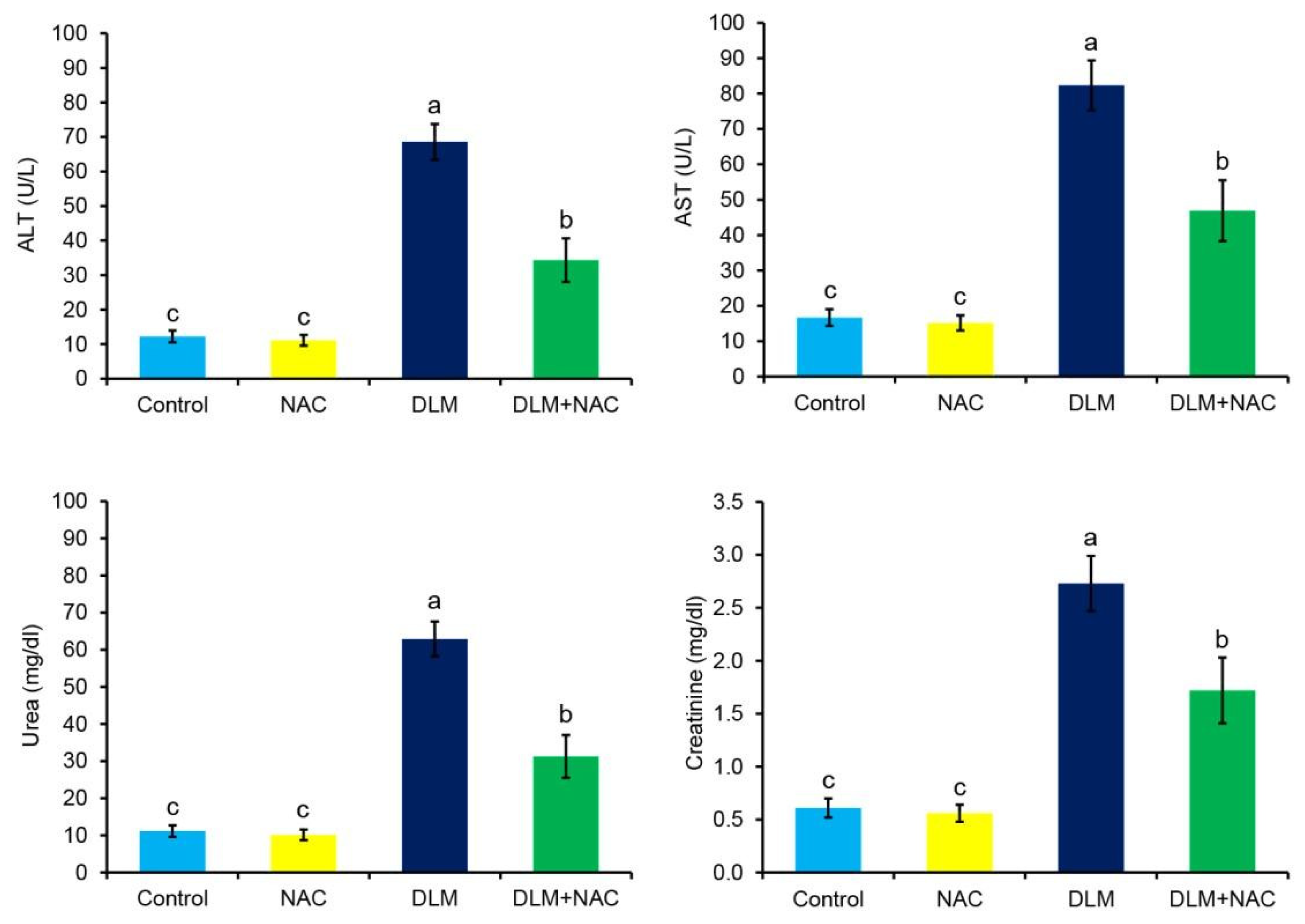
3.2. The Effect of DLM and/or NAC on Biochemical Markers of the Liver and Kidney
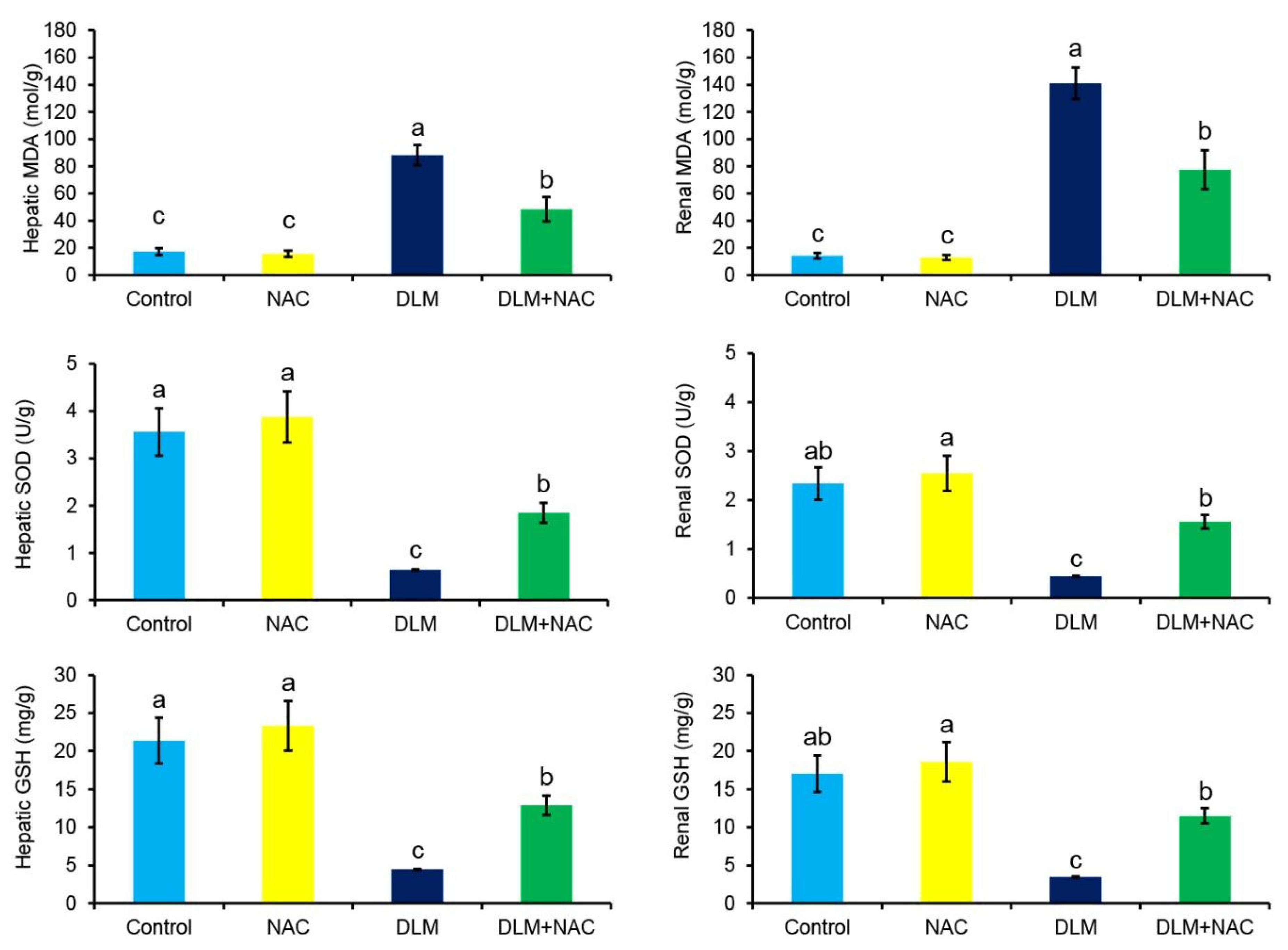
3.3. The Effect of DLM and/or NAC on Oxidative Cascade in Liver and Kidney Tissues
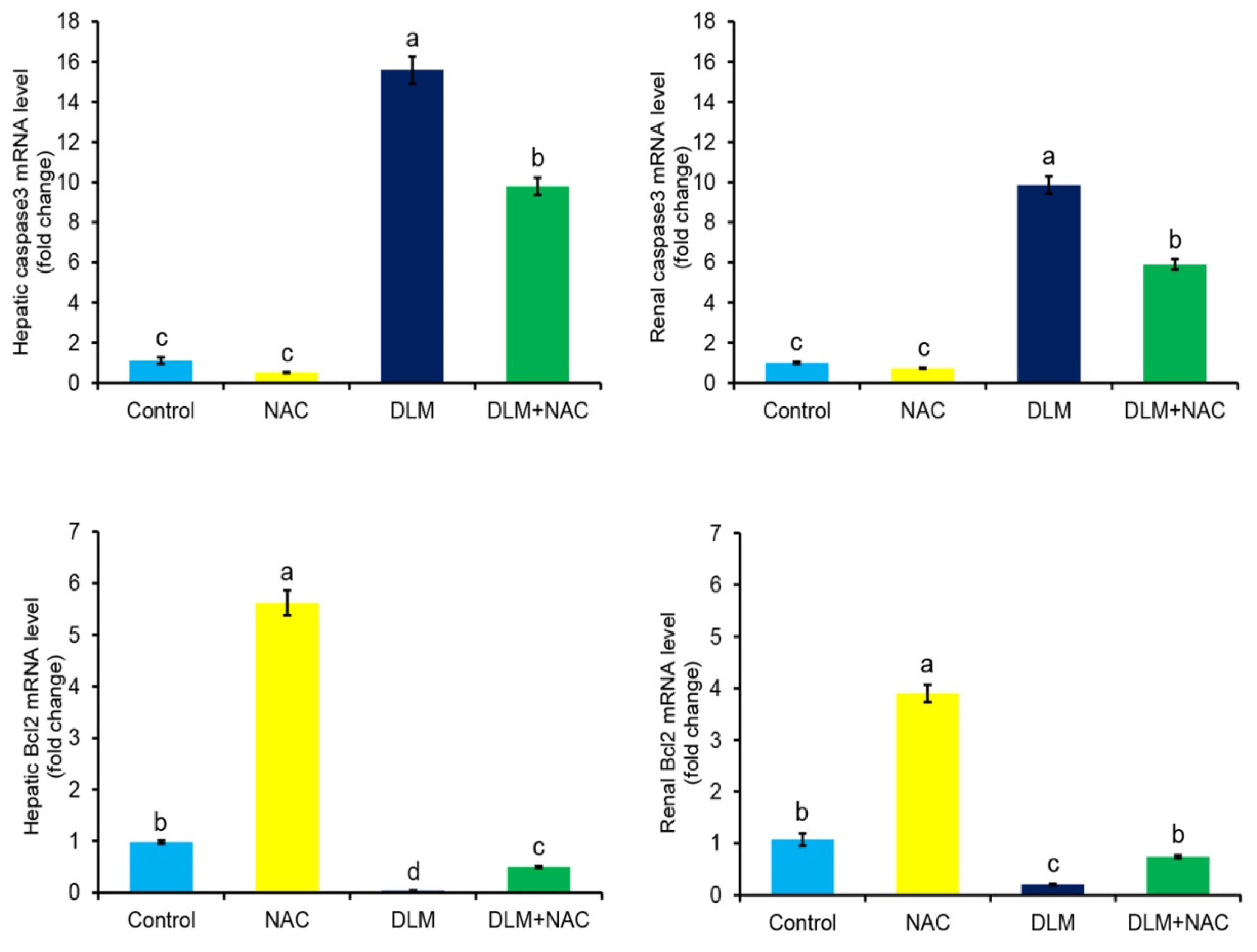
3.4. The Effect of DLM and/or NAC on Caspase-3 and Bcl2 mRNA Expression in Liver and Kidney Tissues
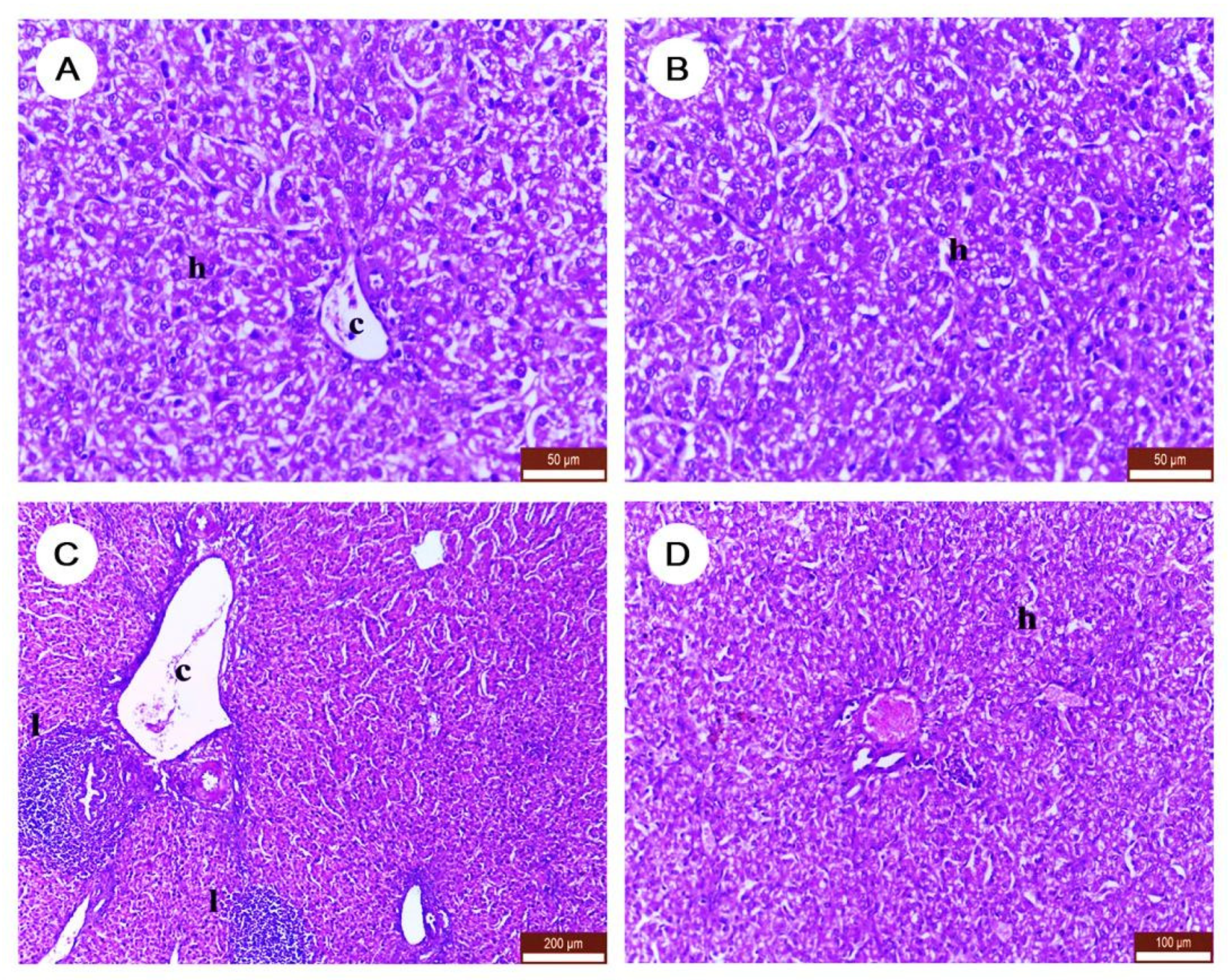
3.5. The Effect of DLM and/or NAC on Histopathological Character of Liver Tissues
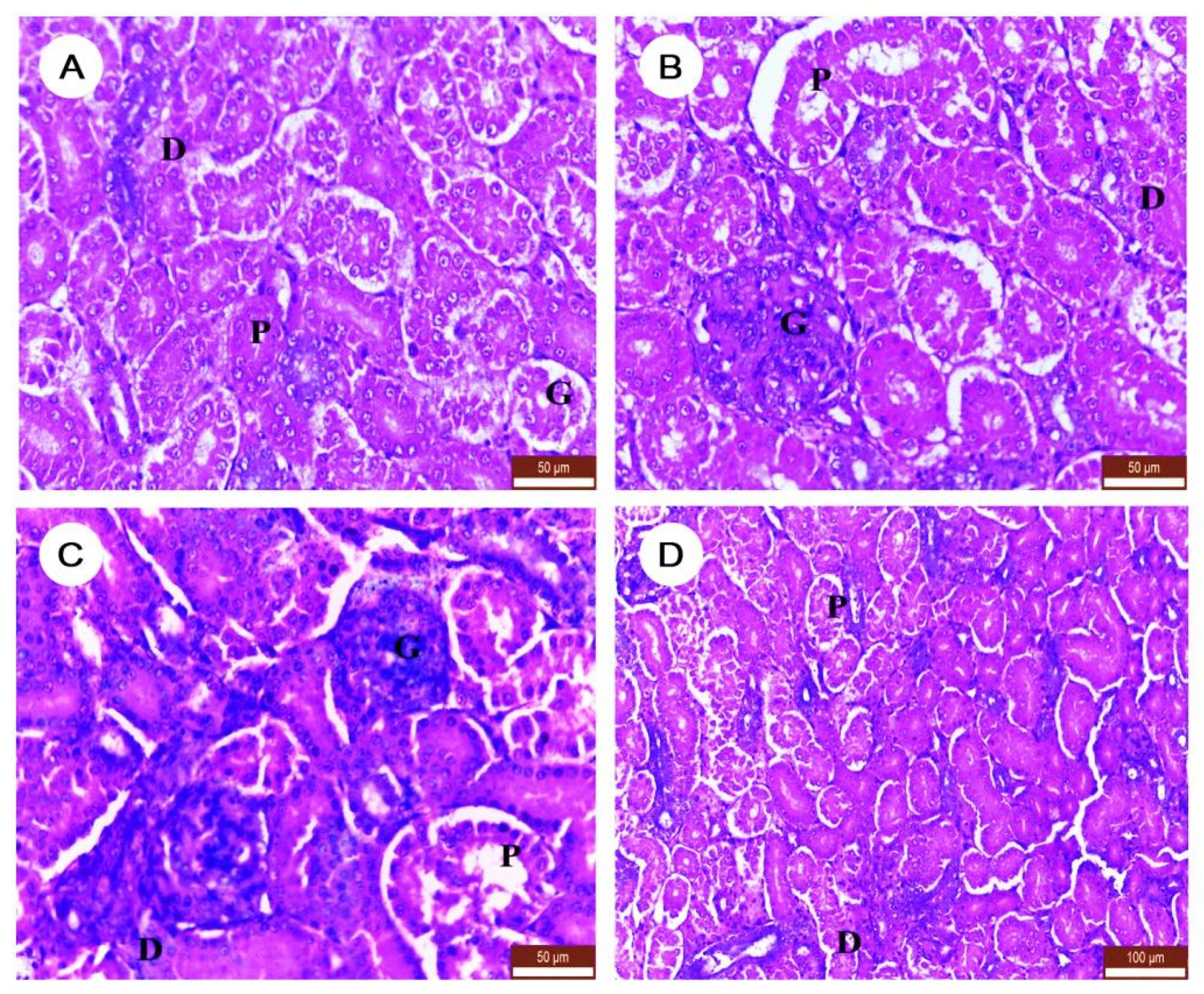
3.6. The Effect of DM and/or NAC on Histopathological Character of Kidney Tissues
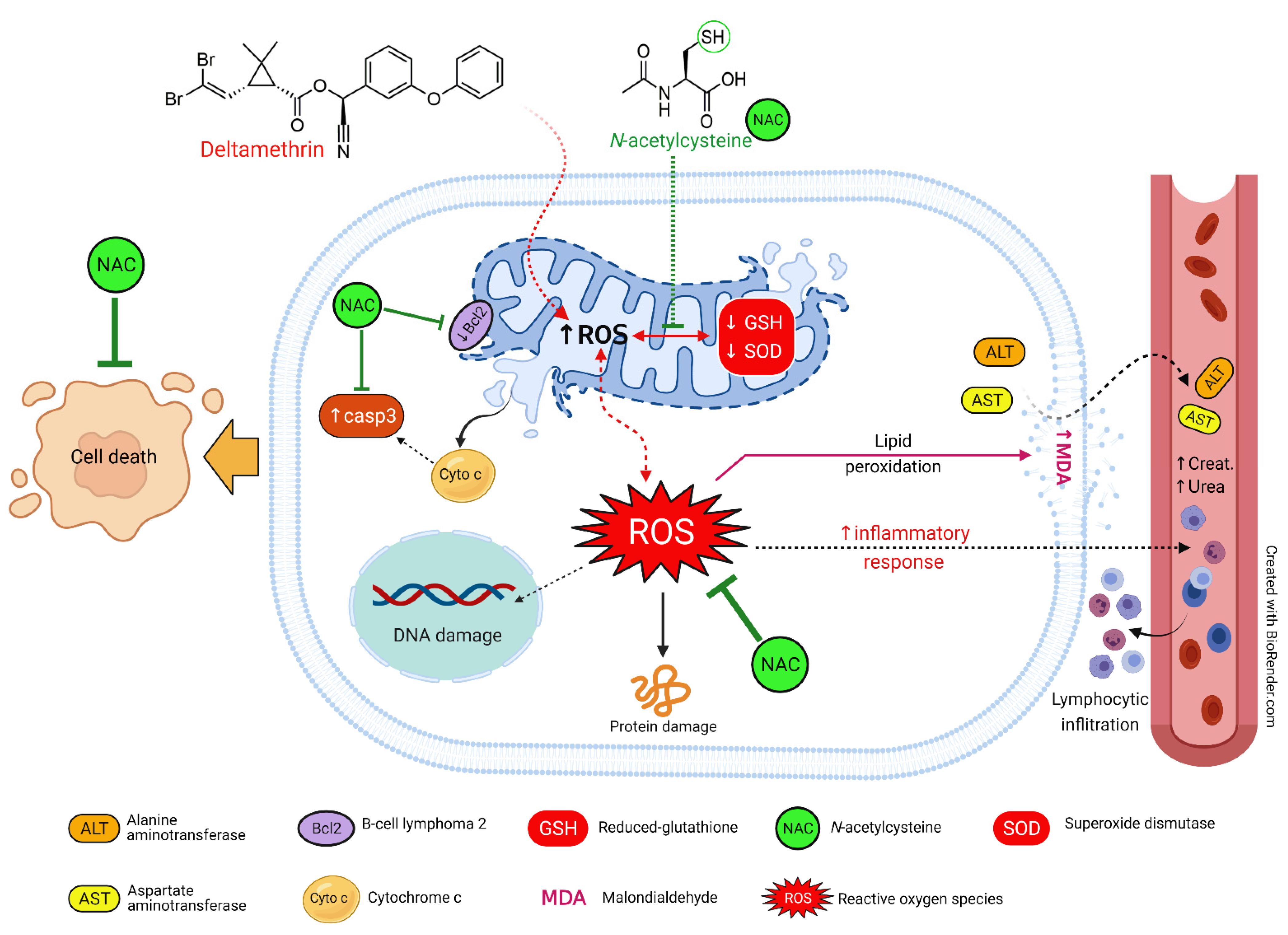
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kurek, M.; Barchańska, H.; Turek, M. Degradation processes of pesticides used in potato cultivations. In Reviews of Environmental Contamination and Toxicology; Springer: Cham, Switzerland, 2016; Volume 242, pp. 105–151. [Google Scholar]
- Lv, Y.; Bing, Q.; Lv, Z.; Xue, J.; Li, S.; Han, B.; Yang, Q.; Wang, X.; Zhang, Z. Imidacloprid-induced liver fibrosis in quails via activation of the TGF-β1/Smad pathway. Sci. Total Environ. 2020, 705, 135915. [Google Scholar] [CrossRef]
- Saoudi, M.; Badraoui, R.; Bouhajja, H.; Ncir, M.; Rahmouni, F.; Grati, M.; Jamoussi, K.; El Feki, A. Deltamethrin induced oxidative stress in kidney and brain of rats: Protective effect of Artemisia campestris essential oil. Biomed. Pharmacother. 2017, 94, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Abdou, R.H.; Abdel-Daim, M.M. Alpha-lipoic acid improves acute deltamethrin-induced toxicity in rats. Can. J. Physiol. Pharmacol. 2014, 92, 773–779. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdelkhalek, N.K.M.; Hassan, A.M. Antagonistic activity of dietary allicin against deltamethrin-induced oxidative damage in freshwater Nile tilapia; Oreochromis niloticus. Ecotoxicol. Environ. Saf. 2015, 111, 146–152. [Google Scholar] [CrossRef]
- Barlow, S.M.; Sullivan, F.M.; Lines, J. Risk assessment of the use of deltamethrin on bednets for the prevention of malaria. Food Chem. Toxicol. 2001, 39, 407–422. [Google Scholar] [CrossRef]
- Salim, G.; Rachid, R.; Mohamed, K. Deltamethrin induced neurodegeneration and behavioral effect by dysfunction cytosolic antioxidant system in rats brain. Algerian J. Nat. Sci. 2016, 1, 14–22. [Google Scholar]
- Chargui, I.; Grissa, I.; Bensassi, F.; Hrira, M.Y.; Haouem, S.; Haouas, Z.; Bencheikh, H. Oxidative stress, biochemical and histopathological alterations in the liver and kidney of female rats exposed to low doses of deltamethrin (DM): A molecular assessment. Biomed. Environ. Sci. 2012, 25, 672–683. [Google Scholar] [PubMed]
- Abdollahi, M.; Ranjbar, A.; Shadnia, S.; Nikfar, S.; Rezaie, A. Pesticides and oxidative stress: A review. Med. Sci. Monit. 2004, 10, 141–148. [Google Scholar]
- Vanova, N.; Pejchal, J.; Herman, D.; Dlabkova, A.; Jun, D. Oxidative stress in organophosphate poisoning: Role of standard antidotal therapy. J. Appl. Toxicol. 2018, 38, 1058–1070. [Google Scholar] [CrossRef]
- Anand, S.S.; Bruckner, J.V.; Haines, W.T.; Muralidhara, S.; Fisher, J.W.; Padilla, S. Characterization of deltamethrin metabolism by rat plasma and liver microsomes. Toxicol. Appl. Pharmacol. 2006, 212, 156–166. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abuzead, S.M.M.; Halawa, S.M. Protective role of Spirulina platensis against acute deltamethrin-induced toxicity in rats. PLoS ONE 2013, 8, e72991. [Google Scholar]
- Cankayali, L.; Demirag, K.; Eris, O.; Ersoz, B.; Moral, A.R. The effects of N-acetylcysteine on oxidative stress in organophosphate poisoning model. Adv. Ther. 2005, 22, 107–116. [Google Scholar] [CrossRef]
- El-Ebiary, A.A.; Elsharkawy, R.E.; Soliman, N.A.; Soliman, M.A.; Hashem, A.A. N-acetylcysteine in acute organophosphorus pesticide poisoning: A randomized, clinical trial. Basic Clin. Pharmacol. Toxicol. 2016, 119, 222–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Sasmal, D.; Bhaskar, A.; Mukhopadhyay, K.; Thakur, A.; Sharma, N. First record of the ground beetle Trechoblemus postilenatus (Coleoptera, Carabidae) in Primorskii krai. Environ. Toxicol. 2016, 31, 808–819. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Håkansson, N.; Wolk, A. Dietary cysteine and other amino acids and stroke incidence in women. Stroke 2015, 46, 922–926. [Google Scholar] [CrossRef] [Green Version]
- Mokhtari, V.; Afsharian, P.; Shahhoseini, M.; Kalantar, S.M.; Moini, A. A review on various uses of N-acetyl cysteine. Cell J. 2017, 19, 11. [Google Scholar]
- Millea, P.J. N-acetylcysteine: Multiple clinical applications. Am. Fam. Physician 2009, 80, 265–269. [Google Scholar]
- Valdivia, A.G.; Martinez, A.; Damian, F.J.; Quezada, T.; Ortiz, R.; Martinez, C.; Llamas, J.; Rodriguez, M.L.; Yamamoto, L.; Jaramillo, F.; et al. Efficacy of N-acetylcysteine to reduce the effects of aflatoxin B1 intoxication in broiler chickens. Poult. Sci. 2001, 80, 727–734. [Google Scholar] [CrossRef]
- Yi, D.; Hou, Y.; Tan, L.; Liao, M.; Xie, J.; Wang, L.; Ding, B.; Yang, Y.; Gong, J. N-acetylcysteine improves the growth performance and intestinal function in the heat-stressed broilers. Anim. Feed Sci. Technol. 2016, 220, 83–92. [Google Scholar] [CrossRef]
- Elmasry, D.; Hegazi, S.M.; El-Sabagh, M.; Nasef, S.A. Impact of N-acetyl cysteine (NAC) chit Nanocomposite as antioxidant on broiler chicks. Afr. J. Biol. Sci. 2020, 16, 33–42. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, S.S.; Elsabagh, R.; Allam, A.; Youssef, G.; Fadl, S.E.; Abdelhiee, E.Y.; Alkafafy, M.; Soliman, A.; Aboubakr, M. Bioremediation role of Spirulina platensis against deltamethrin-mediated toxicity and its chemical residues in chicken meat. Environ. Sci. Pollut. Res. 2021, 28, 56188–56198. [Google Scholar] [CrossRef]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Cui, H.; Peng, X.; Fang, J.; Zuo, Z.; Deng, J.; Wu, B. The association between splenocyte apoptosis and alterations of Bax, Bcl-2 and caspase-3 mRNA expression, and oxidative stress induced by dietary nickel chloride in broilers. Int. J. Environ. Res. Public Health 2013, 10, 7310–7326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Zhao, H.; Wang, Y.; Shao, Y.; Li, J.; Xing, M. Alterations of antioxidant indexes and inflammatory cytokine expression aggravated hepatocellular apoptosis through mitochondrial and death receptor-dependent pathways in Gallus gallus exposed to arsenic and copper. Environ. Sci. Pollut. Res. 2018, 25, 15462–15473. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.M.; Guo, Y.M.; Yang, Y.; Wang, Z.H. Characterization of fatty acid digestion of Beijing Fatty and Arbor Acres Chickens. Asian-Australas. J. Anim. Sci. 2007, 20, 1222–1228. [Google Scholar] [CrossRef]
- Wang, X.H.; Souders II, C.L.; Xavier, P.; Li, X.Y.; Yan, B.; Martyniuk, C.J. The pyrethroid esfenvalerate induces hypoactivity and decreases dopamine transporter expression in embryonic/larval zebrafish (Danio rerio). Chemosphere 2020, 243, 125416. [Google Scholar] [CrossRef]
- Liu, G.; Yan, X.; Sedykh, A.; Pan, X.; Zhao, X.; Yan, B.; Zhu, H. Analysis of model PM2. 5-induced inflammation and cytotoxicity by the combination of a virtual carbon nanoparticle library and computational modeling. Ecotoxicol. Environ. Saf. 2020, 191, 110216. [Google Scholar] [CrossRef]
- Sibiya, I.; Poma, G.; Cuykx, M.; Covaci, A.; Adegbenro, P.D.; Okonkwo, J. Targeted and non-target screening of persistent organic pollutants and organophosphorus flame retardants in leachate and sediment from landfill sites in Gauteng Province, South Africa. Sci. Total Environ. 2019, 653, 1231–1239. [Google Scholar] [CrossRef]
- Prohaszka, L.; Hajdu, E.; Dworschak, E.; Rozsnyai, T. Growth depression in broiler chicks caused by incompatibility of feed ingredients. Acta Vet. Hung. 1987, 35, 349–358. [Google Scholar]
- Abdel-Daim, M.; El-Bialy, B.E.; Rahman, H.G.A.; Radi, A.M.; Hefny, H.A.; Hassan, A.M. Antagonistic effects of Spirulina platensis against sub-acute deltamethrin toxicity in mice: Biochemical and histopathological studies. Biomed. Pharmacother. 2016, 77, 79–85. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abd Eldaim, M.A.; Mahmoud, M.M. Trigonella foenum-graecum protection against deltamethrin-induced toxic effects on haematological, biochemical, and oxidative stress parameters in rats. Can. J. Physiol. Pharmacol. 2014, 92, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, N.K.M.; Ghazy, E.W.; Abdel-Daim, M.M. Pharmacodynamic interaction of Spirulina platensis and deltamethrin in freshwater fish Nile tilapia, Oreochromis niloticus: Impact on lipid peroxidation and oxidative stress. Environ. Sci. Pollut. Res. 2015, 22, 3023–3031. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abdeen, A. Protective effects of rosuvastatin and vitamin E against fipronil-mediated oxidative damage and apoptosis in rat liver and kidney. Food Chem. Toxicol. 2018, 114, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Mossa, A.-T.H.; Refaie, A.A.; Ramadan, A.; Bouajila, J. Amelioration of prallethrin-induced oxidative stress and hepatotoxicity in rat by the administration of Origanum majorana essential oil. BioMed Res. Int. 2013, 2013, 859085. [Google Scholar] [CrossRef] [Green Version]
- Yang, D.; Tan, X.; Lv, Z.; Liu, B.; Baiyun, R.; Lu, J.; Zhang, Z. Regulation of Sirt1/Nrf2/TNF-α signaling pathway by luteolin is critical to attenuate acute mercuric chloride exposure induced hepatotoxicity. Sci. Rep. 2016, 6, 37157. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, S.; Jiang, H.; Liu, B.; Lv, Z.; Guo, C.; Zhang, H. Effects of selenium on apoptosis and abnormal amino acid metabolism induced by excess fatty acid in isolated rat hepatocytes. Mol. Nutr. food Res. 2017, 61, 1700016. [Google Scholar] [CrossRef]
- Abdel-Daim, M.; Dawood, M.A.O.; AlKahtane, A.A.; Abdeen, A.; Abdel-Latif, H.M.R.; Senousy, H.H.; Aleya, L.; Alkahtani, S. Spirulina platensis mediated the biochemical indices and antioxidative function of Nile tilapia (Oreochromis niloticus) intoxicated with aflatoxin B1. Toxicon 2020, 184, 152–157. [Google Scholar] [CrossRef]
- Abdeen, A.; Abdelkader, A.; Elgazzar, D.; Aboubakr, M.; Abdulah, O.A.; Shoghy, K.; Abdel-Daim, M.; El-Serehy, H.A.; Najda, A.; El-Mleeh, A. Coenzyme Q10 supplementation mitigates piroxicam-induced oxidative injury and apoptotic pathways in the stomach, liver, and kidney. Biomed. Pharmacother. 2020, 130, 110627. [Google Scholar] [CrossRef]
- Aboubakr, M.; Elshafae, S.M.; Abdelhiee, E.Y.; Fadl, S.E.; Soliman, A. Antioxidant and Anti-Inflammatory Potential of Thymoquinone and Lycopene Mitigate the Chlorpyrifos-Induced Toxic Neuropathy. Pharmaceuticals 2021, 14, 940. [Google Scholar] [CrossRef]
- Abdeen, A.; Aboubakr, M.; Elgazzar, D.; Abdo, M.; Abdelkader, A.; Ibrahim, S.; Elkomy, A. Rosuvastatin attenuates piroxicam-mediated gastric ulceration and hepato-renal toxicity in rats. Biomed. Pharmacother. 2019, 110, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Abdeen, A.; Abou-Zaid, O.A.; Abdel-Maksoud, H.A.; Aboubakr, M.; Abdelkader, A.; Abdelnaby, A.; Abo-Ahmed, A.I.; El-Mleeh, A.; Mostafa, O.; Abdel-Daim, M.; et al. Cadmium overload modulates piroxicam-regulated oxidative damage and apoptotic pathways. Environ. Sci. Pollut. Res. 2019, 26, 25167–25177. [Google Scholar] [CrossRef]
- Abdeen, A.; Samir, A.; Elkomy, A.; Aboubaker, M.; Habotta, O.A.; Gaber, A.; Alsanie, W.F.; Abdullah, O.; Elnoury, H.A.; Baioumy, B.; et al. The potential antioxidant bioactivity of date palm fruit against gentamicin-mediated hepato-renal injury in male albino rats. Biomed. Pharmacother. 2021, 143, 112154. [Google Scholar] [CrossRef]
- Abdeen, A.; Abdelkader, A.; Abdo, M.; Wareth, G.; Aboubakr, M.; Aleya, L.; Abdel-Daim, M. Protective effect of cinnamon against acetaminophen-mediated cellular damage and apoptosis in renal tissue. Environ. Sci. Pollut. Res. 2019, 26, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Sayed, A.A.; Abdeen, A.; Aleya, L.; Ali, D.; Alkahtane, A.A.; Alarifi, S.; Alkahtani, S. Piperine enhances the antioxidant and anti-inflammatory activities of thymoquinone against microcystin-LR-induced hepatotoxicity and neurotoxicity in mice. Oxid. Med. Cell. Longev. 2019, 2019, 1309175. [Google Scholar] [CrossRef]
- Wong, C.K.; Ooi, V.E.C.; Wong, C.K. Protective effects of N-acetylcysteine against carbon tetrachloride-and trichloroethylene-induced poisoning in rats. Environ. Toxicol. Pharmacol. 2003, 14, 109–116. [Google Scholar] [CrossRef]
- Iyaswamy, A.; Wankhar, D.; Loganathan, S.; Shanmugam, S.; Rajan, R.; Rathinasamy, S. Disruption of redox homeostasis in liver function and activation of apoptosis on consumption of aspartame in folate deficient rat model. J. Nutr. Intermed. Metab. 2017, 8, 41–50. [Google Scholar] [CrossRef]
- Simon, H.-U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef]
- Pelicano, H.; Carney, D.; Huang, P. ROS stress in cancer cells and therapeutic implications. Drug Resist. Updat. 2004, 7, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Khalaf, A.A.; Galal, M.K.; Ibrahim, M.A.; Abd Allah, A.A.; Afify, M.M.; Refaat, R. The Terminalia laxiflora modulates the neurotoxicity induced by fipronil in male albino rats. Biosci. Rep. 2019, 39, BSR20181363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aminzadeh, A.; Dehpour, A.R.; Safa, M.; Mirzamohammadi, S.; Sharifi, A.M. Investigating the protective effect of lithium against high glucose-induced neurotoxicity in PC12 cells: Involvements of ROS, JNK and P38 MAPKs, and apoptotic mitochondria pathway. Cell. Mol. Neurobiol. 2014, 34, 1143–1150. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (kg/100 kg) | Starter Diet (0 to 21 Days) | Finisher Diet (22–35 Days) |
|---|---|---|
| Yellow corn | 58.57 | 64 |
| Soyabean meal (48%) | 32 | 25 |
| Corn gluten (60%) | 5 | 5 |
| Soybean oil | 0.7 | 2.5 |
| Di-calcium phosphate (22% Ca and 19% Ph) | 1.5 | 0.95 |
| Limestone (35% Ca) | 1.7 | 1.74 |
| Common salts | 0.2 | 0.15 |
| Methionine (95%) | 0.13 | 0.36 |
| Lysine (98%) | 0 | 0.1 |
| Vitamins and mineral premix * | 0.2 | 0.2 |
| Calculated composition | ||
| Protein (%) | 23 | 20 |
| k. calory ME/kg | 2950 | 3120 |
| Calcium (%) | 1 | 0.9 |
| Phosphorus (%) | 0.48 | 0.35 |
| Gene Name | Primer Sequence (5′-3′) | Reference |
|---|---|---|
| Caspase3 | F: TGGCCCTCTTGAACTGAAAG R: TCCACTGTCTGCTTCAATACC | [24] |
| Bcl2 | F: ATCGTCGCCTTCTTCGAGTT R: ATCCCATCCTCCGTTGTTCT | [25] |
| ß-actin | F: CCACCGCAAATGCTTCTAAAC R: AAGACTGCTGCTGACACCTTC | [26] |
| Parameters | Control | NAC | DLM | DLM+NAC |
|---|---|---|---|---|
| B. wt (7 days; g) | 143.80 ± 0.50 a | 144.13 ± 0.97 a | 139.27 ± 0.41 b | 143.27 ± 1.01 a |
| B. wt (35 days; g) | 1938.6 ± 2.11 b | 2119.6 ± 1.73 a | 1526.7 ± 8.70 d | 1761.9 ± 8.92 c |
| Weight gain (g) | 1794.8 ± 2.28 b | 1975.5 ± 1.52 a | 1387.5 ± 8.76 d | 1618.6 ± 9.08 c |
| Feed intake (g) | 3254.3 ± 2.48 c | 3383.3 ± 7.17 a | 3931.7 ± 2.63 d | 3286.3 ± 13.10 b |
| FCR (%) | 1.81 ± 0.002 c | 1.71 ± 0.004 d | 2.11 ± 0.013 a | 2.03 ± 0.016 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allam, A.; Abdeen, A.; Devkota, H.P.; Ibrahim, S.S.; Youssef, G.; Soliman, A.; Abdel-Daim, M.M.; Alzahrani, K.J.; Shoghy, K.; Ibrahim, S.F.; et al. N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues. Int. J. Environ. Res. Public Health 2022, 19, 638. https://doi.org/10.3390/ijerph19020638
Allam A, Abdeen A, Devkota HP, Ibrahim SS, Youssef G, Soliman A, Abdel-Daim MM, Alzahrani KJ, Shoghy K, Ibrahim SF, et al. N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues. International Journal of Environmental Research and Public Health. 2022; 19(2):638. https://doi.org/10.3390/ijerph19020638
Chicago/Turabian StyleAllam, Ali, Ahmed Abdeen, Hari Prasad Devkota, Samar S. Ibrahim, Gehan Youssef, Ahmed Soliman, Mohamed M. Abdel-Daim, Khalid J. Alzahrani, Khaled Shoghy, Samah F. Ibrahim, and et al. 2022. "N-Acetylcysteine Alleviated the Deltamethrin-Induced Oxidative Cascade and Apoptosis in Liver and Kidney Tissues" International Journal of Environmental Research and Public Health 19, no. 2: 638. https://doi.org/10.3390/ijerph19020638